Estrogen plays an important role in the female
health and fertility status. It is mainly derived from 3 sources:
Estrone is mainly converted from androstenedione of the adrenal
gland through the skin and adipose tissue (9,10); estradiol (E2), the most widely
effective estrogen, is mainly produced by GCs in the ovaries, and
is the main estrogen product synthesized before menopause; estriol
(E3), the weakest estrogen, is mainly synthesized in the placenta.
Since the ovaries are the main organs which secrete estrogen, the
normal expression of aromatase is of utmost importance. Presently,
the association between aromatase and the ovarian GCs, as well as
the regulatory mechanisms of this enzyme in GCs remain undefined.
The present review thus focused on aromatase expression and the
molecular regulatory mechanisms in ovarian GCs in order to help
interpret estrogen disorders. Potential aromatase inhibitors (AIs)
are also discussed an effort to open new research avenues for
hormone-dependent diseases and fertility treatment influenced by
estrogen-secretion disorders (Table
I).
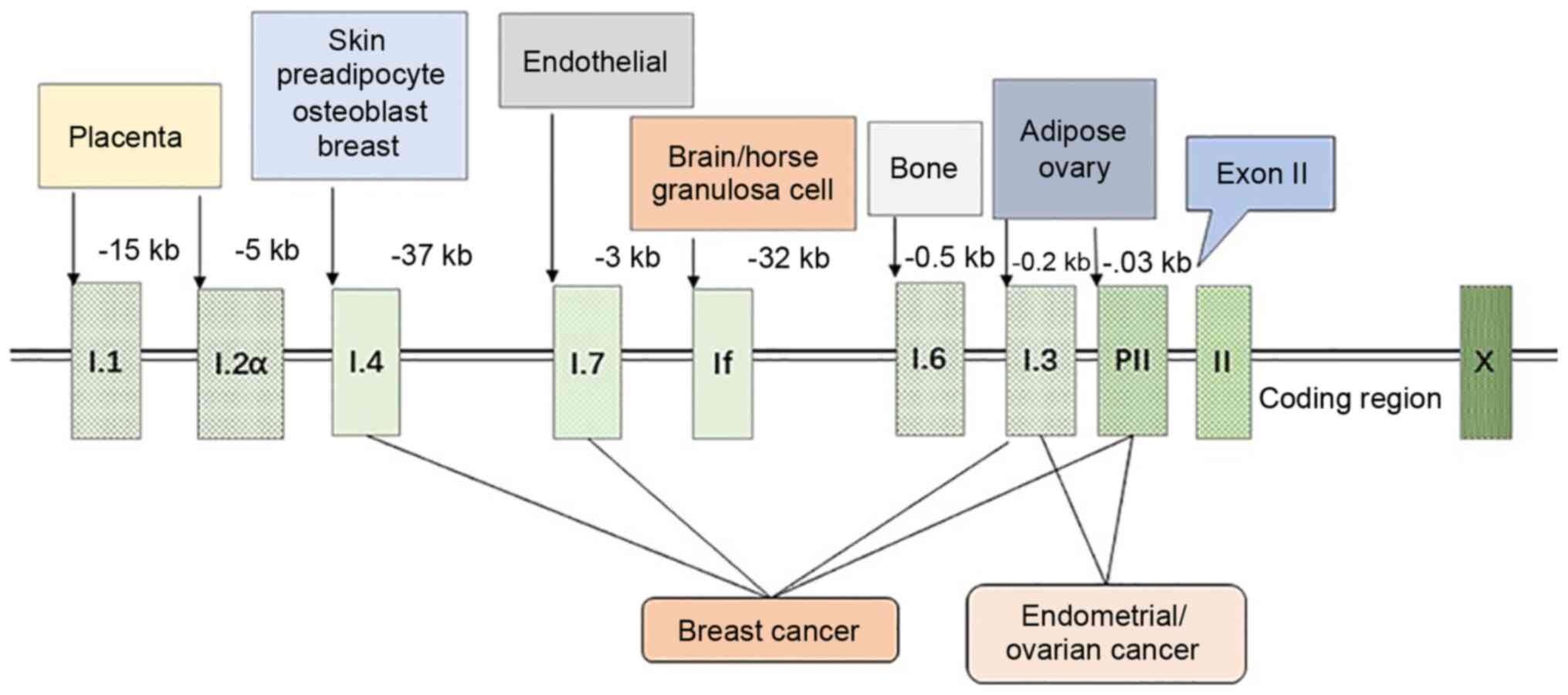
The expression of aromatase in different tissues is
controlled by distinct promoters, and there are >10 promoters
that function for selective purposes and tissue specificity. These
promoters control the expression of aromatase by recruiting
different transcription factors and combining with the
cis-regulatory elements (Fig.
1) (11). The transcription
of its genes is mainly controlled by the distal promoter I.1 of the
placenta (at 40-kb upstream of the translation start site) and the
proximal promoter II of the ovary. The aromatase promoter I.4 plays
an important role in regulating aromatase expression in the skin;
aromatase expression in the adipose tissue is controlled by the
ovarian promoter II, as well as by the promoters between I.1 and II
(i.e., I.4 and I. 3) (8). The
ovaries (promoter II), brain, tissue-specific promoter (If) and
bone (promoter I.4 and I.6) are mainly involved in the
transcription of the aromatase gene (12,13).
The abnormal regulation of promoters can result in
the development of various diseases. For example, promoter
transition from I.4 to I.3/II causes the excessive production of
estrogen in the breast, rendering breast epithelial cells
cancerous, thereby causing breast cancer (11). Similarly, immoderate estrogen
secretion due to the mutation in the promoter I.3/II results in
endometrial cancer and ovarian cancer, and a cAMP-responsive
element (CRE) binding protein (CREB) between promoter I.3/II
regulates aromatase through the cAMP/PKA-dependent pathway, which
results in the disordered expression of aromatase (14,15).
As mentioned above, the transcription of the
aromatase gene is regulated by tissue-specific promoters. This
point raises the question of how aromatase is expressed and
localized in GCs. The ability of GCs to produce steroids is mainly
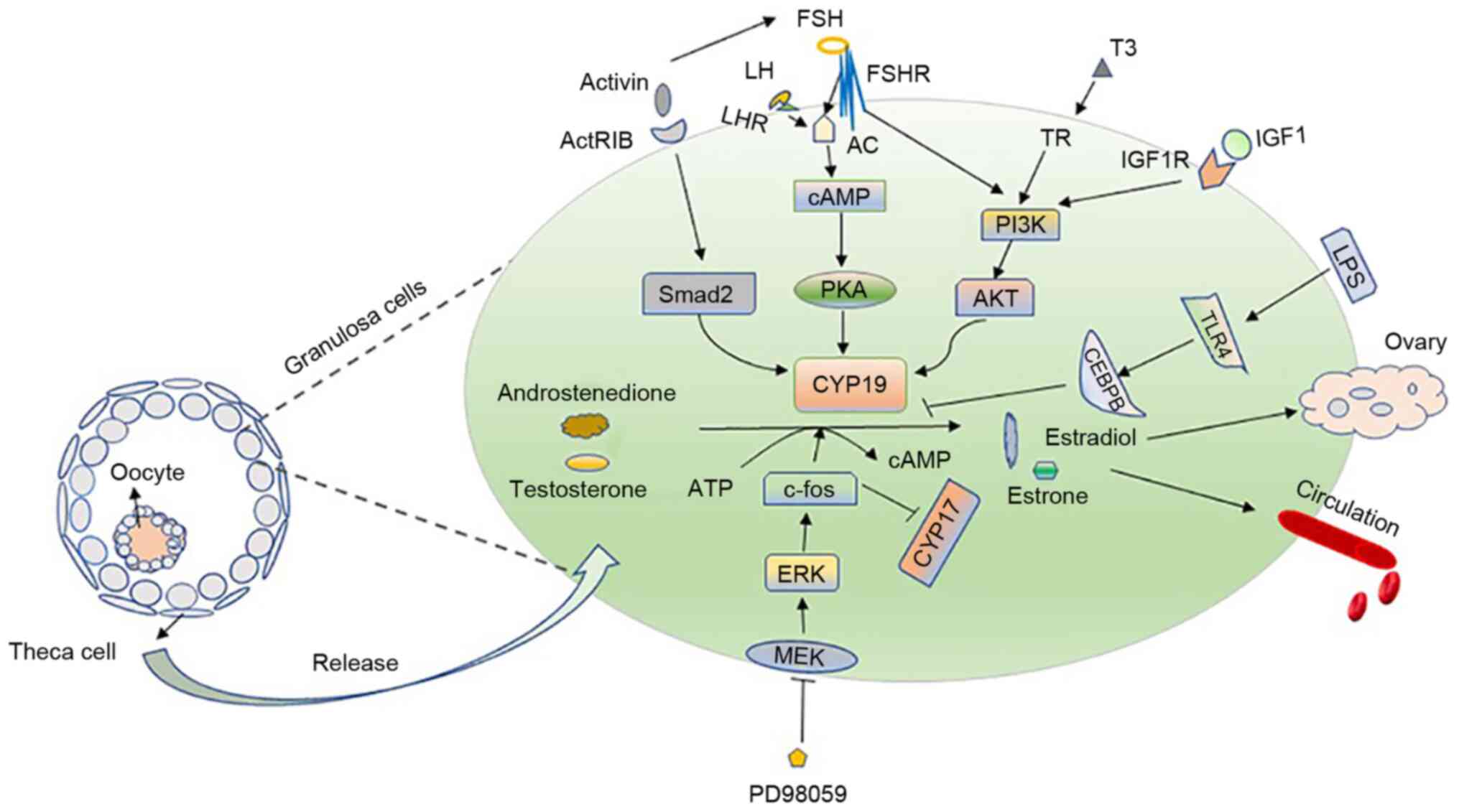
controlled by FSH. FSH binds to its receptor, and this receptor
interacts with adenylate cyclase (AC) to regulate aromatase through
the protein kinase A (PKA) pathway (22). In the presence of steroids, the
expression of P450arom mRNA in GCs is increased rapidly under the
effect of FSH/cAMP (23). The
literature confirms that the addition of the C19 steroid substrates
(androstenedione and testosterone) to the culture medium of GCs can
accelerate the synthesis of estradiol in a concentration- and
time-dependent manner (24).
FSH can regulate the maturation and proliferation of
ovarian GCs and binds to the specific receptor FSHR of Sertoli
cells and GCs (29). Previous
studies have reported that FSH can activate cAMP signaling in
vitro or in vivo to enhance the expression of aromatase
in ovarian GCs (30-32). FSH is the main factor that
stimulates the expression of aromatase CYP19A1. In GCs, it
can upregulate the expression of CYP19A1 by activating the
transcription factor GATA4. Simultaneously, the activation of GATA4
requires the activation of other kinases, such as ERK1/2, PKA and
PI3K (33). Aromatase activity
of immature rat GCs increases under the action of FSH and LH
(32). E2 can enhance this
effect of FSH, which supports the '2-cell, 2-gonadotropin
hypothesis' of ovarian synthesis of estrogen. In addition, FSH not
only enhances the activity of aromatase in a time-dependent manner,
but also enhances the aromatase complex component-flavin protein
NADPH-cytochrome P-450 reductase (34). Previous research has reported
that Anti-Müllerian hormone (AMH) (35) and virus analog (36) inhibit the increase in aromatase
expression and estrogen production induced by FSH. The presence of
diethylstilbestrol and androstenedione can enable FSH to increase
the expression of aromatase in a dose-dependent manner, leading to
increased estrogen secretion. According to the stimulating effect
of FSH on the aromatase activity of rat GCs, it can be used as a
biological assay for GC aromatase (37). Moreover, the bone morphogenetic
protein (BMP) family plays an important role in human ovarian
development; for example, BMP-2 (38), BMP-6 (39) and BMP-7 (40) can reduce the quantity of LH
receptor in hGCs and increase the expression of FSH receptor, while
BMP-2 can increase the expression of aromatase in GC cells.
Cyclic adenosine monophosphate (cAMP) can
transcriptionally regulate aromatase. The inhibitory effect of
progesterone and R5020 on GCs aromatase in rat ovaries has been
found to be dose-dependent; moreover, HCG combined with FSH can
significantly increase the amount of cAMP and aromatase activity is
enhanced at the appropriate concentrations (41). After adenylate cyclase is
activated by forskolin, it rapidly, but transiently induces
LH-responsive genes [such as the nuclear receptor 4A subfamily;
e.g., nerve growth factor IB (NGFI-B), nuclear receptor-related 1
(NURR1) and neuron-derived orphan receptor 1 (NOR-1)]. However, the
ectopic expression of NURR1 or NGFI-B can attenuate the
cAMP-responsive activation of the aromatase promoter in KGN cells.
Therefore, the expression of aromatase mRNA may be closely related
to the induction of nuclear receptor subfamily 4A (42). In addition,
luteolin-7-methylether (XLY29) can inhibit the phosphorylation of
the cAMP response element binding protein by regulating the
expression of aromatase promoter 1.3/II, thereby reducing mouse
serum 17β-estradiol levels (43). The expression of CYP19A1
is thus closely related to the regulation of FSH/cAMP.
In recent years, miRNAs have become increasingly
popular and discovered as drug targets. KGN cells treated with FSH
siRNA can induce the differential expression of miR-329-3p,
miR-1261, miR-144-5p, miR-130a-3p, miR-185-5p and miR-4463, among
which miR-4463 has been found to target the expression of ESR1 and
CYP19A1 to influence estradiol secretion (44). In addition, miR-378 (45), miR-10b (19)and miR-1275 (46) combine with the 3′ untranslated
region (3′-UTR) to downregulate the effective transcription of
CYP19A1 in ovarian GCs. miR-764-3p can inhibit the
expression of CYP19A1 by targeting steroidogenic factor-1
(SF-1) to reduce the production of 17β-estradiol (47). Through in vitro and in
vivo verifications, miR-326 has been found to activate C/EBP-β
by upregulating CREB, thereby inhibiting the expression of
CYP19A1 in buffalo GCs and causing a decrease in the
17β-estradiol levels (48).
Moreover, miR-214-3p has been found to upregulate the expression of
cell cycle genes in porcine GCs, but to downregulate the mRNA
expression levels of CYP19A1 and steroidogenic acute
regulatory protein (StAR) (49).
The overexpression of miR-29a in cov434 and KGN cells can cause the
expression of aromatase to decrease, affecting the estradiol
secretion and inhibiting the cell proliferation (50). In a previous study, a GC model
treated with DOX led to the increased expression of miR-132-3p in a
dose-dependent manner, as well as in a change in the expression of
CYP19A1 and the secretion of 17β-estradiol (51).
Aromatase activity in GCs is regulated by various
growth factors. Cell location studies have indicated that the
insulin-like growth factor-I (IGF1) gene and its receptor gene are
highly expressed and translated in ovarian GCs (52,53). IGF1 can increase the production
of P-450arom in a concentration-dependent manner and can increase
the activity of aromatase. In addition, IGF1 can cooperate with FSH
and is more obvious than any hormone alone (54). However, epidermal growth factor
(EGF) can inhibit the synthesis of P450 and antagonize FSH. These
changes are likely to be related to the mRNA encoding level of
aromatase (55). In addition,
amphiregulin (AREG) in the EGF-like growth factor induced by HCG
can activate the AKT signaling pathway and upregulate the
expression of aromatase and estrogen in human granulosa lutein
(hGL) cells (56). Moreover,
fibroblast growth factor (FGF) and platelet-derived growth factor
(PDGF) can reduce the effect of cAMP and may act based on the
corresponding tyrosine kinase activity to regulate the synthesis of
cytochrome P450 in GCs (57).
Some growth factors, such as vascular endothelial growth factor A
(VEGF-A) and FGF2 can synergistically promote the expression of
CYP19A1 in buffalo GCs and enhance the extent of estradiol
secretion (58). Hepatocyte
growth factor (HGF) can also inhibit the expression of FSH-induced
cAMP-dependent P450arom and 17β-hydroxysteroid dehydrogenase (HSD),
thereby inhibiting GC steroid production in ovarian GCs (59). Transforming growth factor-β
(TGF-β) has also been reported to enhance the expression of
FSH-induced P450arom mRNA in rat GCs (60), as well as accelerate the
production of E1 and E2 by 1.45- and 2.7-fold. However, leptin has
been reported to antagonize this effect of TGF-β, which reduces the
expression of GC aromatase (61).
An increasing number of studies have indicated that
GCs are easily affected by environmental pollutants in their
ability to secrete estrogen. Bisphenol A (BPA), which is
industrially used as a plasticizer, has been reported to decrease
the aromatase expression in GC cell-line KGN stimulated by FSH to
reduce the estradiol secretion, which may function by inducing
peroxisome proliferator-activated receptor-γ (PPARγ) (62,63). In rats,
2,3,7,8-tetrachlorodibenzo-p-dioxin (TCDD) may reduce the aromatase
mRNA expression in GCs through the AHR/ARNT signaling pathway,
causing a decrease in the estradiol secretion (64,65). TCDD may also directly or
indirectly block the endocrine function of human luteinized GCs
(hLGCs) through the interaction of PTK/MAP2K and PKA signaling
(66,67). Plasticizer mono-(2-ethylhexyl)
phthalate and its metabolite mono-(2-ethylhexyl) phthalate (MEHP)
also reduce the transcription of aromatase in human and rat GCs,
and thereby reduce the secretion of estradiol (68-70). Recently, it was reported that
carbon black nanoparticles (CB NPs) can inhibit the transcription
level of aromatase in GCs by activating the ERK1/2 pathway and
influencing E2 secretion. In addition, PD98059 signals can inhibit
the ERK1/2 signaling pathway to reduce the adverse effects of CB
NPs (71). In addition, prenatal
nicotine exposure (PNE) can reduce histone 3 lysine 27 acetylation
(H3K27ac) and H3K9ac of P450arom in the ovarian GC cell line, KGN,
through nicotine acetylcholine receptor (nAChR), which consequently
results in the reduction of the aromatase expression and estradiol
production in the ovaries (72).
In GCs, lead may affect female fertility by downregulating the
transcription of estrogen receptor β (ERβ) and p450arom
(73).
Metformin is an insulin sensitizer with direct
function in the ovaries, which may downregulate the expression of
aromatase promoter I.4, I.3 and II through the MEK/ERK pathway,
thereby reducing the aromatase activity in GCs (76). In addition, the expression of
aromatase mRNA decreases in human luteinized GCs cultured with
metformin, although it can significantly increase the effect of
insulin on IGF1R mRNA and IR (77). Pioglitazone or rosiglitazone can
inhibit estrogen production in human ovarian GCs by interfering
with the combination of aromatase and androgens, and the effect of
thiazolidinediones (TZDs) in reducing the activity of aromatase may
explain the application of TZD in the treatment of
estrogen-dependent diseases and polycystic ovary syndrome (PCOS)
(78). Troglitazone is also a
compound of TZDs that is likely to exert a direct inhibitory effect
on aromatase in GCs through the nuclear receptor system PPARγ/RXR
heterodimer (79). Insulin
sensitizers thus have a probable potential as effective inhibitors
of aromatase that warrant further investigation (Table II).
The rapid activation of the PKA signaling pathway
also induces other cascading reactions. For example, FSH can
promote the activation of PI3K signaling in the rat GCs and cause
the rapid phosphorylation of its downstream branch point, AKT; as a
result, the expression of ovarian genes such as CYP19A1
increases (80). Moreover, it
can enhance the activity of HIF-1 downstream of the
PI3K/AKT/Raheb/mammalian target of rapamycin (mTOR) pathway in GCs
(83). IGF1 has been confirmed
to be an important molecule for hormone synthesis in GCs (76); it can activate PI3K/Akt, which
upregulates FSH receptor. When Akt and PKA are activated, they
regulate the expression of CYP19A1 (84). It has previously been confirmed
that the molecular mechanism of 3,5,3′-tri-iodothyronine (T3)
regulates the expression of CYP19A1 in GC and promotes
follicle development. The combination of FSH and T3 requires the
activation of the PI3K/Akt pathway to regulate CYP19A1.
Therefore, CYP19A1 is likely to be a downstream effector of
the PI3K/Akt pathway that is activated by FSH and T3, which acts as
a new mediator for FSH and T3 to induce the development of GCs and
follicles, while regulating FSH and TH (85).
The surge of LH prior to ovulation can rapidly
inhibit the proliferation of GCs and differentiate them into luteal
cells. However, the proper dose of LH can also cause the rapid
activation of ERK1/2 when acting on GCs, while FSH also have the
same effect (86). ERK, similar
to PKA, is a downstream signaling molecule involved in the action
of LH and FSH. This point raises a question of whether aromatase is
regulated as a downstream molecule of this signaling pathway. Huang
et al used PD98059 to block the hormone-stimulated MEK1 in
KGN cells and found that 17-hydroxylase/17,20 lyase (CYP17)
expression increased, and that the expression of P450 aromatase
(CYP19) and c-fos also decreased (87). Hence, c-fos is likely to be
downstream of the MEK/ERK pathway, and CYP19 is upregulated
and CYP17 is downregulated in GCs, which together inhibit
the production of estradiol (87).
Aromatase is the only enzyme in vertebrates that
catalyzes the production of estrogen. The disruption of estrogen
secretion can thus easily cause cancer of the breast, endometrium
or ovaries, which are together referred to as 'estrogen-dependent
tumors' (92). These tumors can
also cause female reproductive issues. GCs are the main source of
ovarian estrogen secretion, which not only induces ovarian
development but also releases estrogen into the blood and supply it
to various organs. Therefore, whether the secretion of GCs
aromatase is normal directly affects the health status of various
organs. Nowadays, the inhibition of aromatase-induced estrogen
biosynthesis is considered to be the main strategy for the therapy
of estrogen-dependent diseases. Considering that the aromatase
status of patients with recurrent granulosa cell tumors remains
unaltered, the detection of the expression of aromatase can provide
more accurate information for serum E2 in the primary tumor as a
relapse marker (93).
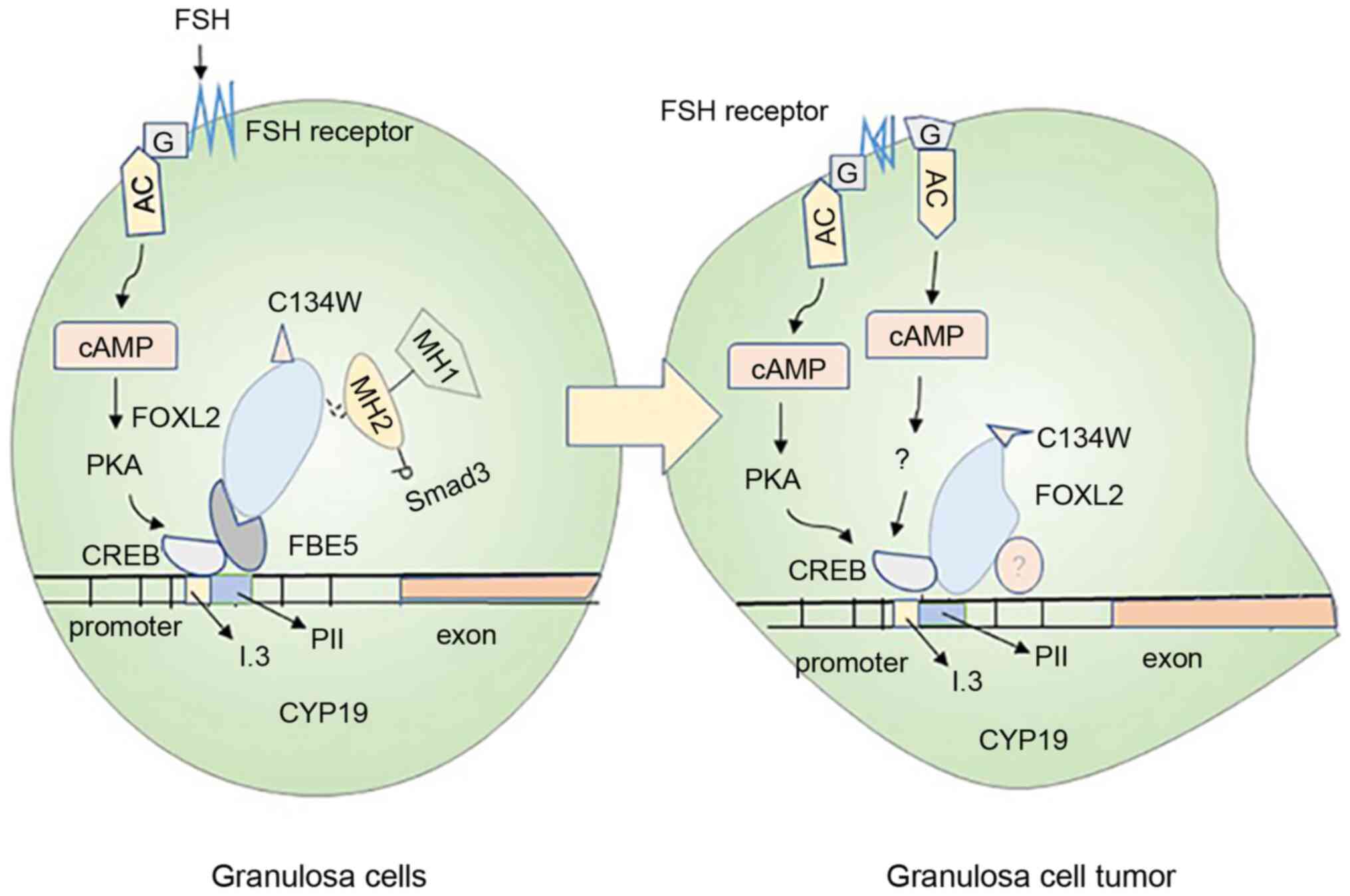
Ovarian GC tumors are one of the most common ovarian
tumors, mainly causing abnormal estrogen secretion (94) (Fig. 3). In normal GCs, FSH binds to its
receptor to activate Gs protein and upregulate the aromatase
expression through the PKA pathway. However, mutations in the Gs
protein on the cell membrane are likely to induce the production of
cancerous cells (95). The
forkhead family FOXL2 is an evolutionarily conserved member of the
transcription factor; its mutation is closely related to the
generation of ovarian GC tumor (96). Smad3 can cooperate with
FOXL2:C134W in combination with the CYP19A1 promoter FBE5 of
the human GC line (HGrC1) to enhance the expression of aromatase,
which is probably the key reason for the large amount of estrogen
secretion by the human GC line (97). The mutation of FOXL2:C134W is the
main reason for the formation of adult ovarian granulosa cell tumor
(GCT); it can enhance the induction of aromatase by FOXL2, and
aromatase is the direct target of FOXL2:C134W (98). This information deepens our
understanding of the research on the pathogenesis of the adult GC
tumor.
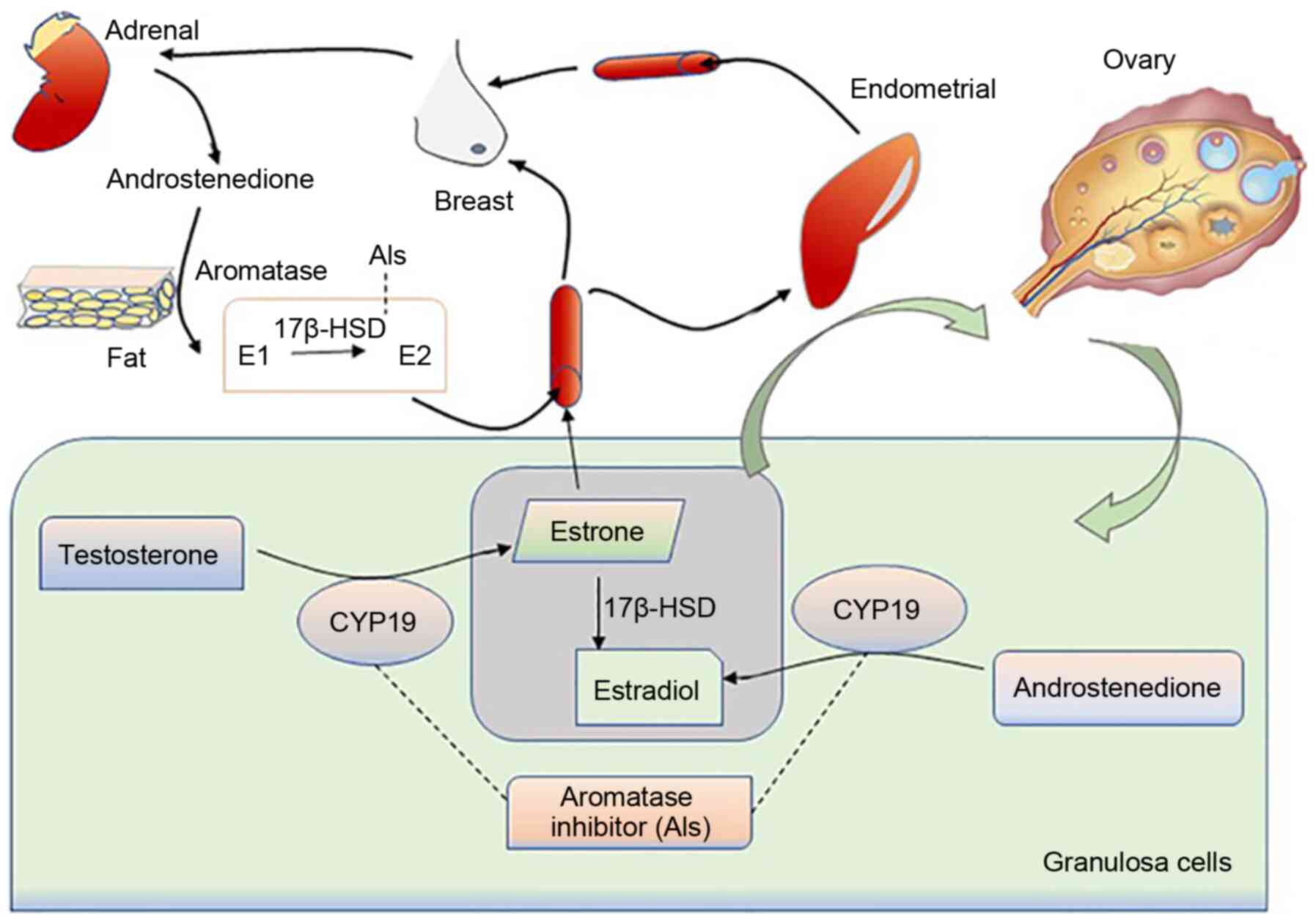
The abnormal secretion of estradiol from the GCs can
result in abnormalities in the target organs, such as the breast
and uterus (Fig. 4). Based on
the literature, the expression levels of aromatase in the diseased
tissues of the endometrium and breast are significantly higher than
those in normal tissues (99),
with adult ovarian cancer patients being more prone to endometrial
and breast cancer (100). In
addition, the increased expression of aromatase can also induce
endometriosis and uterine fibroids in females. In granulosa cells,
SF-1 can bind to the cAMP response element and act on aromatase
promoter II, thereby causing estrogen production (101). Abnormally expressed SF-1 in
endometriosis will compete with the aromatase expression
inhibitor-chicken ovalbumin upstream promoter-transcription factor
(COUP-TF) for the same binding site, thereby stimulating the
activity of aromatase P450 (102). Moreover, glucocorticoids and
IL1βcan affect the secretion of estrogen by regulating the
expression of aromatase promoter 1.4, thereby affecting the
occurrence of uterine fibroids (103).
The estrogen secretion disorder, which induces
increase in the incidence of PCOS, is the main cause of female
infertility (104). PCOS is
mainly manifested as a disease of the reproductive dysfunction and
endocrine disorders, which is mainly characterized by the high
levels of androgens and anovulation. The hyperandrogenic state of
the ovaries of patients with PCOS is the main risk factor for the
decline of aromatase production in luteinized GCs (105). It has been reported that
androgens can enhance the FSH effect in granulosa cells by
increasing FSHR. When androgen secretion is excessive, the high
sensitivity of GCs to FSH causes a large increase in AMH, and AMH
in turn inhibits the effect of FSH on aromatase in follicles
(106). Recently, Che et
al (107) found that the
highly expressed lncRNA HUPCOS of granulosa cells in patients with
PCOS can interact with RNA-binding protein with multiple splicing
(RBPMS) and inhibit the expression of aromatase, thereby mediating
the excess of androgen in the follicular fluid of PCOS patients
Therefore, the decrease in the aromatase activity conversely helps
maintain high androgen levels (102,107). In addition, CYP19A1 and
A-Kinase anchor protein 95 (AKAP95) levels are significantly
decreased in human luteinized GCs of patients with PCOS. AKAP95 is
thus likely to act on FSH-stimulated CYP19A1 (108). All of these participate in the
pathological mechanism of PCOS. Moreover, the abnormal expression
of aromatase mRNA in GCs can also induce obesity in women, which is
an important factor affecting female fertility. It has been
reported that leptin can induce neuropeptide cocaine- and
amphetamine-regulated transcript (CART) in GCs, thereby indirectly
inhibiting the cAMP levels and the aromatase expression in GCs,
causes ovarian dysfunction and reduced fertility (109).
Environmental pollution and modification in diets
are considered to be important elements affecting female fertility
and health issues. Several diseases have been confirmed in relation
to estrogen-secretion disorders in the epidemiology and
experimental studies. In some patients with estrogen-dependent
diseases, high levels of estrogen are accompanied by the
overexpression of aromatase (15). For example, in breast cancer
treatment, multiple AIs have been developed, such as exemestane,
anastrozole, letrozole and vorozole. The current first-line
treatment for breast cancer mainly uses third-generation AIs
(110,111). In addition, AIs have begun to
be used in the treatment of estrogen-regulated diseases, such as
ovarian cancer and endometrial cancer, as well as inducing
ovulation. Presently, AIs combined with progesterone and GnRH
agonists are mainly used to treat women with endometriosis, along
with reducing the risk of ovarian cysts induced by AIs alone.
However, symptoms, such as pelvic pain may recur following AI
treatment. For women with polycystic ovary syndrome and obesity,
letrozole has a higher live birth rate compared with clomiphene
citrate, and it has been regarded as the first-line therapy of
inducing ovulation. Moreover, AIs are also a good first choice for
women with infertility due to the presence of uterine fibroids, the
wish to preserve the uterus, or being unsuitable for surgery
(112).
Although AIs are effective in the treatment of
estrogen-dependent diseases and ovulation induction,
osteoarthropathy, menopausal symptoms, intestinal discomfort and
drug resistance, which are easily induced following treatment with
AIs, remain concerns which require resolutions. A recent study
designed and synthesized a novel aromatase inhibitor based on
triazole and imidazole (113).
However, the current inhibitors are mainly used in the treatment of
breast cancer, albeit for other steroid-dependent diseases and
female infertility, and there are a relatively few effective and
specific drugs available (114,115), which indicates that the search
for novel AIs and other effective drug targets is crucial.
GCs, which are the main site of ovarian estrogen
production, also generate a large amount of aromatase. Aromatase
can promote the biosynthesis of estrogen in GCs, and the production
of estrogen can promote the follicular development of GCs and
inhibit the apoptosis of GCs. The transcription of genes is mainly
controlled by the distal promoter I.1 of the placenta (at 40 kb
upstream of the translation start site) and the proximal promoter
II of the ovary (8). FSH can
regulate the expression of aromatase by activating the PKA
signaling pathway, while simultaneously activating the PI3K
(85), ERK (87) and other signaling pathways,
thereby upregulating the expression of CYP19A1. The
disadvantage of this approach is the lack of substantial research
supporting whether the transient activation of P38MAPK caused by
FSH can also regulate the expression of aromatase. In addition,
owing to biodiversity, individual differences exist in the
regulation of aromatase due to different factors, which may present
new areas of interest in future research. Understanding the
molecular modifications and the mechanisms of action in GCs that
can be targeted in the disorders of aromatase secretion for the
treatment of diseases caused by ovarian hormones is expected to
develop high-efficiency, low-toxic, and side effects of specific
drug targets.
The present article summarized and discussed the
regulatory characteristics of the CYP19A1 promoter in
ovarian GCs, as well as the regulation of aromatase as a downstream
effector through multiple signaling pathways. The expression of
aromatase in GCs is affected by various factors, and it is one of
the main causes of estrogen-dependent diseases and PCOS.
Endocrine-disrupting substances in the environment can cause
alteration in the expression of aromatase and affect the normal
reproduction and sexual differentiation in the human body.
Furthermore, some insulin sensitizers have been developed as AIs
for the clinical treatment. Other substances, such as FSH, IGF-1
and TGFβ have been proven to promote the expression of aromatase
mRNA and protein, although miRNAs, HGF and leptin can partially
inhibit or specifically identify the aromatase promoter and
downregulate the aromatase transcription, leading to the
disturbance in estrogen secretion. Thus, the discovery of potential
aromatase inhibitor targets is expected to provide new directions
for the treatment of estrogen-dependent diseases and PCOS.
Not applicable.
HL, TL and YH designed and wrote the manuscript. All
authors have read and approved the final manuscript.
Not applicable.
Not applicable.
The authors declare that they have no competing
interests.
Not applicable.
The present study was supported by grants from the National
Natural Science Foundation of China (no. 31900852 to HL), Nanchang
University (no. PY201801 to HL) and the Natural Science Foundation
of Jiangxi Province (nos. 2018BAB215012 and 20192ACB21026 to
HL).
|
1
|
Mendelson CR, Jiang B, Shelton JM,
Richardson JA and Hinshelwood MM: Transcriptional regulation of
aromatase in placenta and ovary. J Steroid Biochem Mol Biol.
95:25–33. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Li J and Gibbs RB: Detection of estradiol
in rat brain tissues: Contribution of local versus systemic
production. Psychoneuroendocrinology. 102:84–94. 2019. View Article : Google Scholar
|
|
3
|
Lambard S, Silandre D, Delalande C,
Denis-Galeraud I, Bourguiba S and Carreau S: Aromatase in testis:
Expression and role in male reproduction. J Steroid Biochem Mol
Biol. 95:63–69. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Mahendroo MS, Mendelson CR and Simpson ER:
Tissue-specific and hormonally controlled alternative promoters
regulate aromatase cytochrome P450 gene expression in human adipose
tissue. J Biol Chem. 268:19463–19470. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Wang Y, Pan P, Li X, Zhu Q, Huang T and Ge
RS: Food components and environmental chemicals of inhibiting human
placental aromatase. Food Chem Toxicol. 128:46–53. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Ai A, Tang Z, Liu Y, Yu S, Li B, Huang H,
Wang X, Cao Y and Zhang W: Characterization and identification of
human immortalized granulosa cells derived from ovarian follicular
fluid. Exp Ther Med. 18:2167–2177. 2019.PubMed/NCBI
|
|
7
|
Shoham Z, Jacobs HS and Insler V:
Luteinizing hormone: Its role, mechanism of action, and detrimental
effects when hyper-secreted during the follicular phase. Fertil
Steril. 59:1153–1161. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Nelson LR and Bulun SE: Estrogen
production and action. J Am Acad Dermatol. 45(Suppl 3): pp.
S116–S124. 2001, View Article : Google Scholar
|
|
9
|
Slominski A, Zbytek B, Nikolakis G, Manna
PR, Skobowiat C, Zmijewski M, Li W, Janjetovic Z, Postlethwaite A,
Zouboulis CC and Tuckey RC: Steroidogenesis in the skin:
Implications for local immune functions. J Steroid Biochem Mol
Biol. 137:107–123. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Bulun SE, Chen D, Moy I, Brooks DC and
Zhao H: Aromatase, breast cancer and obesity: A complex
interaction. Trends Endocrinol Metab. 23:83–89. 2012. View Article : Google Scholar :
|
|
11
|
Zhao H, Zhou L, Shangguan AJ and Bulun SE:
Aromatase expression and regulation in breast and endometrial
cancer. J Mol Endocrinol. 57:R19–R33. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Shozu M, Zhao Y and Simpson ER: TGF-beta1
stimulates expression of the aromatase (CYP19) gene in human
osteoblast-like cells and THP-1 cells. Mol Cell Endocrinol.
160:123–133. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Stocco C: Aromatase expression in the
ovary: Hormonal and molecular regulation. Steroids. 73:473–487.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Bulun SE, Chen D, Lu M, Zhao H, Cheng Y,
Demura M, Yilmaz B, Martin R, Utsunomiya H, Thung S, et al:
Aromatase excess in cancers of breast, endometrium and ovary. J
Steroid Biochem Mol Biol. 106:81–96. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Bulun SE and Simpson ER: Aromatase
expression in women's cancers. Adv Exp Med Biol. 630:112–132. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Sharma D, Ghai S and Singh D: Different
promoter usage for CYP19 gene expression in buffalo ovary and
placenta. Gen Comp Endocrinol. 162:319–328. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Solak KA, Wijnolts FMJ, Nijmeijer SM,
Blaauboer BJ, van den Berg M and van Duursen MBM: Excessive levels
of diverse phytoestrogens can modulate steroidogenesis and cell
migration of KGN human granulosa-derived tumor cells. Toxicol Rep.
1:360–372. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Ghosh S, Wu Y, Li R and Hu Y: Jun proteins
modulate the ovary-specific promoter of aromatase gene in ovarian
granulosa cells via a cAMP-responsive element. Oncogene.
24:2236–2246. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Li Q, Du X, Pan Z, Zhang L and Li Q: The
transcription factor SMAD4 and miR-10b contribute to E2 release and
cell apoptosis in ovarian granulosa cells by targeting CYP19A1. Mol
Cell Endocrinol. 476:84–95. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Andrieu T, Féral C, Joubert M, Benhaim A
and Mittre H: The absence of a functional nuclear receptor element
A (NREA) in the promoter II of the aromatase P450 gene in rabbit
granulosa cells. J Steroid Biochem Mol Biol. 101:127–135. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Boerboom D, Kerban A and Sirois J: Dual
regulation of promoter II- and promoter 1f-derived cytochrome P450
aromatase transcripts in equine granulosa cells during human
chorionic gonadotropin-induced ovulation: A novel model for the
study of aromatase promoter switching. Endocrinology.
140:4133–4141. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Simpson ER: Sources of estrogen and their
importance. J Steroid Biochem Mol Biol. 86:225–230. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Miyoshi T, Otsuka F and Shimasaki S: GRK-6
mediates FSH action synergistically enhanced by estrogen and the
oocyte in rat granulosa cells. Biochem Biophys Res Commun.
434:401–406. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Czajka-Oraniec I and Simpson ER: Aromatase
research and its clinical significance. Endokrynol Pol. 61:126–134.
2010.PubMed/NCBI
|
|
25
|
Velthut-Meikas A, Simm J, Tuuri T,
Tapanainen JS, Metsis M and Salumets A: Research resource: Small
RNA-seq of human granulosa cells reveals miRNAs in FSHR and
aromatase genes. Mol Endocrinol. 27:1128–1141. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Mlodawska W and Slomczynska M:
Immunohistochemical localization of aromatase during the
development and atresia of ovarian follicles in prepubertal horses.
Theriogenology. 74:1707–1712. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Naganuma H, Ohtani H, Harada N and Nagura
H: Immunoelectron microscopic localization of aromatase in human
placenta and ovary using microwave fixation. J Histochem Cytochem.
38:1427–1432. 1990. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Shaikh AA: Estrone and estradiol levels in
the ovarian venous blood from rats during the estrous cycle and
pregnancy. Biol Reprod. 5:297–307. 1971. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Szymańska K, Kałafut J, Przybyszewska A,
Paziewska B, Adamczuk G, Kiełbus M and Rivero-Müller A: FSHR
trans-activation and oligomerization. Front Endocrinol (Lausanne).
9:7602018. View Article : Google Scholar
|
|
30
|
Jiang C, Hou X, Wang C, May JV, Butnev VY,
Bousfield GR and Davis JS: Hypoglycosylated hFSH has greater
bioactivity than fully glycosylated recombinant hFSH in human
granulosa cells. J Clin Endocrinol Metab. 100:E852–E860. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Hobeika E, Armouti M, Kala H, Fierro MA,
Winston NJ, Scoccia B, Zamah AM and Stocco C: Oocyte-secreted
factors synergize with FSH to promote aromatase expression in
primary human cumulus cells. J Clin Endocrinol Metab.
104:1667–1676. 2019. View Article : Google Scholar :
|
|
32
|
Parakh TN, Hernandez JA, Grammer JC, Weck
J, Hunzicker- Dunn M, Zeleznik AJ and Nilson JH:
Follicle-stimulating hormone/cAMP regulation of aromatase gene
expression requires beta-catenin. Proc Natl Acad Sci USA.
103:12435–12440. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Kwintkiewicz J, Cai Z and Stocco C:
Follicle-stimulating hormone-induced activation of Gata4
contributes in the up-regulation of Cyp19 expression in rat
granulosa cells. Mol Endocrinol. 21:933–947. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Hong Y, Li H, Yuan YC and Chen S:
Molecular characterization of aromatase. Ann N Y Acad Sci.
1155:112–120. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Li Y, Gao D, Xu T, Adur MK, Zhang L, Luo
L, Zhu T, Tong X, Zhang D, Wang Y, et al: Anti-Müllerian hormone
inhibits luteinizing hormone-induced androstenedione synthesis in
porcine theca cells. Theriogenology. 142:421–432. 2020. View Article : Google Scholar
|
|
36
|
Fang Y, Wang B, Lyu S, Zhang K, Cheng Q
and Zhu Y: Virus analog decreases estradiol secretion in
FSH-treated human ovarian granulosa cells. Gynecol Endocrinol.
36:346–350. 2020. View Article : Google Scholar
|
|
37
|
Kajitani T, Liu S, Maruyama T, Uchida H,
Sakurai R, Masuda H, Nagashima T, Ono M, Arase T and Yoshimura Y:
Analysis of serum FSH bioactivity in a patient with an
FSH-secreting pituitary microadenoma and multicystic ovaries: A
case report. Hum Reprod. 23:435–439. 2008. View Article : Google Scholar
|
|
38
|
Shi J, Yoshino O, Osuga Y, Koga K, Hirota
Y, Nose E, Nishii O, Yano T and Taketani Y: Bone morphogenetic
protein-2 (BMP-2) increases gene expression of FSH receptor and
aromatase and decreases gene expression of LH receptor and StAR in
human granulosa cells. Am J Reprod Immunol. 65:421–427. 2011.
View Article : Google Scholar
|
|
39
|
Shi J, Yoshino O, Osuga Y, Koga K, Hirota
Y, Hirata T, Yano T, Nishii O and Taketani Y: Bone morphogenetic
protein-6 stimulates gene expression of follicle-stimulating
hormone receptor, inhibin/activin beta subunits, and anti-Müllerian
hormone in human granulosa cells. Fertil Steril. 92:1794–1798.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Shi J, Yoshino O, Osuga Y, Nishii O, Yano
T and Taketani Y: Bone morphogenetic protein 7 (BMP-7) increases
the expression of follicle-stimulating hormone (FSH) receptor in
human granulosa cells. Fertil Steril. 93:1273–1279. 2010.
View Article : Google Scholar
|
|
41
|
Overes HW, de Leeuw R and Kloosterboer HJ:
Regulation of aromatase activity in FSH-primed rat granulosa cells
in vitro by follicle-stimulating hormone and various amounts of
human chorionic gonadotrophin. Hum Reprod. 7:191–196. 1992.
View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Wu Y, Ghosh S, Nishi Y, Yanase T, Nawata H
and Hu Y: The orphan nuclear receptors NURR1 and NGFI-B modulate
aromatase gene expression in ovarian granulosa cells: A possible
mechanism for repression of aromatase expression upon luteinizing
hormone surge. Endocrinology. 146:237–246. 2005. View Article : Google Scholar
|
|
43
|
Du BW, Zhang XJ, Shi N, Peng T, Gao JB,
Azimova B, Zhang R, Pu DB, Wang C, Abduvaliev A, et al:
Luteolin-7-methylether from Leonurus japonicus inhibits estrogen
biosynthesis in human ovarian granulosa cells by suppression of
aromatase (CYP19). Eur J Pharmacol. 879:1731542020. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Lee SY, Kang YJ, Kwon J, Nishi Y, Yanase
T, Lee KA and Koong MK: miR-4463 regulates aromatase expression and
activity for 17β-estradiol synthesis in response to
follicle-stimulating hormone. Clin Exp Reprod Med. 47:194–206.
2020. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Xu S, Linher-Melville K, Yang BB, Wu D and
Li J: Micro-RNA378 (miR-378) regulates ovarian estradiol production
by targeting aromatase. Endocrinology. 152:3941–3951. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Liu J, Li X, Yao Y and Li Q, Pan Z and Li
Q: miR-1275 controls granulosa cell apoptosis and estradiol
synthesis by impairing LRH-1/CYP19A1 axis. Biochim Biophys Acta
Gene Regul Mech. 1861:246–257. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Wang L, Li C, Li R, Deng Y, Tan Y, Tong C
and Qi H: MicroRNA-764-3p regulates 17β-estradiol synthesis of
mouse ovarian granulosa cells by targeting steroidogenic factor-1.
In Vitro Cell Dev Biol Anim. 52:365–373. 2016. View Article : Google Scholar
|
|
48
|
Chaurasiya V, Kumari S, Onteru SK and
Singh D: miR-326 down-regulate CYP19A1 expression and estradiol-17b
production in buffalo granulosa cells through CREB and C/EBP-β. J
Steroid Biochem Mol Biol. 199:1056082020. View Article : Google Scholar
|
|
49
|
Shi S, Zhou X, Li J, Zhang L, Hu Y, Li Y,
Yang G and Chu G: MiR-214-3p promotes proliferation and inhibits
estradiol synthesis in porcine granulosa cells. J Anim Sci
Biotechnol. 11:942020. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Li Y, Liu YD, Zhou XY, Chen SL, Chen X,
Zhe J, Zhang J, Zhang QY and Chen YX: MiR-29a regulates the
proliferation, aromatase expression, and estradiol biosynthesis of
human granulosa cells in polycystic ovary syndrome. Mol Cell
Endocrinol. 498:1105402019. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Al-Kawlani B, Murrieta-Coxca JM,
Chaiwangyen W, Fröhlich K, Fritzsche A, Winkler S, Markert UR and
Morales-Prieto DM: Doxorubicin induces cytotoxicity and miR-132
expression in granulosa cells. Reprod Toxicol. 96:95–101. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Ogo Y, Taniuchi S, Ojima F, Hayashi S,
Murakami I, Saito Y, Takeuchi S, Kudo T and Takahashi S: IGF-1 gene
expression is differentially regulated by estrogen receptors α and
β in mouse endometrial stromal cells and ovarian granulosa cells. J
Reprod Dev. 60:216–223. 2014. View Article : Google Scholar :
|
|
53
|
Zhou J, Chin E and Bondy C: Cellular
pattern of insulin-like growth factor-I (IGF-I) and IGF-I receptor
gene expression in the developing and mature ovarian follicle.
Endocrinology. 129:3281–3288. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Mani AM, Fenwick MA, Cheng Z, Sharma MK,
Singh D and Wathes DC: IGF1 induces up-regulation of steroidogenic
and apoptotic regulatory genes via activation of
phosphatidylinositol-dependent kinase/AKT in bovine granulosa
cells. Reproduction. 139:139–151. 2010. View Article : Google Scholar
|
|
55
|
Herrmann M, Scholmerich J and Straub RH:
Influence of cytokines and growth factors on distinct steroidogenic
enzymes in vitro: A short tabular data collection. Ann NY Acad Sci.
966:166–186. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Fang L, Yu Y, Li Y, Wang S, Zhang R, Guo
Y, Li Y, Yan Y and Sun YP: Human chorionic gonadotropin-induced
amphiregulin stimulates aromatase expression in human
granulosa-lutein cells: A mechanism for estradiol production in the
luteal phase. Hum Reprod. 34:2018–2026. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Mendelson CR, Merrill JC, Steinkampf MP
and Simpson ER: Regulation of the synthesis of aromatase cytochrome
P-450 in human adipose stromal and ovarian granulosa cells.
Steroids. 50:51–59. 1987. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Mishra SR, Bharati J, Rajesh G, Chauhan
VS, Taru Sharma G, Bag S, Maurya VP, Singh G and Sarkar M:
Fibroblast growth factor 2 (FGF2) and vascular endothelial growth
factor A (VEGFA) synergistically promote steroidogenesis and
survival of cultured buffalo granulosa cells. Anim Reprod Sci.
179:88–97. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Zachow RJ, Ramski BE and Lee H: Modulation
of estrogen production and 17beta-hydroxysteroid dehydrogenase-type
1, cytochrome P450 aromatase, c-met, and protein kinase Balpha
messenger ribonucleic acid content in rat ovarian granulosa cells
by hepatocyte growth factor and follicle-stimulating hormone. Biol
Reprod. 62:1851–1857. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Chen YJ, Hsiao PW, Lee MT, Mason JI, Ke FC
and Hwang JJ: Interplay of PI3K and cAMP/PKA signaling, and
rapamycin-hypersensitivity in TGFbeta1 enhancement of
FSH-stimulated steroidogenesis in rat ovarian granulosa cells. J
Endocrinol. 192:405–419. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Zachow RJ, Weitsman SR and Magoffin DA:
Leptin impairs the synergistic stimulation by transforming growth
factor-beta of follicle-stimulating hormone-dependent aromatase
activity and messenger ribonucleic acid expression in rat ovarian
granulosa cells. Biol Reprod. 61:1104–1109. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Kwintkiewicz J, Nishi Y, Yanase T and
Giudice LC: Peroxisome proliferator-activated receptor-gamma
mediates bisphenol A inhibition of FSH-stimulated IGF-1, aromatase,
and estradiol in human granulosa cells. Environ Health Perspect.
118:400–406. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Bloom MS, Mok-Lin E and Fujimoto VY:
Bisphenol A and ovarian steroidogenesis. Fertil Steril.
106:857–863. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Dasmahapatra AK, Wimpee BA, Trewin AL and
Hutz RJ: 2,3,7,8-Tetrachlorodibenzo-p-dioxin increases steady-state
estrogen receptor-beta mRNA levels after CYP1A1 and CYP1B1
induction in rat granulosa cells in vitro. Mol Cell Endocrinol.
182:39–48. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Dasmahapatra AK, Wimpee BA, Trewin AL,
Wimpee CF, Ghorai JK and Hutz RJ: Demonstration of
2,3,7,8-tetrachloro-dibenzo-p-dioxin attenuation of P450
steroidogenic enzyme mRNAs in rat granulosa cell in vitro by
competitive reverse transcriptase-polymerase chain reaction assay.
Mol Cell Endocrinol. 164:5–18. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Enan E, Moran F, VandeVoort CA, Stewart
DR, Overstreet JW and Lasley BL: Mechanism of toxic action of
2,3,7,8-tetrachlorodibenzo-p-dioxin (TCDD) in cultured human
luteinized granulosa cells. Reprod Toxicol. 10:497–508. 1996.
View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Baldridge MG, Marks GT, Rawlins RG and
Hutz RJ: Very low-dose (femtomolar)
2,3,7,8-tetrachlorodibenzo-p-dioxin (TCDD) disrupts steroidogenic
enzyme mRNAs and steroid secretion by human luteinizing granulosa
cells. Reprod Toxicol. 52:57–61. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Lovekamp TN and Davis BJ:
Mono-(2-ethylhexyl) phthalate suppresses aromatase transcript
levels and estradiol production in cultured rat granulosa cells.
Toxicol Appl Pharmacol. 172:217–224. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Reinsberg J, Wegener-Toper P, van der Ven
K, van der Ven H and Klingmueller D: Effect of mono-(2-ethylhexyl)
phthalate on steroid production of human granulosa cells. Toxicol
Appl Pharmacol. 239:116–123. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Davis BJ, Weaver R, Gaines LJ and Heindel
JJ: Mono-(2-ethylhexyl) phthalate suppresses estradiol production
independent of FSH-cAMP stimulation in rat granulosa cells. Toxicol
Appl Pharmacol. 128:224–228. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Simon V, Avet C, Grange-Messent V,
Wargnier R, Denoyelle C, Pierre A, Dairou J, Dupret JM and
Cohen-Tannoudji J: Carbon black nanoparticles inhibit aromatase
expression and estradiol secretion in human granulosa cells through
the ERK1/2 pathway. Endocrinology. 158:3200–3211. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Fan G, Zhang Q, Wan Y, Lv F, Chen Y, Ni Y,
Zou W, Zhang W and Wang H: Decreased levels of H3K9ac and H3K27ac
in the promotor region of ovarian P450 aromatase mediated low
estradiol synthesis in female offspring rats induced by prenatal
nicotine exposure as well as in human granulosa cells after
nicotine treatment. Food Chem Toxicol. 128:256–266. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Taupeau C, Poupon J, Treton D, Brosse A,
Richard Y and Machelon V: Lead reduces messenger RNA and protein
levels of cytochrome p450 aromatase and estrogen receptor beta in
human ovarian granulosa cells. Biol Reprod. 68:1982–1988. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Morinaga H, Yanase T, Nomura M, Okabe T,
Goto K, Harada N and Nawata H: A benzimidazole fungicide, benomyl,
and its metabolite, carbendazim, induce aromatase activity in a
human ovarian granulose-like tumor cell line (KGN). Endocrinology.
145:1860–1869. 2004. View Article : Google Scholar
|
|
75
|
Zachow R and Uzumcu M: The methoxychlor
metabolite, 2,2-bis-(p-hydroxyphenyl)-1,1,1-trichloroethane,
inhibits steroidogenesis in rat ovarian granulosa cells in vitro.
Reprod Toxicol. 22:659–665. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
76
|
Rice S, Pellatt L, Ramanathan K, Whitehead
SA and Mason HD: Metformin inhibits aromatase via an extracellular
signal-regulated kinase-mediated pathway. Endocrinology.
150:4794–4801. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
77
|
Fuhrmeister IP, Branchini G, Pimentel AM,
Ferreira GD, Capp E, Brum IS and von Eye Corleta H: Human granulosa
cells: Insulin and insulin-like growth factor-1 receptors and
aromatase expression modulation by metformin. Gynecol Obstet
Invest. 77:156–162. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
78
|
Seto-Young D, Avtanski D, Parikh G,
Suwandhi P, Strizhevsky M, Araki T, Rosenwaks Z and Poretsky L:
Rosiglitazone and pioglitazone inhibit estrogen synthesis in human
granulosa cells by interfering with androgen binding to aromatase.
Horm Metab Res. 43:250–256. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
79
|
Mu YM, Yanase T, Nishi Y, Waseda N, Oda T,
Tanaka A, Takayanagi R and Nawata H: Insulin sensitizer,
troglitazone, directly inhibits aromatase activity in human ovarian
granulosa cells. Biochem Biophys Res Commun. 271:710–713. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
80
|
Gonzalez-Robayna IJ, Falender AE, Ochsner
S, Firestone GL and Richards JS: Follicle-Stimulating hormone (FSH)
stimulates phosphorylation and activation of protein kinase B
(PKB/Akt) and serum and glucocorticoid-lnduced kinase (Sgk):
Evidence for A kinase-independent signaling by FSH in granulosa
cells. Mol Endocrinol. 14:1283–1300. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
81
|
Donadeu FX and Ascoli M: The differential
effects of the gonado- tropin receptors on aromatase expression in
primary cultures of immature rat granulosa cells are highly
dependent on the density of receptors expressed and the activation
of the inositol phosphate cascade. Endocrinology. 146:3907–3916.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
82
|
Riccetti L, Sperduti S, Lazzaretti C,
Casarini L and Simoni M: The cAMP/PKA pathway: Steroidogenesis of
the antral follicular stage. Minerva Ginecol. 70:516–524. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
83
|
Alam H, Maizels ET, Park Y, Ghaey S,
Feiger ZJ, Chandel NS and Hunzicker-Dunn M: Follicle-stimulating
hormone activation of hypoxia-inducible factor-1 by the
phosphatidylinositol 3-kinase/AKT/Ras homolog enriched in brain
(Rheb)/mammalian target of rapamycin (mTOR) pathway is necessary
for induction of select protein markers of follicular
differentiation. J Biol Chem. 279:19431–19440. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
84
|
Zhou Y, Zeng C, Li X, Wu PL, Yin L, Yu XL,
Zhou YF and Xue Q: IGF-I stimulates ERβ and aromatase expression
via IGF1R/PI3K/AKT-mediated transcriptional activation in
endometriosis. J Mol Med (Berl). 94:887–897. 2016. View Article : Google Scholar
|
|
85
|
Liu J, Han Y, Tian Y, Weng X, Hu X, Liu W,
Heng D, Xu K, Yang Y and Zhang C: Regulation by
3,5,3′-tri-iodothyronine and FSH of cytochrome P450 family 19
(CYP19) expression in mouse granulosa cells. Reprod Fertil Dev.
30:1225–1233. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
86
|
Cottom J, Salvador LM, Maizels ET,
Reierstad S, Park Y, Carr DW, Davare MA, Hell JW, Palmer SS, Dent
P, et al: Follicle-stimulating hormone activates extracellular
signal-regulated kinase but not extracellular signal-regulated
kinase kinase through a 100-kDa phosphotyrosine phosphatase. J Biol
Chem. 278:7167–7179. 2003. View Article : Google Scholar
|
|
87
|
Huang X, Jin J, Shen S, Xia Y, Xu P, Zou
X, Wang H, Yi L, Wang Y and Gao Q: Modulation of expression of
17-Hydroxylase/17,20 lyase (CYP17) and P450 aromatase (CYP19) by
inhibition of MEK1 in a human ovarian granulosa-like tumor cell
line. Gynecol Endocrinol. 32:201–205. 2016. View Article : Google Scholar
|
|
88
|
Findlay JK: An update on the roles of
inhibin, activin, and follistatin as local regulators of
folliculogenesis. Biol Reprod. 48:15–23. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
89
|
Nomura M, Sakamoto R, Morinaga H, Wang L,
Mukasa C and Takayanagi R: Activin stimulates CYP19A gene
expression in human ovarian granulosa cell-like KGN cells via the
Smad2 signaling pathway. Biochem Biophys Res Commun. 436:443–448.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
90
|
Yenuganti VR, Ravinder and Singh D:
Endotoxin induced TLR4 signaling downregulates CYP19A1 expression
through CEBPB in buffalo granulosa cells. Toxicol In Vitro.
42:93–100. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
91
|
Wang Y, Lu E, Bao R, Xu P, Feng F, Wen W,
Dong Q, Hu C, Xiao L, Tang M, et al: Notch signalling regulates
steroidogenesis in mouse ovarian granulosa cells. Reprod Fertil
Dev. 31:1091–1103. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
92
|
Manna PR, Molehin D and Ahmed AU:
Dysregulation of aromatase in breast, endometrial, and ovarian
cancers: An overview of therapeutic strategies. Prog Mol Biol
Transl Sci. 144:487–537. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
93
|
Kato N, Uchigasaki S, Fukase M and Kurose
A: Expression of P450 aromatase in granulosa cell tumors and
sertoli-stromal cell tumors of the ovary: Which cells are
responsible for estrogenesis? Int J Gynecol Pathol. 35:41–47. 2016.
View Article : Google Scholar
|
|
94
|
Kitamura S, Abiko K, Matsumura N, Nakai H,
Akimoto Y, Tanimoto H and Konishi I: Adult granulosa cell tumors of
the ovary: A retrospective study of 30 cases with respect to the
expression of steroid synthesis enzymes. J Gynecol Oncol.
28:e312017. View Article : Google Scholar : PubMed/NCBI
|
|
95
|
Hsueh AJ, Adashi EY, Jones PB and Welsh TH
Jr: Hormonal regulation of the differentiation of cultured ovarian
granulosa cells. Endocr Rev. 5:76–127. 1984. View Article : Google Scholar : PubMed/NCBI
|
|
96
|
Cocquet J, Pailhoux E, Jaubert F, Servel
N, Xia X, Pannetier M, De Baere E, Messiaen L, Cotinot C, Fellous M
and Veitia RA: Evolution and expression of FOXL2. J Med Genet.
39:916–921. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
97
|
Belli M, Iwata N, Nakamura T, Iwase A,
Stupack D and Shimasaki S: FOXL2C134W-induced CYP19 expression via
cooperation with SMAD3 in HGrC1 cells. Endocrinology.
159:1690–1703. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
98
|
Fleming NI, Knower KC, Lazarus KA, Fuller
PJ, Simpson ER and Clyne CD: Aromatase is a direct target of FOXL2:
C134W in granulosa cell tumors via a single highly conserved
binding site in the ovarian specific promoter. PLoS One.
5:e143892010. View Article : Google Scholar : PubMed/NCBI
|
|
99
|
Leung K:
(S)-6-[(4-Chlorophenyl)(1H-1,2,4-triazol-1-yl)
methyl]-1-[(11)C]methyl-1H-benzotriazole. Molecular imaging and
contrast agent database (MICAD). National Center for Biotechnology
Information; Bethesda, MD: 2004
|
|
100
|
Moro F, Leombroni M, Pasciuto T,
Trivellizzi IN, Mascilini F, Ciccarone F, Zannoni GF, Fanfani F,
Scambia G and Testa AC: Synchronous primary cancers of endometrium
and ovary vs endometrial cancer with ovarian metastasis: An
observational study. Ultrasound Obstet Gynecol. 53:827–835.
2019.PubMed/NCBI
|
|
101
|
Michael MD, Kilgore MW, Morohashi K and
Simpson ER: Ad4BP/SF-1 regulates cyclic AMP-induced transcription
from the proximal promoter (PII) of the human aromatase P450
(CYP19) gene in the ovary. J Biol Chem. 270:13561–13566. 1995.
View Article : Google Scholar : PubMed/NCBI
|
|
102
|
Panghiyangani R, Soeharso P, Andrijono,
Suryandari DA, Wiweko B, Kurniati M and Pujianto DA: CYP19A1 gene
expression in patients with polycystic ovarian syndrome. J Hum
Reprod Sci. 13:100–103. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
103
|
Shozu M, Sumitani H, Segawa T, Yang HJ,
Murakami K, Kasai T and Inoue M: Overexpression of aromatase P450
in leiomyoma tissue is driven primarily through promoter I.4 of the
aromatase P450 gene (CYP19). J Clin Endocrinol Metab. 87:2540–2548.
2002. View Article : Google Scholar : PubMed/NCBI
|
|
104
|
Jamnongjit M and Hammes SR: Ovarian
steroids: The good, the bad, and the signals that raise them. Cell
Cycle. 5:1178–1183. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
105
|
Yang F, Ruan YC, Yang YJ, Wang K, Liang
SS, Han YB, Teng XM and Yang JZ: Follicular hyperandrogenism
downregulates aromatase in luteinized granulosa cells in polycystic
ovary syndrome women. Reproduction. 150:289–296. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
106
|
Dewailly D, Robin G, Peigne M, Decanter C,
Pigny P and Catteau-Jonard S: Interactions between androgens, FSH,
anti-Müllerian hormone and estradiol during folliculogenesis in the
human normal and polycystic ovary. Hum Reprod Update. 22:709–724.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
107
|
Che Q, Liu M, Zhang D, Lu Y, Xu J, Lu X,
Cao X, Liu Y, Dong X and Liu S: Long noncoding RNA HUPCOS promotes
follicular fluid androgen excess in PCOS patients via aromatase
inhibition. J Clin Endocrinol Metab. 105:dgaa0602020. View Article : Google Scholar : PubMed/NCBI
|
|
108
|
Gu Y, Xu W, Zhuang B and Fu W: Role of
A-kinase anchoring protein 95 in the regulation of cytochrome P450
family 19 subfamily A member 1 (CYP19A1) in human ovarian granulosa
cells. Reprod Fertil Dev. 30:1128–1136. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
109
|
Ma X, Hayes E, Prizant H, Srivastava RK,
Hammes SR and Sen A: Leptin-induced CART (cocaine- and
amphetamine-regulated transcript) is a novel intraovarian mediator
of obesity-related infertility in females. Endocrinology.
157:1248–1257. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
110
|
Turkistani A and Marsh S: Pharmacogenomics
of third-generation aromatase inhibitors. Expert Opin Pharmacother.
13:1299–1307. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
111
|
Kharb R, Haider K, Neha K and Yar MS:
Aromatase inhibitors: Role in postmenopausal breast cancer. Arch
Pharm (Weinheim). 353:e20000812020. View Article : Google Scholar
|
|
112
|
Usluogullari B, Duvan C and Usluogullari
C: Use of aromatase inhibitors in practice of gynecology. J Ovarian
Res. 8:42015. View Article : Google Scholar : PubMed/NCBI
|
|
113
|
Ammazzalorso A, Gallorini M, Fantacuzzi M,
Gambacorta N, De Filippis B, Giampietro L, Maccallini C, Nicolotti
O, Cataldi A and Amoroso R: Design, synthesis and biological
evaluation of imidazole and triazole-based carbamates as novel
aromatase inhibitors. Eur J Med Chem. 211:1131152021. View Article : Google Scholar
|
|
114
|
Haltia UM, Pihlajoki M, Andersson N,
Mäkinen L, Tapper J, Cervera A, Horlings HM, Turpeinen U, Anttonen
M, Bützow R, et al: Functional profiling of FSH and estradiol in
ovarian granulosa cell tumors. J Endocr Soc. 4:bvaa0342020.
View Article : Google Scholar : PubMed/NCBI
|
|
115
|
Ghosh D, Lo J and Egbuta C: Recent
progress in the discovery of next generation inhibitors of
aromatase from the structure-function perspective. J Med Chem.
59:5131–5148. 2016. View Article : Google Scholar :
|
|
116
|
Steinkampf MP, Mendelson CR and Simpson
ER: Effects of epidermal growth factor and insulin-like growth
factor I on the levels of mRNA encoding aromatase cytochrome P-450
of human ovarian granulosa cells. Mol Cell Endocrinol. 59:93–99.
1988. View Article : Google Scholar : PubMed/NCBI
|