1. Introduction
According to data obtained from the World Health
Organization, cardiovascular diseases accounted for 31% of all
causes of death in 2016, and they are considered as the leading
cause of mortality worldwide (1).
One of the risk factors for cardiovascular diseases is metabolic
syndrome, a multifactorial entity that includes variables such as
obesity, dyslipidemia, hypertension and glucose metabolism
dysfunctions (2). Evidence
connects these variables to increased oxidative stress, with
mitochondrial dysfunction, activation of enzymes that produce
reactive oxygen and nitrogen species (RONS), and impairment of the
activity of antioxidant systems acting as the main triggering
mechanisms (2-4).
As oxidative stress and inflammation are
interrelated and contribute to the initial events of cardiovascular
diseases, antioxidants that modulate redox balance, such as
astaxanthin, may be considered as important regulators of
inflammatory responses (5) and
have become the focus of research to evaluate whether and how they
prevent these diseases. Astaxanthin is closely associated with
other well-known carotenoids, such as beta-carotene, lutein and
zeaxanthin, sharing various of the physiological and metabolic
functions attributed to these compounds (6). The two oxygenated groups, hydroxyl
(OH) and carbonyl (C=O), in each of its ionone rings explains some
of the unique characteristics of astaxanthin, such as its more
potent anti-oxidant activity and polar configuration when compared
to other carotenoids (6-8). In addition to its antioxidant
action, astaxanthin has anti-inflammatory properties and the
ability to modulate lipid and glucose metabolism (6,7,9),
which are beneficial in the cardiovascular system, preventing
disorders such as atherosclerosis, arterial hypertension and
dyslipidemia (10-20). Given the extensive evidence
supporting its health-promoting properties and safety, astaxanthin
was approved as a nutraceutical by the United States Food and Drug
Administration in 1999 (21).
Considering the safety of using astaxanthin as a
nutraceutical and the scientific evidence of its beneficial effects
on cardiovascular physiology, the aim of the present review was to
describe the molecular and cellular mechanisms underlying the
antioxidant and anti-inflammatory role of astaxanthin in the
prevention of cardiovascular diseases.
2. Mechanisms of action of astaxanthin
Antioxidant effect
Cell membrane systems are particularly vulnerable to
RONS attacks due to their content of polyun-saturated fatty acids
(PUFAs) and their metabolic activities, which endogenously generate
other oxidizing metabolites (22).
Astaxanthin protects cell membranes against RONS and
oxidative damage. Due to its chemical structure, its polar groups
overlap the polar regions of the cell membrane, while the central
non-polar region of the molecule fits into the inner non-polar
region of the membrane. Thus, this carotenoid may take on a
transmembrane alignment in biological membranes, helping to
maintain the membrane structure and decrease membrane fluidity, and
acting as an antioxidant (17,23,24).
Astaxanthin scavenges RONS and other reactive
species (sulfur and carbon) directly, both by donating electrons
and by bonding with the free radical to form a non-reactive product
(25). In addition, the presence
of a series of conjugated bonds in the central non-polar region of
astaxanthin enables the molecule to remove free radicals
(high-energy electrons) from the cell interior by transporting them
along its own carbon chain, resembling a 'lightning rod' for these
electrons, so that these are neutralized by other antioxidants
located outside the cell membrane, such as vitamin C (25).
The increased susceptibility of membrane lipids and
low-density lipoprotein (LDL) to oxidation may trigger the
formation of thrombi and the development of atherosclerosis
(5). One of the reactive species
that induces lipid peroxidation and LDL oxidation is peroxynitrite
(ONOO−) (26,27), which is neutralized by astaxanthin
to form 15-nitroastaxanthin, a compound that also has important
antioxidant action (28).
The LDL oxidation time in the presence of
astaxanthin has been analyzed in vitro and ex vivo.
In the in vitro assays, astaxanthin prolonged LDL oxidation
in a dose-dependent manner, in addition to being more effective
compared with lutein and α-tocopherol. In turn, the blood samples
of individuals who were supplemented daily with 1.8, 3.6, 14.4, or
21.6 mg astaxanthin for 14 days evidenced a significant delay in
LDL oxidation when compared to samples collected before
supplementation, the greatest effect being obtained with the dose
of 14.4 mg (oxidation time increased by 5.0, 26.2, 42.3 and 30.7%
with 1.8, 3.6, 14.4 and 21.6 mg astaxanthin, respectively)
(Table I) (10). Thus, it was demonstrated that the
intake of astaxanthin delayed LDL oxidation, one of the key factors
involved in the process of atherosclerosis.
 | Table IClinical studies that have
demonstrated the potential beneficial effects of oral astaxanthin
supplementation on cardiovascular physiology. |
Table I
Clinical studies that have
demonstrated the potential beneficial effects of oral astaxanthin
supplementation on cardiovascular physiology.
| Study type | Subjects (age) | Intervention (no.
of subjects per group) | Mechanism of action
evaluated | Main findings | (Refs.) |
|---|
| Open-label | 24 healthy
volunteers (mean, 28.2±7.8 years) | Control
(n=6)a; AST 1.8 mg/day (n=5);
AST 3.6 mg/day (n=5); AST 14.4 mg/day (n=3);AST 21.6 mg/day (n=5);
14 days | Antioxidant
effect | Delayed LDL
oxidation time | (10) |
| Randomized,
double-blind, placebo-controlled | 27 overweight and
obese adults (BMI>25 kg/m2) (20-55 years) | Placebo (n=13); AST
20 mg/day (n=14); 12 weeks | Antioxidant
effect | Reduced LDL, ApoB
and ApoA1/ApoB ratio relative to baseline; increased TAC and SOD
compared to baseline; reduced lipid peroxidation biomarkers (MDA
and ISP) compared to baseline. | (11) |
| Randomized,
double-blind, placebo-controlled | 30 healthy
individuals (50-69 years) | Placebo (n=10); AST
6 mg/day(n=10); AST 12 mg/day (n=10); 12 weeks | Antioxidant
effect | Reduced
phospholipid hydroperoxide levels in erythrocytes | (12) |
| Randomized,
double-blind, placebo-controlled | 39 healthy men
(19-33 years) | Placebo (n=19); AST
8 mg/day (n=20); 3 months | Antioxidant
effect | Reduced plasma
lipid peroxidation, particularly 12-hydroxy and 15-hydroxy fatty
acids | (13) |
| Open-label,
uncontrolled study | 20 healthy
postmenopausal women with high oxidative stress levels (mean,
55.7±4.8 years) | AST 12 mg/day
(n=20); 8 weeks | Antioxidant
effect | Lowered blood
pressure; increased antioxidant capacity; reduced vascular
resistance in lower limbs and serum adiponectin | (14) |
| Randomized | 39 smokers (≥20
cigarettes per day) and 39 non-smokers (21-43 years) | Control
(n=39)b; AST 5 mg/day (n=13);
AST 20 mg/day (n=13); AST 40 mg/day (n=13); 3 weeks | Antioxidant
effect | Reduced MDA and
ISP; increased SOD and TAC | (15) |
| Randomized,
double-blind | 23 overweight
(25<BMI≤29.9 kg/m2) and obese (BMI>30
kg/m2) subjects (mean, 25.1±3.7 years) | Control
(n=10)c; AST 5 mg/day (n=12);
AST 20 mg/day (n=11); 3 weeks | Antioxidant
effect | Reduced MDA and
ISP; increased SOD and TAC | (16) |
| Randomized,
double-blind, placebo-controlled | 61 healthy subjects
with triglyceride levels 120-200 mg/dl (25-60 years) | Placebo (n=15); AST
6 mg/day (n=15); AST 12 mg/day (n=15); AST 18 mg/day (n=16); 12
weeks | Lipid
metabolism | Reduced
triglyceride levels; increased HDL; increased adiponectin | (20) |
| Randomized,
double-blind, placebo-controlled | 42 healthy young
women (20-22 years) | Placebo (n=14); AST
2 mg/day (n=14); AST 8 mg/day (n=14); 8 weeks |
Anti-inflammatoryand antioxidant
effect | Increased total
number of T and B lymphocytes; increased cytotoxic activity of
natural killer cells; reduced biomarkers of oxidative damage
8-hydroxy-2′-deoxyguanosine and C-reactive protein | (17) |
| Single-blind | 20 healthy adult
men (37-67 years) | Placebo (n=10); AST
6 mg/day (n=10); 10 days | Blood rheology | Reduced blood
transit time | (18) |
| Randomized,
double-blind, placebo-controlled | 43 participants
with type 2 diabetes (46-62 years) | Placebo (n=21); AST
8 mg/day (n=22); 8 weeks | Lipid and glucose
metabolism | Increased
adiponectin; reduced visceral fat mass, triglycerides, VLDL,
fructosamine and systolic blood pressure | (19) |
LDL oxidation is also associated with the
development of endothelial dysfunction in patients with diabetes
mellitus, which increases the risk of cardiovascular complications
(29). Endothelial dysfunction
consists of impaired vessel relaxation dependent on endothelial
factors, such as the production of nitric oxide (NO) by endothelial
NO synthase (eNOS) (30). One of
the pathways responsible for this type of dysfunction is the
binding of oxidized LDL to its endothelial receptor, lectin-like
ox-LDL receptor 1 (LOX-1), thus favoring oxidative stress, which
leads to increased lipid peroxidation and eNOS inactivation
(31). Supplementation with 10
mg/kg astaxanthin for 42 days in a diabetic rat model increased
artery relaxation by significantly lowering oxidized the levels of
LDL, LOX-1 receptor and lipid peroxidation in the aorta, in
addition to increasing eNOS, demonstrating that astaxanthin may
have therapeutic potential for the treatment of endothelial
dysfunction in diabetic patients (32).
Erythrocytes have a large amount of PUFAs and high
concentrations of oxygen and ferrous ions (Fe2+), which
makes these cells more susceptible to oxidative changes in the
lipid bilayer, leading to compromised cell stability, oxygen
transport and blood rheological properties (33). The oxidation of erythrocytes is
associated with both the formation of atheromas and the occurrence
of intraplate hemorrhage during the development of atherosclerosis
(34). As regards lipid
peroxidation in erythrocytes, daily supplementation with 6 or 12 mg
of astaxanthin for 12 weeks in healthy individuals demonstrated
that this carotenoid is incorporated and distributed into these
blood cells. When incorporated into erythrocytes, astaxanthin
exerted antioxidant effects on the cell membrane by significantly
reducing the levels of phospholipid hydroperoxides, which are the
primary products of phospholipid oxidation (14.9 pmol/ml for the
placebo group and 8.0 and 9.7 pmol/ml for the 6 and 12 mg
astaxanthin groups, respectively). In that study, the two doses of
astaxanthin used had similar effects when compared to placebo,
suggesting that the intake of 6 mg of this carotenoid is sufficient
to inhibit oxidative stress in erythrocytes (Table I) (12). A significant reduction in
oxidative damage by lipid peroxidation was also observed, as the
plasma levels of 12- and 15-hydroxy fatty acids (P=0.048 and
P=0.047, respectively) decreased after 3 months of supplementation
with 8 mg astaxanthin in healthy men (Table I) (13).
Blood rheology is important for cardiovascular
homeostasis. Astaxanthin was shown to reduce blood transit time in
a hypertensive rat model (35) as
well as in humans (18). In the
latter study, individuals receiving supplementation with 6 mg
astaxanthin for 10 days had a significantly faster blood transit
time compared with that before supplementation and with that in the
placebo group (47.6±4.2 vs. 54.2±6.7 sec for the treated and
placebo groups, respectively; P<0.05; Table I) (18). One hypothesis for the improvement
of blood rheology by astaxanthin is its antioxidant effect on the
intra- and extracellular environment and the consequent increase in
the flexibility of the erythrocyte membrane conferred by the
structural arrangement of astaxanthin in the membrane.
Other factors that affect the blood flow velocity,
such as plasma viscosity and vasodilation, affect peripheral
vascular resistance and may contribute to hypertension and its main
cardiac complication, myocardial hypertrophy (36). Studies of spontaneously
hypertensive rats (SHR) reported that astaxanthin supplementation
significantly reduced systolic pressure and induced significant
histological changes in the aorta associated with decreased
vascular stiffness and blood pressure (37-39). This response was caused by
increased endothelial cell-dependent vasodilation due to the
greater bioavailability of NO, as well as the remodeling of the
arteries. The increase in NO was caused by the reduced production
of superoxide anion radicals released by NADPH oxidase, which is
one of the antioxidant effects of astaxanthin (37,38). Chen et al (2020) reported
that this carotenoid also participated in the remodeling of the
smooth muscle cells of the vessels, reducing their proliferation
and the damage caused by oxidative stress. Astaxanthin lowered RONS
levels by increasing the activity of antioxidant enzymes and
regulating mitochondrial dynamics, mitophagy and mitochondrial
biogenesis, which are important for the maintenance of
mitochondrial and cellular metabolism (39).
The high bioavailability of NO due to lower
oxidative stress promoted by astaxanthin was also associated with
its antithrombogenic effects. In a stroke-prone SHR model,
astax-anthin significantly downregulated the oxidative stress
marker 8-hydroxy-2′-deoxyguanosine (8-OHdG) in the urine, lowered
systolic blood pressure and suppressed thrombogenesis in the
cerebral veins. The observed antithrombogenic effect may have been
due to vasodilation and inhibition of platelet aggregation caused
by an increased bioavailability of NO (40). In a murine model of thrombosis
treated with astaxanthin in the form of the CDX-085 prodrug, an
increase of ~20% in the blood flow of the carotid artery was
observed before the occurrence of endothelial dysfunction, as was a
delay in the formation of occlusive thrombi. These results were due
to the increase in NO and the decrease in ONOO−. The
authors suggested that the increase in blood flow was due to
vasodilation caused by increased release of NO by endothelial cells
and reduced platelet activation triggered by the antioxidant effect
(41).
Some of the vascular benefits promoted by the
antioxidant effect of astaxanthin were reported in an open study of
20 postmenopausal women who had a high rate of oxidative stress
(14). After 8 weeks of
supplementation with 12 mg astaxanthin, there was a significant
reduction of 4.64 and 6.93% in the systolic and diastolic pressure
values, respectively, possibly resulting from the reduction in the
vascular tone due to the action of the carotenoid in the
endothelium. Reduced vascular resistance in the lower limbs (3.7%
increase in the ankle-brachial pressure index), a 4.58% increase in
antioxidant capacity, and improvement of some physical and mental
symptoms, such as tired eye sensation and difficulty sleeping, were
also observed (14).
In addition to the protective role of astaxanthin in
the lipid oxidation process, this carotenoid affects the activity
of antioxidant enzymes involved in lipid metabolism, such as
thioredoxin reductase (TrxR) and paraoxonase-1. TrxR is an
antioxidant enzyme involved in the reduction of thioredoxin, lipid
hydroperoxides and hydrogen peroxide (42). A previous study demonstrated that
thioredoxin in its oxidized form was associated with the degree of
severity of chronic heart failure and with the resulting oxidative
stress (43). Paraoxonase-1 binds
to serum high-density lipoprotein (HDL) and is responsible for
protecting both LDL and HDL from oxidation, as well as for breaking
down oxidized lipids (44). The
effect of astaxanthin on these two enzymes was evaluated in rabbits
fed a cholesterol-rich diet (45). The authors found that astaxanthin
(100 and 500 mg/100 g of feed) reduced the amount of oxidized
protein, possibly due to changes in the activities of TrxR and
paraoxonase-1, but the in vitro evaluation only demonstrated
the direct action of this molecule on TrxR (45).
Choi et al (11,16) also conducted two randomized and
double-blind clinical studies on overweight or obese individuals to
demonstrate the antioxidant effects of astaxanthin (Table I). In one study, volunteers who
received 5 and 20 mg astaxanthin for 3 weeks exhibited lower
oxidative stress biomarkers associated with lipid peroxidation
compared with prior to treatment, with a 34.6 and 35.2% reduction
in malondialdehyde (MDA) levels, and 64.9 and 64.7% reduction in
isoprostane (ISP) levels, respectively. An increase in the activity
of the antioxidant defense system was also observed, with a 193 and
194% increase in superoxide dismutase (SOD) and a 121 and 125%
increase in total antioxidant capacity at the doses of 5 and 20 mg,
respectively, compared with the data prior to treatment (16). No significant differences were
observed between the results obtained with the two doses,
indicating that the clinical effects of this carotenoid are not
dose-dependent. Later, the same authors analyzed lipid profile,
oxidative stress and antioxidant system parameters (11). After 12 weeks of supplementation
with 20 mg astaxanthin, the same results in oxidative stress and
the antioxidant system as in the previous study were observed.
Regarding the lipid profile, there was a significant reduction of
10.4% in the LDL concentration, 7.59% in ApoB and 8.22% in the
ApoA1/ApoB ratio (considered an index of the risk of heart attack)
compared to the placebo group values (11). Therefore, these studies
demonstrated that astaxanthin reduces oxidative stress and
modulates the lipid profile in overweight and obese individuals,
mitigating the risk of developing cardiovascular diseases.
Astaxanthin also has potent detoxifying and
antioxidant effects in smokers (15). The free radicals induced by
smoking have been strongly associated with increased oxidative
stress, contributing to the increased susceptibility of smokers to
the pathogenesis of cardiovascular diseases. This group of
individuals requires a higher daily intake of antioxidants compared
with non-smokers to reduce the consequences of prolonged exposure
to toxins present in cigarettes. After 3 weeks, supplementation
with different doses of astaxanthin (5, 20 and 40 mg) in active
smokers prevented oxidative damage by suppressing lipid
peroxidation and stimulating the activity of the antioxidant system
(Table I) (15). This effect was confirmed by the
significant reduction in serum MDA and ISP levels and the increased
SOD activity and total antioxidant capacity in the three
astaxanthin groups compared with the indices prior to treatment
(15). The authors also observed
that the serum concentration of astaxanthin in the groups treated
with 20 and 40 mg was similar, showing that there was saturation of
its absorption and that smaller doses, such as 5 mg, may have the
necessary antioxidant effect for these individuals. However,
placebo-controlled studies with larger groups and longer
interventions may help determine the optimal dosage for
smokers.
Several preclinical studies have demonstrated that
astaxanthin also exerts an indirect antioxidant effect by
activating transcription factor nuclear factor erythroid 2-related
factor 2 (Nrf2), and increasing the expression of its antioxidant
target genes, such as phase II biotransformation enzymes (46-52). A study with a model of coronary
microembolization in rats revealed that supplementation with
astaxanthin drastically attenuated the induction of cardiac
dysfunction, myocardial infarction and cardiomyocyte apoptosis,
which was associated with the suppression of oxidative stress via
activation of Nrf2/heme oxygenase-1 signaling (47).
Thus, astaxanthin may accumulate in the blood plasma
and, through its antioxidant action, it helps reduce the levels of
RONS responsible for LDL oxidation and lipid peroxidation; it
increases the bioavailability of NO, enabling its vasodilator and
antithrombogenic effects; it increases the activity of antioxidant
enzymes; and it ensures the stability of blood rheological
properties, thus avoiding the loss of erythrocyte flexibility and
the increase in plasma viscosity, factors that affect the blood
flow velocity. These actions of astaxanthin against early events of
atherosclerotic plaque formation and arterial dysfunction may delay
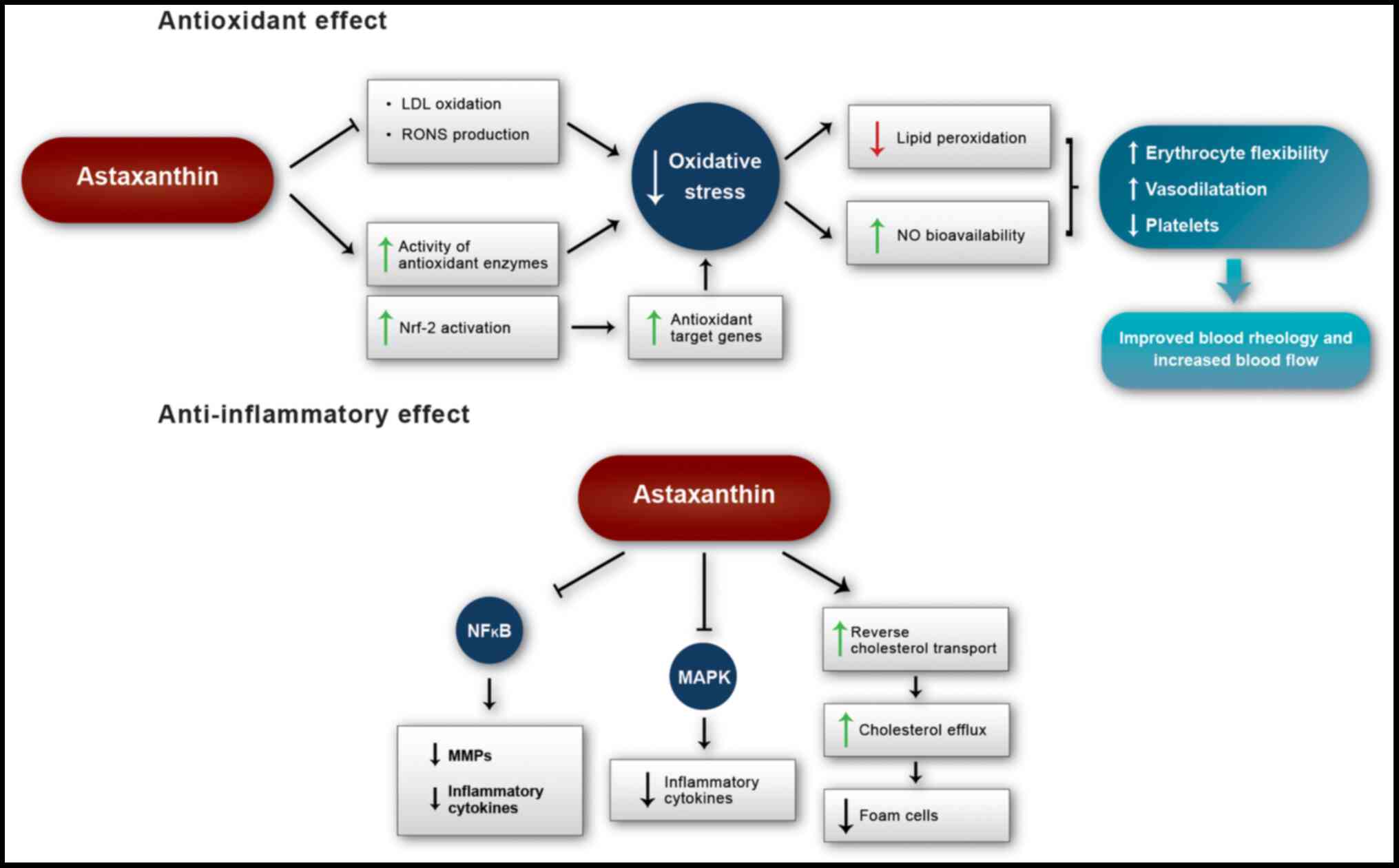
the progression of cardiovascular diseases (Fig. 1).
Anti-inflammatory effect
Inflammation plays an important role in the
development of cardiovascular diseases and other comorbidities,
such as hypertension, hypercholesterolemia, type 2 diabetes,
chronic kidney disease and obesity (53). Astaxanthin exerts a marked
anti-inflammatory effect, which may be interrelated with its
antioxidant effect and contributes to physiological changes that
benefit cardiovascular function (Fig.
1) (53-58).
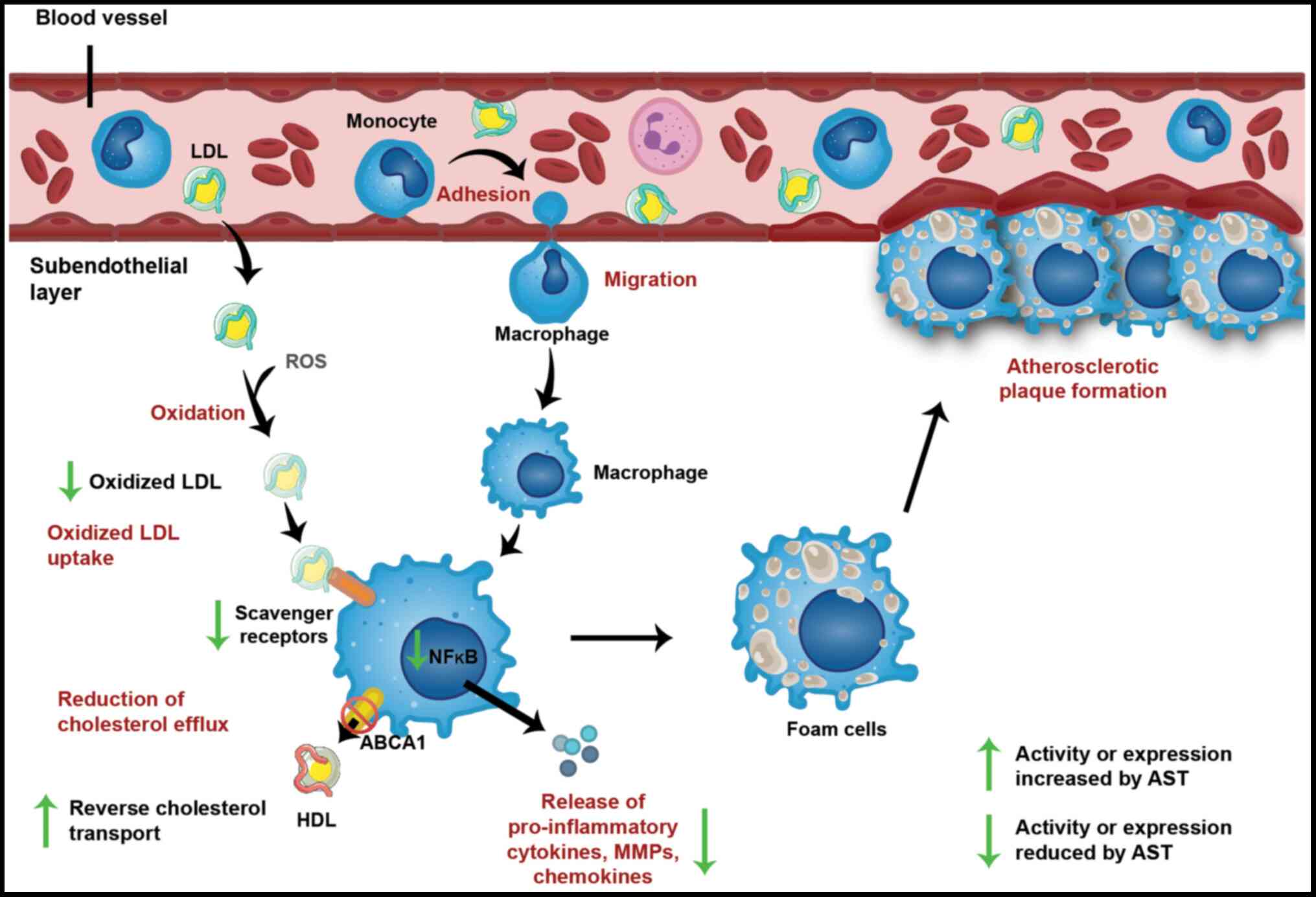
Atherosclerosis is a degenerative and chronic
disease that affects large- and medium-caliber arteries.
Atherogenesis, the initial phase of the atherosclerotic process, is
character-ized by the accumulation of LDL in the subendothelial
layer of the vascular wall, which is responsible for inflammation
mediated by the innate and adaptive immune responses (59). The anti-inflammatory effects
promoted by astaxanthin are evidenced in its role in
atherosclerosis prevention, as will be detailed below.
The epitopes generated from enzymatic or
non-enzymatic oxidation of LDL are the main damage-associated
molecular patterns recognized by macrophages and are responsible
for the onset of the inflammatory cascade, with the release of
cytokines and chemokines that recruit more resident vascular
macrophages and monocytes from the blood. Macrophages bind to
oxidized LDL via scavenger receptors, such as SR-A, SR-B2 (CD36)
and LOX-1 (59). The expression
of these receptors is controlled by nuclear factor-κB (NF-κB), the
main mediator of the inflammatory response, which is activated by
pattern recognition receptors and pro-inflammatory cytokines
(60). In inflammatory states,
macrophages produce excessive amounts of pro-inflammatory
mediators, such as cytokines, chemokines, NO, cyclooxygenase-2
(COX-2) and matrix metalloproteinases (MMPs). MMPs are responsible
for the degradation of most extracellular matrix proteins and
mediate the tissue remodeling associated with atherosclerosis
(5).
In vitro and in vivo studies have
evaluated the effect of astaxanthin on the formation of
atherosclerotic plaques (54-58,61-82). Supplementation with 10 µM
astaxanthin significantly reduced the expression of the SR-A and
CD36 scavenger receptors in the THP-1 macrophage line and reduced
the total activity of MMPs, as reflected by reduced protein
expression of MMP-9 and MMP-2 and of the mRNA levels of five MMPs
(62). Astaxanthin at this
concentration also reduced the gene expression of pro-inflammatory
markers, such as interleukin (IL)-1β, IL-6, tumor necrosis factor-α
(TNF-α), inducible nitric oxide synthase (iNOS) and COX-2 (62). These results corroborate those of
other studies indicating that astaxanthin reduces the expression of
pro-inflammatory mediators in macrophages (62-65) and other cell types, such as
microglia, endothelial vascular cells and human neutrophils
(66-69).
The significant decreases in the levels of MMPs and
proinflammatory cytokines may result from the suppression of the
NF-κB transcription factor by astaxanthin (54-58,62,64,68,70,71). NF-κB is frequently activated at
inflammation sites associated with various pathologies,
particularly cardiovascular diseases, in the etiology of which the
increased expression of its pro-inflammatory target genes plays a
key role (72). The inflammatory
pathway of NF-κB is, at least in part, regulated by oxidative
stress (72). Astaxanthin
inhibits the activity of IκB kinase, a complex responsible for the
control of NF-κB activation. This maintains NF-κB inactive in the
cell cytoplasm, and its pro-inflammatory target genes, such as
TNF-α, IL-1β and iNOS, are downregulated (64).
Cholesterol uptake is balanced by the transfer of
this molecule from macrophages to free apolipoproteins A1 or to
HDL, the latter being responsible for the reverse cholesterol
transport process. When cholesterol uptake exceeds cholesterol
efflux in macrophages, lipid droplets accumulate in the cytoplasm,
forming foam cells, the main markers of atherosclerosis (Fig. 2) (59). The progression of cholesterol
accumulation may lead to its precipitation in the form of crystals,
which activate the inflammasome, leading to cell death by apoptosis
or necrosis (83). The
atherosclerotic plaque is separated from the bloodstream by a
fibrous layer, which, upon rupture, initiates intraluminal
thrombosis, the initial event of stroke and other coronary
syndromes (59).
Reverse cholesterol transport consists in HDL
removing excess cholesterol from peripheral tissues and
transporting it to the liver, where it is degraded by bile juice
and excreted in the feces, thus preventing the accumulation of
cholesterol in macrophages (79).
Both the liver and intestine synthesize apolipoprotein A-I (apoA-I)
and apoA-II in the plasma, which incorporate free cholesterol and
phospholipids through the ATP-binding cassette A1 (ABCA1) hepatic
transporter, originating from nascent HDL. In peripheral tissues,
nascent HDL molecules recruit free cholesterol from foam cells via
the macrophage ABCA1 transporter (80). Of note, this reverse cholesterol
transport was also observed in the lymphatic system, which is
largely responsible for the removal of cholesterol from different
tissues (79). Finally, mature
HDL can transport cholesterol directly to the liver via the SR-B1
scavenger receptor or can transfer cholesteryl esters to very
low-density lipoprotein (VLDL) through the cholesteryl ester
transfer protein (81). These
lipoproteins are absorbed in the liver by their specific receptors,
which is likely the predominant pathway in humans. Once in the
liver, cholesterol is secreted into the bile via the ABCG5 and
ABCG8 transporters. Some of these molecules can be reabsorbed by
the intestine and reach the bloodstream again, while the rest is
excreted in the feces (80).
In atherosclerosis, apolipoproteins are oxidized by
the enzyme myeloperoxidase, which is expressed in macrophages
during the inflammatory process, compromising cholesterol efflux
via ABCA1 (81). In individuals
with heart disease, elevated levels of apoA-I modified by
myeloperoxidase were identified, and their HDL molecules were
dysfunctional in performing reverse cholesterol transport. In
addition, an association was observed between increased cholesterol
efflux via ABCA1 and reduced risk of cardiovascular diseases
(OR=0.30; 95% CI: 0.14-0.66; P<0.003) (82). Thus, the oxidation of
apolipoproteins by macrophage myeloperoxidase is a determining
factor in HDL dysfunction in cholesterol transport and, therefore,
in the risk of cardiovascular diseases.
The effects of astaxanthin on reverse cholesterol
transport have been demonstrated in vivo. In wild-type and
ApoE−/− mice, astaxanthin increased cholesterol efflux
from peripheral tissues to the liver and its subsequent excretion
in feces (61). In addition, in
the ApoE−/− model, this carotenoid promoted a
significant decline in plasma total cholesterol, triglycerides and
non-HDL cholesterol, and reduced the atherosclerotic plaque area of
the aortic sinus and the cholesterol concentration in the aorta
compared to controls. Thus, astaxanthin may exert
antiatherosclerotic effects by increasing the activity of the
reverse cholesterol transport pathway, but the molecular mechanisms
underlying this action remain elusive (61).
In addition to its function in reverse cholesterol
transport, astaxanthin is involved in certain lipid metabolism
steps, a finding corroborated by a randomized, placebo-controlled
clinical study of 61 adult individuals with moderate hyperlipidemia
(Table I) (20). In that study, daily
supplementation with 6, 12 or 18 mg astaxanthin for 12 weeks led to
an improvement in the lipid profile of the patients. Triglycerides
were reduced by 25.2 and 23.8% (P<0.05) by doses of 12 and 18
mg, respectively, whereas HDL was increased by 10.6% (P<0.05)
and 15.4% (P<0.01) by doses of 6 and 12 mg, respectively. Doses
of 12 and 18 mg also significantly increased serum adiponectin
(P<0.01 and P<0.05, respectively), a protein secreted by
adipocytes with important functions in the cardiovascular and
endocrine systems associated with its anti-inflammatory,
atheroprotective and insulin-sensitizing actions (20).
Inflammation is also involved in the pathophysiology
of metabolic syndrome, a multifactorial disorder associated with
glucose and lipid metabolism disorders. This disease has risk
factors that are also strongly associated with the development of
cardiovascular complications, including type 2 diabetes,
dyslipidemia, hypertension and abdominal fat deposition (84). In this context, astaxanthin has
been found to be promising in the improvement of glucose and lipid
metabolism in a randomized, placebo-controlled clinical study with
43 diabetic patients aged 46-62 years (Table I) (19). In agreement with a previous
clinical study (20),
supplementation with astaxanthin (8 mg/day for 8 weeks)
significantly increased serum adiponectin (47±14 vs. 45±13 and
36±15 µg/ml compared with placebo and base-line,
respectively; P<0.05) and improved the lipid profile of the
patients, as shown by the reductions in the levels of triglycerides
(128±52 vs. 150±85 and 156±90 mg/dl compared with placebo and
baseline, respectively; P<0.05) and VLDL (27±16 vs. 31±16 mg/dl
compared with placebo; P<0.05). Furthermore, astaxanthin
marginally reduced fasting glucose levels (8.3±2.7 vs. 9.4±3.2
mmol/l compared with placebo; P=0.057) and significantly increased
serum fructosamine levels (5.8±3.8 vs. 7.32±4.31 and 7.36±4.2
µmol/l compared with placebo and baseline, respectively;
P<0.05), an important marker in the control of diabetes that
reflects the mean concentration of blood glucose. Patients
receiving astaxanthin supplementation also exhibited lower visceral
fat deposition (11.2±3.4 vs. 11.85±3.8% compared with placebo;
P<0.05) and systolic blood pressure (132±18 vs. 133±19 and
143±27 mmHg compared with placebo and baseline, respectively;
P<0.05) (19).
Disorders characterized by ischemia/reperfusion,
including myocardial infarction, stroke and peripheral vascular
disease, are among the most frequent causes of morbidity and
mortality worldwide (85).
Ischemia/reperfusion is a complex inflammatory process associated
with high levels of oxidative stress in the affected tissue
(86). In rodents with hepatic
lesions induced by ischemia/reperfusion, astaxanthin not only
reduced oxidative stress and histopathological damage (87,88) but also exerted a significant
anti-inflammatory effect, attenuating the release of inflammatory
cytokines through the mitogen-activated protein kinase (MAPK)
pathway (88,89). Furthermore, astaxanthin exerted an
anti-inflammatory and antioxidant effect in the context of
myocardial injury due to ischemia/reperfusion in rabbits by
significantly reducing the activation of the complement system
associated with the reduced deposition of C-reactive protein and
the membrane attack complex in the injured area of the myocardium
(90).
In mice with non-alcoholic steatohepatitis (NASH)
induced by a high-lipid diet, supplementation with astaxanthin
(0.02% in the diet, ~20 mg/kg body weight) significantly improved
several liver parameters: it reduced liver inflammation, decreased
the proportion of pro-inflammatory M1-type macrophages, reduced
stellate cell activation, and attenuated liver fibrosis, the
accumulation and peroxidation of hepatic lipids and insulin
resistance (91). Additionally,
astaxanthin was more effective in preventing and treating NASH and
improving liver inflammation and fibrosis compared with vitamin E
(standard NASH treatment). The same study also corroborated the
potential of astaxanthin to improve NASH in 12 individuals
receiving this carotenoid as a supplement (12 mg/day; control:
placebo) for 24 weeks (91).
The effects of astaxanthin on the relief of liver
injury was shown to be correlated to its positive effects on the
intestinal microbiota and consequent reduction of inflammation
(92). In fact, a growing body of
evidence indicates that gut microbiota plays a key role in the
pathogenesis of inflammatory disorders and cardiovascular diseases,
and alterations in its composition (dysbiosis) have been associated
with heart failure, hypertension, atherosclerosis and metabolic
syndrome (93-95). Several recent in vivo
studies revealed that astaxanthin supplementation improved gut
microbiota composition, which may contribute to its local and
systemic anti-inflammatory and antioxidant effects (92,96-99). The beneficial effect of
astaxanthin on gut microbiota has been shown to be correlated with
the mitigation of cardiovascular disease-related pathologies and
risk factors, such as obesity (100), insulin resistance (99) and alcoholic fatty liver disease
(92).
The anti-inflammatory and antioxidant effects of
astaxanthin were also confirmed by a randomized clinical trial in
42 healthy young women receiving placebo or astaxanthin at 2 or 8
mg/day (Table I) (17). Compared with placebo, after 8
weeks of treatment, 2 mg astaxanthin significantly lowered the
levels of the plasma inflammatory marker C-reactive protein
(unspecified values; P<0.05). Astaxanthin improved the immune
responses of the participants, as evidenced by the increased
cytotoxic activity of natural killer cells (8 mg dose, 67.9±3.0 vs.
57.8±2.7% lysis; P<0.05), levels of T and B lymphocytes (2 mg
dose, 75.7±1.6 vs. 70.6±1.5 and 13.1±0.5 vs. 10.7±0.5%,
respectively; P<0.05), and the production of interferon (IFN)-γ
and IL-6 (dose of 8 mg, 9.55 vs. 4.68 and 25.2 vs. 13.6 pg/ml,
respectively; P<0.05). Finally, starting at 4 weeks, both doses
drastically reduced the plasma levels of 8-OHdG (unspecified
values; P<0.01), a biomarker of oxidative DNA damage (17).
Although macrophages are the main type of immune
cell found in atherosclerotic plaques, T lymphocytes also
contribute to the development of the disease (101). In fact, the inflammatory
response mediated by T lymphocytes plays a crucial role in the
etiology of cardiovascular diseases, contributing to
atherosclerosis, heart failure and myocardial infarction (102-108). For example, T helper cells can
be activated by LDL particles in the arterial wall and trigger
inflammation through an autoimmune response, contributing to the
development of atherosclerotic plaques (106,108,109). Similarly, self-reactive T helper
cells may target cardiomyocytes, contributing to the development of
heart failure (103).
Astaxanthin was shown to be effective not only in
preventing oxidative stress in T lymphocytes (110-115), but also in modulating their
activity (115-123). In the aforementioned clinical
study on healthy young women (17), astaxanthin supplementation
stimulated mitogen-induced lymphoproliferation and increased the
subpopulation of T lymphocytes, without changing the populations of
T killer or T helper cells, as well as increased the response to
tuberculin, an indicator of T lymphocyte function. In a mouse model
of NASH, astaxanthin reduced T helper and T killer cell recruitment
to the liver, contributing to the improvement of inflammation and
insulin resistance (91).
Supplementation for 45 days with fish oil containing astaxanthin (1
mg/kg of body weight) significantly reduced the proliferative
capacity of T lymphocytes in response to mitogens and RONS
production when compared with fish oil alone (115). In in vitro studies with
peripheral blood mononuclear cells from patients with asthma and
allergic rhinitis, it was demonstrated that astaxanthin
significantly suppressed the activation of T lymphocytes induced by
phytohemagglutinin (116,122).
Another in vitro and ex vivo study with cultured
lymphocytes demonstrated that astaxanthin stimulated their immune
response and increased the production of IL-2 and IFN-γ, without
inducing cytotoxicity (117).
The administration of astaxanthin in mice prevented renal fibrosis
by mechanisms involving stimulation of T killer cell recruitment
and increased production of IFN-γ (118). In cats, astaxanthin increased
the immune response mediated by total T lymphocytes and T helper
cells (119).
Therefore, astaxanthin was shown to exert a clear
modulatory effect on T lymphocytes, overall improving their immune
response or downregulating their potentially pathological immune
activation. However, the role of T lymphocyte modulation by
astaxanthin in the risk and progression of cardiovascular diseases
remain to be fully elucidated.
In summary, inflammation plays a key role in the
pathophysiology of cardiovascular diseases and their risk factors,
while astaxanthin exerts beneficial anti-inflammatory effects. The
mechanism of action of this carotenoid involves inhibition of the
NF-κB and MAPK signaling pathways, which suppresses the
inflammatory process and stimulates reverse cholesterol transport,
thereby attenuating the formation of foam cells (Fig. 1).
3. Clinical evidence of the prevention of
cardiovascular diseases by astaxanthin
The potential beneficial effects of oral astaxanthin
supplementation on cardiovascular physiology were evidenced in 11
clinical studies, summarized in Table
I. Of these 11 studies, 6 were randomized placebo-controlled
studies (11-13,17,19,20), 1 was single-blinded (18), 2 were open-label (10,14), and 2 were randomized but lacked a
placebo group (15,16). A total of 6 studies evaluated the
metabolic and oxidative changes promoted by astaxanthin in healthy
individuals, while 5 investigated individuals who had one element
of the metabolic syndrome, namely obesity, dyslipidemia or type 2
diabetes. In addition, the dose of astaxanthin ranged between 1.8
and 40 mg, and the duration of supplementation varied between 10
days and 12 weeks, reflecting a wide variety of these two
parameters. Despite encompassing a small population with a total of
417 individuals, the results of those studies indicated that the
beneficial effect of astaxanthin on cardiovascular health was
mainly due to its antioxidant and anti-inflammatory properties, its
ability to modulate lipid and glucose metabolism, and its role in
the maintenance of blood rheological properties.
4. Conclusion
Based on preclinical and clinical evidence, the
antioxidant and anti-inflammatory effects of astaxanthin appear to
delay the progression of cardiovascular diseases. As an
antioxidant, astaxanthin reduces oxidative stress, increases the
bioavailability of NO and the activity of antioxidant enzymes, and
maintains the rheological properties of the blood. Its
anti-inflammatory properties involve modulating the NF-κB and MAPK
signaling pathways, reducing the release of pro-inflammatory
cytokines and increasing reverse cholesterol transport by HDL,
thereby attenuating the accumulation of cholesterol in foam cells
and the formation of atherosclerotic plaques. These properties of
astaxanthin, together with its favorable safety profile, make this
compound a promising option for the prevention and/or adjuvant
treatment of cardiovascular diseases.
Funding
No funding was received.
Availability of data and materials
Not applicable.
Authors' contributions
CPMP and JJN contributed to the conception of the
study and critically reviewed the article. CPMP contributed to the
design of the manuscript and figure preparation and editing. CPMP,
ACRS, ARV, PSP and JJN contributed to the data acquisition and
analysis and drafted the manuscript. All authors have approved all
aspects of the present study and agree to be fully accountable for
ensuring the integrity and accuracy of the work. All the authors
have read and approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
All the authors declare that they have no competing
interests.
Acknowledgments
The authors would like to thank Paula Mitie Hirata
for the technical assistance with figure editing.
References
|
1
|
World Health Organization Cardiovascular
diseases (CVDs): Journal. 2017, https://www.who.int/en/news-room/fact-sheets/detail/cardiovascular-diseases-(cvds)urisimplehttps://www.who.int/en/news-room/fact-sheets/detail/cardiovascular-diseases-(cvds).
Accessed June 10, 2020.
|
|
2
|
Spahis S, Borys JM and Levy E: Metabolic
syndrome as a multi-faceted risk factor for oxidative stress.
Antioxid Redox Signal. 26:445–461. 2017. View Article : Google Scholar
|
|
3
|
Vona R, Gambardella L, Cittadini C,
Straface E and Pietraforte D: Biomarkers of oxidative stress in
metabolic syndrome and associated diseases. Oxid Med Cell Longev.
2019:82672342019. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Puddu P, Puddu GM, Galletti L, Cravero E
and Muscari A: Mitochondrial dysfunction as an initiating event in
atherogenesis: A plausible hypothesis. Cardiology. 103:137–141.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Kishimoto Y, Yoshida H and Kondo K:
Potential anti-atherosclerotic properties of astaxanthin. Mar
Drugs. 14:352016. View Article : Google Scholar :
|
|
6
|
Guerin M, Huntley ME and Olaizola M:
Haematococcus astaxanthin: Applications for human health and
nutrition. Trends Biotechnol. 21:210–216. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Hussein G, Sankawa U, Goto H, Matsumoto K
and Watanabe H: Astaxanthin, a carotenoid with potential in human
health and nutrition. J Nat Prod. 69:443–449. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Zhang L and Wang H: Multiple mechanisms of
anti-cancer effects exerted by astaxanthin. Mar Drugs.
13:4310–4330. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Maoka T and Etoh H: Some biological
functions of carotenoids in Japanese food. Functional Foods of the
East. Shi J, Ho CT and Shahidi F: CRC Press; Boca Raton, FL: pp.
85–97. 2010, View
Article : Google Scholar
|
|
10
|
Iwamoto T, Hosoda K, Hirano R, Kurata H,
Matsumoto A, Miki W, Kamiyama M, Itakura H, Yamamoto S and Kondo K:
Inhibition of low-density lipoprotein oxidation by astaxanthin. J
Atheroscler Thromb. 7:216–222. 2000. View Article : Google Scholar
|
|
11
|
Choi HD, Youn YK and Shin WG: Positive
effects of astaxanthin on lipid profiles and oxidative stress in
overweight subjects. Plant Foods Hum Nutr. 66:363–369. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Nakagawa K, Kiko T and Miyazawa T,
Carpentero Burdeos G, Kimura F, Satoh A and Miyazawa T: Antioxidant
effect of astaxanthin on phospholipid peroxidation in human
erythrocytes. Br J Nutr. 105:1563–1571. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Karppi J, Rissanen TH, Nyyssönen K,
Kaikkonen J, Olsson AG, Voutilainen S and Salonen JT: Effects of
astaxanthin supplementation on lipid peroxidation. Int J Vitam Nutr
Res. 77:3–11. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Iwabayashi M, Fujioka N, Nomoto K,
Miyazaki R, Takahashi H, Hibino S, Takahashi Y, Nishikawa K,
Nishida M and Yonei Y: Efficacy and safety of eight-week treatment
with astaxanthin in individuals screened for increased oxidative
stress burden. Anti Aging Med. 6:15–21. 2009. View Article : Google Scholar
|
|
15
|
Kim JH, Chang MJ, Choi HD, Youn YK, Kim
JT, Oh JM and Shin WG: Protective effects of Haematococcus
astaxanthin on oxidative stress in healthy smokers. J Med Food.
14:1469–1475. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Choi HD, Kim JH, Chang MJ, Kyu-Youn Y and
Shin WG: Effects of astaxanthin on oxidative stress in overweight
and obese adults. Phytother Res. 25:1813–1818. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Park JS, Chyun JH, Kim YK, Line LL and
Chew BP: Astaxanthin decreased oxidative stress and inflammation
and enhanced immune response in humans. Nutr Metab (Lond).
7:182010. View Article : Google Scholar
|
|
18
|
Miyawaki H, Takahashi J, Tsukahara H and
Takehara I: Effects of astaxanthin on human blood rheology. J Clin
Biochem Nutr. 43:69–74. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Mashhadi NS, Zakerkish M, Mohammadiasl J,
Zarei M, Mohammadshahi M and Haghighizadeh MH: Astaxanthin improves
glucose metabolism and reduces blood pressure in patients with type
2 diabetes mellitus. Asia Pac J Clin Nutr. 27:341–346.
2018.PubMed/NCBI
|
|
20
|
Yoshida H, Yanai H, Ito K, Tomono Y,
Koikeda T, Tsukahara H and Tada N: Administration of natural
astaxanthin increases serum HDL-cholesterol and adiponectin in
subjects with mild hyperlipidemia. Atherosclerosis. 209:520–523.
2010. View Article : Google Scholar
|
|
21
|
Lorenz RT and Cysewski GR: Commercial
potential for Haematococcus microalgae as a natural source of
astaxanthin. Trends Biotechnol. 18:160–167. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Hulbert AJ, Pamplona R, Buffenstein R and
Buttemer WA: Life and death: Metabolic rate, membrane composition,
and life span of animals. Physiol Rev. 87:1175–1213. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Barros MP, Pinto E, Colepicolo P and
Pedersén M: Astaxanthin and peridinin inhibit oxidative damage in
Fe(2+)-loaded liposomes: Scavenging oxyradicals or changing
membrane permeability? Biochem Biophys Res Commun. 288:225–232.
2001. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
McNulty HP, Byun J, Lockwood SF, Jacob RF
and Mason RP: Differential effects of carotenoids on lipid
peroxidation due to membrane interactions: X-ray diffraction
analysis. Biochim Biophys Acta. 1768:167–174. 2007. View Article : Google Scholar
|
|
25
|
Kidd P: Astaxanthin, cell membrane
nutrient with diverse clinical benefits and anti-aging potential.
Altern Med Rev. 16:355–364. 2011.
|
|
26
|
Liaudet L, Rosenblatt-Velin N and Pacher
P: Role of peroxynitrite in the cardiovascular dysfunction of
septic shock. Curr Vasc Pharmacol. 11:196–207. 2013.PubMed/NCBI
|
|
27
|
Halliwell B: Free radicals and
antioxidants: A personal view. Nutr Rev. 52:253–265. 1994.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Maoka T, Tokuda H, Suzuki N, Kato H and
Etoh H: Anti-oxidative, anti-tumor-promoting, and
anti-carcinogensis activities of nitroastaxanthin and nitrolutein,
the reaction prod-ucts of astaxanthin and lutein with
peroxynitrite. Mar Drugs. 10:1391–1399. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Donahoe SM, Stewart GC, McCabe CH,
Mohanavelu S, Murphy SA, Cannon CP and Antman EM: Diabetes and
mortality following acute coronary syndromes. JAMA. 298:765–775.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Cai H and Harrison DG: Endothelial
dysfunction in cardiovas-cular diseases: The role of oxidant
stress. Circ Res. 87:840–844. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Xu X, Gao X, Potter BJ, Cao JM and Zhang
C: Anti-LOX-1 rescues endothelial function in coronary arterioles
in atheroscle-rotic ApoE knockout mice. Arterioscler Thromb Vasc
Biol. 27:871–877. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Zhao ZW, Cai W, Lin YL, Lin QF, Jiang Q,
Lin Z and Chen LL: Ameliorative effect of astaxanthin on
endothelial dysfunction in streptozotocin-induced diabetes in male
rats. Arzneimittelforschung. 61:239–246. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
da Silva Garrote-Filho M, Bernardino-Neto
M and Penha-Silva N: Influence of erythrocyte membrane stability in
atherosclerosis. Curr Atheroscler Rep. 19:172017. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Pasterkamp G and Virmani R: The
erythrocyte: A new player in atheromatous core formation. Heart.
88:115–116. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Hussein G, Goto H, Oda S, Iguchi T,
Sankawa U, Matsumoto K and Watanabe H: Antihypertensive potential
and mechanism of action of astaxanthin: II. Vascular reactivity and
hemorheology in spontaneously hypertensive rats. Biol Pharm Bull.
28:967–971. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Becker RC: The role of blood viscosity in
the development and progression of coronary artery disease. Cleve
Clin J Med. 60:353–358. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Hussein G, Goto H, Oda S, Sankawa U,
Matsumoto K and Watanabe H: Antihypertensive potential and
mechanism of action of astaxanthin: III. Antioxidant and
histopathological effects in spontaneously hypertensive rats. Biol
Pharm Bull. 29:684–688. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Monroy-Ruiz J, Sevilla MÁ, Carrón R and
Montero MJ: Astaxanthin-enriched-diet reduces blood pressure and
improves cardiovascular parameters in spontaneously hypertensive
rats. Pharmacol Res. 63:44–50. 2011. View Article : Google Scholar
|
|
39
|
Chen Y, Li S, Guo Y, Yu H, Bao Y, Xin X,
Yang H, Ni X, Wu N and Jia D: Astaxanthin attenuates hypertensive
vascular remodeling by protecting vascular smooth muscle cells from
oxidative stress-induced mitochondrial dysfunction. Oxid Med Cell
Longev. 2020:46291892020. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Sasaki Y, Kobara N, Higashino S, Giddings
JC and Yamamoto J: Astaxanthin inhibits thrombosis in cerebral
vessels of stroke-prone spontaneously hypertensive rats. Nutr Res.
31:784–789. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Khan SK, Malinski T, Mason RP, Kubant R,
Jacob RF, Fujioka K, Denstaedt SJ, King TJ, Jackson HL, Hieber AD,
et al: Novel astaxanthin prodrug (CDX-085) attenuates thrombosis in
a mouse model. Thromb Res. 126:299–305. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Nordberg J and Arnér ES: Reactive oxygen
species, antioxidants, and the mammalian thioredoxin system. Free
Radic Biol Med. 31:1287–1312. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Jekell A, Hossain A, Alehagen U, Dahlström
U and Rosén A: Elevated circulating levels of thioredoxin and
stress in chronic heart failure. Eur J Heart Fail. 6:883–890. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Aviram M: Introduction to the serial
review on paraoxonases, oxidative stress, and cardiovascular
diseases. Free Radic Biol Med. 37:1301–1303. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Augusti PR, Quatrin A, Somacal S,
Conterato GM, Sobieski R, Ruviaro AR, Maurer LH, Duarte MM, Roehrs
M and Emanuelli T: Astaxanthin prevents changes in the activities
of thioredoxin reductase and paraoxonase in hypercholesterolemic
rabbits. J Clin Biochem Nutr. 51:42–49. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Cui G, Li L, Xu W, Wang M, Jiao D, Yao B,
Xu K, Chen Y, Yang S, Long M, et al: Astaxanthin protects
ochratoxin a-induced oxidative stress and apoptosis in the heart
via the Nrf2 pathway. Oxid Med Cell Longev. 2020:76391092020.
View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Xue Y, Sun C, Hao Q and Cheng J:
Astaxanthin ameliorates cardiomyocyte apoptosis after coronary
microembolization by inhibiting oxidative stress via Nrf2/HO-1
pathway in rats. Naunyn Schmiedebergs Arch Pharmacol. 392:341–348.
2019. View Article : Google Scholar
|
|
48
|
Wu Q, Zhang XS, Wang HD, Zhang X, Yu Q, Li
W, Zhou ML and Wang XL: Astaxanthin activates nuclear factor
erythroid-related factor 2 and the antioxidant responsive element
(Nrf2-ARE) pathway in the brain after subarachnoid hemorrhage in
rats and attenuates early brain injury. Mar Drugs. 12:6125–6141.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Kavitha K, Thiyagarajan P, Rathna Nandhini
J, Mishra R and Nagini S: Chemopreventive effects of diverse
dietary phyto-chemicals against DMBA-induced hamster buccal pouch
carcinogenesis via the induction of Nrf2-mediated cytoprotective
antioxidant, detoxification, and DNA repair enzymes. Biochimie.
95:1629–1639. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Tripathi DN and Jena GB: Astaxanthin
intervention ameliorates cyclophosphamide-induced oxidative stress,
DNA damage and early hepatocarcinogenesis in rat: Role of Nrf2,
p53, p38 and phase-II enzymes. Mutat Res. 696:69–80. 2010.
View Article : Google Scholar
|
|
51
|
Saw CL, Yang AY, Guo Y and Kong AN:
Astaxanthin and omega-3 fatty acids individually and in combination
protect against oxidative stress via the Nrf2-ARE pathway. Food
Chem Toxicol. 62:869–875. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Wen X, Huang A, Hu J, Zhong Z, Liu Y, Li
Z, Pan X and Liu Z: Neuroprotective effect of astaxanthin against
glutamate-induced cytotoxicity in HT22 cells: Involvement of the
Akt/GSK-3β pathway. Neuroscience. 303:558–568. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Visioli F and Artaria C: Astaxanthin in
cardiovascular health and disease: Mechanisms of action,
therapeutic merits, and knowledge gaps. Food Funct. 8:39–63. 2017.
View Article : Google Scholar
|
|
54
|
Li J, Dai W, Xia Y, Chen K, Li S, Liu T,
Zhang R, Wang J, Lu W, Zhou Y, et al: Astaxanthin inhibits
proliferation and induces apoptosis of human hepatocellular
carcinoma cells via inhibition of NF-κB P65 and Wnt/B-catenin in
vitro. Mar Drugs. 13:6064–6081. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Kochi T, Shimizu M, Sumi T, Kubota M,
Shirakami Y, Tanaka T and Moriwaki H: Inhibitory effects of
astaxanthin on azoxymethane-induced colonic preneoplastic lesions
in C57/BL/KsJ-db/db mice. BMC Gastroenterol. 14:2122014. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Kavitha K, Kowshik J, Kishore TK, Baba AB
and Nagini S: Astaxanthin inhibits NF-κB and Wnt/β-catenin
signaling path-ways via inactivation of Erk/MAPK and PI3K/Akt to
induce intrinsic apoptosis in a hamster model of oral cancer.
Biochim Biophys Acta. 1830:4433–4444. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Yasui Y, Hosokawa M, Mikami N, Miyashita K
and Tanaka T: Dietary astaxanthin inhibits colitis and
colitis-associated colon carcinogenesis in mice via modulation of
the inflammatory cyto-kines. Chem Biol Interact. 193:79–87. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Nagendraprabhu P and Sudhandiran G:
Astaxanthin inhibits tumor invasion by decreasing extracellular
matrix production and induces apoptosis in experimental rat colon
carcinogenesis by modulating the expressions of ERK-2, NFkB and
COX-2. Invest New Drugs. 29:207–224. 2011. View Article : Google Scholar
|
|
59
|
Moroni F, Ammirati E, Norata GD, Magnoni M
and Camici PG: The role of monocytes and macrophages in human
atherosclerosis, plaque neoangiogenesis, and atherothrombosis.
Mediators Inflamm. 2019:74343762019. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Hashizume M and Mihara M: Blockade of IL-6
and TNF-α inhibited oxLDL-induced production of MCP-1 via scavenger
receptor induction. Eur J Pharmacol. 689:249–254. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Zou TB, Zhu SS, Luo F, Li WQ, Sun XR and
Wu HF: Effects of astaxanthin on reverse cholesterol transport and
atherosclerosis in mice. Biomed Res Int. 2017:46259322017.
View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Kishimoto Y, Tani M, Uto-Kondo H, Iizuka
M, Saita E, Sone H, Kurata H and Kondo K: Astaxanthin suppresses
scavenger receptor expression and matrix metalloproteinase activity
in macrophages. Eur J Nutr. 49:119–126. 2010. View Article : Google Scholar
|
|
63
|
Santos SD, Cahú TB, Firmino GO, de Castro
CC, Carvalho LB Jr, Bezerra RS and Filho JL: Shrimp waste extract
and astaxanthin: Rat alveolar macrophage, oxidative stress and
inflammation. J Food Sci. 77:H141–H146. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Lee SJ, Bai SK, Lee KS, Namkoong S, Na HJ,
Ha KS, Han JA, Yim SV, Chang K, Kwon YG, et al: Astaxanthin
inhibits nitric oxide production and inflammatory gene expression
by suppressing I(kappa)B kinase-dependent NF-kappaB activation. Mol
Cells. 16:97–105. 2003.PubMed/NCBI
|
|
65
|
Franceschelli S, Pesce M, Ferrone A, De
Lutiis MA, Patruno A, Grilli A, Felaco M and Speranza L:
Astaxanthin treatment confers protection against oxidative stress
in U937 cells stimulated with lipopolysaccharide reducing
O2-production. PLoS One. 9:e883592014. View Article : Google Scholar
|
|
66
|
Macedo RC, Bolin AP, Marin DP and Otton R:
Astaxanthin addition improves human neutrophils function: In vitro
study. Eur J Nutr. 49:447–457. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Choi SK, Park YS, Choi DK and Chang HI:
Effects of astaxanthin on the production of NO and the expression
of COX-2 and iNOS in LPS-stimulated BV2 microglial cells. J
Microbiol Biotechnol. 18:1990–1996. 2008.
|
|
68
|
Kim YJ, Kim YA and Yokozawa T: Protection
against oxidative stress, inflammation, and apoptosis of
high-glucose-exposed proximal tubular epithelial cells by
astaxanthin. J Agric Food Chem. 57:8793–8797. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Abdelzaher LA, Imaizumi T, Suzuki T,
Tomita K, Takashina M and Hattori Y: Astaxanthin alleviates
oxidative stress insults-related derangements in human vascular
endothelial cells exposed to glucose fluctuations. Life Sci.
150:24–31. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Speranza L, Pesce M, Patruno A,
Franceschelli S, de Lutiis MA, Grilli A and Felaco M: Astaxanthin
treatment reduced oxidative induced pro-inflammatory cytokines
secretion in U937: SHP-1 as a novel biological target. Mar Drugs.
10:890–899. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Jones WK, Brown M, Wilhide M, He S and Ren
X: NF-kappaB in cardiovascular disease: Diverse and specific
effects of a 'general' transcription factor? Cardiovasc Toxicol.
5:183–202. 2005. View Article : Google Scholar
|
|
72
|
Pashkow FJ, Watumull DG and Campbell CL:
Astaxanthin: A novel potential treatment for oxidative stress and
inflammation in cardiovascular disease. Am J Cardiol. 101:58D–68D.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Ghosh S, May MJ and Kopp EB: NF-kappa B
and Rel proteins: Evolutionarily conserved mediators of immune
responses. Annu Rev Immunol. 16:225–260. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Woronicz JD, Gao X, Cao Z, Rothe M and
Goeddel DV: IkappaB kinase-beta: NF-kappaB activation and complex
formation with IkappaB kinase-alpha and NIK. Science. 278:866–869.
1997. View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Mercurio F, Zhu H, Murray BW, Shevchenko
A, Bennett BL, Li J, Young DB, Barbosa M, Mann M, Manning A and Rao
A: IKK-1 and IKK-2: Cytokine-activated IkappaB kinases essential
for NF-kappaB activation. Science. 278:860–866. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
76
|
Zandi E, Rothwarf DM, Delhase M, Hayakawa
M and Karin M: The IkappaB kinase complex (IKK) contains two kinase
subunits, IKKalpha and IKKbeta, necessary for IkappaB
phosphorylation and NF-kappaB activation. Cell. 91:243–252. 1997.
View Article : Google Scholar : PubMed/NCBI
|
|
77
|
DiDonato JA, Hayakawa M, Rothwarf DM,
Zandi E and Karin M: A cytokine-responsive IkappaB kinase that
activates the transcription factor NF-kappaB. Nature. 388:548–554.
1997. View Article : Google Scholar : PubMed/NCBI
|
|
78
|
Ghosh S and Karin M: Missing pieces in the
NF-kappaB puzzle. Cell. 109(Suppl): S81–S96. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
79
|
Wang HH, Garruti G, Liu M, Portincasa P
and Wang DQ: Cholesterol and lipoprotein metabolism and
atherosclerosis: Recent advances in reverse cholesterol transport.
Ann Hepatol. 16(Suppl 1): S27–S42. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
80
|
Khera AV and Rader DJ: Future therapeutic
directions in reverse cholesterol transport. Curr Atheroscler Rep.
12:73–81. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
81
|
Tall AR and Yvan-Charvet L: Cholesterol,
inflammation and innate immunity. Nat Rev Immunol. 15:104–116.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
82
|
Shao B, Tang C, Sinha A, Mayer PS,
Davenport GD, Brot N, Oda MN, Zhao XQ and Heinecke JW: Humans with
atherosclerosis have impaired ABCA1 cholesterol efflux and enhanced
high-density lipoprotein oxidation by myeloperoxidase. Circ Res.
114:1733–1742. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
83
|
Clarke MC and Bennett MR: Cause or
consequence: What does macrophage apoptosis do in atherosclerosis?
Arterioscler Thromb Vasc Biol. 29:153–155. 2009. View Article : Google Scholar
|
|
84
|
Monteiro R and Azevedo I: Chronic
inflammation in obesity and the metabolic syndrome. Mediators
Inflamm. 2010:2896452010. View Article : Google Scholar : PubMed/NCBI
|
|
85
|
Kalogeris T, Baines CP, Krenz M and
Korthuis RJ: Cell biology of ischemia/reperfusion injury. Int Rev
Cell Mol Biol. 298:229–317. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
86
|
Sanderson TH, Reynolds CA, Kumar R,
Przyklenk K and Hüttemann M: Molecular mechanisms of
ischemia-reperfusion injury in brain: Pivotal role of the
mitochondrial membrane potential in reactive oxygen species
generation. Mol Neurobiol. 47:9–23. 2013. View Article : Google Scholar :
|
|
87
|
Curek GD, Cort A, Yucel G, Demir N, Ozturk
S, Elpek GO, Savas B and Aslan M: Effect of astaxanthin on
hepatocellular injury following ischemia/reperfusion. Toxicology.
267:147–153. 2010. View Article : Google Scholar
|
|
88
|
Li J, Wang F, Xia Y, Dai W, Chen K, Li S,
Liu T, Zheng Y, Wang J, Lu W, et al: Astaxanthin pretreatment
attenuates hepatic ischemia reperfusion-induced apoptosis and
autophagy via the ROS/MAPK pathway in mice. Mar Drugs.
13:3368–3387. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
89
|
Cai X, Chen Y, Xie X, Yao D, Ding C and
Chen M: Astaxanthin prevents against lipopolysaccharide-induced
acute lung injury and sepsis via inhibiting activation of
MAPK/NF-κB. Am J Transl Res. 11:1884–1894. 2019.
|
|
90
|
Lauver DA, Lockwood SF and Lucchesi BR:
Disodium disuccinate astaxanthin (Cardax) attenuates complement
activation and reduces myocardial injury following
ischemia/reperfusion. J Pharmacol Exp Ther. 314:686–692. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
91
|
Ni Y, Nagashimada M, Zhuge F, Zhan L,
Nagata N, Tsutsui A, Nakanuma Y, Kaneko S and Ota T: Astaxanthin
prevents and reverses diet-induced insulin resistance and
steatohepatitis in mice: A comparison with vitamin E. Sci Rep.
5:171922015. View Article : Google Scholar : PubMed/NCBI
|
|
92
|
Liu H, Liu M, Fu X, Zhang Z, Zhu L, Zheng
X and Liu J: Astaxanthin prevents alcoholic fatty liver disease by
modulating mouse gut microbiota. Nutrients. 10:12982018. View Article : Google Scholar :
|
|
93
|
Lyu Y, Wu L, Wang F, Shen X and Lin D:
Carotenoid supple-mentation and retinoic acid in immunoglobulin A
regulation of the gut microbiota dysbiosis. Exp Biol Med (Maywood).
243:613–620. 2018. View Article : Google Scholar
|
|
94
|
Tang WH, Kitai T and Hazen SL: Gut
microbiota in cardiovascular health and disease. Circ Res.
120:1183–1196. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
95
|
Saad MJ, Santos A and Prada PO: Linking
gut microbiota and inflammation to obesity and insulin resistance.
Physiology (Bethesda). 31:283–293. 2016.
|
|
96
|
Wu L, Lyu Y, Srinivasagan R, Wu J, Ojo B,
Tang M, El-Rassi GD, Metzinger K, Smith BJ, Lucas EA, et al:
Astaxanthin-shifted gut microbiota is associated with inflammation
and metabolic homeostasis in mice. J Nutr. 150:2687–2698. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
97
|
Ou W, Liao Z, Yu G, Xu H, Liang M, Mai K
and Zhang Y: The effects of dietary astaxanthin on intestinal
health of juvenile tiger puffer takifugu rubripes in terms of
antioxidative status, inflammatory response and microbiota.
Aquaculture Nutrition. 25:466–476. 2018.
|
|
98
|
Zhang L, Cao W, Gao Y, Yang R, Zhang X, Xu
J and Tang Q: Astaxanthin (ATX) enhances the intestinal mucosal
functions in immunodeficient mice. Food Funct. 11:3371–3381. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
99
|
Gao Y, Yang L, Chin Y, Liu F, Li RW, Yuan
S, Xue C, Xu J and Tang Q: Astaxanthin n-octanoic acid diester
ameliorates insulin resistance and modulates gut microbiota in
high-fat and high-sucrose dietfed mice. Int J Mol Sci. 21:21492020.
View Article : Google Scholar
|
|
100
|
Wang J, Liu S, Wang H, Xiao S, Li C, Li Y
and Liu B: Xanthophyllomyces dendrorhous-derived astaxanthin
regulates lipid metabolism and gut microbiota in obese mice induced
by a high-fat diet. Mar Drugs. 17:3372019. View Article : Google Scholar :
|
|
101
|
Linton MRF, Yancey PG, Davies SS, Jerome
WG, Linton EF, Song WL, Doran AC and Vickers KC: The role of lipids
and lipoproteins in atherosclerosis. Endotext Feingold KR, Anawalt
B, Boyce A, Chrousos G, Dungan K, Grossman A, Hershman JM, Kaltsas
G, Koch C, Kopp P, et al: MDText Com, Inc; South Dartmouth, MA:
2000
|
|
102
|
Strassheim D, Dempsey EC, Gerasimovskaya
E, Stenmark K and Karoor V: Role of inflammatory cell subtypes in
heart failure. J Immunol Res. 2019:21640172019. View Article : Google Scholar : PubMed/NCBI
|
|
103
|
Gröschel C, Sasse A, Röhrborn C, Monecke
S, Didié M, Elsner L, Kruse V, Bunt G, Lichtman AH, Toischer K, et
al: T helper cells with specificity for an antigen in
cardiomyocytes promote pressure overload-induced progression from
hypertrophy to heart failure. Sci Rep. 7:159982017. View Article : Google Scholar : PubMed/NCBI
|
|
104
|
Fukunaga T, Soejima H, Irie A, Sugamura K,
Oe Y, Tanaka T, Nagayoshi Y, Kaikita K, Sugiyama S, Yoshimura M, et
al: Relation between CD4+ T-cell activation and severity of chronic
heart failure secondary to ischemic or idiopathic dilated
cardio-myopathy. Am J Cardiol. 100:483–488. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
105
|
Kallikourdis M, Martini E, Carullo P,
Sardi C, Roselli G, Greco CM, Vignali D, Riva F, Ormbostad Berre
AM, Stølen TO, et al: T cell costimulation blockade blunts pressure
overload-induced heart failure. Nat Commun. 8:146802017. View Article : Google Scholar : PubMed/NCBI
|
|
106
|
Gisterå A and Hansson GK: The immunology
of atherosclerosis. Nat Rev Nephrol. 13:368–380. 2017. View Article : Google Scholar
|
|
107
|
Swirski FK and Nahrendorf M: Leukocyte
behavior in athero-sclerosis, myocardial infarction, and heart
failure. Science. 339:161–166. 2013. View Article : Google Scholar
|
|
108
|
Stemme S, Faber B, Holm J, Wiklund O,
Witztum JL and Hansson GK: T lymphocytes from human atherosclerotic
plaques recognize oxidized low density lipoprotein. Proc Natl Acad
Sci USA. 92:3893–3897. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
109
|
Frostegård J, Ulfgren AK, Nyberg P, Hedin
U, Swedenborg J, Andersson U and Hansson GK: Cytokine expression in
advanced human atherosclerotic plaques: Dominance of
pro-inflammatory (Th1) and macrophage-stimulating cytokines.
Atherosclerosis. 145:33–43. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
110
|
Tripathi DN and Jena GB: Intervention of
astaxanthin against cyclophosphamide-induced oxidative stress and
DNA damage: A study in mice. Chem Biol Interact. 180:398–406. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
111
|
Bolin AP, Guerra BA, Nascimento SJ and
Otton R: Changes in lymphocyte oxidant/antioxidant parameters after
carbonyl and antioxidant exposure. Int Immunopharmacol. 14:690–697.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
112
|
Bolin AP, Macedo RC, Marin DP, Barros MP
and Otton R: Astaxanthin prevents in vitro auto-oxidative injury in
human lymphocytes. Cell Biol Toxicol. 26:457–467. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
113
|
Campoio TR, Oliveira FA and Otton R:
Oxidative stress in human lymphocytes treated with fatty acid
mixture: Role of carotenoid astaxanthin. Toxicol In Vitro.
25:1448–1456. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
114
|
Pilinska MA, capital Ka CD, Rushkovsky SR
and Dybska OB: Genoprotective properties of astaxanthin revealed by
ionizing radiation exposure in vitro on human peripheral blood
lymphocytes. Probl Radiac Med Radiobiol. 21:141–148. 2016.In
English, Ukrainian. View Article : Google Scholar : PubMed/NCBI
|
|
115
|
Otton R, Marin DP, Bolin AP, de Cássia
Santos Macedo R, Campoio TR, Fineto C Jr, Guerra BA, Leite JR,
Barros MP and Mattei R: Combined fish oil and astaxanthin
supplementation modulates rat lymphocyte function. Eur J Nutr.
51:707–718. 2012. View Article : Google Scholar
|
|
116
|
Mahmoud FF, Haines DD, Abul HT, Abal AT,
Onadeko BO and Wise JA: In vitro effects of astaxanthin combined
with ginkgolide B on T lymphocyte activation in peripheral blood
mononuclear cells from asthmatic subjects. J Pharmacol Sci.
94:129–136. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
117
|
Lin KH, Lin KC, Lu WJ, Thomas PA,
Jayakumar T and Sheu JR: Astaxanthin, a carotenoid, stimulates
immune responses by enhancing IFN-γ and IL-2 secretion in primary
cultured lymphocytes in vitro and ex vivo. Int J Mol Sci.
17:442015. View Article : Google Scholar
|
|
118
|
Diao W, Chen W, Cao W, Yuan H, Ji H, Wang
T, Chen W, Zhu X, Zhou H, Guo H and Zhao X: Astaxanthin protects
against renal fibrosis through inhibiting myofibroblast activation
and promoting CD8+ T cell recruitment. Biochim Biophys
Acta Gen Subj. 1863:1360–1370. 2019. View Article : Google Scholar
|
|
119
|
Park JS, Mathison BD, Hayek MG, Massimino
S, Reinhart GA and Chew BP: Astaxanthin stimulates cell-mediated
and humoral immune responses in cats. Vet Immunol Immunopathol.
144:455–461. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
120
|
Chew BP, Wong MW, Park JS and Wong TS:
Dietary beta-carotene and astaxanthin but not canthaxanthin
stimulate splenocyte function in mice. Anticancer Res.
19:5223–5227. 1999.
|
|
121
|
Jyonouchi H, Hill RJ, Tomita Y and Good
RA: Studies of immunomodulating actions of carotenoids. I. Effects
of beta-carotene and astaxanthin on murine lymphocyte functions and
cell surface marker expression in in vitro culture system. Nutr
Cancer. 16:93–105. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
122
|
Mahmoud FF, Haines D, Al-Awadhi R,
Arifhodzic N, Abal A, Azeamouzi C, Al-Sharah S and Tosaki A: In
vitro suppression of lymphocyte activation in patients with
seasonal allergic rhinitis and pollen-related asthma by cetirizine
or azelastine in combination with ginkgolide B or astaxanthin. Acta
Physiol Hung. 99:173–184. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
123
|
Jyonouchi H, Zhang L and Tomita Y: Studies
of immunomodulating actions of carotenoids. II. Astaxanthin
enhances in vitro antibody production to T-dependent antigens
without facilitating polyclonal B-cell activation. Nutr Cancer.
19:269–280. 1993. View Article : Google Scholar : PubMed/NCBI
|