Introduction
Epilepsy is a prevalent brain disease characterized
by long-term vulnerability to seizures and emotional and cognitive
impairment, affecting approximately 50 million individuals
worldwide (1). Approximately 40%
of epileptic patients have structural or metabolic causes due to
different brain injuries (2) and
develop depression and anxiety (3). Previous research has indicated that
the incidence of epilepsy in developing nations is significantly
higher than that in developed countries (4). Recently, the average annual cost of
each epilepsy patient was reported to be approximately 5,253 USD in
China, accounting for more than half of their annual income
(5). Additionally, it has been
scientifically confirmed that factors, such as chronic stress,
brain abnormalities and interactions between underlying genetic
factors and the environment may lead to epilepsy (6,7).
Head injury, infection, trauma, malnutrition, and poor prenatal and
perinatal care are associated with an increased risk of seizures
and epilepsy (4). Additionally,
the abnormal accumulation of damaged mitochondria in the central
nervous system, particularly in the hippocampus, has been reported
to contribute to mitochondrial autophagy defects and may be
involved in the pathology of epilepsy (8). Therefore, the present study focused
on mitochondrial autophagy to investigate the underlying molecular
mechanisms in epilepsy.
MicroRNAs (miRNAs or miRs) are a class of small,
well-preserved noncoding RNAs that monitor numerous biological
processes and cellular functions during the development of the
central nervous system, and the dysfunction of miRNAs contributes
to neurological diseases (9,10). miR-142 is a highly conservative
miR in many invertebrates and vertebrates, and it has been widely
studied in the hematopoietic system, cancers, the inflammatory
response and immune tolerance (11). Junker et al reported that
miR-142 expression was more abundant in active multiple sclerosis
brain lesions than in normal white matter, suggesting that miR-142
may be involved in brain inflammatory and degenerative diseases
(12). However, the role of
miR-142 in epilepsy has not been extensively studied. In the
present study, it was found that miR-142 could target phosphatase
and tensin homolog deleted on chromosome 10 (PTEN)-induced putative
kinase 1 (PINK1). PINK1 is a mitochondrial targeting kinase that
accumulates on damaged mitochondria and protects cells from
stress-induced mitochondrial dysfunction (13). As a neuroprotective kinase, PINK1
is processed by healthy mitochondria and is released to trigger
neuron differentiation (14).
PTEN deficiency can lead to severe brain dysfunction, including
ataxia, seizures and brain enlargement (15). However, the mechanisms of PINK1
in epilepsy remain largely unknown. Considering the above-mentioned
findings, the present study examined the effects of miR-142 and
PINK1 on mitochondrial autophagy in a rat model of epilepsy.
Materials and methods
Animal treatment and grouping
The study was approved and supervised by the Ethics
Committee of Guangzhou Women and Children's Medical Center. The
protocol was also approved by the Institutional Animal Care and Use
Committee of Guangzhou Women and Children's Medical Center.
Significant efforts were made to minimize the number of animals
used and their pain. A total of 96 healthy male Wistar rats (6-8
weeks old, weighing 220±20 g) purchased from Dongzhimen Hospital,
Beijing University of Chinese Medicine [Certificate no. SYXK (Jing)
2015-0001, Beijing, China] were raised under specific pathogen-free
conditions at a constant temperature (20±2°C) and humidity (50-60%)
with free access to sterilized drinking water and standard feed.
The rats were randomly split into the normal group, epilepsy group,
antagomir-negative control (NC) group, antagomir-miR-142 group,
lentivirus vector (LV)-NC group, LV-PINK1 group, antagomir-miR-142
+ siRNA-NC group, and antagomir-miR-142 + siRNA-PINK1 group, with
12 rats in each group. Following the establishment of the model of
epilepsy, the rats were administered a corresponding stereotaxic
injection of antagomir-NC, antagomir-miR-142, LV-NC, LV-PINK1,
antagomir-miR-142 + siRNA-NC and antagomir-miR-142 + siRNA-PINK1
into the brain as described below. The injectants were all from
Shanghai GenePharma Co., Ltd.
Establishment of model of epilepsy
The rats were administered 125 mg/kg of lithium
chloride via intraperitoneal injection, followed by 20 mg/kg of
pilocarpine (both from Sigma-Aldrich; Merck KGaA) 18-20 h later, as
previously described (16). At
30 min prior to the pilocarpine administration, the rats were
administered 1 mg/kg of atropine (Sigma-Aldrich; Merck KGaA) to
alleviate the peripheral cholinergic effect of pilocarpine. Rats in
the normal group were injected with an equal amount of saline.
Following observation for 30 min, according to the Racine (17) grading standard, the model of
epilepsy was successfully established when the seizure activity
reached grade IV or above and lasted for >30 min. To control the
seizure intensity and reduce mortality, 10 mg/kg of diazepam
(Sigma-Aldrich; Merck KGaA) was injected intraperitoneally 90 min
following the onset of the epilepticus status. The
electroencephalogram (EEG, Medelec) data of the rats were recorded
before epilepsy and at 3 h after epilepsy; the data of normal rats
were also collected.
Stereotaxic injection into the rat
brain
Epileptic rats were anesthetized with 1%
pentobarbital sodium (45 mg/kg) through intraperitoneal injection.
The incisor teeth of the rats were fixed using a maxillary fixator
of a stereotaxic instrument (Shanghai Medical Instruments Co.,
Ltd.). Subsequently, 1 ear stick was pushed into the external
auditory canal of the rats to locate the rat's head in the middle
of the 2 slip paths, and the ear stick on the other side was pushed
in to fix the head. The scalp of the rats was shaved to determine
the bregma of the stereotaxic localization of the brain. The right
lateral ventricle was determined as the injection site (AP, -1 mm;
LL, 1.5 mm; V, -4.5 mm), and 10 µl antagomir-NC (4 nmol),
antagomir-miR-142 (4 nmol) (18), LV-NC (6×104 TU),
LV-PINK1 (6×104 TU), antagomir-miR-142 + siRNA-NC (0.5
µg) or antagomir-miR-142 + siRNA-PINK1 (0.5 µg) were
injected into the brains of the rats at 0.25 µl/min using a
Hamilton syringe (Hamilton Company). After the injection, the
needle was pulled out; the skull was sealed with bone wax and the
skin was sutured and disinfected. The rats were allowed to recover
for 5 days after the injection. During this period, the health
status of the rats was detected every day, and no deaths occurred.
Following recovery, samples were collected for use in subsequent
experiments.
Tissue collection
Following anesthesia (45 mg/kg, 1% pentobarbital
sodium through intraperitoneal injection), the rats were rapidly
decapitated, and the hippocampal tissues were rapidly separated
from the ice. In each group, 3 hippocampal tissues were made into a
homogenate and stored at −80°C until analysis, and another 3
hippocampal tissues (1 mm3) were used for transmission
electron microscope (TEM) observation, and the remaining 6
hippocampal tissues were fixed with 10% formalin for 24 h. The
hippocampal tissues were dehydrated, sliced into 4-µM-thick
sections, and were then embedded in paraffin for tissue
staining.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
Total RNA was extracted from the tissue samples
using the TRIzol kit (Invitrogen; Thermo Fisher Scientific, Inc.)
and was then reverse transcribed into cDNA using the Rever Tra
Ace® aPCR RT Master Mix (Toyobo Co., Ltd.). The designed
primers were synthesized by Takara Holdings Inc. (Table I). Reverse transcription was
performed in a 10-µl volume in a 37°C water bath for 15 min,
followed by incubation at 50°C for 5 min and then incubation at
98°C for 5 min for reverse transcriptase inactivation according to
the manufacturer's instructions. The reaction solution was
subjected to quantitative fluorescence following the instructions
of the SYBR® Premix Ex Taq™ II kit in the ABI
PRISM® 7300 system. The 50-µl reaction system
comprised 25 µl of SYBR® Premix Ex Taq™ II (2X),
2 µl of PCR forward primers, 2 µl of PCR reverse
primers, l µl of ROX Reference Dye (50 X), 4 µl of
DNA template and 16 µl of ddH2O. The reaction
conditions were as follows: Pre-denaturation at 95°C for 5 min, and
40 cycles of denaturation at 95°C for 40 sec, annealing at 60°C for
30 sec, and extension at 72°C for 1 min. U6 served as the internal
reference for miR-142 and glyceraldehyde-3-phosphate dehydrogenase
(GAPDH) for PINK1. The relative transcription levels of target
genes were calculated using the 2−ΔΔCt relative
quantitative method (19).
ΔΔCt=ΔCt epilepsy group-ΔCt control group, ΔCt=Ct (target gene)-Ct
(internal reference).
 | Table ISequences of primers used for
RT-qPCR. |
Table I
Sequences of primers used for
RT-qPCR.
| Gene | Sequence
(5′-3′) |
|---|
| miR-142 | F:
5′-GGCGCTAGCAAGAGAGGGG-3′ |
| R:
5′-GGCGGATCCTCGTCCTTGACGCTG-3′ |
| U6 | F:
5′-CTCGCTTCGGCAGCACA-3′ |
| R:
5′-AACGCTTCACGAATTTGCGT-3′ |
| PINK1 | F:
5′-ATCAGTAGCATCTAGCATAC-3′ |
| R:
5′-GATCACTGATCAGATCTATCC-3′ |
| GAPDH | F:
5′-TCTCCCTCACAATTTCCATCCC-3′ |
| R:
5′-TTTTTGTGGGTGCAGCGAAC-3′ |
Western blot analysis
Total proteins were extracted from the hippocampal
tissues according to the instructions provided with TRIzol reagent
(Invitrogen; Thermo Fisher Scientific, Inc.). The protein
concentration was measured and adjusted with deionized water to
ensure an equal loading amountu sing the bicinchoninic acid assay
kit (Jiancheng Bioengineering Institute). Subsequently, 10%
separating gels and stacking gels were prepared. The samples were
mixed with the loading buffer, boiled at 100°C for 5 min, cooled on
ice, centrifuged at 15,000 × g and 4°C for 5 min, and loaded into
each lane at equal amounts using a pipette for electrophoretic
separation. Subsequently, the proteins (40 µg) on the gel
were transferred to nitrocellulose membranes and blocked with 5%
non-fat dry milk at 4°C overnight. Thereafter, the membranes were
incubated with the primary antibodies (all from Abcam Inc.) PINK1
(ab23707; 1:1,000), translocase of the inner membrane 23 (TIM23;
ab230253; 1:1,000), C-III core 1 (ab2740; 1 µg/ml),
cytochrome c oxidase IV (COX IV; ab33985; 1 µg/ml),
mitochondrial fusion protein 1 (MFN1; ab104274; 1 µg/ml),
B-cell lymphoma-2 (Bcl-2; ab182858; 1:2,000) and Bcl-2-associated X
(Bax; ab32503; 1:1,000) at 4°C overnight. The membranes were then
washed with phosphate-buffered saline (PBS) at room temperature 3
times, each for 5 min. Subsequently, the membranes were incubated
with the horseradish peroxidase-labeled secondary antibody
immunoglobulin G (1:1,000; ab6702, or 1:3,000; ab6708; Abcam) at
37°C for 1 h and then were washed with PBS again at room
temperature 3 times, each for 5 min. The membranes were then rinsed
in enhanced chemiluminescence reagent (Pierce; Thermo Fisher
Scientific, Inc.) at room temperature for 1 min. Following liquid
removal, the membranes were covered with food cling wraps, exposed
in the dark, developed and fixed to observe the results. GAPDH was
used as an internal reference. The protein marker was purchased
from Pierce; Thermo Fisher Scientific, Inc. The western blots were
analyzed using ImageJ2x V2.1.4.7 (Rawak Software, Inc.).
Hematoxylin and eosin (H&E)
staining
Tissue sections were dewaxed in xylene I and II for
5 min and were then separately rinsed in 100, 95, 80 and 75%
ethanol for 2 min, and then in PBS (2×5 min). The sections were
then stained with hematoxylin at room temperature for 5 min, washed
in running water for 3 min, color-separated with 1% hydrochloric
ethanol for 10 sec, and turned to blue with 1% ammonia for 30 sec.
Following a 2-min wash in running water, the sections were rinsed
in 50, 70 and 80% ethanol for 2 min and were then stained with
eosin at room temperature for 2 min. Thereafter, the sections were
washed in running water for 3 min, rinsed in 95% ethanol, absolute
ethanol I and II for 3 min each, followed by rinsing in xylene I
and II for 5 min each. Finally, the sections were sealed with
neutral gum and observed under a BX-51 optical microscope (Olympus
Optical Co., Ltd.) by two pathologists in a blinded manner.
Nissl staining
Paraffin-embedded sections were dewaxed with 0.5%
cresyl violet at room temperature, hydrated and stained with Nissl
dye (G1434; Beijing Solarbio Science & Technology Co., Ltd.) at
room temperature for 10 min. The sections were dehydrated and
sealed. Finally, each section was observed by 2 pathologists under
a BX-51 optical microscope (Olympus Optical Co., Ltd.) and the
Nissl-positive cells were counted.
Terminal deoxynucleotidyl transferase
(TdT)-mediated dUTP nick end labeling (TUNEL) assay
After the paraffin-embedded sections were dewaxed
and dehydrated, apoptosis was detected using the TUNEL detection
kit (Beyotime Institute of Biotechnology). The sections were
incubated with protease K (diluted with 10 mM Tris-HCl) at 37°C for
25 min. Following 3 PBS washes and drying, the sections were added
to mounting fluid and incubated at room temperature for 20 min. The
sections were then washed again with PBS in triplicate and
incubated with 50 µl of biotin labeling solution at 37°C for
60 min. Thereafter, the sections were washed 3 times with PBS and
2,4-diaminobutyric acid reagent was added to observe coloration
under the BX-51 optical microscope (Olympus Optical Co., Ltd.).
Color development was terminated by adding water, and the nuclei
were stained in hematoxylin at room temperature for 1 min,
dehydrated and cleared, and were then sealed with neutral gum. Each
section was observed by two pathologists under a BX-51 optical
microscope (Olympus Optical Co., Ltd.) and the number of
TUNEL-positive cells stained brown yellow in 5 random fields was
counted.
Hoechst 33258 staining
The paraffin-embedded sections were dewaxed with
xylene and gradient ethanol, bathed at 97°C in citrate buffer for
15 min, and washed twice with PBS, 3 min each time. The sections
were then stained with Hoechst 33258 dye (Invitrogen; Thermo Fisher
Scientific, Inc.) at room temperature for 5 min, and were then
washed twice with PBS (3 min/time) following dye removal. A drop of
anti-quenching sealing liquid was added to the sections, which were
then covered with a clean cover slip while avoiding bubbles as much
as possible. Finally, the sections were photographed (excitation
wavelength, 350 nm; emission wavelength, 460 nm) under a BX-60
fluorescence microscope (Olympus Corporation).
Fluoro-Jade C (FJC) staining
Hippocampal tissue sections were immersed in 1%
NaOH-80% ethanol mixture for 5 min, transferred to 70% ethanol for
2 min, and then immersed in distilled water for 2 min, 0.06%
potassium permanganate solution for 10 min, followed by washing in
distilled water for 2 min. Additionally, the sections were evenly
dripped with 0.0001% FJC dye (containing 0.1% acetic acid; Chemicon
International, Inc.) at room temperature for 10 min and were then
rinsed 3 times in distilled water for 1 min each time. The slices
were dried, cleared with xylene for 1 min, and sealed with neutral
gum. The blue filter (excitation wavelength 450-490 nm) was used to
observe and collect images under the BX-60 fluorescence microscope
(Olympus Corporation).
Thiobarbituric acid (TBA) method and
water-soluble tetrazolium salt-8 (WST-8) method
The hippocampus tissue homogenate was centrifuged at
2,000 × g and 4°C for 15 min with the supernatant absorbed. The
malondialdehyde (MDA) content and superoxide dismutase (SOD)
activity were determined using the TBA method and WAT-8 method,
respectively, according to the manufacturer's instructions
(Beyotime Institue of Biotechnology).
TEM observation
The hippocampal tissue sections at 1 mm3
were fixed in 40 g/l of glutaraldehyde for 1 h and washed 3 times
with 0.1 mol/l of phosphate buffer (pH 7.4) for 5 min each time.
The tissue sections were fixed with 1% osmium acid for 2-3 h and
were washed 3 times with 0.1 mol/l of phosphate buffer (pH 7.4) for
5 min each time. The tissue sections were dehydrated by gradient
ethanol, immersed in a mixture of acetone and an equal amount of
Epon812 for 3 h, embedded in Epon812, and polymerized at 60°C for
48 h. Thereafter, the sections were stained with 3% uranium acetate
and lead citrate (all from Sangon Biotech) at room temperature for
15 min, respectively, and were observed and photographed under a
TEM (JEM-1200 EX; JEOL, Ltd.).
Detection of mitochondrial transmembrane
potential (MTP) by JC-10 staining
The MTP in hippocampal homogenates was detected
using JC-10 fluorescent probes (Beijing Solarbio Science &
Technology Co., Ltd.). The hippocampal homogenates were incubated
with 500 µl of JC-10 dye solution at 37°C for 20 min.
Following incubation, the sections were centrifuged at 600 × g and
4°C for 4 min and the supernatants were discarded, and the sections
were then washed twice with JC-10 staining buffer. The MTP was
measured using a flow cytometer (Guava, Easycyte™ 8; EMD Millipore)
and a Guava PCA flow cytometry system (EMD Millipore) following
suspension.
MitoSOX-based flow cytometry to detect
mitochondrial reactive oxygen species (ROS)
The hippocampal homogenates were incubated in 0.01
µm MitoSOX dye (Invitrogen; Thermo Fisher Scientific, Inc.)
diluted in PBS at a ratio of 1:1 for 30 min at 37°C. The
supernatant was then removed, and the cells were detached using
ethylene diamine tetraacetic acid-free trypsin, followed by
centrifugation at 1,000 × g and 4°C for 5 min. Finally, the cells
were washed 3 times with 200 ml of PBS and the ROS content was
detected by flow cytometry (Guava, Easycyte™ 8; EMD Millipore) with
the Guava PCA flow cytometry system (EMD Millipore).
Dual luciferase reporter gene assay
Bioinformatics software TargetScan (http://www.targetscan.org) was used to predict the
targeting association and binding sites between miR-142 and PINK1.
The PINK1 3′UTR promoter sequence containing the binding site of
miR-142 was synthesized and inserted into the pMIR-REPORT™
Luciferase vector plasmid (Ambion, Inc.) to construct the PINK1
3′UTR wild-type (PINK1-WT) plasmid and PINK1 3′UTR mutant type
(PINK1-MUT) plasmid. Plasmid extraction was carried out according
to the manufacturer's instructions (Promega Corporation). PINK1-WT
and PINK1-MUT were mixed with 100 nM mimic NC and mimic miR-142
(GenePharma), respectively, and were then co-transfected into 293T
cells [American Type Culture Collection (ATCC)] using
Lipofectamine™ 2000 (Invitrogen; Thermo Fisher Scientific, Inc.).
After 48 h, the luciferase activity was detected using a luciferase
detection kit (BioVision, Inc.) and a Glomax 20/20 luminometer
fluorescence detector (Promega Corporation). The ratio of firefly
luciferase activity to Renilla luciferase activity was
calculated as the relative activity. The experiment was repeated 3
times.
Statistical analysis
Statistical analysis was conducted using SPSS 21.0
(IBM Corp.). All the data were examined using the
Kolmogorov-Smirnov test. Measurement data are expressed as the
means ± standard deviation. A t-test was used for comparisons
between 2 groups, and one-way analysis of variance (ANOVA) or
two-way ANOVA was used for comparisons among multiple groups.
Tukey's multiple comparisons test was used as a post hoc test.
P-values were obtained by a two-tailed test, and P<0.05 was
considered to indicated a statistically significant difference.
Results
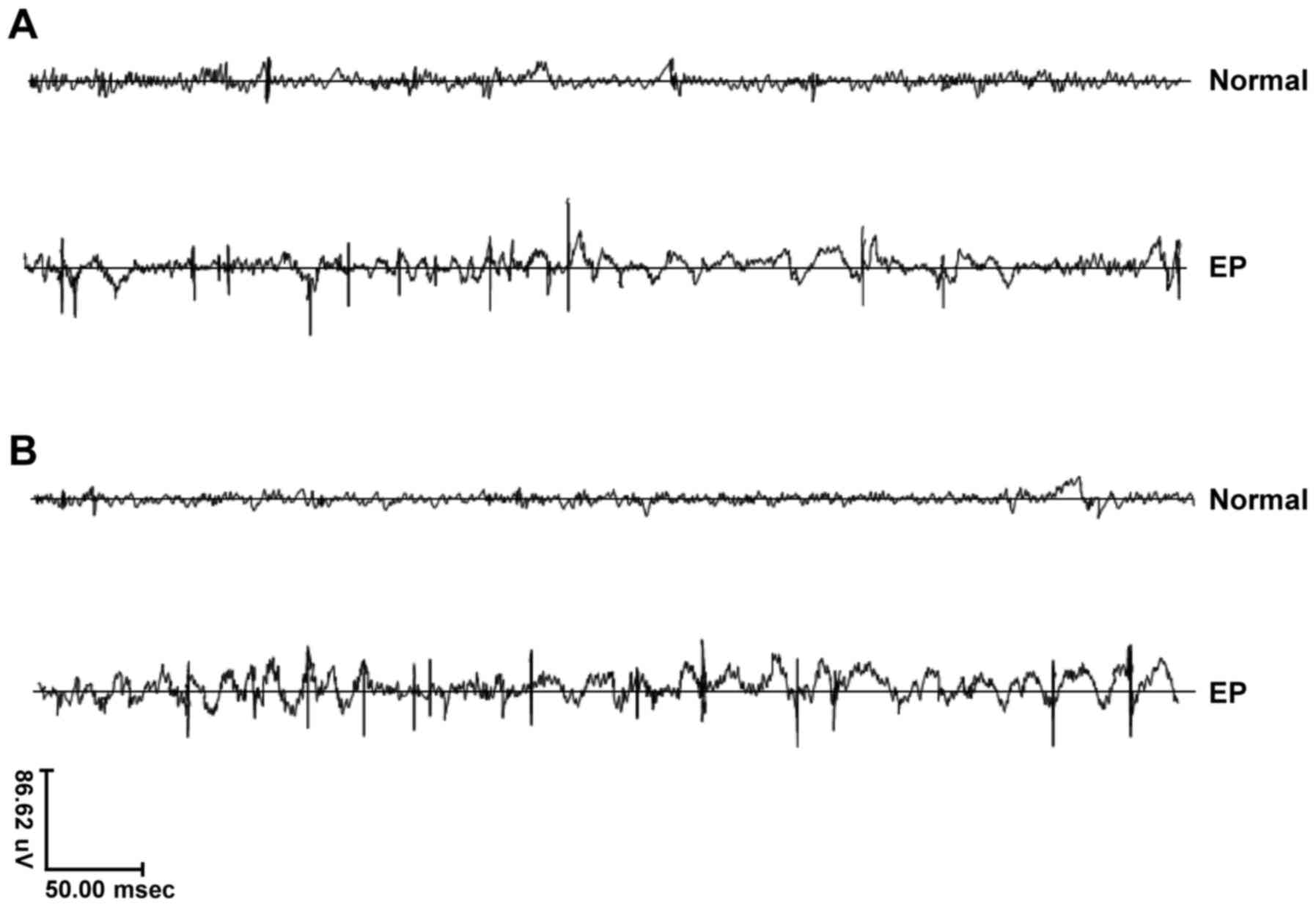
Epileptic rats exhibit obvious
epileptiform discharges, spikes and sharp waves
The normal rats exhibited normal behavior and no
epileptic seizures. According to the Racine grading standard,
epileptic seizures in epileptic rats reached grade IV-V. EEG
mapping results revealed that the normal rats had no cluster spike
discharges, while the epileptic rats exhibited high cluster
amplitudes and high-frequency multi-spike discharges (Fig. 1). These manifestations suggested
the epilepsy model was successfully established.
miR-142 inhibition reduces ROS generation
and apoptosis in hippocampal neurons in epileptic rats
Following the successful establishment of the model
of epilepsy, miR-142 expression was measured in rats subjected to
different treatments. miR-142 expression was upregulated in the
epileptic rats compared with that in normal rats, and was decreased
in rats treated with antagomir-miR-142 compared that in rats
treated with antagomir-NC (both P<0.01; Fig. 2A). H&E staining (Fig. 2B) and Nissl staining (Fig. 2C) revealed that the hippocampal
neurons in the normal rats were in band distribution. A number of
dense vertebral cells were observed in the CA1 and CA3 regions,
which were well arranged, complete in shape, clear in the edge and
nucleolus, transparent in the cytoplasm, round or oval in the
nucleus, and uniform in chromatin distribution, with a normal shape
and abundant Nissl bodies in the cytoplasm. Additionally, the
hippocampus of the epileptic rats exhibited obvious neuronal
degeneration, cell structure disintegration, edema and
proliferation in astrocytes. In the CA1 and CA3 regions, the
structure of nerve cells was incomplete, with swelling and
ruptures, a blurred outline and unclear boundaries, disordered
arrangement, enlarged cell spacing, chromatin clumping in the
edges, cytoplasmic edema, and enlargement of cell bodies and
decreased cytoplasmic Nissl bodies. Following antagomir-miR-142
treatment, most of the pyramidal cells in the CA1 and CA3 regions
of the hippocampus of rats had clear boundaries and normal
morphology, with broken structures, blurred contours and unclear
boundaries in a few neurons.
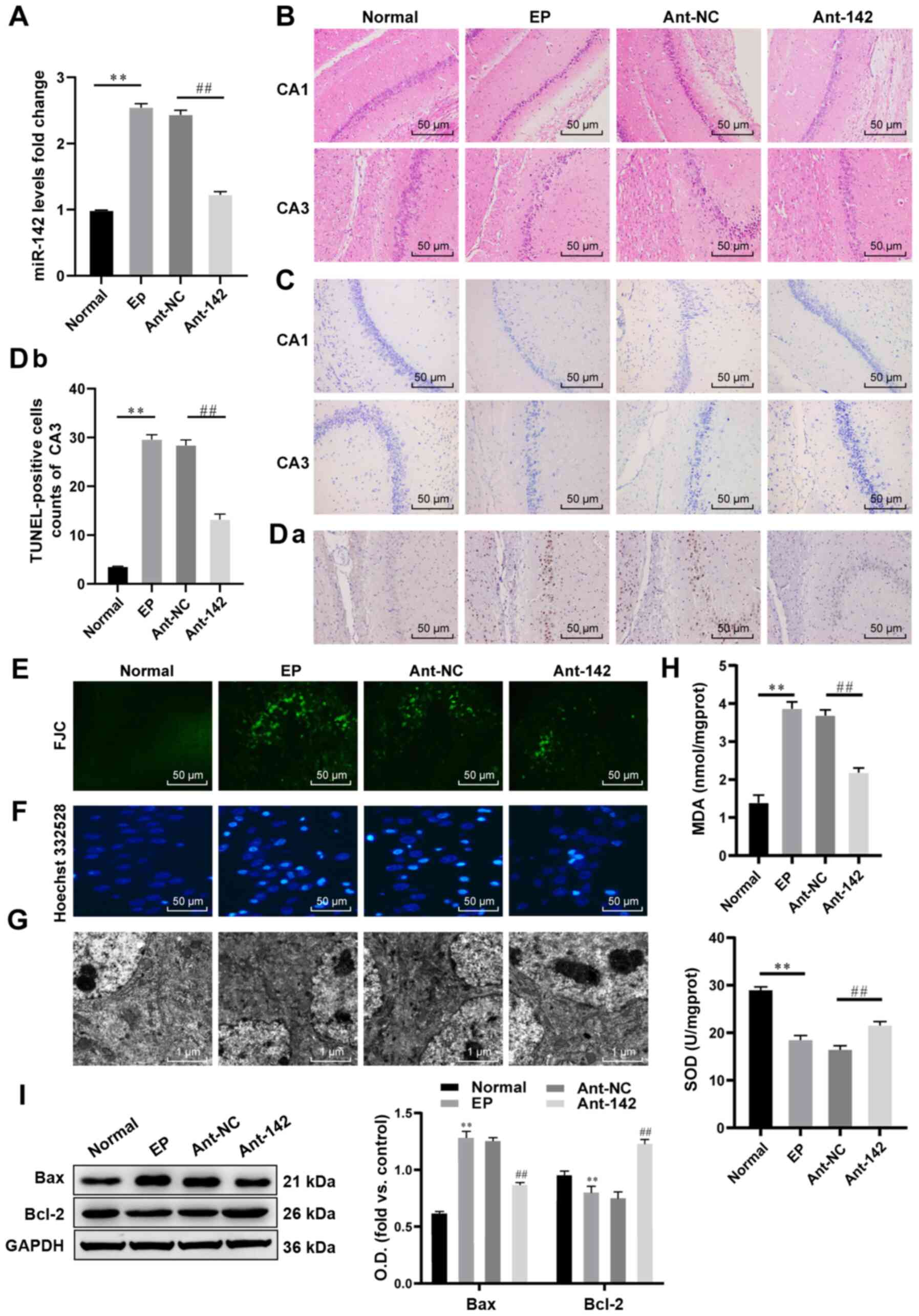 | Figure 2miR-142 inhibition reduces ROS and
apoptosis in hippocampal neurons in epileptic rats. (A) Relative
miR-142 expression in normal rats and epileptic rats measured by
RT-qPCR, n=3. (B and C) Representative images of the
histopathological sections of the hippocampus in epileptic rats
detected by H&E staining and Nissl staining, n=6. (D-F)
Representative images of apoptosis and degeneration in the
hippocampal CA3 region of epileptic rats detected by the TUNEL
assay, FJC staining and Hoechst 332528 staining, n=6. (G)
Representative images of the neuron structure in the hippocampus of
epileptic rats observed under TEM, n=3. (H) Relative MDA and SOD
contents in normal rats and epileptic rats measured by the TBA and
WST-8 methods, n=3. (I) Protein levels of Bcl-2 and Bax in the
hippocampus of normal rats and epileptic rats measured by western
blot analysis, n=3. The data were analyzed using one-way ANOVA and
Tukey's multiple comparisons test as a post hoc test.
**P<0.01, compared with the normal group;
##P<0.01, compared with the antagomir-NC group.
miR-142, microRNA-142; ROS, reactive oxygen species; RT-qPCR,
reverse transcription-quantitative polymerase chain reaction;
TUNEL, terminal deoxynucleotidyl transferase (TdT)-mediated dUTP
nick end labeling; H&E, hematoxylin and eosin; FJC, Fluoro-Jade
C; MDA, malondialdehyde; SOD, superoxide dismutase; TBA,
thiobarbituric acid; WST-8, water-soluble tetrazolium salt-8;
Bcl-2, B-cell lymphoma-2; Bax, Bcl-2-associated X; ANOVA, analysis
of variance. |
In the CA3 region of the hippocampus of the normal
rats, only a few TUNEL-positive cells (dark brown cells; Fig. 2D) and FJC-positive cells
(Fig. 2E) were observed. In the
epileptic rats, the number of TUNEL-positive cells in the
hippocampal CA3 region significantly increased (P<0.01; Fig. 2D) and the bright yellow-green
fluorescent FJC-positive cells with a neuron-like shape were
observed with clear cell bodies and neurites. Hoechst 332528
staining (Fig. 2F) revealed that
the nuclei of apoptotic cells in epileptic rats exhibited
coagulation and bright blue fluorescence compared with the normal
rats. Following antagomir-miR-142 treatment, the number of
TUNEL-positive apoptotic cells and FJC-positive cells decreased
significantly in the hippocampal CA3 region in the rats, and the
numbers of cells with bright blue fluorescent decreased
(P<0.01).
Under the TEM (Fig.
2G), the structure of hippocampal neurons in normal rats was
clear and complete, the nuclear structure was normal, the nuclear
pore complex was clear, chromatin was evenly distributed, and the
nucleolus was located in the middle. In hippocampal neurons of
epileptic rats, the cell body was shrunken, and there were small
and irregular clots and chromatin margination, rough endoplasmic
reticulum and mitochondrion swelling, the loss of mitochondrial
cristae, organelle reduction, structural destruction, the formation
of cavity and vacuoles of different sizes in the cytoplasm, and
cell membrane rupture. However, these morphologies were reversed by
antagomir-miR-142. Compared with the normal rats, the MDA content
increased, SOD activity decreased significantly (Fig. 2H), the expression of Bax in the
hippocampus increased significantly, and Bcl-2 protein decreased
significantly in the epileptic rats (all P<0.01; Fig. 2I). Following the inhibition of
miR-142 expression, the above-mentioned trends were all
reversed.
miR-142 targets PINK1
These results indicated that miR-142 was involved in
the biological process of epileptic rats in the current research.
Subsequently, the present study wished to identify its downstream
target gene. Using online analysis software, a specific binding
region was found between the PINK1 sequence and miR-142 sequence
(Fig. 3A), indicating that PINK1
is the target gene of miR-142. The dual luciferase report gene
assay was used to further verify the results. Compared with the NC
group, the luciferase activity of the WT-PINK1/miR-142
co-transfection group decreased (P<0.05), while that of
MUT-PINK1 exhibited no significant difference (P>0.05),
indicating that miR-142 could specifically bind to PINK1 (Fig. 3B). Subsequently, miR-142 mimic
was constructed and the overexpression efficiency was detected. The
results revealed that the overexpression of miR-142 significantly
decreased the mRNA and protein levels of PINK1 (Fig. 3C).
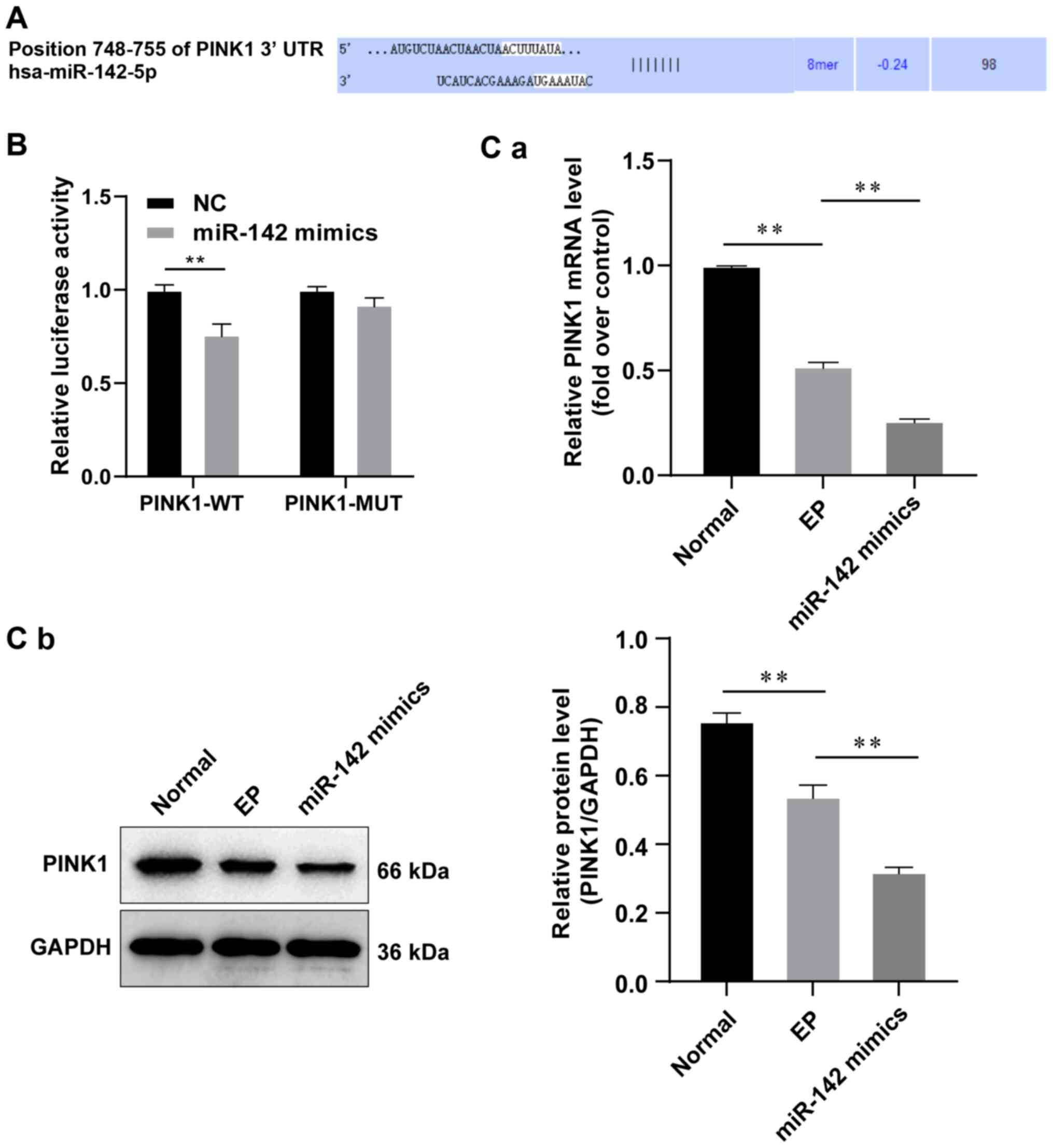 | Figure 3PINK1 is the target gene of miR-142.
(A) Binding sequence of miR-142 and PINK1 was analyzed using
bioinformatics software. (B) The targeting association between
miR-142 and PINK1 was validated by the dual luciferase report gene
assay. The data were analyzed by two-way ANOVA, and the pairwise
comparisons following ANOVA were analyzed by Sidak's multiple
comparisons test. **P<0.01, compared with the NC
group. (C) Relative levels of PINK1 were detected by (a) RT-qPCR
and (b) western blot analysis, n=3. Data were analyzed by one-way
ANOVA, and pairwise comparisons following ANOVA were analyzed by
Tukey's multiple comparisons test. **P<0.01, compared
with the epilepsy group. miR-142, microRNA-142; PINK1, phosphatase
and tensin homolog deleted on chromosome 10 (PTEN)-induced putative
kinase 1; RT-qPCR, reverse transcription-quantitative polymerase
chain reaction; ANOVA, analysis of variance; NC, negative
control. |
Overexpression of PINK1 attenuates
hippocampal damage in epileptic rats
As PINK1 is the target gene of miR-142, it may also
affect the behavior of epileptic rats. Thus, the gain- and
loss-of-functions of PINK1 were investigated to determine its roles
in epilepsy. LV-PINK1 transfection significantly upregulated PINK1
expression in the epileptic rats, while the epileptic rats treated
with anti-miR-142 and PINK1 together exhibited a downregulated
PINK1 expression (both P<0.05; Fig. 4A). Through H&E staining
(Fig. 4B) and Nissl staining
(Fig. 4C), it was observed that
the neurons in the epileptic rats injected with LV-PINK had a
complete morphology, a distinct structure, a normal nuclear
structure, a regular arrangement of vertebral cells, abundant Nissl
bodies in the cytoplasm, and fewer TUNEL-positive cells (Fig. 4D) and FJC-positive cells
(Fig. 4E) (all P<0.05).
Hoechst 33258 nuclear staining (Fig.
4F) revealed that most of the cells displayed uniform light
blue fluorescence and only a few apoptotic cells were observed. The
inhibition of miR-142 and PINK1 together revealed that some neurons
in the hippocampus were damaged more severely and were arranged
disorderly and sparsely, with reduced vertebral cells and Nissl
bodies, and increased TUNEL-positive cells, FJC-positive cells and
bright blue cells. Briefly, the overexpression of PINK1 attenuated
hippocampal damage in epileptic rats and PINK1 downregulation
suppressed the protective effects of antagomir-miR-142.
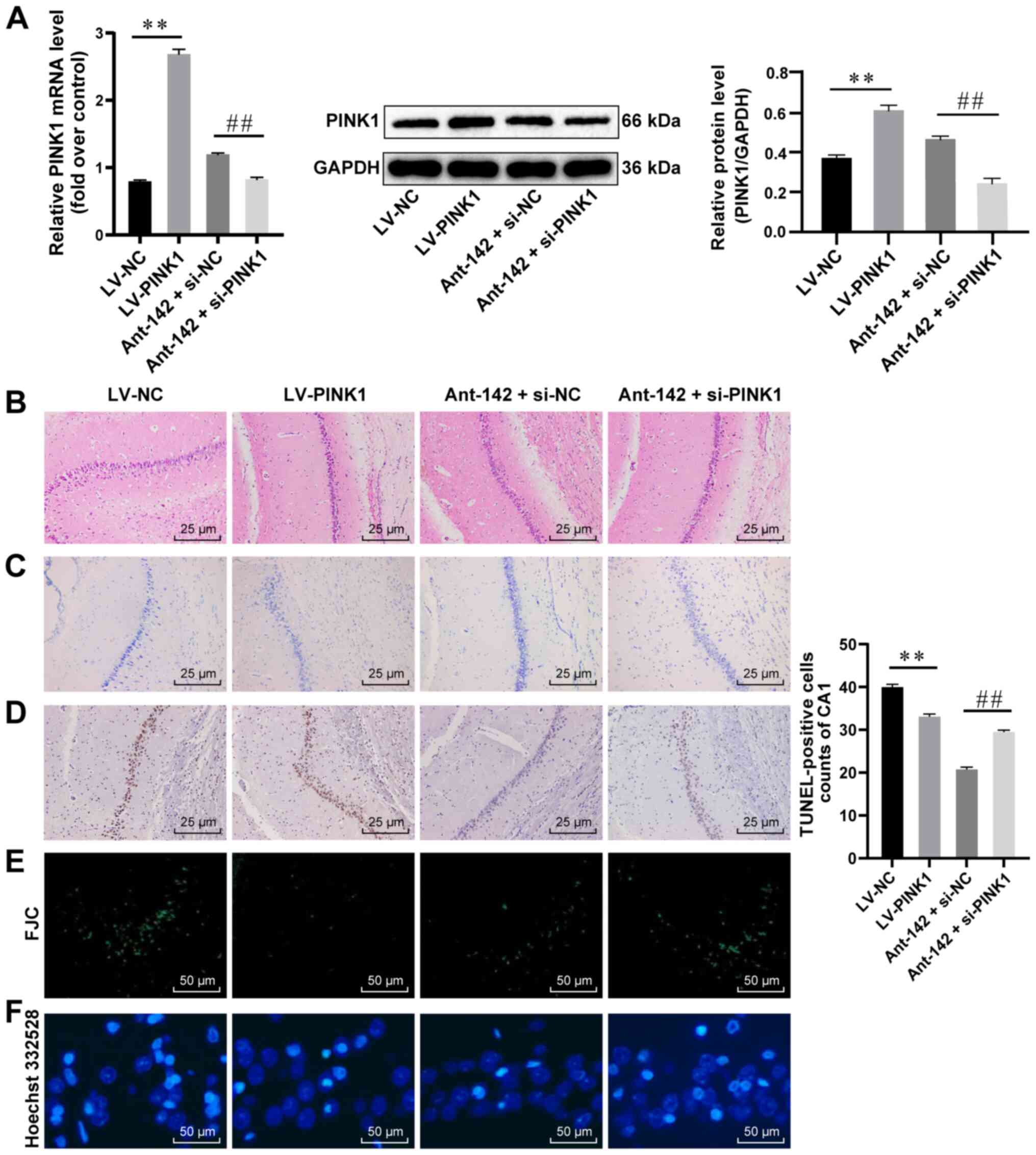 | Figure 4PINK1 overexpression is protective
for hippocampal neurons in epileptic rats. (A) Relative mRNA and
protein levels of PINK1 in the hippocampus of epileptic rats
detected by RT-qPCR and western blot analysis, n=3. (B-F)
Representative images of H&E staining, Nissl staining, TUNEL
assay, EJC staining and Hoechst 33258 nuclear staining, n=6. Data
were analyzed by one-way ANOVA, and the pairwise comparisons
following ANOVA were analyzed by Tukey's multiple comparisons test.
**P<0.01, compared with the LV-NC group;
##P<0.01, compared with the antagomir-miR-142 +
siRNA-NC group. miR-142, microRNA-142; PINK1, phosphatase and
tensin homolog deleted on chromosome 10 (PTEN)-induced putative
kinase 1; RT-qPCR, reverse transcription quantitative polymerase
chain reaction; H&E, hematoxylin and eosin; ANOVA, analysis of
variance; LV, lentivirus vector; NC, negative control. |
miR-142 inhibition promotes mitochondrial
autophagy in the hippocampal tissue of epileptic rats by targeting
PINK1
The above-mentioned findings revealed that miR-142
downregulated PINK1 expression, and it has been shown that PINK1
can promote mitochondrial autophagy (20). Therefore, the present study
further investigated the regulatory effects of miR-142 on
PINK1-mediated mitochondrial autophagy in epileptic rats. Compared
with the normal rats, the number of mitochondria in the epileptic
rats decreased and many damaged mitochondria were encapsulated by
the bilayer membrane. The mitochondria were swollen, and the
mitochondrial ridge was disordered, broken or disappeared with
obvious vacuolar degeneration. Mitochondrial morphology was
restored without vacuole mitochondria following the inhibition of
miR-142 expression or the overexpression of PINK1 in the epileptic
rats. However, mitochondrial morphology and structure were
destroyed with severe vacuolar degeneration following interference
with miR-142 and PINK1 (Fig.
5A).
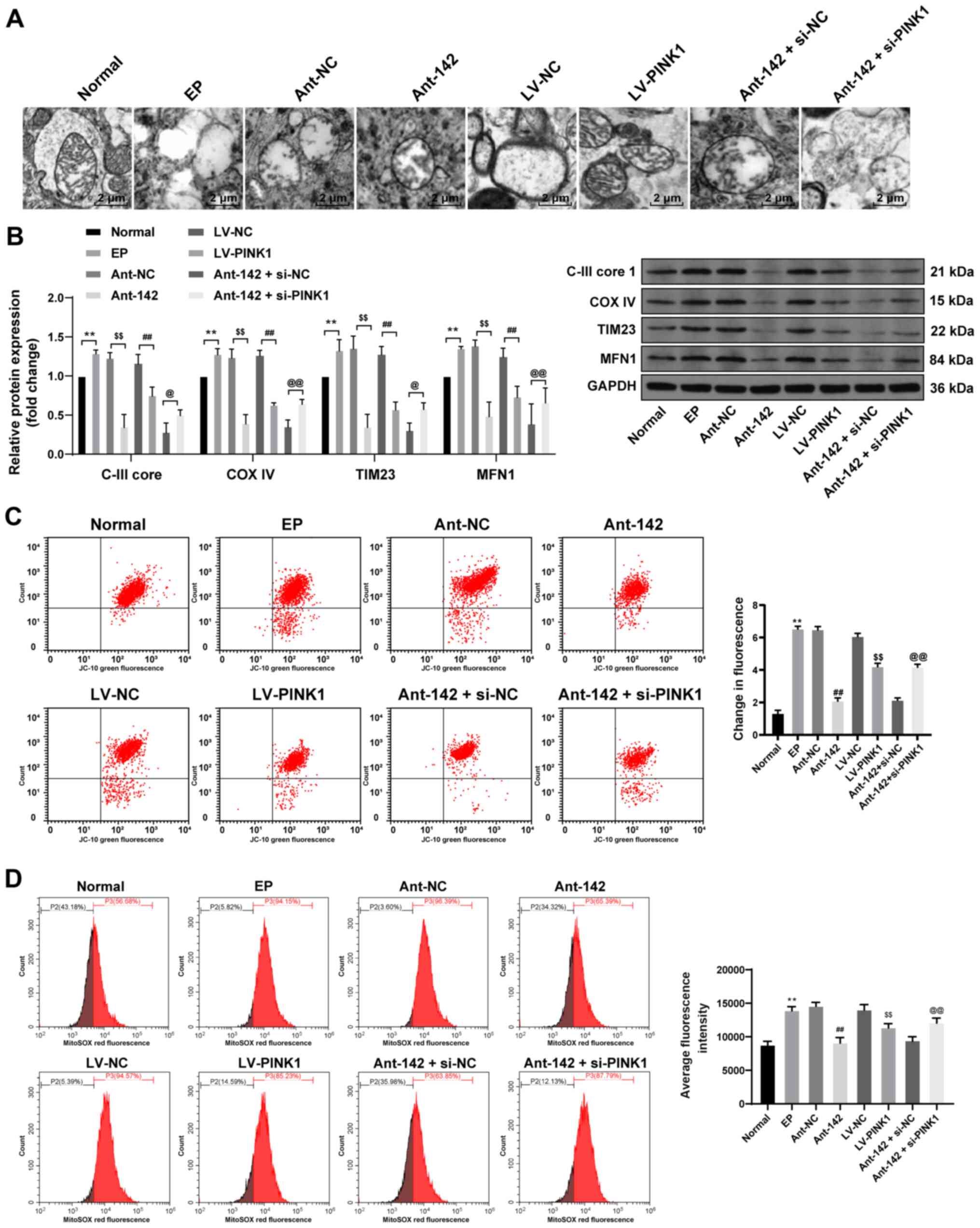 | Figure 5miR-142 inhibition promotes
mitochondrial autophagy in epileptic rats by upregulating PINK1.
(A) Representative images of the mitochondrial structure in the
hippocampus of epileptic rats were observed by TEM, n=3. (B)
Protein levels of C-III core 1, COX IV, TIM23 and MFN1 in the
hippocampus of epileptic rats were measured by western blot
analysis, n=3; (C and D) Relative MTP and ROS generation in the
hippocampus of epileptic rats were measured by JC-10 staining and
MitoSOX-based flow cytometry, n=3. Data were analyzed by one-way
ANOVA, and pairwise comparisons following ANOVA were analyzed by
Tukey's multiple comparisons test. **P<0.01, compared
with the normal group; ##P<0.01, compared with the
LV-NC group; $$P<0.01, compared with the antagomir-NC
group; @P<0.05, @@P<0.01, compared with
the antagomir-miR-142 + siRNA-NC group. miR-142, microRNA-142;
PINK1, phosphatase and tensin homolog deleted on chromosome 10
(PTEN)-induced putative kinase 1; TEM, transmission electron
microscope; COX IV, cytochrome C oxidase IV; TIM23, translocase of
the inner membrane 23; MFN1, mitochondrial fusion protein 1; MTP,
mitochondrial transmembrane potential; ROS, reactive oxygen
species; ANOVA, analysis of variance; LV, lentivirus vector; NC,
negative control. |
The protein levels of C-III core 1, COX IV, TIM23
and MFN1 were upregulated in the epileptic rats; however, opposite
trends were observed following the inhibition of miR-142 expression
or the overexpression of PINK1. Additionally, intervention with
miR-142 and PINK1 further increased the levels of the
above-mentioned proteins in the epileptic rats (all P<0.01;
Fig. 5B).
In the epileptic rats, MTP decreased significantly
and the number of green fluorescent cells increased, indicating
that increased numbers of cells were apoptotic, while orange
fluorescence exhibited normal living cells, and ROS generation
increased significantly. The inhibition of miR-142 expression or
the overexpression of PINK1 maintained MTP and reduced ROS
generation in the epileptic rats, effects that were reversed by
combined intervention with miR-142 and PINK1 (all P<0.01;
Fig. 5C and D).
Discussion
Despite several new improvements in this field over
the past years, ~30% of all epilepsy cases remain refractory to
medication, and seizures persist even after receiving optimum drug
treatment (21). It has been
revealed that a series of pathophysiological changes occur in the
CA1 and CA3 in regions in the hippocampus during epilepsy (22-24). Previous evidence has supported
the important roles of miRNAs in hippocampal neuronal death, brain
injuries, inflammation and the immune response in the pathogenesis
of epilepsy (25). Therefore,
the present study focused on the pathophysiological processes in
the CA1 and CA3 regions in the hippocampus to explore the effects
of miR-142 on epileptic rats. Collectively, it was concluded that
miR-142 knockdown stimulated mitochondrial autophagy and alleviated
hippocampal damage in epileptic rats by targeting PINK1.
First, miR-142 expression was upregulated in
epileptic rats, and rats treated with antagomir-miR-142 exhibited a
normal structure of hippocampal neurons, the alleviation of
hippocampal damage and mitochondrial dysfunction, a decreased MDA
content, an increased SOD activity, a downregulated Bax expression
and an upregulated Bal-2 expression. A recent study demonstrated
that changes in miR-142 expression in the brain can lead to an
increased risk of Alzheimer's disease, and elevated levels of
miR-142-3p are a risk factor for Alzheimer's disease (26). MDA, a key marker of oxidative
stress (OS), is expressed in the process of oxidative degradation
of polyunsaturated lipids (27).
ROS-induced lipid peroxidation and MDA production, in turn,
contribute to neurotoxicity and cell death (28). Lengthy seizure activities lead to
increased ROS production, OS) and mitochondrial dysfunction, which
may cause severe brain damage (29). OS and mitochondrial dysfunction
destroy the balance of the intracellular environment, leading to
neuroexcitability and neuronal death in epilepsy (30). OS can damage the mitochondrial
respiratory chain and lead to the excessive production of ROS, the
accumulation of which can inhibit the activity of the mitochondrial
respiratory enzyme complex, leading to cell death in the epileptic
area (16). Additionally, a
reduction in OS protects the hippocampus from abnormal mossy fiber
sprouting, thus lessening recurrent seizures and repressing
hippocampal neuronal loss in a rat model of temporal lobe epilepsy
(31). Furthermore, an increased
MDA content is assocaited with the early age of epilepsy onset
(32). SOD1 levels are
downregulated in the cerebrospinal fluid of patients with epilepsy,
particularly those with refractory epilepsy; thus, the decreased
SOD1 level may predict antiepileptic drug resistance in epileptic
patients (33). Consistent with
the results of the present study, a previous study displayed
significantly enhanced OS in the hippocampus injected with lithium
chloride-pilocarpine, as evidenced by a a markedly increased MDA
content and a decreased SOD activity (16). Partially coinciding with the
current findings, the elevation of SOD activity and the
downregulation of the MDA content has been reported to inhibit OS
in Parkinson's disease, thereby protecting against dopaminergic
neuronal cell apoptosis (34).
Notably, a recent study revealed that, partly in line with the
current observation, namely, the inhibition of miR-142-5p
attenuated oxygen-glucose deprivation and reoxygenation
(OGD/R)-induced cell injury in hippocampal neurons and OS,
indicated by decreased MDA levels and increased SOD levels, and
promoted cell viability and inhibited apoptosis (35).
Furthermore, PINK1 is the target gene of miR-142,
and the overexpression of PINK1 attenuated hippocampal damage in
epileptic rats, and PINK1 downregulation attenuated the protective
effects of antagomir-miR-142. In humans, PINK1 is highly expressed
in the hippocampus, and it antagonizes mitochondrial dysfunction
and protects neurons against OS and neurotoxins (36). Additionally, mice with a
deficiency in PINK1 exhibit increased oxidative damage and
mitochondrial dysfunction in the striatum (37). In primary cultured rat
hippocampal neurons, the overexpression of PINK1 has been shown to
lead to increased mitochondrial, and a diminished mitochondrial
size and proportion in neuronal processes (38). Moreover, the present study found
that the protein levels of C-III core 1, COX IV, TIM23 and MFN1
were upregulated, the MTP was decreased and ROS production
increased significantly in epileptic rats; however, opposite trends
were observed following the inhibition of miR-142 expression or the
overexpression of PINK1, indicating enhanced mitochondrial
autophagy. C-III core 1, COX IV, TIM23 and MFN1 are well-known
indicators of mitochondrial autophagy, among which MFN1 promotes
mitochondrial elongation and activity (39). PINK1 is specifically activated by
MTP depolarization, and the overexpression of PINK1 enhances
mitochondrial fission (40).
PINK1 deficiency results in mitochondrial fragmentation, ROS
production, decreased MTP and mitochondrial cristae density, and
the inhibition of mitochondrial autophagy (41). Importantly, the inhibition of
miR-142-5p can promote PTEN expression and increase autophagy
levels (42).
Taken together, the present study demonstrates that
miR-142 expression is upregulated in epileptic rats, and that
miR-142 inhibition can stimulate mitochondrial autophagy and
alleviate hippocampal damage and ROS in epileptic rats by targeting
PINK1. Additionally, the pathological and physiological changes in
the dentate gyrus area are an important characterization in
epileptic rats; thus, the authors aim to conduct a further in-depth
investigations on the changes in the dentate gyrus area in
epileptic rats in the future. The findings of the present study may
provide a novel understanding of the mechanisms of epilepsy,
enabling the identification of more effective therapeutic
approaches. Further research is warranted to further validate the
current results and apply the results to clinical settings.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
All authors are the guarantors of the integrity of
the entire study. DX, JL and ZZ contributed to the study concept
and design. YL and YZ contributed to the study design and
experiments. CLu, BQ, CLi and LQ were responsible for the
acquisition and analysis of data. DX, JL, ZZ, BQ and CLi
contributed to the manuscript preparation and manuscript editing.
All the authors read and approved the final manuscript.
Ethics approval and consent to
participate
The present study was approved and supervised by the
Ethics Committee of Guangzhou Women and Children's Medical Center.
The protocol was also approved by the Institutional Animal Care and
Use Committee of Guangzhou Women and Children's Medical Center.
Significant efforts were made to minimize the number of animals
used and their pain.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Acknowledgments
Not applicable.
Funding
No funding was received.
References
|
1
|
Vezzani A, French J, Bartfai T and Baram
TZ: The role of inflammation in epilepsy. Nat Rev Neurol. 7:31–40.
2011. View Article : Google Scholar
|
|
2
|
Vezzani A, Fujinami RS, White HS, Preux
PM, Blümcke I, Sander JW and Löscher W: Infections, inflammation
and epilepsy. Acta Neuropathol. 131:211–234. 2016. View Article : Google Scholar :
|
|
3
|
Moon J, Lee ST, Choi J, Jung KH, Yang H,
Khalid A, Kim JM, Park KI, Shin JW, Ban JJ, et al: Unique
behavioral characteristics and microRNA signatures in a drug
resistant epilepsy model. PLoS One. 9:e856172014. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Newton CR and Garcia HH: Epilepsy in poor
regions of the world. Lancet. 380:1193–1201. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Gu L, Liang B, Chen Q, Long J, Xie J, Wu
G, Yan Y, Tan J, Dou W, Chen W, et al: Prevalence of epilepsy in
the People's Republic of China: A systematic review. Epilepsy Res.
105:195–205. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Guerrini R, Marini C and Mantegazza M:
Genetic epilepsy syndromes without structural brain abnormalities:
Clinical features and experimental models. Neurotherapeutics.
11:269–285. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
van Campen JS, Jansen FE, de Graan PN,
Braun KP and Joels M: Early life stress in epilepsy: A seizure
precipitant and risk factor for epileptogenesis. Epilepsy Behav.
38:160–171. 2014. View Article : Google Scholar
|
|
8
|
Wu M, Liu X, Chi X, Zhang L, Xiong W,
Chiang SMV, Zhou D and Li J: Mitophagy in refractory temporal lobe
epilepsy patients with hippocampal sclerosis. Cell Mol Neurobiol.
38:479–486. 2018. View Article : Google Scholar
|
|
9
|
Cao DD, Li L and Chan WY: MicroRNAs: Key
regulators in the central nervous system and their implication in
neurological diseases. Int J Mol Sci. 17:8422016. View Article : Google Scholar :
|
|
10
|
Gorter JA, Iyer A, White I, Colzi A, van
Vliet EA, Sisodiya S and Aronica E: Hippocampal subregion-specific
microRNA expression during epileptogenesis in experimental temporal
lobe epilepsy. Neurobiol Dis. 62:508–520. 2014. View Article : Google Scholar
|
|
11
|
Shrestha A, Mukhametshina RT, Taghizadeh
S, Vásquez-Pacheco E, Cabrera-Fuentes H, Rizvanov A, Mari B,
Carraro G and Bellusci S: MicroRNA-142 is a multifaceted regulator
in organogenesis, homeostasis, and disease. Dev Dyn. 246:285–290.
2017. View Article : Google Scholar
|
|
12
|
Junker A, Krumbholz M, Eisele S, Mohan H,
Augstein F, Bittner R, Lassmann H, Wekerle H, Hohlfeld R and Meinl
E: MicroRNA profiling of multiple sclerosis lesions identifies
modulators of the regulatory protein CD47. Brain. 132:3342–3352.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Lin W and Kang UJ: Structural determinants
of PINK1 topology and dual subcellular distribution. BMC Cell Biol.
11:902010. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Dagda RK, Pien I, Wang R, Zhu J, Wang KZQ,
Callio J, Banerjee TD, Dagda RY and Chu CT: Beyond the
mitochondrion: Cytosolic PINK1 remodels dendrites through protein
kinase A. J Neurochem. 128:864–877. 2014. View Article : Google Scholar :
|
|
15
|
Kwak YD, Wang B, Pan W, Xu H, Jiang X and
Liao FF: Functional interaction of phosphatase and tensin homologue
(PTEN) with the E3 ligase NEDD4-1 during neuronal response to zinc.
J Biol Chem. 285:9847–9857. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Sun J, Gao X, Meng D, Xu Y, Wang X, Gu X,
Guo M, Shao X, Yan H, Jiang C and Zheng Y: Antagomirs targeting
MiroRNA-134 attenuates epilepsy in rats through regulation of
oxidative stress, mitochondrial functions and autophagy. Front
Pharmacol. 8:5242017. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Racine RJ: Modification of seizure
activity by electrical stimulation: Cortical areas.
Electroencephalogr Clin Neurophysiol. 38:1–12. 1975. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Liao Y, Huang Y, Liu X, Luo C, Zou D, Wei
X, Huang Q and Wu Y: MicroRNA-328a regulates water maze performance
in PTZ-kindled rats. Brain Res Bull. 125:205–210. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
20
|
Ma P, Yun J, Deng H and Guo M: Atg1
mediated autophagy suppresses tissue degeneration in pink1/parkin
mutants by promoting mitochondrial fission in drosophila. Mol Biol
Cell. 29:3082–3092. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Jacobs J, Staba R, Asano E, Otsubo H, Wu
JY, Zijlmans M, Mohamed I, Kahane P, Dubeau F, Navarro V and Gotman
J: High-frequency oscillations (HFOs) in clinical epilepsy. Prog
Neurobiol. 98:302–315. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Feng L, Shu Y, Wu Q, Liu T, Long H, Yang
H, Li Y and Xiao B: EphA4 may contribute to microvessel remodeling
in the hippocampal CA1 and CA3 areas in a mouse model of temporal
lobe epilepsy. Mol Med Rep. 15:37–46. 2017. View Article : Google Scholar :
|
|
23
|
Liu Z, Yang C, Meng X, Li Z, Lv C and Cao
P: Neuroprotection of edaravone on the hippocampus of
kainate-induced epilepsy rats through Nrf2/HO-1 pathway. Neurochem
Int. 112:159–165. 2018. View Article : Google Scholar
|
|
24
|
Saffarzadeh F, Eslamizade MJ, Mousavi SM,
Abraki SB, Hadjighassem MR and Gorji A: TRPV1 receptors augment
basal synaptic transmission in CA1 and CA3 pyramidal neurons in
epilepsy. Neuroscience. 314:170–178. 2016. View Article : Google Scholar
|
|
25
|
Henshall DC: MicroRNA and epilepsy:
Profiling, functions and potential clinical applications. Curr Opin
Neurol. 27:199–205. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Ghanbari M, Munshi ST, Ma B, Lendemeijer
B, Bansal S, Adams HH, Wang W, Goth K, Slump DE, van den Hout MCGN,
et al: A functional variant in the miR-142 promoter modulating its
expression and conferring risk of Alzheimer disease. Hum Mutat.
40:2131–2145. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Qiang M, Xu Y, Lu Y, He Y, Han C, Liu Y
and He R: Autofluorescence of MDA-modified proteins as an in vitro
and in vivo probe in oxidative stress analysis. Protein Cell.
5:484–487. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Roomruangwong C, Barbosa DS, de Farias CC,
Matsumoto AK, Baltus TH, Morelli NR, Kanchanatawan B, Duleu S,
Geffard M and Maes M: Natural regulatory IgM-mediated autoimmune
responses directed against malondialdehyde regulate oxidative and
nitrosative pathways and coupled with IgM responses to nitroso
adducts attenuate depressive and physiosomatic symptoms at the end
of term pregnancy. Psychiatry Clin Neurosci. 72:116–130. 2018.
View Article : Google Scholar
|
|
29
|
Dillioglugil MO, Kir HM, Demir C, Ilbay G,
Sahin D, Dillioglugil O, Bambal G, Mekik H and Ates N: Effect of
pentylenetetrazole and sound stimulation induced single and
repeated convulsive seizures on the MDA, GSH and NO levels, and SOD
activities in rat liver and kidney tissues. Brain Res Bull.
83:356–359. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Chang SJ and Yu BC: Mitochondrial matters
of the brain: Mitochondrial dysfunction and oxidative status in
epilepsy. J Bioenerg Biomembr. 42:457–459. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Baluchnejadmojarad T and Roghani M:
Coenzyme q10 ameliorates neurodegeneration, mossy fiber sprouting,
and oxidative stress in intrahippocampal kainate model of temporal
lobe epilepsy in rat. J Mol Neurosci. 49:194–201. 2013. View Article : Google Scholar
|
|
32
|
Lorigados Pedre L, Gallardo JM, Morales
Chacon LM, García AV, Flores-Mendoza M, Neri-Gómez T, Díaz BE,
Cruz-Xenes RM, Fuentes NP and Orozco-Suárez S: Oxidative stress in
patients with drug resistant partial complex seizure. Behav Sci
(Basel). 8. pp. 592018, View Article : Google Scholar
|
|
33
|
Chen D, Lu Y, Yu W, Luo J, Xiao Z, Xiao F
and Wang X: Clinical value of decreased superoxide dismutase 1 in
patients with epilepsy. Seizure. 21:508–511. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Li J, Wang LN, Xiao HL, Li X and Yang JJ:
Effect of electroacupuncture intervention on levels of SOD, GSH,
GSH-Px, MDA, and apoptosis of dopaminergic neurons in substantia
Nigra in rats with Parkinson's disease. Zhen Ci Yan Jiu.
39:185–191. 2014.In Chinese. PubMed/NCBI
|
|
35
|
Wang N, Zhang L, Lu Y, Zhang M, Zhang Z,
Wang K and Lv J: Down-regulation of microRNA-142-5p attenuates
oxygen-glucose deprivation and reoxygenation-induced neuron injury
through up-regulating Nrf2/ARE signaling pathway. Biomed
Pharmacother. 89:1187–1195. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Priyadarshini M, Orosco LA and Panula PJ:
Oxidative stress and regulation of Pink1 in zebrafish (Danio
rerio). PLoS One. 8:e818512013. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Narendra DP, Jin SM, Tanaka A, Suen DF,
Gautier CA, Shen J, Cookson MR and Youle RJ: PINK1 is selectively
stabilized on impaired mitochondria to activate Parkin. PLoS Biol.
8:e10002982010. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Yu W, Sun Y, Guo S and Lu B: The
PINK1/Parkin pathway regulates mitochondrial dynamics and function
in mammalian hippocampal and dopaminergic neurons. Hum Mol Genet.
20:3227–3240. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
White JP, Baltgalvis KA, Puppa MJ, Sato S,
Baynes JW and Carson JA: Muscle oxidative capacity during
IL-6-dependent cancer cachexia. Am J Physiol Regul Integr Comp
Physiol. 300:R201–R211. 2011. View Article : Google Scholar :
|
|
40
|
Kondapalli C, Kazlauskaite A, Zhang N,
Woodroof HI, Campbell DG, Gourlay R, Burchell L, Walden H,
Macartney TJ, Deak M, et al: PINK1 is activated by mitochondrial
membrane potential depolarization and stimulates Parkin E3 ligase
activity by phosphorylating Serine 65. Open Biol. 2:1200802012.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Cherra SJ III, Dagda RK, Tandon A and Chu
CT: Mitochondrial autophagy as a compensatory response to PINK1
deficiency. Autophagy. 5:1213–1214. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Chen J, Cui Y, Zhang N, Yao X, Wang Z and
Yang L: Oleanolic acid attenuated diabetic mesangial cell injury by
activation of autophagy via miRNA-142-5p/PTEN signaling.
Cytotechnology. 71:925–933. 2019. View Article : Google Scholar : PubMed/NCBI
|