Introduction
As a type of primary liver cancer, hepatocellular
carcinoma (HCC) has a high mortality rate with 782,000 deaths
annually worldwide (1-3). Although significant progress has
been achieved in HCC treatment, the overall prognosis is still not
optimal due to the high metastasis rate and high recurrence rate
(4,5). For patients with HCC, surgical
resection is the most effective treatment method (6). It has been reported that anesthetics
can affect the treatment of multiple human cancer types, such as
osteosarcoma (7), ovarian cancer
(8) and breast cancer (9). Therefore, it is important to
investigate the underlying mechanism of action of anesthetics
affecting the development of cancer.
Propofol is a short-acting intravenous anesthetic
that is widely used to relieve the pain of patients during surgery
(10). Previous studies have
revealed that propofol is associated with the progression of
numerous human tumors. For instance, Zhang et al (11) revealed that propofol induced cell
proliferation and invasion, but restrained cell apoptosis in
gallbladder cancer. Moreover, Wang et al (12) showed that propofol suppressed cell
proliferation and metastasis in glioma, while Liu et al
(13) reported that propofol
served a tumor suppression role in pancreatic cancer. In addition,
Ou et al (14)
demonstrated that propofol repressed HCC cell proliferation and
metastasis, as well as induced apoptosis. These findings suggest
that propofol serves different roles in human cancer types.
However, the exact function and mechanism of propofol in HCC
requires further investigation.
As a family of non-coding transcripts that are
>200 nucleotides in length, long non-coding RNAs (lncRNAs)
participate in various biological processes, such as
differentiation, cell development, survival and apoptosis (15,16). Previous studies have reported that
lncRNAs, such as antisense noncoding RNA in the INK4 locus
(17), taurine upregulated 1
(18) and DiGeorge syndrome
critical region gene 5 (19),
could be dysregulated by propofol treatment in human cancer types.
Moreover, multiple lncRNAs have been demonstrated to serve vital
roles in HCC. For example, MYD88 innate immune signal transduction
adaptor can promote HCC cell proliferation and metastasis (20). Furthermore, E74-like ETS
transcription factor 209 could suppress tumor progression via
inhibiting cell metastasis in HCC (21). HOMEOBOX A11 (HOXA11) antisense RNA
(HOXA11-AS) has also been identified to be associated with HCC
(22). However, the regulatory
mechanism of HOXA11-AS in HCC is not fully characterized, and
whether there is an association between propofol and HOXA11-AS is
yet to be elucidated.
MicroRNAs (miRNAs/miRs), a family of endogenous RNAs
with 19-22 nucleotides, have crucial roles in human cancer,
including HCC (23). In recent
decades, numerous miRNAs have been identified to be involved in the
promotion of HCC. For example, Wang et al (24) identified that miR-194-5p repressed
HCC cell proliferation and induced cell apoptosis. Moreover, Kabir
et al (25) reported that
miR-7 affected cell viability and metastasis in HCC. miR-4458 has
also been shown to exert an anti-tumor effect in HCC (26). Thus, as lncRNAs can regulate miRNA
expression levels and activities by sponging to miRNAs (27), whether HOXA11-AS can target
miR-4458 in HCC requires further investigation.
The present study aimed to evaluate the functions of
propofol in tumor progression in HCC. In addition, the influences
of propofol on HOXA11-AS and miR-4458 were investigated, as well as
the roles of HOXA11-AS and miR-4458 in HCC cell proliferation,
apoptosis and metastasis.
Materials and methods
Cell culture
HCC cell lines Hep3B (cat. no. SCSP-5045) and Huh-7
(cat. no. SCSP-526) were purchased from the Type Culture Collection
of the Chinese Academy of Sciences. HCC cells were cultured in DMEM
(cat. no. 10099-141; Gibco; Thermo Fisher Scientific, Inc.)
supplemented with 10% FBS (cat. no. 12483-012; Gibco; Thermo Fisher
Scientific, Inc.) and 1% penicillin/streptomycin (cat. no.
15140-122; Gibco; Thermo Fisher Scientific, Inc.) in an incubator
at 37°C with 5% CO2.
Propofol treatment
Propofol (cat. no. BP1031 MSDS; Sigma-Aldrich; Merck
KGaA) was dissolved in DMSO (40 mg/ml; cat. no. D8371; Beijing
Solarbio Science & Technology Co., Ltd.) and diluted in the
culture medium at 37°C for 15 min to achieve final concentrations
of 2.5, 5 and 10 µg/ml for the experiments. Then, HCC cells
were exposed to different concentrations of propofol (2.5, 5 or 10
µg/ml) at 37°C for 24 h according to previous studies
(28,29). The control groups were treated
with equal volume of 0.2% DMSO (500 µl; cat. no. D8371;
Beijing Solarbio Science & Technology Co., Ltd.) at 37°C for 24
h.
Cell transfection
The overexpression plasmid of HOXA11-AS and
corresponding empty plasmid of pcDNA 3.1 (Vector); small
interfering RNA targeting HOXA11-AS (si-HOXA11-AS#1, 5′-CTA CCA TCC
CTG AGC CTT A-3′; si-HOXA11-AS#2, 5′- CAG AAG AAT GGT ACA ATC CAA
G-3′; si-HOXA11-AS#3, 5′-AGG ATG AGA TTC AGA ATA TGA AG-3′) and its
matched negative control (si-NC; 5′-CCT ATC TGG TCA ACA CGT
ATT-3′); mimics of miR-4458 (miR-4458; 5′-AGA GGU AGG UGU GGA AGA
A-3′) and its corresponding NC (miR-NC; 5′-UUC UCC GAA CGU GUC ACG
UUU -3′); inhibitors of miR-4458 (anti-miR-4458; 5′-UUC UUC CAC ACC
UAC CUC U-3′) and its NC (anti-miR-NC; 5′-CAG UAC UUU UGU GUA GUA
CAA -3′) were all purchased from Sangon Biotech Co., Ltd.
Hep3B and Huh-7 cells were seeded into 6-well plates
at a density of 2.0×104 cells/well and transfected with
50 nM synthetic oligonucleotides or vectors using
Lipofectamine® 2000 (cat. no. 11668-109; Invitrogen;
Thermo Fisher Scientific, Inc.). After HCC cells were transfected
for 12 h and transfection efficiencies were measured (Fig. S1), cells were treated with 5
µg/ml propofol for 24 h at 37°C.
MTT assay
Cell proliferation was determined using a MTT assay.
Hep3B and Huh-7 cells were seeded into 96-well plates at a density
of 1×103 cells/well and incubated overnight. After the
aforementioned transfection and treatment, 20 µl MTT (5
mg/ml; cat. no. M1020; Beijing Solarbio Science & Technology
Co., Ltd.) was added into each well at 24, 48 and 72 h, and then
further incubated at 37°C for an additional 4 h, followed by
centrifugation at 37°C for 10 min at a speed of 1,000 × g.
Subsequently, the supernatant was discarded and 150 µl DMSO
was added to dissolve the formazan crystals. The optical density
(OD) value at 570 nm was measured via a microplate reader (BioTek
Instruments, Inc.).
Flow cytometry analysis
The apoptosis of HCC cells was assessed via an
Annexin V- FITC/propidium iodide (PI) Apoptosis Detection kit (cat.
no. A211-01/02; Vazyme Biotech Co., Ltd.). After relevant
transfection and treatment, HCC cells were harvested and
resuspended at a concentration of 1.0×106 cells/ml.
Then, 5 µl AnnexinV-FITC and 5 µl PI were added into
the cell suspension to the stain of cells. After 15 min of
incubation in the dark at room temperature, the number of apoptotic
cells was analyzed within 1 h via a flow cytometry (FACSCalibur; BD
Biosciences) with software FlowJo 7.6.1 (FlowJo LLC). The number of
apoptotic cells was the sum of the early apoptosis number (the
lower right quadrant) and the late apoptosis number (the upper
right quadrant).
Western blotting
After relevant transfection and treatment, Hep3B and
Huh-7 cells were harvested and lysed in RIPA buffer (cat. no.
R0010; Beijing Solarbio Science & Technology Co., Ltd.)
containing protease inhibitor (cat. no. P8340 MSDS; Sigma-Aldrich;
Merck KGaA) and phosphatase inhibitor (cat. no. P2745 MSDS;
Sigma-Aldrich; Merck KGaA) to isolate total protein. Then, a
bicinchoninic acid protein quantification kit (cat. no. E112-01/02;
Vazyme Biotech Co., Ltd.) was used to examine the concentration of
proteins. Next, a total of 50 µg proteins were separated by
10% SDS-PAGE (cat. no. P1200; Beijing Solarbio Science &
Technology Co., Ltd.) and transferred onto PVDF membranes (cat. no.
PVM020C-160; Pall Corporation). The membranes were blocked with 5%
non-fat milk for 2 h at room temperature and then incubated with
primary antibodies against Bcl-2 (cat. no. ab182858; 1:2,000;
Abcam), cleaved-caspase 3 (c-caspase 3; cat. no. ab2302; 1:2,000;
Abcam), proliferating cell nuclear antigen (PCNA; cat. no.
ab152114; 1:2,000; Abcam), cyclinD1 (cat. no. ab190564; 1:5,000;
Abcam), C-Myc (cat. no. Ab39688; 1:1,000; Abcam), Bax (cat. no.
ab59348; 1:1,000; Abcam), c-poly(ADP-ribose) polymerase 1 (PARP;
cat. no. ab4380; 1:1,000; Abcam), E-cadherin (cat. no. ab76055;
1:1,000; Abcam), N-cadherin (cat. no. ab18203; 1:1,000; Abcam),
Vimentin (cat. no. ab45939; 1:1,000; Abcam) or GAPDH (cat. no.
ab8245; 1:5,000; Abcam) overnight at 4°C. Subsequently, the
membranes were probed with corresponding horseradish peroxidase
(HRP)-conjugated secondary antibody (cat. no. ab150117; 1:5,000;
Abcam) for 1 h at room temperature. The protein bands were
determined using an enhanced chemiluminescence reagent (cat. no.
E411-03/04/05; Vazyme Biotech Co., Ltd.) and analyzed using the
software ImageJ v1.8.0 (National Institutes of Health).
Transwell assay
Following relevant transfection and treatment, HCC
cells were collected. For the detection of cell migration,
collected HCC cells were suspended in medium without serum and 500
µl cell suspension (5.0×105 cells/ml) was seeded
into the upper chamber of a 24-well Transwell (cat. no. 3379; 8
µm; Corning, Inc.). The bottom chamber was plated with 600
µl DMEM with 10% FBS. After 24 h of incubation at 37°C,
cells that remained on the upper chamber were removed using a
cotton swab and cells that migrated were fixed with 90% methanol
for 30 min at 37°C and stained with 0.1% crystal violet for 15 min
at 37°C (cat. no. IC0600; Beijing Solarbio Science & Technology
Co., Ltd.). The migrated cells were counted under a light
microscope at ×100 magnification.
For the detection of cell invasion, the upper
chamber was coated with Matrigel (cat. no. 356234; Beijing Solarbio
Science & Technology Co., Ltd.) for 30 min at 37°C and the
other procedures were the same as aforementioned. The numbers of
migrated and invaded cells were represented by the average number
of cells in five randomly selected fields.
In vivo experiment
The suspension of Hep3B cells (1×106
cells) was subcutaneously injected into the right leg near the
abdomen of BALB/c nude mice (n=12; Shanghai SLAC Laboratory Animal
Co., Ltd.; age, 5 weeks; sex, female; weight, 19-21 g). The mice
were housed under pathogen-free conditions with a 12-h light/dark
cycle at 27°C and 45% humidity and fed sterile fodder and drinking
water. All inoculations were performed under anesthesia with
isoflurane. The parameters of anesthesia were monitored as follows:
i) Ocular reflex, nystagmus indicated if the anesthesia was too
shallow, the pupil dilated during anesthesia excitement period and
the would pupil dilate excessively if the anesthesia was excessive;
ii) eyelid reflex, when the medial or lateral canthus was touched,
the mice blinked and this reflex disappeared during surgical
anesthesia; iii) pinch reflex, in the absence of deep anesthesia,
pinching or clamping the tail of the mouse would result in a flick
of the tail and occasional sound. When the tumor volume reached
70-80 mm3, the mice were randomly divided into two
groups: Mock group (n=6) and propofol group (n=6). The propofol
group was intraperitoneally injected with 100 mg/kg propofol
(Sigma-Aldrich; Merck KGaA) every day for 28 days, and the Mock
group was intraperitoneally injected with an equal volume of saline
(30). No side effects were
observed after relevant administration. The tumor volume was
examined every 4 days and calculated with the following formula:
(Length × width2)/2. After 28 days, the mice were
euthanized by cervical dislocation, and the tumor tissues were
collected and weighed. Approval was obtained from the Ethics
Committee of Animal Research of the Third Affiliated Hospital of
Sun Yat-Sen University.
Reverse transcription-quantitative PCR
(RT-qPCR)
After relevant transfection and treatment, total RNA
was isolated from Hep3B and Huh-7 cells using TRIzol®
reagent (cat. no. R0016; Beyotime Institute of Biotechnology). The
concentrations and quality of RNAs were examined using NanoDrop
2000c spectrophotometer (Thermo Fisher Scientific, Inc.). For RNA
extracted from Hep3B cells, the concentration was 324 ng/µl
(OD260/OD280=1.86; OD260/OD230=2.26). For RNA extracted from Huh-7
cells, the concentration was 258 ng/µl (OD260/OD280=1.94;
OD260/OD230=2.19). The RT experiment was performed using
PrimeScript™ RT reagent kit (cat. no. 6215A; Takara Biotechnology
Co., Ltd.) or miRNA 1st Strand cDNA Synthesis kit (cat. no.
MR101-01/02; Vazyme Biotech Co., Ltd.) under the conditions of 50°C
for 15 min followed by 85°C for 5 sec. Then, RT-qPCR was conducted
using BeyoFast™ SYBR Green qPCR Mix (cat. no. D7260; Beyotime
Institute of Biotechnology) on the ABI 7500 RT PCR system (cat. no.
4351104; Applied Biosystems; Thermo Fisher Scientific, Inc.). The
thermocycling conditions of qPCR reaction were: Initial
denaturation at 95°C for 5 min, followed by 40 cycles of 95°C for
30 sec and 60°C for 45 sec. The expression levels of HOXA11-AS and
miR-4458 were measured using the 2−ΔΔCq method (31). GAPDH or small nuclear RNA U6 were
used as an internal reference. The primers were: HOXA11-AS forward
(F), 5′-GAG TGT TGG CCT GTC CTC -3′ and reverse (R), 5′-TTG TGC CCA
GTT GCC TGT AT-3′; miR-4458 F, 5′-AGA GGT AGG TGT GGA AGA A-3′ and
R, 5′-GCG AGC ACA GAA TTA ATA CGA C-3′; GAPDH F, 5′-CGA GCC ACA TCG
CTC AGA CA-3′ and R, 5′-GTG GTG AAG ACG CCA GTG GA-3′; and U6 F,
5′-AGA GCC TGT GGT GTC CG-3′ and R, 5′-CAT CTT CAA AGC ACT TCC
CT-3′.
Dual-luciferase reporter assay
The potential target of HOXA11-AS was predicted
using software starBase 2. 0 (Sun Yat-sen University) The fragments
of HOXA11-AS containing the predicated wild-type (WT) or mutant
(MUT) complementary sequences of miR-4458 were amplified and cloned
into pmirGLO plasmids (cat. no. E1330; Promega Corporation) to
construct luciferase reporter vectors HOXA11-AS WT and HOXA11-AS
MUT, respectively. HCC cells (2×104) were seeded in
24-well plates and 100 ng indicated vector was transfected into HCC
cells in combination with 50 nM miR-4458 or miR-NC using
Lipofectamine® 2000 (cat. no. 11668-109; Invitrogen;
Thermo Fisher Scientific, Inc.). After co-transfection for 48 h, a
dual-luciferase reporter assay kit (cat. no. DL101-01; Vazyme
Biotech Co., Ltd.) was used to determine luciferase activity.
Renilla luciferase activity was normalized to Firefly luciferase
activity.
RNA pull-down assay
The Pierce Magnetic RNA-protein pull-down kit (cat.
no. 20164; Thermo Fisher Scientific, Inc.) was used to conduct RNA
pull-down assay according to the manufacturer's instructions. Then,
50 µM biotinylated miR-4458 (Bio-miR-4458) or its control
(Bio-NC) containing putative binding sites of HOXA11-AS were
transfected into HCC cells using Lipofectamine® 2000
(cat. no. 11668-109; Invitrogen; Thermo Fisher Scientific, Inc.).
After 24 h, these cells (1×106 cells) were incubated
with streptavidin-coated magnetic beads (Invitrogen; Thermo Fisher
Scientific, Inc.) for 2 h at room temperature. Bound RNAs were
isolated using TRIzol® reagent after biotin-coupled RNA
complex was pulled down. The enrichment of HOXA11-AS was determined
via RT-qPCR as aforementioned.
Statistical analysis
The experiments in this study were performed three
times and all data are presented as the mean ± SD. Data analysis
was performed using GraphPad Prism 7 software (GraphPad Software,
Inc.). Unpaired Student's t-test or one-way ANOVA followed by
Tukey's test were used to analyze the data. P<0.05 was
considered to indicate a significantly significant difference.
Results
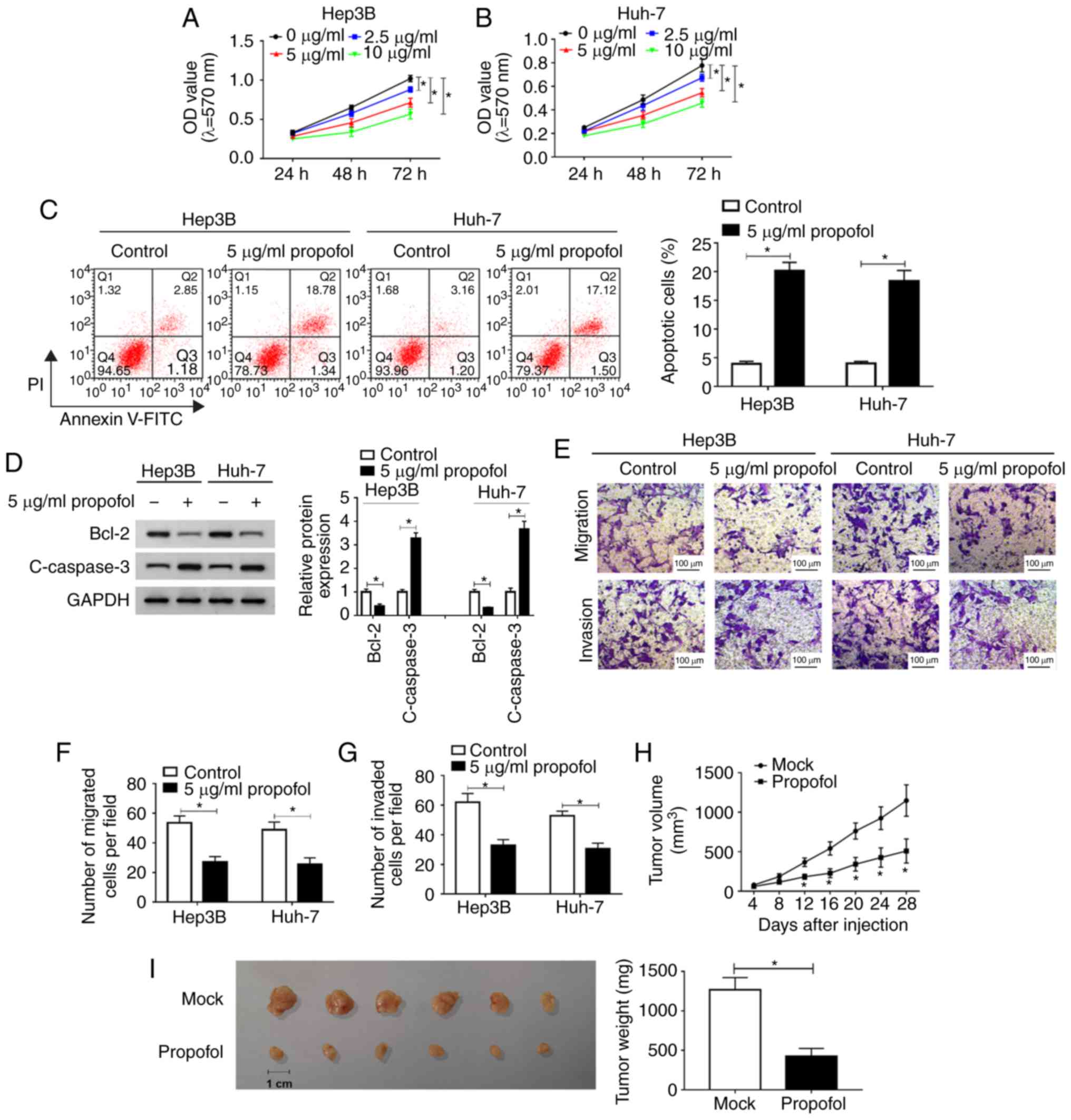
Propofol suppresses HCC cell
proliferation, migration and invasion, and promotes apoptosis in
vitro, as well as inhibits tumor growth in vivo
To identify the function of propofol in HCC in
vitro, Hep3B cells and Huh-7 cells were untreated or treated
with different concentrations of propofol (2.5, 5 or 10
µg/ml) and then an MTT assay was conducted. The
proliferation of Hep3B and Huh-7 cells was significantly inhibited
by propofol in a dose-dependent manner (Fig. 1A and B). It was also found that
cell proliferation was ~80% of that in the control group after 5
µg/ml propofol treatment. Thus, 5 µg/ml propofol was
selected to treated Hep3B and Huh-7 cells in subsequent
experiments.
The results of flow cytometry analysis indicated
that cell apoptosis was significantly promoted by propofol
treatment in Hep3B and Huh-7 cells (Fig. 1C). Moreover, the expression levels
of apoptotic-related proteins Bcl-2 and c-caspase 3 were determined
using western blotting. It was demonstrated that Bcl-2 expression
was significantly decreased, while c-caspase 3 expression was
increased significantly after propofol treatment in both Hep3B and
Huh-7 cells (Fig. 1D). Propofol
treatment also significantly suppressed the migration and invasion
of Hep3B and Huh-7 cells, as identified by the Transwell assay
(Fig. 1E-G).
Then, the effect of propofol was evaluated in
vivo. After propofol treatment, tumor volume was significantly
decreased; in Mock group, the diameter ranged between 1.17-1.44 and
the volume ranged between 682.3-1,274.9 mm3, while in
propofol group, the diameter ranged between 0.71-1.12 cm and the
volume was 174.0-584.1 mm3 (Fig. 1H). The mice were euthanized after
Hep3B cells were injected for 28 days and tumor weight was
measured. The results suggested that the tumor weight was
significantly reduced after propofol treatment, compared with the
Mock group (Fig. 1I).
Collectively, these findings indicated that propofol treatment
inhibited HCC cell progression in vitro and tumor growth
in vivo.
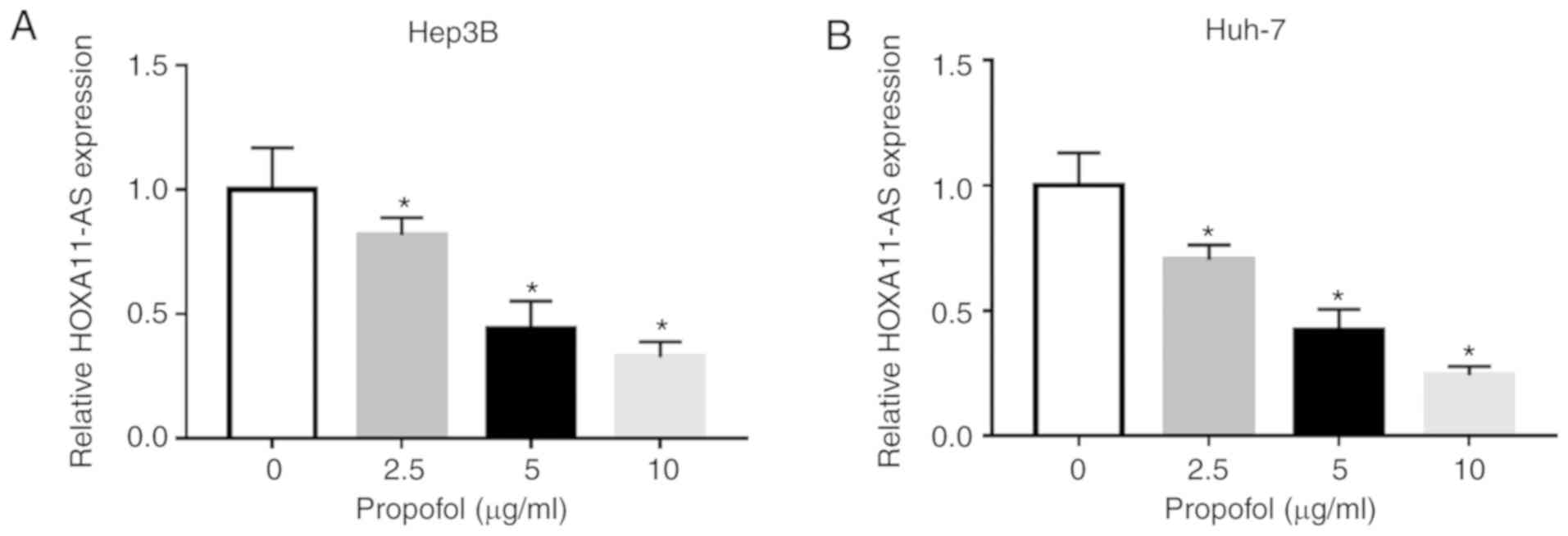
Propofol treatment significant decreases
HOXA11-AS expression in HCC cells
To assess the potential mechanism of propofol on the
features of HCC cells, Hep3B and Huh-7 cells were untreated or
treated with different doses of propofol (2.5, 5 or 10
µg/ml) for 24 h. It was identified that the expression of
HOXA11-AS was significantly decreased after propofol treatment in a
concentration-dependent manner in both Hep3B and Huh-7 cells
compared with untreated cells (Fig.
2A and B). Thus, it was speculated that propofol may restrain
cell proliferation, migration and invasion, as well as induce cell
apoptosis via the downregulation of HOXA11-AS in HCC.
HOXA11-AS overexpression weakens the effects on cell
proliferation, apoptosis, migration and invasion mediated by
propofol in HCC. It was demonstrated that HOXA11-AS trans-fection
resulted in a significantly elevation in HOXA11-AS expression in
both Hep3B and Huh-7 cells (Fig.
S1A). Next, to investigate whether propofol could regulate the
features of HCC cells by downregulating HOXA11-AS, Hep3B and Huh-7
cells were untreated or treated with propofol, propofol + Vector or
propofol + HOXA11-AS. RT-qPCR results suggested that the reduction
of HOXA11-AS expression caused by propofol was partially reversed
after HOXA11-AS overexpression in both Hep3B and Huh-7 cells
(Fig. 3A).
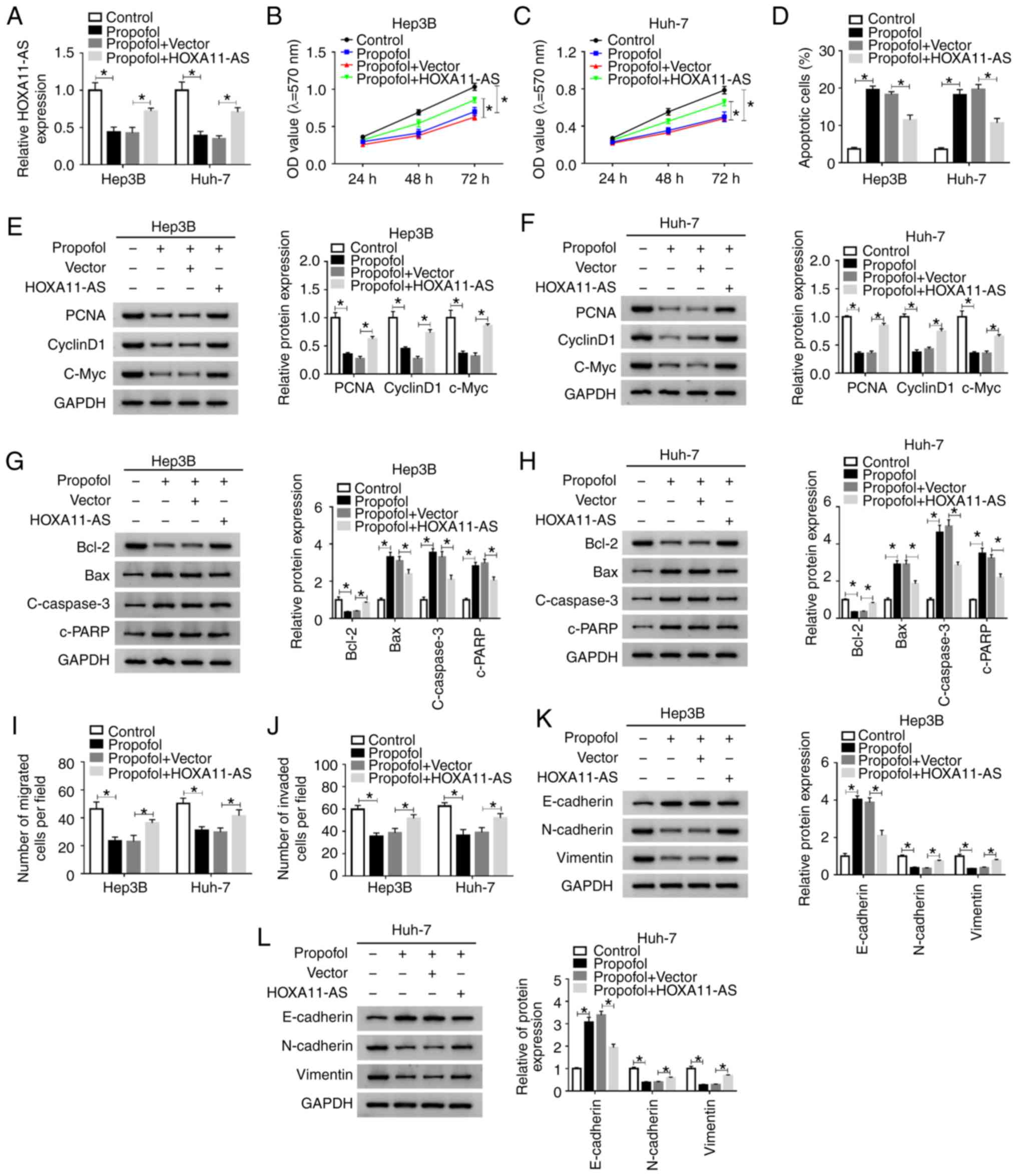 | Figure 3HOXA11-AS restores the effects of
propofol on cell proliferation, apoptosis, migration and invasion
in HCC. Hep3B and Huh-7 cells were untreated or treated with
propofol, propofol + Vector or propofol + HOXA11-AS. (A) Expression
of HOXA11-AS in Hep3B and Huh-7 cells was determined using reverse
transcription-quantitative PCR. Proliferation of (B) Hep3B and (C)
Huh-7 cells was evaluated using MTT assay. (D) Apoptosis of Hep3B
and Huh-7 cells was analyzed via flow cytometry analysis. Protein
expression levels of PCNA, cyclinD1 and C-Myc in (E) Hep3B and (F)
Huh-7 cells were analyzed using western blotting. Protein
expression levels of Bcl-2, Bax, c-caspase3 and c-PRRP in (G) Hep3B
and (H) Huh-7 cells were analyzed using western blotting. (I)
Migration and (J) invasion of Hep3B and Huh-7 cells were assessed
using Transwell assay. Protein expression levels of E-cadherin,
N-cadherin and Vimentin in (K) Hep3B and (L) Huh-7 cells were
analyzed using western blotting. Experiments were repeated three
times. *P<0.05. HOXA11-AS, HOMEOBOX A11 antisense
RNA; c-, cleaved; PCNA, proliferating cell nuclear antigen; PARP,
poly(ADP-ribose) polymerase 1. |
MTT assay results indicated that cell proliferation
was significantly suppressed by propofol treatment, but this effect
was partly reversed by HOXA11-AS transfection in Hep3B and Huh-7
cells (Fig. 3B and C). Moreover,
as indicated by flow cytometry analysis, the promotional effect on
cell apoptosis mediated by propofol was partially abolished by
HOXA11-AS transfection in Hep3B and Huh-7 cells (Figs. 3D and S2A). It was also demonstrated that the
protein expression levels of PCNA, cyclinD1, c-Myc and Bcl-2 were
significantly reduced, while the protein expression levels of Bax,
c-caspase 3 and c-PARP were significantly elevated by propofol in
Hep3B and Huh-7 cells; however, these effects were all reversed by
HOXA11-AS transfection (Fig.
3E-H). The inhibitory effects of propofol treatment on cell
migration and invasion were also partly abrogated after the
overexpression of HOXA11-AS in Hep3B and Huh-7 cells, as indicated
by Transwell assay (Figs. 3I and
J, S2B and C). Moreover, it
was found that propofol treatment increased E-cadherin expression
and decreased N-cadherin and Vimentin expression levels in Hep3B
and Huh-7 cells, while the effects were reversed by HOXA11-AS
(Fig. 3K and L). Therefore, these
results suggested that propofol suppressed cell progression and
metastasis by downregu-lating HOXA11-AS in HCC cells.
HOXA11-AS negatively regulates the
expression of miR-4458 via direct interaction in HCC cells
By searching the online software starBase 2.0, it
was found that miR-4458 was a target of HOXA11-AS, and their
complementary binding sites are presented in Fig. 4A. RT-qPCR assay results
demonstrated that miR-4458 transfected led to a significant
elevation in miR-4458 expression in Hep3B and Huh-7 cells,
indicating the successful transfection of miR-4458 (Fig. S1C). To assess this prediction, a
dual-luciferase reporter assay was conducted. The results suggested
that the luciferase activity was significantly suppressed in
HOXA11-AS WT + miR-4458 co-transfected Hep3B and Huh-7 cells
compared with HOXA11-AS WT + miR-NC co-transfected cells, while
co-transfection of HOXA11-AS MUT + miR-4458 or miR-NC did not
affect the luciferase activity (Fig.
4B and C). RNA pull-down assay identified that Bio-miR-4458
pulled down HOXA11-AS to a greater extent compared with Bio-NC in
Hep3B and Huh-7 cells, which further support the prediction
interaction (Fig. 4D).
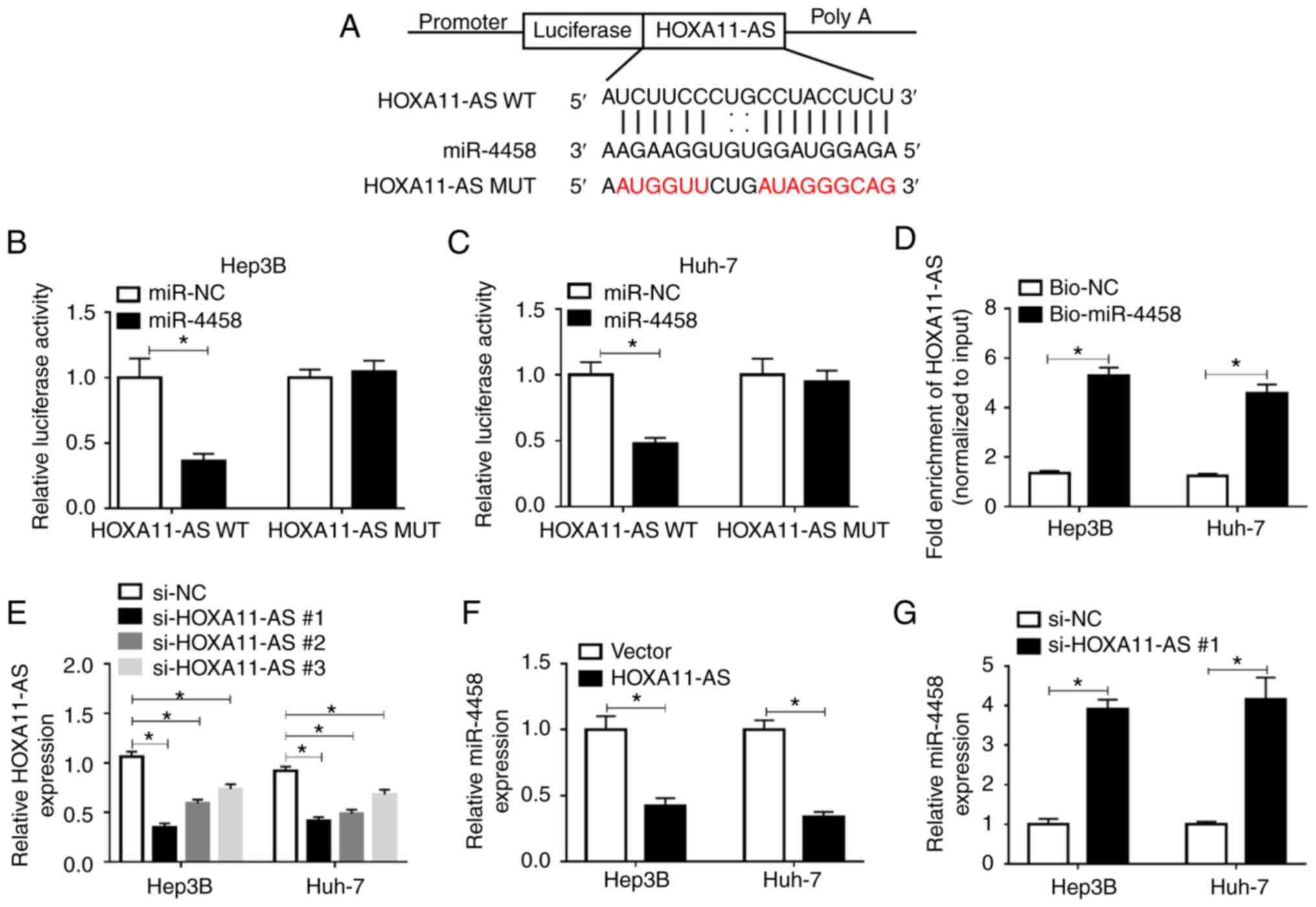 | Figure 4HOXA11-AS negatively regulates
miR-4458 expression via direct targeting in HCC cells. (A) Putative
binding sites between HOXA11-AS and miR-4458 were predicted using
starBase 2.0. Luciferase activity in HOXA11-AS WT or HOXA11-AS MUT
and miR-4458 or miR-NC co-transfected (B) Hep3B and (C) Huh-7 cells
was detected via dual-luciferase reporter assay. (D) RNA pull-down
assay was conducted to assess the interaction between HOXA11-AS and
miR-4458, and the expression of HOXA11-AS was detected using
RT-qPCR after Bio-miR-4458 or Bio-NC was transfected into Hep3B and
Huh-7 cells. (E) Expression of HOXA11-AS in Hep3B and Huh-7 cells
transfected with si-NC, si-HOXA11-AS#1, si-HOXA11-AS#2 or
si-HOXA11-AS#3 was determined via RT-qPCR. (F) Expression of
miR-4458 in Hep3B and Huh-7 cells transfected with Vector or
HOXA11-AS was determined via RT-qPCR. (G) Expression of miR-4458 in
Hep3B and Huh-7 cells transfected with si-NC or si-HOXA11-AS#1 was
measured using RT-qPCR. Experiments were repeated three times.
*P<0.05. HOXA11-AS, HOMEOBOX A11 antisense RNA; NC,
negative control; miR, microRNA; WT, wild-type; MUT, mutant;
RT-qPCR, reverse transcription-quantitative PCR; bio, Biotinylated;
si, small interfering RNA. |
Subsequently, three HOXA11-AS knockdown vectors
(si-HOXA11-AS#1, si-HOXA11-AS#2 and si-HOXA11-AS#3) were
constructed and then transfected into Hep3B and Huh-7 cells.
RT-qPCR was performed to evaluate the knockdown efficiency. The
data suggested that HOXA11-AS was significantly downregulated after
HOXA11-AS knockdown vectors were transfected in Hep3B and Huh-7
cells compared with si-NC groups; si-HOXA11-AS#1 was selected for
the subsequent experiments due to its stronger inhibitory effects
on HOXA11-AS expression compared with si-HOXA11-AS#2 and
si-HOXA11-AS#3 (Fig. 4E).
HOXA11-AS overexpression led to a significant reduction in miR-4458
expression, while HOXA11-AS downregulation resulted in a
significant elevation in miR-4458 expression in both Hep3B and
Huh-7 cells (Fig. 4F and G).
Thus, HOXA11-AS could target miR-4458 and negatively regulate
miR-4458 expression in HCC cells.
miR-4458 inhibition attenuates the
effects of HOXA11-AS knockdown on cell proliferation, apoptosis,
migration and invasion in HCC cells
Anti-miR-4458 transfection significantly decreased
the expression of miR-4458 in Hep3B and Huh-7 cells (Fig. S1B). Subsequently, in order to
identify whether HOXA11-AS could affect cell progression by
regulating the expression of miR-4458 in HCC, si-NC,
si-HOXA11-AS#1, si-HOXA11-AS#1 + anti-miR-NC or si-HOXA11-AS#1 +
anti-miR-4458 were transfected into Hep3B and Huh-7 cells. The
transfection efficiency was determined using RT-qPCR, and the
results demonstrated that si-HOXA11-AS#1 led to a significant
increase in miR-4458 expression, while anti-miR-4458 reversed this
effect in Hep3B and Huh-7 cells (Fig.
5A).
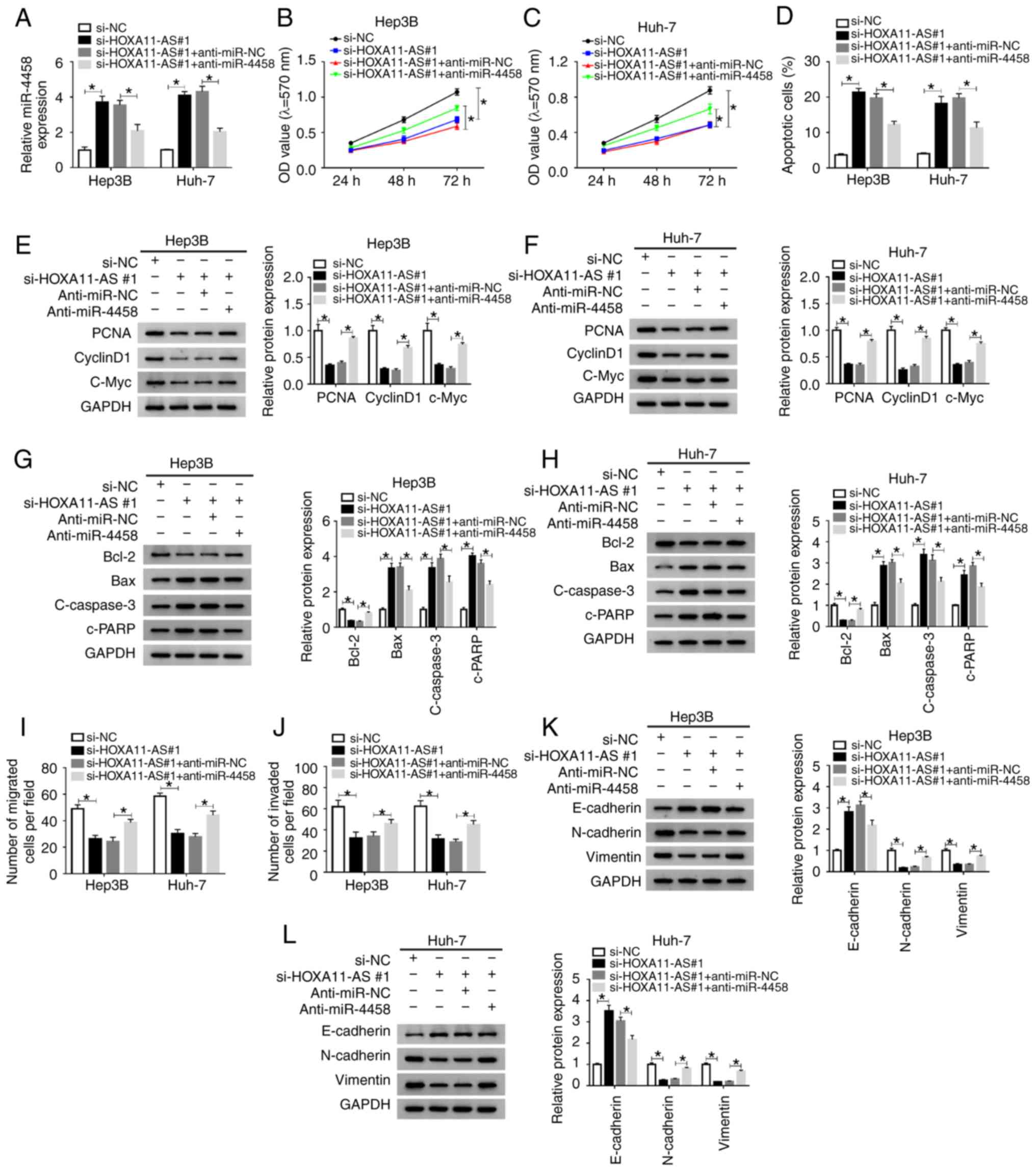 | Figure 5miR-4458 inhibition abolishes the
effects of HOXA11-AS knockdown on cell progression in HCC cells.
Hep3B and Huh-7 cells were transfected with si-NC, si-HOXA11-AS#1,
si-HOXA11-AS#1+anti-miR-NC or si-HOXA11-AS#1+anti-miR-4458. (A)
Expression of miR-4458 in Hep3B and Huh-7 cells was measured using
reverse transcription-quantitative PCR. Cell proliferation was
analyzed by MTT assay in (B) Hep3B and (C) Huh-7 cells. (D) Cell
apoptosis was detected via flow cytometry analysis. Protein
expression levels of PCNA, cyclinD1 and C-Myc in (E) Hep3B and (F)
Huh-7 cells were analyzed using western blotting. Protein
expression levels of Bcl-2, Bax, c-caspase3 and c-PRRP in (G) Hep3B
and (H) Huh-7 cells were analyzed using western blotting. Cell (I)
migration and (J) invasion were assessed using Transwell assay.
Protein expression levels of E-cadherin, N-cadherin and Vimentin in
(K) Hep3B and (L) Huh-7 cells were analyzed using western blotting.
Experiments were repeated three times. *P<0.05.
HOXA11-AS, HOMEOBOX A11 antisense RNA; NC, negative control; miR,
microRNA; si, small interfering RNA; OD, optical density. |
MTT assay results indicated that knockdown of
HOXA11-AS significantly suppressed cell proliferation, while
miR-4458 knockdown reversed this suppression in Hep3B and Huh-7
cells (Fig. 5B and C). Flow
cytometry analysis indicated that propofol promoted the apoptosis
of Hep3B and Huh-7 cells, while HOXA11-AS overexpression reversed
the impact. Moreover, the promotional effect on cell apoptosis
caused by si-HOXA11-AS#1 was significantly rescued by anti-miR-4458
in Hep3B and Huh-7 cells (Figs.
5D and S3A). The protein
expression levels of PCNA, CyclinD1, c-Myc and Bcl-2 were
significantly reduced and the protein expression levels of Bax,
c-caspase 3 and c-PARP were significantly elevated by silencing of
HOXA11-AS in Hep3B and Huh-7 cells, but these effects were reversed
by inhibition of miR-4458 (Fig.
5E-H). The Transwell assay results suggested that propofol
treatment repressed the migration and invasion of Hep3B and Huh-7
cells, while the overexpression of HOXA11-AS reversed the effects.
Furthermore, the inhibition of cell migration and invasion mediated
by si-HOXA11-AS#1 was effectively reversed by anti-miR-4458 in
Hep3B and Huh-7 cells (Figs. 5I and
J, S3B and C). In addition,
it was demonstrated that HOXA11-AS knockdown increased E-cadherin
expression and decreased N-cadherin and Vimentin expression levels
in Hep3B and Huh-7 cells compared with si-NC control groups, while
miR-4458 inhibition restored the impacts (Fig. 5K and L). These data indicated that
HOXA11-AS regulated cell progression by targeting miR-4458 in HCC
cells.
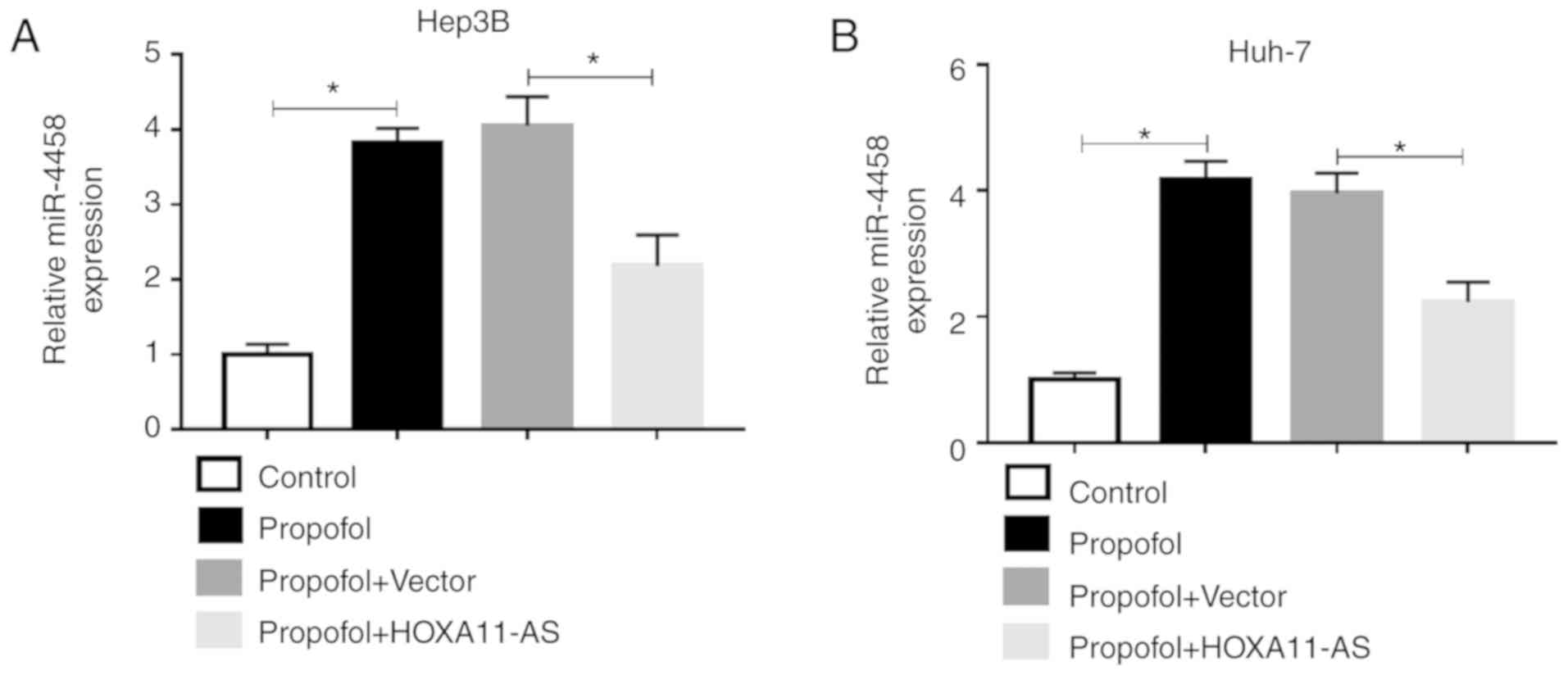
HOXA11-AS reverses the upregulation of
miR-4458 caused by propofol in HCC cells
To investigate the effect of propofol on miR-4458
expression in HCC cells and the relationship between the expression
of HOXA11-AS1 and miR-4458 in HCC cells treated with propofol,
Hep3B and Huh-7 cells were untreated or treated with propofol,
propofol + Vector or propofol + HOXA11-AS. RT-qPCR results
demonstrated that the expression of miR-4458 was significantly
increased by propofol, while this effect was partly reversed by
HOXA11-AS overexpression in Hep3B and Huh-7 cells (Fig. 6A and B). Therefore, these results
suggested that propofol could regulate miR-4458 expression via
HOXA11-AS in HCC cells.
Discussion
The metastasis and recurrence of cancer types are
the most fundamental factors affecting prognosis (32). Numerous studies have reported that
propofol affects the metastasis and recurrence of tumors in
multiple cancer types, including HCC (14,33). Therefore, investigating the
functions of propofol in the proliferation, metastasis and
apoptosis of HCC cells is not only of great theoretical
significance, but also important with regards to scientific
research and clinical application. In the present study, it was
found that propofol could suppress cell progression via
downregulating HOXA11-AS and upregulating miR-4458 in HCC.
Ou et al (14) reported that propofol led to an
inhibition in HCC cell proliferation and metastasis and a promotion
in HCC cell apoptosis. Furthermore, Zhang et al (33) demonstrated that propofol could
suppress cell proliferation and induced cell apoptosis in HCC,
while Liu et al (34) also
revealed that propofol suppressed HCC cell proliferation and
metastasis, and induced HCC apoptosis. Consistent with these
reports, the present results suggested that there were significant
suppressive effects on cell proliferation and metastasis, as well
as a significant promotional effect on cell apoptosis after
propofol treatment in vitro in HCC. Moreover, tumor growth
was significantly suppressed in vivo. Collectively, these
findings indicated that propofol has a tumor suppressor effect on
HCC.
Reduced expression levels of lncRNAs mediated by
propofol have been revealed in human cancer types. For instance,
HOXA11-AS expression is reduced by propofol in colorectal cancer
(35). Propofol also induces the
apoptosis of cervical cancer cells by repressing HOX transcript
antisense RNA (36). In addition,
Zhan et al (22) reported
that HOXA11-AS exerted its promotional role in HCC progression by
targeting miR-214-3p. Yu et al (37) have also shown that HOXA11-AS
silencing could repress HCC cell viability and induce apoptosis
in vitro, as well as inhibit tumor growth in vivo.
Based on these findings, it was hypothesized that HOXA11-AS
expression could be regulated by propofol treatment and that
propofol-induced HOXA11-AS could regulate HCC progression. The
current results indicated that HOXA11-AS was suppressed by propofol
treatment in a dose-dependent manner. Moreover, HOXA11-AS
upregulation reduced the effects of propofol on HCC features,
suggesting that HOXA11-AS could promote the development of HCC
cells; these present study findings were consistent with previous
reports.
To the best of our knowledge, the current study was
the first to identify miR-4458 as a target of HOXA11-AS in HCC
using dual-luciferase reporter and RNA pull-down assays. A previous
study showed that miR-4458 was weakly expressed in HCC and its
overexpression inhibited HCC cell proliferation and metastasis, as
well as contributed to HCC cell apoptosis (26). The present results indicated that
the inhibition of miR-4458 significantly attenuated the inhibitory
effect of HOXA11-AS knockdown on cell progression in HCC,
suggesting that miR-4458 may serve a tumor suppression effect in
HCC. Previous studies have reported that several miRNAs are
elevated by propofol in HCC, such as miR-219-5p (28), miR-199a (33) and miR-142-3p (38). Moreover, the current study
identified that miR-4458 was stimulated by propofol in HCC cells,
while HOXA11-AS reversed this effect.
However, the present study had some limitations. For
example, it was not verified whether HOXA11-AS regulated
tumorigenesis of HCC in vivo. Moreover, the potential target
genes of miR-4458 should be examined in future experiments.
In conclusion, propofol inhibited cell progression
in vitro and tumor growth in vivo in HCC.
Furthermore, propofol affected the features of HCC cells via the
HOXA11-AS/miR-4458 axis in HCC. Therefore, this study demonstrated
the potential of propofol during the surgery of patients with HCC
and may provide a novel insight for HCC treatment in the
future.
Supplementary Data
Acknowledgments
Not applicable.
Funding
No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
FS and YJ conceived and designed the study. JL and
YF performed data analyses and interpretation. FS and YF co-wrote
the manuscript. All authors have read and approved the final
manuscript.
Ethics approval and consent to
participate
Ethical approval was obtained from the Ethics
Committee of Animal Research of the Third Affiliated Hospital of
Sun Yat-Sen University.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Bruix J and Sherman M; American
Association for the Study of Liver Diseases: Management of
hepatocellular carcinoma: An update. Hepatology. 53:1020–1022.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Mittal S and El-Serag HB: Epidemiology of
HCC: Consider the population. J Clin Gastroenterol. 47:S2–S6. 2013.
View Article : Google Scholar :
|
|
3
|
Bray F, Ferlay J, Soerjomataram I, Siegel
RL, Torre LA and Jemal A: Global cancer statistics 2018: GLOBOCAN
estimates of incidence and mortality worldwide for 36 cancers in
185 countries. CA Cancer J Clin. 68:394–424. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Cidon EU: Systemic treatment of
hepatocellular carcinoma: Past, present and future. World J
Hepatol. 9:797–807. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Yu JJ, Xiao W, Dong SL, Liang HF, Zhang
ZW, Zhang BX, Huang ZY, Chen YF, Zhang WG, Luo HP, et al: Effect of
surgical liver resection on circulating tumor cells in patients
with hepatocellular carcinoma. BMC Cancer. 18:8352018. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Suh SW and Choi YS: Influence of liver
fibrosis on prognosis after surgical resection for resectable
single hepatocellular carcinoma. ANZ J Surg. 89:211–215. 2019.
View Article : Google Scholar
|
|
7
|
Ye Z, Jingzhong L, Yangbo L, Lei C and
Jiandong Y: Propofol inhibits proliferation and invasion of
osteosarcoma cells by regulation of microRNA-143 expression. Oncol
Res. 21:201–207. 2013. View Article : Google Scholar
|
|
8
|
Huang X, Teng Y, Yang H and Ma J: Propofol
inhibits invasion and growth of ovarian cancer cells via regulating
miR-9/NF-kappaB signal. Braz J Med Biol Res. 49:e57172016.
View Article : Google Scholar
|
|
9
|
Meng C, Song L, Wang J, Li D, Liu Y and
Cui X: Propofol induces proliferation partially via downregulation
of p53 protein and promotes migration via activation of the nrf2
pathway in human breast cancer cell line MDA-MB-231. Oncol Rep.
37:841–848. 2017. View Article : Google Scholar
|
|
10
|
Chidambaran V, Costandi A and D'Mello A:
Propofol: A review of its role in pediatric anesthesia and
sedation. CNS Drugs. 29:543–563. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Zhang L, Wang N, Zhou S, Ye W, Jing G and
Zhang M: Propofol induces proliferation and invasion of gallbladder
cancer cells through activation of Nrf2. J Exp Clin Cancer Res.
31:662012. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Wang XY, Li YL, Wang HY, Zhu M, Guo D,
Wang GL, Gao YT, Yang Z, Li T, Yang CY and Chen YM: Propofol
inhibits invasion and proliferation of C6 glioma cells by
regulating the Ca2+ permeable AMPA receptor-system xc-
pathway. Toxicol In Vitro. 44:57–65. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Liu Z, Zhang J, Hong G, Quan J, Zhang L
and Yu M: Propofol inhibits growth and invasion of pancreatic
cancer cells through regulation of the miR-21/slug signaling
pathway. Am J Transl Res. 8:4120–4133. 2016.PubMed/NCBI
|
|
14
|
Ou W, Lv J, Zou X, Yao Y, Wu J, Yang J,
Wang Z and Ma Y: Propofol inhibits hepatocellular carcinoma growth
and invasion through the HMGA2-mediated wnt/β-catenin pathway. Exp
Ther Med. 13:2501–2506. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Fatica A and Bozzoni I: Long non-coding
RNAs: New players in cell differentiation and development. Nat Rev
Genet. 15:7–21. 2014. View
Article : Google Scholar
|
|
16
|
Ponting CP, Oliver PL and Reik W:
Evolution and functions of long noncoding RNAs. Cell. 136:629–641.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Chen F, Li M and Zhu X: Propofol
suppresses proliferation and migration of papillary thyroid cancer
cells by down-regulation of lncRNA ANRIL. Exp Mol Pathol.
107:68–76. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Ming N, Na HST, He JL, Meng QT and Xia ZY:
Propofol alleviates oxidative stress via upregulating
lncRNA-TUG1/Brg1 pathway in hypoxia/reoxygenation hepatic cells. J
Biochem. 166:415–421. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Sun Y and Sun H: Propofol exerts
anticancer activity on hepatocellular carcinoma cells by raising
lncRNA DGCR5. J Cell Physiol. 235:2963–2972. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Xu X, Yin Y, Tang J, Xie Y, Han Z, Zhang
X, Liu Q, Qin X, Huang X and Sun B: Long non-coding RNA myd88
promotes growth and metastasis in hepatocellular carcinoma via
regulating myd88 expression through H3K27 modification. Cell Death
Dis. 8:e31242017. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Yang Y, Chen Q, Piao HY, Wang B, Zhu GQ,
Chen EB, Xiao K, Zhou ZJ, Shi GM, Shi YH, et al: HNRNPAB-regulated
lncRNA-ELF209 inhibits the malignancy of hepatocellular carcinoma.
Int J Cancer. 146:169–180. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Zhan M, He K, Xiao J, Liu F, Wang H, Xia
Z, Duan X, Huang R, Li Y, He X, et al: Lnc RNA HOXA 11-AS promotes
hepatocellular carcinoma progression by repressing miR-214-3p. J
Cell Mol Med. 22:3758–3767. 2018. View Article : Google Scholar :
|
|
23
|
Borel F, Konstantinova P and Jansen PL:
Diagnostic and therapeutic potential of miRNA signatures in
patients with hepa-tocellular carcinoma. J Hepatol. 56:1371–1383.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Wang Y, Yang L, Chen T, Liu X, Guo Y, Zhu
Q, Tong X, Yang W, Xu Q, Huang D and Tu K: A novel lncRNA
MCM3AP-AS1 promotes the growth of hepatocellular carcinoma by
targeting miR-194-5p/FOXA1 axis. Mol Cancer. 18:282019. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Kabir TD, Ganda C, Brown RM, Beveridge DJ,
Richardson KL, Chaturvedi V, Candy P, Epis M, Wintle L, Kalinowski
F, et al: A microRNA-7/growth arrest specific 6/TYRO3 axis
regulates the growth and invasiveness of sorafenib-resistant cells
in human hepatocellular carcinoma. Hepatology. 67:216–231. 2018.
View Article : Google Scholar
|
|
26
|
Tang D, Sun B, Yu H, Yang Z and Zhu L:
Tumor-suppressing effect of miR-4458 on human hepatocellular
carcinoma. Cell Physiol Biochem. 35:1797–1807. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Wang P, Ning S, Zhang Y, Li R, Ye J, Zhao
Z, Zhi H, Wang T, Guo Z and Li X: Identification of
lncRNA-associated competing triplets reveals global patterns and
prognostic markers for cancer. Nucleic Acids Res. 43:3478–3489.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Gong T, Ning X, Deng Z, Liu M, Zhou B,
Chen X, Huang S, Xu Y, Chen Z and Luo R: Propofol-induced
miR-219-5p inhibits growth and invasion of hepatocellular carcinoma
through suppression of GPC3-mediated wnt/β-catenin signalling
activation. J Cell Biochem. 120:16934–16945. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Xu J, Xu W and Zhu J: Propofol suppresses
proliferation and invasion of glioma cells by upregulating
microRNA-218 expression. Mol Med Rep. 12:4815–4820. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Liu Y, Zhang N, Cao Q, Cui X, Zhou Q and
Yang C: The effects of propofol on the growth behavior of hepatoma
xenografts in balb/c mice. Biomed Pharmacother. 90:47–52. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
32
|
Jiang WG, Sanders AJ, Katoh M, Ungefroren
H, Gieseler F, Prince M, Thompson SK, Zollo M, Spano D, Dhawan P,
et al: Tissue invasion and metastasis: Molecular, biological and
clinical perspectives. Semin Cancer Biol. 35:S244–S275. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Zhang J, Wu GQ, Zhang Y, Feng ZY and Zhu
SM: Propofol induces apoptosis of hepatocellular carcinoma cells by
upregulation of microRNA-199a expression. Cell Biol Int.
37:227–232. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Liu SQ, Zhang JL, Li ZW, Hu ZH, Liu Z and
Li Y: Propofol inhibits proliferation, migration, invasion and
promotes apoptosis through down-regulating miR-374a in
hepatocarcinoma cell lines. Cell Physiol Biochem. 49:2099–2110.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Ren YL and Zhang W: Propofol promotes
apoptosis of colorectal cancer cells via alleviating the
suppression of lncRNA HOXA11-AS on miRNA let-7i. Biochem Cell Biol.
98:90–98. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Zhang D, Zhou XH, Zhang J, Zhou YX, Ying
J, Wu GQ and Qian JH: Propofol promotes cell apoptosis via
inhibiting HOTAIR mediated mTOR pathway in cervical cancer. Biochem
Biophys Res Commun. 468:561–567. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Yu J, Hong JF, Kang J, Liao LH and Li CD:
Promotion of LncRNA HOXA11-AS on the proliferation of
hepatocellular carcinoma by regulating the expression of LATS1. Eur
Rev Med Pharmacol Sci. 21:3402–3411. 2017.PubMed/NCBI
|
|
38
|
Zhang J, Shan WF, Jin TT, Wu GQ, Xiong XX,
Jin Hy and Zhu SM: Propofol exerts anti-hepatocellular carcinoma by
microvesicle-mediated transfer of miR-142-3p from macrophage to
cancer cells. J Transl Med. 12:2792014. View Article : Google Scholar : PubMed/NCBI
|