Background
To meet the demand for kidney transplants (KTx),
organs are frequently retrieved not only from standard criteria
donors (SCD; a donor who is aged <50 years and suffered brain
death from any number of causes, such as traumatic injuries or a
stroke) but also from expanded criteria donors [ECDs; any donor
aged >60 years or a donors aged ≥50 years with two of the
following: a history of high blood pressure, a creatinine serum
level ≥1.5 mg/dl or death resulting from a stroke]. This comes at
the cost of a higher risk of primary non-function (PNF; the
permanent hyperkalemia, hyperuremia and fluid overload that result
in the need for continuous dialysis after KTx), delayed graft
function (DGF; the need for dialysis session at least once during
the first week after KTx), earlier graft loss and urinary
complications (vesico-ureteral reflux, obstruction of the
vesico-ureteral anastomosis, urine leakage) (1-3).
An increased risk of graft failure and graftectomy can result in
recipient immunization, leading to increasingly difficult donor
selection and higher risk of consecutive graft failure (4,5).
However, there is also evidence that graft survival is unaffected
by the procurement of kidneys from ECDs compared to kidneys
retrieved from SCDs (6,7).
Currently, there is no commercially available
diagnostic tool for the quality assessment of the kidney before KTx
(8). To this end, the majority
of medical centers will typically perform a 0-h biopsy (9,10), but the results only become
available days after surgery and assessment is limited to a limited
part of the kidney. A 0-h biopsy does not provide an insight into
the condition of the entire organ and cannot be used to predict its
function after KTx. The period of kidney storage should be as short
as possible and the decision on KTx should be made during this
period. However, predictive models based on clinical data, such as
the Kidney Donor Profile Index (KDPI) (11,12), the continuous kidney donor risk
index (13) and deceased donor
score (14), are insufficient
for assessing the condition of potentially transplanted kidneys. A
number of preservation and perfusion solutions have been assessed
as possible novel biomarkers of kidney quality and predictors of
short- and long-term outcomes. These include metabolites, enzymes,
complement-related molecules, antibodies, histones and cytoskeletal
elements released by tubular, interstitial, immunological and
epithelial cells (15-17). However, none of these previous
studies have contributed to the identification of biomarkers that
are beneficial for assessing the value of the kidney. This is due
to the lack of randomized controlled trials in this area to confirm
the utility of potential biomarkers. Some of the markers, such as
lactate dehydrogenase (LDH) or neutrophil gelatinase-associated
lipocalin (NGAL), are not tissue specific (18,19). In addition, the methods used to
detect the majority of the candidates are time-consuming, requiring
qualified personnel, expensive equipments and reagents. Therefore,
there remain to be a demand for a rapid, non-invasive diagnostic
tool and reliable predictive model for assessing the
post-transplant function of a sub-optimal donor kidney.
Studies on single protein biomarkers
The analysis of biological samples inevitably
involves the detection of a wide range of proteins. In such
scenarios, two primary pathways for analysis emerge, namely
targeted and untargeted. The former involves targeting specific
groups of proteins, such as immunoglobulins or individual proteins
based on existing knowledge, assessing their relevance to the
phenomena in question. To detect these known individual particles,
targeted analytical methods were used in previous studies. These
included ELISA, western blotting and enzymatic/cytotoxic
assays.
Proteins released after cell
disruption
LDH
LDH was one of the first proteins to be studied in
the renal perfusate. This cytosolic enzyme is responsible for the
conversion of pyruvate to lactate by NADH under normoxic and
hypoxic conditions (20). In
total, five tetrameric isoenzymes consisting of LDHA and LDHB
subunits have been previously described (18,21,22). They differ in their degree of
substrate affinity, electrophoretic mobility, inhibition constants
and tissue distribution (18).
In an experimental rat model, LDHA was previously found to be
mainly present in the proximal renal tubules, whilst LDHB was found
to be mainly expressed in the distal part of the nephron (23).
In relation to KTx, LDH has been documented to serve
as a non-specific marker of cell injury (18). Perfusate LDH activity is not
applicable in the assessment of renal function prior to KTx.
Despite the studies showing a positive association between LDH
concentration and DGF (15,24-26), there are no randomized controlled
trials that have confirmed the viability of LDH measurements for
the assessment of kidney quality. Furthermore, LDH is not a
kidney-specific biomarker (18,27,28). Guzzi et al (15) previously found that besides
glutathione S-transferase (GST), LDH levels were the most commonly
reported parameter in the renal perfusate. Elevated LDH levels have
been reported to associate positively with DGF in cases of donation
after cardiac death (DCDs) (24-26), but not in all cases of donations
after brain death (DBDs) (29,30). According to previous studies,
(15,24-26,31,32) where LDH levels were measured only
in perfusate from DCDs, LDH levels were found to be positively
associated with DGF in all cases. By contrast, in studies
describing the measurement of LDH levels in the perfusate of
kidneys from DBDs, a positive association between DGF and LDH
levels was only found in two (29,30) of the four selected studies found
(15). In addition, this
meta-analysis (15) also showed
that LDH activity should not be considered as a marker of PNF in
patients with DCDs grafts. Further analysis of cases of DBDs and
DCDs demonstrated that although the LDH levels were associated
positively with DGF in the majority of the included studies,
following multivariable analysis only one study showed a weak
association (32). In another
meta-analysis previously conducted by Bhangoo et al
(16), perfusate LDH activity
was found to be a predictor of DGF, PNF and graft failure (GF).
However, this previous analysis lacked a donor-type subgroup
assessment (16). A number of
the included studies revealed a significant association between LDH
activity and DGF, PNF (31,33). Among the different types of
donors, LDH activity was the highest in kidneys from DCD cases,
which reflected the greatest extent of damage prior to KTx among
the studied organs (34).
Perfusate assessment has been used to estimate renal cell injury
after ischemia in a previous study (29), where LDH levels were previously
found to be increased during the ischemic period (29). This phenomenon also associated
positively with damage to renal tubular cells detected by electron
microscopy. The lesions found include mitochondrial swelling and
loss of detail in the inner cristae, swelling and detachment of the
basolateral tubular membrane connective tissue (29).
GSTs
GSTs form one of the most frequently studied groups
of biomarkers obtained from the renal hypothermic machine perfusate
(15,16). In humans, the GST family consists
of three subpopulations according to their localization: Cytosolic,
mitochondrial and microsomal (35). GSTs can be divided into
subclasses, designated by Greek letters that differ in their levels
of expression in humans. α-GST is abundantly expressed in liver and
renal proximal tubular cells, whereas distal tubular cells mainly
express the p-GST isoform (36,37). Although GSTs mainly function as
catabolic agents to catalyze the detoxification of xenobiotics
within glutathione, they have also been reported to mediate cell
signaling. p-GST isoform can inhibit the function of kinases
involved in the MAPK pathway, which regulates cell proliferation
and cell death (38).
GSTs are considered one of the most valuable and
reliable non-invasive markers for pre-transplant DGF prediction,
but their utility in forecasting long-term outcomes, PNF and
discarding kidneys from transplantation remains controversial
(15,16). Significant differences were
observed in the accuracy of the predictive values of different
isoforms for DGF. Hall et al (39) previously found that p-GST
measured at baseline and at the end of hypothermic machine perfused
(HMP) kidneys from deceased donors was associated positively with
DGF. The relative risk ratio of DGF for each log-unit increase at
baseline and post-perfusion p-GST concentration after adjustment
(for black ethnicity, male sex, history of previous kidney
transplant, diabetes as the cause of end-stage renal disease, need
for blood transfusion before KTx, number of human leukocyte antigen
mismatches, body mass index in kg/m2 and duration of
dialysis before transplant) was found to be 1.14 (95% CI,
1.00-1.28) and 1.33 (95% CI, 1.02-1.72), respectively. α-GST did
not associate with DGF. In addition, the concentrations of both
iso-enzymes did not differ between the transplanted and discarded
organs at start and just before end of perfusion, showing that GSTs
levels could not be used as a tool for discarding kidneys from
KTx.
Perfusate GST levels have been previously compared
with HMP parameters to improve the predictive value of GST.
However, the value of this prospect remains controversial, where no
clear association has yet been established. In a previous study by
Moers et al (33), a
correlation between the studied biomarkers [such as LDH, aspartate
aminotransferase, N-Acetyl-/β-glucosaminidase, GSTs, alanine
aminopeptidase (Ala-AP) and heart fatty acid binding proteins
(H-FABP)] and pump parameters (such as cold ischemia time and renal
vascular resistance) was not confirmed. Hall et al (39) also previously found no
correlation between pump parameters (such as renal resistance at 4
h of perfusion and perfusate flow at 4 h) and concentrations of GST
isoforms. By contrast, Qiao et al (40) found that the model combining
end-of-perfusion vascular resistance and GST [area under the curve
(AUC)=0.888; 95% CI, 0.842-0.933] significantly improved DGF
predictability compared to using terminal resistance (AUC=0.756;
95% CI, 0.693-0.818) or GST alone (AUC= 0.729, 95% CI,
0.591-0.806).
Perfusate GST activity has also been investigated
using other potential biomarkers of DGF. Gok et al (41) reported that GST with Ala-AP
activity or GST activity with FABP concentration corresponded
positively to renal injury during ischemia. However, its role in
predicting the occurrence of PNF and in kidney discards for
transplantation remains unclear (33).
Extracellular histones
Ischemic injury during HMP leads to the release of
histones from disrupted cells (42), including extracellular histones
H2A, H2B, H3 and H4. Each nucleated cell contains nucleosomes built
from these four elements in an equimolar manner (43). Following the induction of
pathological conditions, such as ischemia, histones are released
from dying cells into the extracellular space (44,45). Using an experimental porcine
model of warm ischemia and machine perfusion of retrieved kidneys,
extracellular histones were reported to be cytotoxic to endothelial
and renal epithelial cells in a temperature-dependent manner
(46). Furthermore, histone
release is higher during sub normothermic machine perfusion (SNMP)
at 28°C compared with hypothermic machine perfusion (HMP) at 4°C
(46). Under normal conditions,
histones are responsible for nucleosome assembly (47), chromatin stability (47) and epigenetic modifications in
transcription, replication and DNA repair (48). However, in the extracellular
space, they function as damage-associated molecular pattern
proteins (DAMPs) (49).
A previous retrospective study of 309 kidneys from
DCDs was conducted on extracellular histones in the renal
perfusate. van Smaalen et al (42) measured histone H3 levels at 1, 2
and 4 h of HMP. In uncontrolled DCD kidneys (Maastricht categories
1 and 2), no difference in H3 levels could be found compared with
the PNF, DGF and IF (immediate function) groups. In the control
cohort (Maastricht categories 3 and 4), a higher H3 concentration
in perfusate was associated with DGF [univariable odds ratio
(OR)=3.071; 95% CI, 1.588-5.941; multivariable OR=2.433; 95% CI,
1.253-4.723]. When comparing two groups of patients with histone H3
concentrations above and below the median, Kaplan-Meier analysis
revealed a significant difference in graft survival in favor of
lower concentrations. In particular, the 1- and 5-year survival
rates were significantly superior in the lower H3 group (at 1 year,
84 vs. 72%; at 5 years, 79 vs. 66%) (50).
FABPs
Ischemia leads to the overloading of proximal
tubular cells with free fatty acids (FFAs) and tubulo-interstitial
damage (51,52). During oxidative stress, FFAs bind
to fatty acid binding proteins (FABPs), thereby preventing their
intracellular accumulation and further oxidation (53). In addition, they can regulate the
expression of genes involved in FFA metabolism and promote their
β-oxidation in peroxisomes (53). To date, two isoforms have been
described in human proximal tubular cells, namely liver (L)-FABPs
and heart (H)-FABPs, which are also present in distal tubular cells
(54). Under normal conditions,
FABPs are involved in the cellular uptake and utilization of FFAs
from the plasma by rapid incorporation into triacylglycerols and
phospholipids, promotion of FFA metabolism in mitochondria or
peroxisomes, regulation of intracellular cholesterol metabolism,
regulation of gene expression involved in lipid metabolism and
modulation of cell proliferation (54,55).
Perfusate FABPs have been reported to associate with
DGF. Moers et al (33)
found that H-FABP was an independent predictor of DGF but only had
a moderate prognostic value. In another previous study, L-FABP
levels were reported to be associated with estimated glomerular
filtration rate (eGFR) 6 months after KTx (56). To the best of our knowledge, the
association between FABP levels in the renal perfusate and PNF has
not been studied. Sun et al (57) could not confirm the predictive
value of FABPs for DGF and 3-year post-transplant function
(expressed by eGFR value) in a retrospective study of cases of
DCDs. Perfusate FABPs levels have been shown to associate with high
vascular resistance and early graft dysfunction (33). Fig. 1 presents a list of molecules that
can be released after renal tubular cells disruption upon necrosis
and apoptosis resulting from ischemia.
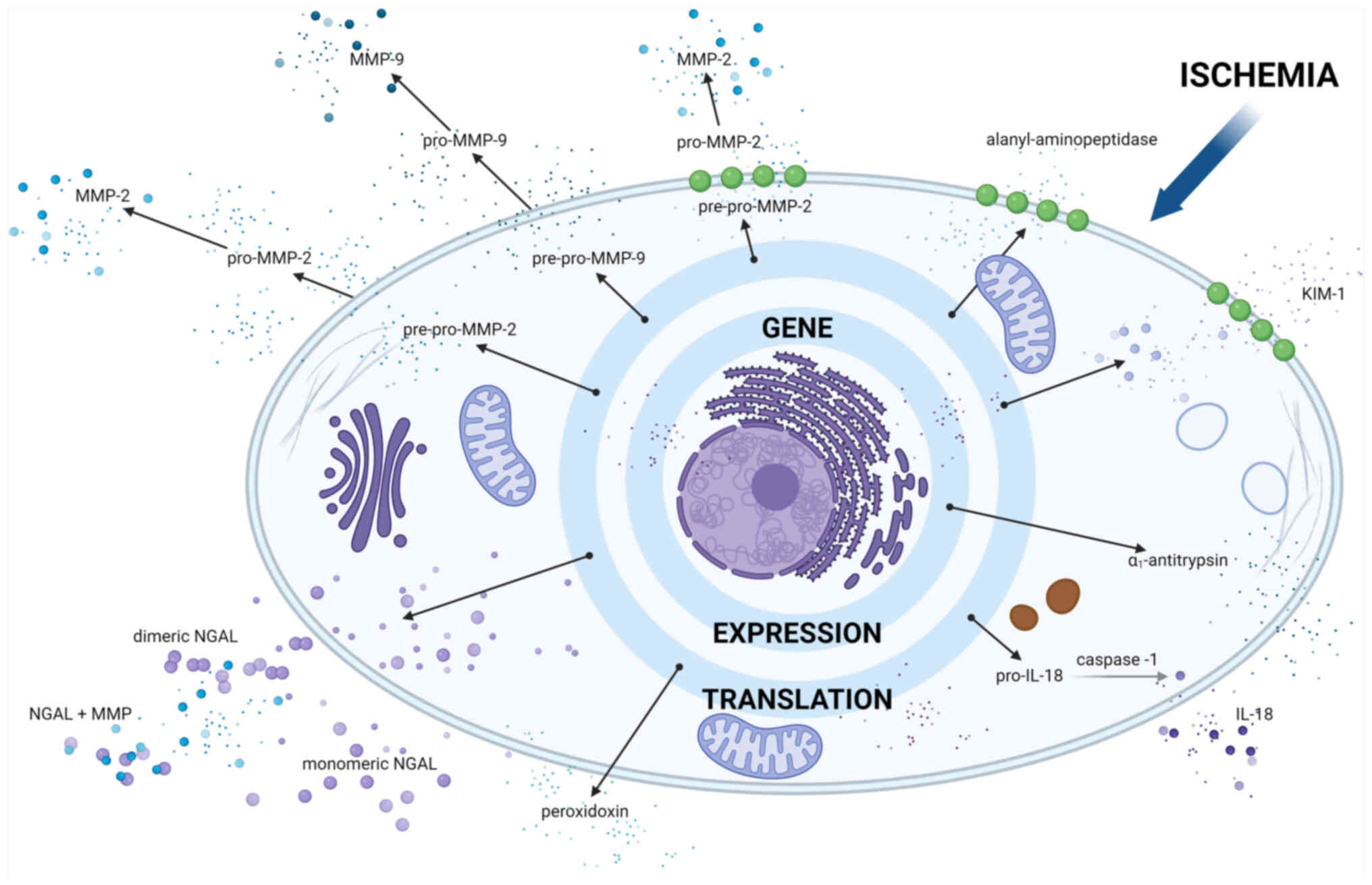
 | Figure 1Proteins released after cell
disruption. Metabolism switches from aerobic to anaerobic during
ischemia, resulting in lactate production and intracellular
acidosis. These conditions cause the inhibition of
Na+/K+-dependent ATPase activity, leading to
the accumulation of sodium ions and cellular edema. Other
consequences of ischemia include leakage of lysosomal enzymes,
cytoskeletal breakdown and decreased calcium excretion. Excess
calcium leads to the generation of ROS. Ischemia also triggers the
expression and redistribution of microtubule cytoskeletal elements.
Cell-cell integrity is altered, leading to loss of cell polarity
and redistribution of the basolateral membrane. All of the
aforementioned conditions can lead to cell death. The figure shows
preformed proteins that are released extracellularly after cell
death. The figure was created with BioRender® Science
Suite, Inc. ROS, reactive oxygen species; NAG,
N-Acetyl-/β-glucosaminidase; FABP, fatty acid binding protein; LDH,
lactate dehydrogenase; AST, aspartate aminotransferase; GST,
glutathione S-transferases. |
Proteins secreted during ischemia in
HMP
NGAL
NGAL is a protein that is originally synthesized in
the bone marrow and is stored in neutrophil granules (58). NGAL has been previously detected
in the lung, colon, trachea and renal epithelium (19). NGAL is capable of binding
siderophores. Therefore, it has been postulated that NGAL can serve
as a bacteriostatic agent by sequestering iron (59) and as a regulator of extracellular
iron-induced injury (60).
Production in non-hematopoietic locations has been associated with
systemic inflammation, where one such stimuli is IL-1β (61). NGAL is typically released in
response to ischemia (62) and
nephrotoxic agent (such as cisplatin) contact (63). In addition, ATP depletion induces
NGAL mRNA expression, leading to the production of NGAL (62). In this context, necrosis and cell
apoptosis is a factor that can trigger NGAL mRNA expression in
renal tubular cells (63).
In a previous systematic review, Guzzi et al
(15) found three studies on
NGAL (29,32,56), but only one (56) showing a positive association
between NGAL concentration in the HMP kidney perfusate and DGF
occurrence. The study by Moser et al (29) found that NGAL levels are not
significantly different between the DGF and non-DGF groups. In
another study by Hoogland et al (32), NGAL concentrations measured 4 h
after initiation of perfusion were not reported to be significantly
associated with the risk of DGF or PNF. Furthermore, the post-HMP
NGAL concentration was found to be associated with 6-month eGFR but
not with PNF (56). In another
study on discarded human kidneys, NGAL levels were found to be
markedly higher after 6 h of normothermic machine perfusion (NMP)
administration in organs from donors with higher serum creatinine
levels (sCr) at the time of retrieval (64).
However, care must be taken when examining the
perfusate for the presence of NGAL and interpreting the available
data. NGAL is secreted in different forms, including monomeric,
dimeric and conjugated with MMP-9 (65). Available assays utilizing
antibodies have not been suitably adapted to measure the monomeric
and heterodimeric forms released by injured or stressed tubular
cells (19). Another important
issue is the timing of sample collection from the perfusion
machine. NGAL synthesis and release are considered to be altered by
hypothermia (15). To determine
the most appropriate time interval, measuring NGAL in the renal
perfusate not only after 4 h, but also after 14-16 h of HMP, are of
importance to confirm the effect of hypothermia and time of
perfusion on its concentrations.
MMPs
MMPs have been recognized for their roles in
ischemia and reperfusion processes. They have been studied in the
context of ischemia-reperfusion injury in the central nervous
system (66), lungs (67), liver (68), heart, (69,70), skeletal muscle (71), retina (72) and kidney (73-77). MMPs, also known as matrixins, are
zinc-dependent enzymes that are located mainly in the extracellular
space (78). However, MMP-1,
MMP-2 and MMP-11 have been found to function intracellularly. To
date, 23 MMPs have been identified in humans (79). They are first synthetized as
pre-proenzymes and the majority are secreted into the extracellular
space. MMPs are typically produced after stimuli, such as by
inflammatory cytokines (TNF-α and IL-1), free radicals, reactive
oxygen species (ROS), oncogenic cellular transformation, physical
stress and chemical agents (78). In addition, a number of soluble
factors can induce the expression of MMPs and molecules on the cell
surface, including leukocyte function-associated antigen-1,
intercellular adhesion molecule-1-mediated cell adhesion very late
antigen 4 and vascular cell adhesion molecule (VCAM)-1 (80). The interaction between gp39-CD40
in monocytes and α5β1 integrin-fibronectin can also lead to MMP
expression (80). Secreted
pre-proenzymes are activated in the extracellular space by tissue,
plasma or opportunistic bacterial proteinases. In particular, the
urokinase-type plasminogen activator/plasmin system has been
reported to serve a role in MMP activation (81). Pro-MMP-2 has been reported to be
activated on the cell surface (82). Pro-MMP-2 is localized in
podosomes or the invadopodia (83,84). This activation process requires
the activation of the first membrane-type MMP (MT1-MMP), and the
tissue inhibitor of MMP-2 (TIMP-2)-bound MT1-MMP (78). The TIMP-2 in the latter complex
binds through its C-terminal domain to the hemopexin domain of
pro-MMP-2, which is assumed to localize the zymogen close to the
active MT1-MMP (78). It is also
activated by plasmin produced by urokinase-type plasmin activator
(from an inactive form), which is attached to a high-affinity cell
membrane binding site by a specific N-terminal sequence of its
non-catalytic chain (82).
MMP-2 and MMP-9, also known as gelatinases, are the
most frequently investigated MMPs in KTx and organ preservation.
MMP-2 activity contributes to the proteolysis of collagen types I,
IV, V, VII, X, XI and XIV, gelatin, elastin, fibronectin, laminin
and aggrecan (69,85). By contrast, MMP-9 targets include
collagen types IV, V, VII, X and XIV, gelatin, aggrecan, elastin,
entactin and fibronectin (86).
Data on the presence of MMPs in the renal perfusate and their
predictive value remain scarce. Moser et al (29) previously investigated the
activity of NGAL, LDH and MMPs in renal perfusates from DBDs and
donors after controlled cardiac death (cDCDs). MMPs could be
detected in all samples and a significant positive association
between perfusate MMP levels and the occurrence of DGF was
reported. However, this association was not observed for NGAL and
LDH levels. Additionally, higher concentrations of MMP-2 and MMP-9
were observed in the cDCDs group compared with those in the DBDs
group. In another previous proteomic analysis on three types of
donors, namely living kidney donors (LKDs), DBDs and cDCDs, the
levels of MMPs were found to be ranked in the following manner:
LKDs < DBDs < cDCDs (34).
Previous studies using animal models have shown that
different storage methods may influence MMP secretion. In one such
study, it was observed that HMP reduced MMP-9/NF-κB-dependent
expression compared with static cold storage (SCS) in a rabbit
model (87). Notably, HMP did
not affect MMP-2 expression. In another study performed by
Sulikowski et al (88) on
a rat model, the authors investigated the impact of University of
Wisconsin solution (UW) and EuroCollins solution (EC) on the gene
expression of MMP-2 and tissue inhibitor of MMP-2 (TIMP-2). After
24 h of cold ischemia time (CIT), the gene expression levels of
MMP-2 and TIMP-2 were found to be decreased in kidneys perfused
with UW, whereas they were increased in kidneys perfused with EC.
After warm ischemia, the levels of MMP-2 and TIMP-2 gene expression
were increased in kidneys perfused with UW, whereas they were
significantly lower in kidneys perfused with EC.
Kidney injury molecule-1 (KIM-1)
KIM-1, also known as hepatitis A virus cellular
receptor 1 and T cell immunoglobulin- and mucin-domain-containing
molecule, is considered to be a biomarker of ischemia-mediated
renal injury during preservation (56). Its attractiveness stems from its
specific expression pattern and function in renal proximal tubular
cells. KIM-1 is a cell membrane glycoprotein (89), which is comprised of an
extracellular portion, a six-cysteine immunoglobulin-like domain,
two N-glycosylation sites and a thrombospondin-rich domain, which
is characteristic of mucin-like O-glycosylated proteins (90). KIM-1 is predominantly localized
on the apical surface of renal proximal tubular epithelial cells
(91). It exhibits homology to a
hepatitis A virus receptor (89). KIM-1 expression is typically
absent under healthy conditions, but is activated under various
conditions and diseases, such as cell dedifferentiation, ischemic
injury (89), toxic injury
(92), autosomal dominant
polycystic kidney disease (93)
and renal cell carcinoma (94).
In the renal proximal epithelium, KIM-1 serves as a
phosphatidylserine receptor that can recognize apoptotic cells and
direct them into lysosomes (95). It can also bind to oxidized
lipoproteins in injured cells. In addition, KIM-1 has been observed
to mediate the transformation of epithelial cells into phagocytic
cells (96).
A previous study on 671 kidneys retrieved from
standard criteria donors (SCDs) and ECDs failed to demonstrate the
utility of KIM-1 as a biomarker, predictor of DGF or long-term
outcomes (56). KIM-1
concentrations were measured at the beginning and end of the
perfusion. The mean difference between the KIM-1 concentrations at
these points was not found to be associated with DGF. In an
experimental model of NMP of discarded human kidneys, KIM-1 was
found to be released at lower levels in kidneys with a higher urine
output (64). In a previous
study, in the DCDs kidney group, perfusate KIM-1 was proposed as an
independent marker of DGF and was correlated with 3-month eGFR
(57).
IL-18
IL-18 is a cytokine that belongs to the IL-1 family
and has been detected during hypothermic machine perfusion
(25,56). IL-18 precursors are present in
monocytes, macrophages, dendritic cells, intestinal cells,
keratocytes, epithelial cells and renal tubular cells (97), in addition to being readily
released by dying cells (97).
Transcription of the IL-18 precursor is stimulated by the binding
of Toll-like receptors to PAMPs, which then signals through the
NF-κB pathway (97). Activation
of the IL-18 precursor leads to its intracellular conversion into
its mature form by caspase-1 (98). Extracellular activation
mechanisms involve various proteases, such as neutrophil proteinase
3, granzyme B and chymase (99,100). In renal epithelial cells, the
IL-18 precursor can be activated and converted into its mature form
by the MMP meprin B (101).
IL-18 was originally identified to be an
IFN-γ-inducing factor. It serves an important role in both naive
and adaptive immunity (102).
Among IL-12 and IL-15, which can upregulate IL-18 receptor
expression, IL-18 induces IFN-γ production in CD4, CD8 T cells and
natural killer (NK) cells (103). In addition, IL-18 can directly
upregulate perforin- and FasL-dependent cytotoxicity in NK and CD8
T cells (103). IL-1β/IL-18
signaling initiates the transcription of several inflammatory
factors, including GM-CSF, IL-4, histamine and TNFα, which
facilitate leukocyte infiltration (104). IL-18, in the absence of IL-12
and IL-15, is responsible for the differentiation of naive T cells
into T helper 2 cells, which produce IL-4 and IL-13, (105,106). Upon IL-18 stimulation, mast
cells and basophils can produce IL-4 and IL-13 (107), whilst γδ T-cells produce IL-17
(97).
Previous studies on kidneys during storage using
perfusate IL-18 could not confirm its predictive value for DGF
(25,56). Its concentration was higher in
samples from kidneys developing DGF, but no statistical
significance could be found. In addition, although IL-18 associated
with PNF in kidneys from DCDs, the diagnostic accuracy of IL-18
alone was poor (56). Similarly,
in kidneys with a KDPI >80, IL-18 provided no predictive value
for PNF, DGF or 1-year post-transplant outcomes according to
another previous study (108).
Table I shows a list of protein
biomarkers detected using untargeted analysis methods. Fig. 2 shows molecules secreted during
ischemia in HMP.
 | Table IProtein biomarkers of kidney quality
detected in human perfusate. |
Table I
Protein biomarkers of kidney quality
detected in human perfusate.
| Protein class | Name | Type of donor | (Refs.) |
|---|
| Cytoplasmic
enzymes | Lactate
dehydrogenase | DBDs, DCDs and
LKDs | (15,16,26,29,34) |
|
Glutathione-S-transferases and its
isoforms | DBDs and DCDs | (15,16) |
| Glutathione
peroxidase | DBDs | (145) |
| Peroxidoxin-2 | DBDs, DCDs and
LKDs | (34) |
| Alanine
aminopeptidase (cytosolic form) | DCDs | (41) |
| Protein deglycase
DJ-1 | DCDs, DBDs and
LKDs | (34) |
| Alcohol
dehydrogenase, subunit β | DBDs and DCDs | (17) |
| Aldose
reductase | DBDs and DCDs | (17) |
| α-enolase | DBDs and DCDs | (17) |
| Aminoacylase | DBDs and DCDs | (17) |
| Superoxide
dismutase | DBDs | (144) |
| Mitochondrial
enzymes | Aspartate
aminotransferase | DBDs and DCDs | (33) |
| GAPDH | DBDs and DCDs | (17) |
| Superoxide
dismutase | DBDs | (144) |
| Lyzosomal
enzymes |
N-Acetyl-/β-glucosaminidase | DBDs and DCDs | (33) |
| Proteins secreted
extracellularly | MMP-2 | DCDs, DBDs and
LKDs | (34) |
| Neutrophil
gelatinase-associated lipocalin | DBDs, DCDs and
LKDs | (34) |
| α1-antitrypsin | DBDs, DCDs and
LKDs | (34) |
| Superoxide
dismutase | DBDs | (144) |
| Proteins attached
to cellular membrane | Kidney injury
molecule 1 | DBDs and DCDs | (56) |
| Alanine
aminopeptidase (membrane form) | DCDs | (41) |
| MMP-2 (membrane
attached form) | DBDs, DCDs and
LKDs | (29,34) |
| Nucleus
components | Histones H2A, H2B,
H3 and H4 | DCDs | (42) |
| Complement
elements | C3a, c3d, and
soluble c5b-9 | DCDs | (128) |
| C1q subcomponent
subunit B, C1s subcomponent, C1r subcomponent and C4b-binding
protein α chain | DBDs | (116) |
| Cytoskeletal
elements | Actinin-1 | DBDs | (116) |
| Talin-1 | DBDs | (116) |
| Desmoplakin | DBDs | (116) |
| Collagen α-1(I)
chain | DCDs, DBDs and
LKDs | (34) |
| Proteasome
elements | Liver-FABP | DCDs, DBDs and
LKDs | (34) |
| Heart-FABP | DBDs and DCDs | (56) |
| Leucyl
aminopeptidase | DCDs | (143) |
| Pyroglutamyl
aminopeptidase | DCDs | (143) |
| Arginyl
aminopeptidase | DCDs | (143) |
| Dipeptidyl
IV-aminopeptidase | DCDs | (143) |
| Peroxisomal
elements | Catalase | DBDs | (144) |
| Immunological
components | Immunoglobulin
heavy variable 2-26 | DBDs | (116) |
| Immunoglobulin
κ-chain | DBDs and DCDs | (17) |
| Immunoglobulin
γ1-chain | DBDs and DCDs | (17) |
| Immunoglobulin
γ2-chain | DBDs and DCDs | (17) |
| Immunoglobulin
γ3-chain | DBDs and DCDs | (17) |
| Immunoglobulin
γ3-chain | DBDs and DCDs | (17) |
| Vascular cell
adhesion molecule | DBDs | (141) |
| Intercellular
adhesion molecule | DBDs | (141) |
| TNF-α | DBDs | (141) |
| IFN-γ | DBDs | (141) |
| IL-1β, -2, -6,
-18 | DBDs and DCDs | (56,141) |
| IL-4, -8, -10 | Not mentioned | (145,146) |
| IL-1Rα | Not mentioned | (145) |
| IL-6R | DBDs | (141) |
| Macrophage
inflammatory protein-1α | Not mentioned | (145) |
| Monocyte
chemoattractant protein-1 | Not mentioned | (145) |
| Chemokine (C-X-C
motif) ligand 1 | DBDs | (141) |
| Cellular iron
homeostasis | Transferrin | DBDs and DCDs | (17) |
| Hemopexin | DBDs and DCDs | (17) |
| Haptoglobin | DBDs and DCDs | (17) |
| Oxyhemoglobin | DCDs | (117) |
| Blood
coagulation | Fibrinogen
γ-chain | DBDs and DCDs | (17) |
| Fibrinogen
α-chain | DCDs | (117) |
| Annexin A5 | DBDs and DCDs | (17) |
| Plasminogen
activator inhibitor-1 | DBDs and DCDs | (17) |
| Transport
proteins | Transthyretin | DBDs and DCDs | (17) |
| Albumin | DBDs and DCDs | (17) |
| Vitamin D binding
protein | DBDs and DCDs | (17) |
| High density
lipoprotein | DCDs | (117) |
Novel directions and highlights in proteomic
studies
Whole-proteome analysis
Studies conducted over the past decade have provided
novel insights and techniques into the proteomic analysis of the
renal perfusate, where untargeted protein analysis techniques,
including two-dimensional-polyacrylamide gel electrophoresis
(2D-PAGE) and liquid chromatography-mass spectrometry (LC-MS), have
been devised.
2D-PAGE (109)
is a potent method for detecting nearly all protein elements in
tested samples. According to this technique, proteins are separated
according to their isoelectric points by isoelectric focusing,
followed by SDS-PAGE, which segregates proteins based on their
molecular weight and electrical charge (110). Staining with Coomassie Blue or
silver helps to identify the target protein fractions, which can
then be excised for further analysis (111).
LC-MS is currently the most accurate method for
proteomic investigation (112).
This tool involves digesting the protein sample with proteolytic
enzymes, which is most commonly trypsin, to generate unique protein
mass value. These peptides are then separated by high-performance
liquid chromatography. After separation, MS is used to analyze each
peak, converting the samples to the gas phase and transferring them
to the ionization chamber. Electrospray ionization then ionizes the
samples into cations, which are subsequently separated based on
their mass-to-charge ratio using a mass analyzer. The detector
identifies and quantifies each ion and the results are compared
with database information (113). Proteomics most frequently uses
the following two modifications of the MS element known as the
ionizer: i) Matrix-assisted laser desorption/ionization (MALDI),
which is laser desorption with the participation of a matrix; and
ii) electrospray ionization. Other modifications can also be made
to mass analyzers. They include quadrupole mass analyzers, Fourier
transform ion cyclotron mass analyzer, Orbitrap mass analyzers, ion
trap mass analyzer and TOF mass analyzer (114). At present, the most commonly
applied method in proteome research is the time-of-flight (TOF)
analyzer combined with a MALDI ionizer (113). TOF allows for the study of the
flight time of ions between the ionizer and the detector, which
depends on the ratio of the mass to the number of ion charges
(112). The greatest advantage
of MALDI-TOF is the direct detection of the composition of a
complex mixture of proteins and other molecules (113,115). The underlying principles of
both 2D-PAGE and LC-MS are shown in Fig. 3.
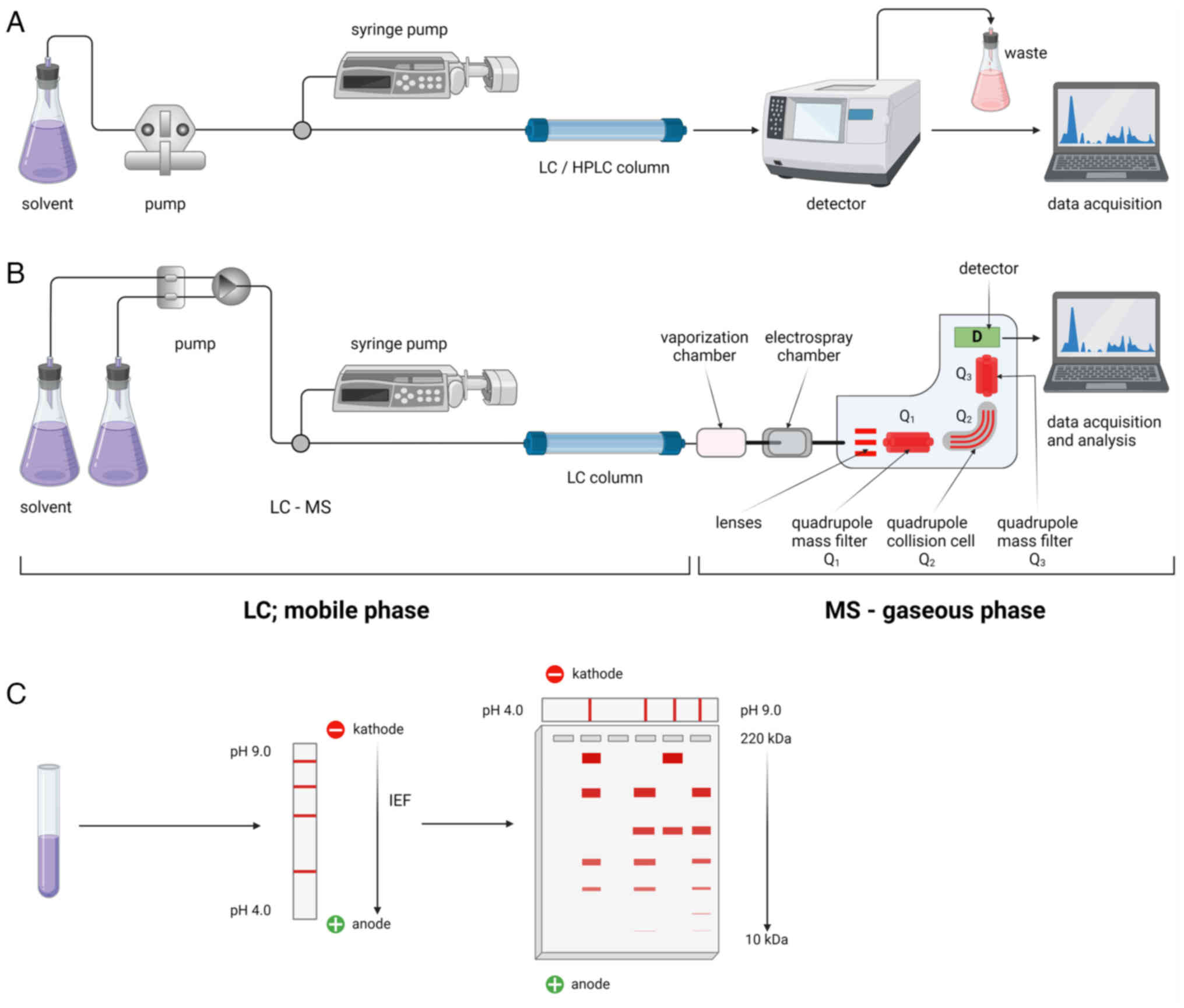 | Figure 3Whole-proteome analysis methods.
LC-MS serves as a vital tool for whole-proteome analysis. This
method involves digesting the protein sample with proteolytic
enzymes, commonly trypsin, generating unique protein mass value.
These peptides are then subjected to HPLC for separation. (A) After
separation, MS analyzes each peak, converting samples to the gas
phase and transferring them to the ionization chamber. (B)
Electrospray ionization ionizes the samples into cations, which are
subsequently separated based on their mass-to-charge ratio in a
mass analyzer. The detector identifies and quantifies each ion,
with results compared to database information. (C) 2D-PAGE is a
potent method for detecting nearly all protein elements in tested
samples. Proteins are separated according to their isoelectric
points through isoelectric focusing, followed by SDS-PAGE, which
segregates proteins based on their molecular mass and electrical
charge. Staining with Coomassie Blue or silver helps to identify
the target protein fractions, which can then be excised for further
analysis. The figure was created with BioRender® Science
Suite Inc. LC, liquid chromatography; HPLC, high performance LC;
MS, mass spectrometry; IEF, isoelectic focusing. |
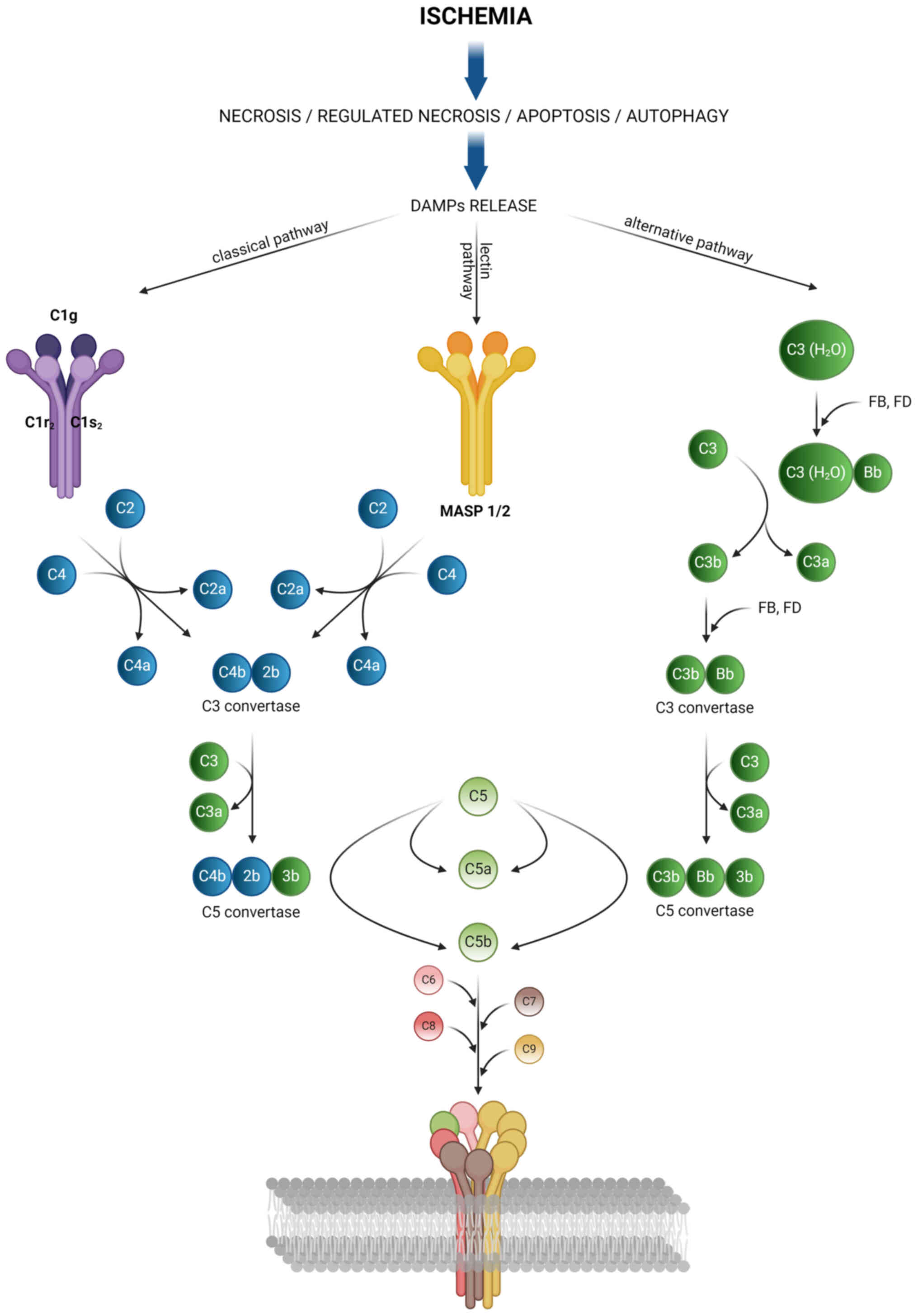
Complement elements
Previous studies on LC-MS analysis have highlighted
the role of the complement system in renal injury during HMP and
oxygenated-HMP (116,117). The complement system forms a
part of the innate immune response that contributes to extensive
endothelial cell damage (118).
This system contains soluble proteins (such as C3a, C3b and C5a)
and membrane-bound proteins (C5b, C6, C7, C8 and C9). Under
physiological conditions, regulatory proteins control the
degradation of C3 and C5 convertases to prevent overreactions
(118-120). Different elements are involved
in three pathways: Classical, alternative and lectin pathways. Both
ischemia and reperfusion are important factors that can trigger the
activation of the complement system (121). In addition, preformed
donor-specific antibodies can activate the complement system by
binding to C1q or C3d (122,123). Activation by DAMPs released
during ischemia and reperfusion leads to the formation of a
membrane-associated complex (MAC). This complex is comprised of
C5b-9 subunits and has been implicated in apoptosis induction
(124,125), direct cell injury, necrosis and
neutrophil influx (126). MAC
can also induce a pro-inflammatory response through the NF-κB
signaling pathway in endothelial cells (127). Fig. 4 presents the different pathways
of the complement system.
A previous study was performed on 22 kidneys
retrieved from DBDs stored in HMP (116). Perfusate samples that were
obtained after 15 min of HMP in patients with a good outcome (GO)
at 1-year post-transplantation (eGFR ≥45 ml/min/1.73 m2
and sCr level ≤120 μmol/l) were found to contain more
complement C1q subcomponent subunit B, complement C1s subcomponent
(C1S), complement C1r subcomponent (C1R) and C4b binding protein α
chain compared with those in perfusate samples of patients with a
suboptimal outcome (SO) at 1-year post-transplantation (eGFR ≤45
ml/min/1.73 m2 and sCr level ≥120 μmol/l).
Similar observations were made for samples collected after 4 h of
HMP. In the GO group, complement C1r subcomponent-like protein, C1S
and C1R were also found to be significantly upregulated compared
with those in the SO group. The authors showed that the
statistically significant difference in the concentration of
complement elements can distinguish between GO and SO at 1-year
post-transplantation (116).
Another study by Mulvey et al (117) focused on the proteomic
investigation of the perfusate from 67 paired DCDs kidneys stored
in oxygenated and non-oxygenated HMP, yielding 1,716 proteins
including complement cascade, platelet aggregation and
detoxification of reactive oxygen species. The most abundant
proteins identified were complement C3, apolipoprotein A1 and
fibrinogen α chain. One of the most abundant proteins was found to
be C3. In addition, complement cascade proteins (such as C1R, C1QC,
C3, C5, C7, C8A and C8G) were correlated with 12-month eGFR, where
their abundance decreased during HMP (117). Oxygenation did not affect the
proteome profile, including that of the complement cascade
(117). Using porcine kidneys
and discarded human kidneys (128), perfusion was previously
conducted with a blood-based solution for 4 and 6 h. C3a, C3d and
soluble C5b-9 (sC5b-9) were then measured in perfusate samples. In
porcine kidneys, the presence of sC5b-9 was reported to contribute
to lower creatinine clearance. It was then observed that the levels
of complement products in the perfusate were higher in the kidneys
from DBDs compared with those from DCDs.
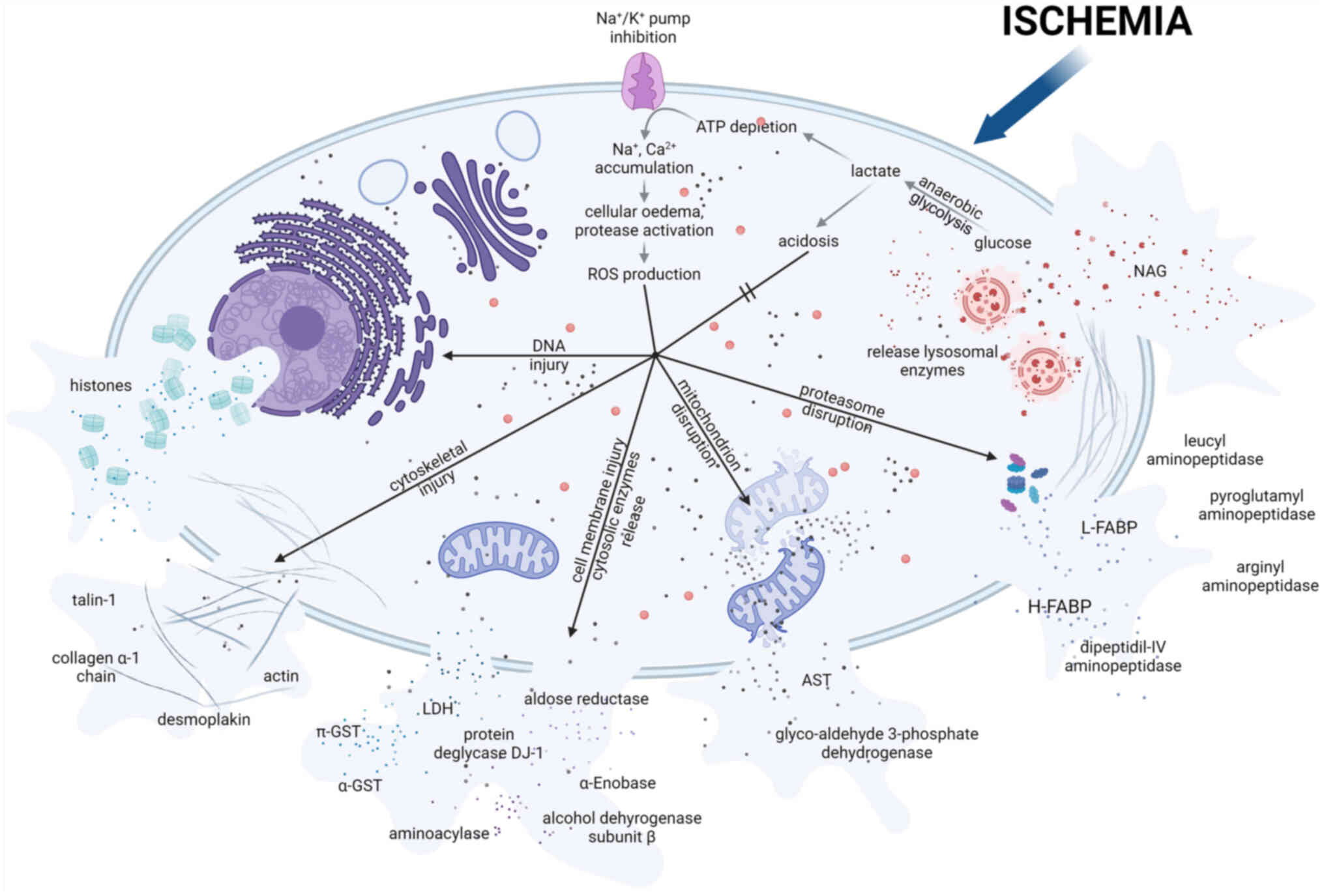
Cytoskeletal structural elements
To identify novel biomarkers, cytoskeletal
structural elements have been previously studied. These biomarkers
tend to be associated with non-specific cellular damage (34,116,129), unlike the aforementioned
markers associated with specific pathways or systems. During
ischemia, metabolism switches from aerobic to anaerobic conditions
(130). This results in
increased lactate production and intracellular acidosis. These
conditions inhibit Na+/K+-dependent ATPase
activity, leading to the accumulation of sodium ions and cellular
edema (131). Other
consequences of ischemia include leakage of lysosomal enzymes,
cytoskeletal breakdown and decreased calcium excretion (130). In addition, the excess calcium
accumulation that ensues leads to the generation of ROS (132), but the quantity of ROS
generated is not as potent as that during reperfusion. This is due
to the reduction in the activity of cytochromes, nitric oxide
synthases, xanthine oxidase and reduced nicotinamide adenine
dinucleotide phosphate oxidase activity (130). Ischemia has been shown to
trigger the expression and redistribution of microtubule
cytoskeletal elements (such as actin, fodrin, tubulin and
uvomorulin) (133,134). Cell integrity is then altered,
leading to loss of cell polarity and redistribution of the
basolateral membrane (135).
These conditions can lead to various forms of cell death, including
necrosis, apoptosis, autophagy and programmed necrosis (136).
A study by Coskun et al (129) revealed a group of structural
proteins associated with cellular damage in the kidney. In
particular, the preservation solution from 25 kidneys retrieved
from DBDs stored in SCS was analyzed. Among the 206 proteins
identified, a subset that associated significantly with donor
characteristics was identified. Specifically, perlecan levels were
correlated positively with sCr and blood urea nitrogen (BUN)
levels, increased level of talin-1 was related positively to CIT,
keratin type II cytoskeletal 8 was associated positively with
recipient sCr, whilst profilin was associated positively with donor
age (DA) and recipient BUN levels, myosin 6, 9 and 11 were
correlated with DA, collagen VI from epithelial cells was
associated positively with DA and recipient BUN levels, whereas
microfibril-associated glycoprotein 4 was associated positively
with DA and recipient sCr (113). Another study also to previously
investigated HMP perfusates for the presences of structural
components. van Leeuwen et al (116) studied 22 DBD kidneys and found
a positive correlation between actinin-1 and talin-1 concentration
in the perfusate and the 1-year post-transplant function expressed
by eGFR. Upregulated cytoskeletal proteins actinin-1 and talin-1 in
the HMP perfusate after 4 h of HMP discriminated kidneys with GO
and SO after 1-year post-transplantation. It was therefore
hypothesized that lower eGFR in the SO group was a result of the
loss of cellular integrity and disruption in the podocyte
architecture (116). In
addition, this previous study (116) reported that the presence of
desmoplakin quantified within LC-MS in the HMP perfusate after 4 h
of perfusion was associated with SO. Desmoplakin is one of the most
abundant components in desmosomes and cellular junctions (137). Together with immunoglobulin
heavy variable 2-26, desmoplakin had a predictive value of 86% for
SO occurrence in the first year after transplantation (116). The designs and results of
whole-proteome studies are listed in Tables II and III.
| Table IIWhole proteome analysis study
designs. |
Table II
Whole proteome analysis study
designs.
| Author, year | Analytical
method | Type of donor | Kidney amount | Outcome | Time of sample
acquistion | (Refs.) |
|---|
| Snoeijs et
al, 2013 | i) 2D-DIGE; ii)
LC-MS (spots identification) | DCD and DBD | 18 (6 uncontrolled
DCD; 6 controlled DCD; 6 DBD) | Immediate
function/DGF/PNF | 1 h of HMP | (17) |
| Moser et al,
2017 | i) 2D-DIGE; ii) MS
(spots identification); iii) Western blotting (results
confirmation) | DCD, DBD, LKD | 41 (9 DCD, 16 DBD,
16 LKD) | DGF/SGF (SGF was
defined as serum creatinine >265 μmol/l on
post-transplant day 5 but no need for dialysis by day 5) | End of HMP | (34) |
| van Leeuwen et
al, 2021 | LC-MS | DBD | 22 | Suboptimal outcome
at 1-year post-transplantation (kidney function with an eGFR ≥45
ml/min/1.73 m2 and a serum creatinine level of ≤120
μmol/l); good outcome at1-year post-transplantation (kidneys
with an eGFR ≤45 ml/min/1.73 m2 and a serum creatinine
level of ≥120 μmol/l 1-year post-transplantation) | 15 min of HMP (T1)
and just before ending HMP (T2) | (116) |
| Mulvey et
al, 2023 | LC-MS | DCD | 137 | eGFR measured one
year after kidney transplant | 15 min of HMP; just
before leaving donor's hospital; end of HMP | (117) |
| Table IIIWhole proteome analysis study
results. |
Table III
Whole proteome analysis study
results.
| Authors, year | Number of detected
proteins | Number of selected
proteins | Clinical
outcome | Analytical
method | Significance | (Refs.) |
|---|
| Snoeijs et
al, 2013 | 64 | 19 | Uncontrolled DCD, 4
DGF and 2 PNF; Controlled DCD, 3 IF and3 DGF; DBD, 3 IF and 3
DGF | i) 2D-DIGE; ii)
LC-MS (spots identification) | i) α1-antitrypsin
was upregulated in kidneys (considering all donor types) with DGF;
ii) haptoglobin was downregulated in the perfusate of DCD kidneys
(compared with DBD kidneys) | (17) |
| Moser et al,
2017 | Not mentioned | 7 | Not mentioned | i) 2D-DIGE; ii) MS
(spots identification) Western Blot (results confirmation) | i) total protein
concentration-DCD > DBD > LKD; ii) 7 protein significantly
different among the 3 groups, namely serum albumin, α-1-antitrypsin
peroxiredoxin-2, heavy chain of immuno-globulin, fragment of
collagen 1 and fatty acid binding protein, protein deglycase | (34) |
| van Leeuwen et
al, 2021 | 1,255 | T1, 28; T2, 48 | GO, 11 [12-month
eGFR 67.4 (15.5)]; SO, 11 [12-month eGFR 27.6 (9.7)] | LC-MS | T1: i) 18 proteins
significantly upregulated in GO vs. SO.; ii) all significant
proteins are located in extracellular regions, 5 of them belong to
complement and coagulation cascades, including complement C1q
subcomponent subunit B, complement C1s subcomponent, complement C1r
subcomponent and C4b-binding protein α chain; iii) at T1,
ATP-citrate synthase together with fatty acid binding protein 5
provided a predictive value of 91%, corresponding to a ROC with AUC
of 0.97; and iv) 10 proteins significantly downregulated in GO vs.
SO, 2 of them, fatty acid binding protein 4 and fatty acid binding
protein 5, are involved in Peroxisome proliferator-activated
receptor signalling. T2: i) 22 proteins significantly upregulated
in GO vs. SO, 5 of them belong to complement and coagulation
cascades, of which 4 are involved in complement activation of the
classical pathway, namely complement C1r subcomponent-like protein,
C1S and C1; ii) 26 proteins significantly downregulated in GO vs.
SO, 14 of them are cytoskeletal proteins, including desmoplakin,
talin-1 and α-actinin-1, 13 of the downregulated proteins are
affiliated with the immune system; and iii) immunoglobulin heavy
variable 2-26 togetherx heavy desmoplakine provided a predictive
value of 86%, corresponding to ROC with AUC of 0.95 for 26
proteins. | (116) |
| Mulvey et
al, 2023 | 1,716 | 137 | Mean eGFR in HMP +
O2 group, 41 ml/min/1, 73 m2; In HMP without
O2 group, 40 ml/min/1, 73 m2 | LC-MS | i) Overall HMP
duration associated with increasing levels of a subgroup of
proteins; ii) HDL and complement cascade proteins are associated
with 12-month eGFR; iii) blood-derived proteins are enriched in the
perfusate of kidneys that developed acute rejection, these proteins
belonged to 'regulation of response to wound healing' and
'metabolic processes contributed to ATP generation and nucleotide
phosphorylation'; iv) the most abundant proteins in all samples
were complement C3, apolipoprotein A1 and fibrinogen α-chain | (117) |
Other directions in proteomic
investigations
Both 2D-PAGE and LC-MS have certain limitations.
2D-PAGE lacks specificity and frequently requires additional
methods, such as western blotting, to confirm protein identity
(111). LC-MS in contrast
commands substantial financial investment, specialized equipment
and trained personnel. Sample preparation for quantification is
also time-consuming (112).
Modern methods, such as microarrays or protein
chips, have emerged for the analysis of large protein panels
(138). These microarrays
contain predefined panels of proteins or antibodies tailored to the
objectives of the experiment, such as analysis of specific protein
fractions, cancer markers or virus antigens. Upon addition of the
sample to the microarray plate, proteins will bind to the
antibodies if present. Antibodies conjugated to fluorochromes are
then introduced and bind to the previously formed complexes. This
process distinguishes between capture, reverse and functional
arrays, each designed for distinct analytical purposes. Fluorescent
detectors then provide quantitative and qualitative results,
enabling the simultaneous measurement of multiple compounds from a
single sample (139). When the
focus is on a limited number of proteins or a single protein, light
scattering-based protein analysis techniques come into play. These
include batch dynamic light scattering, static light scattering,
charge and ζ potential measurements, in addition to circular
dichroism spectrometry and isothermal titration calorimetry. These
methods provide valuable information regarding protein size,
abundance, charge, chirality, interactions and binding to other
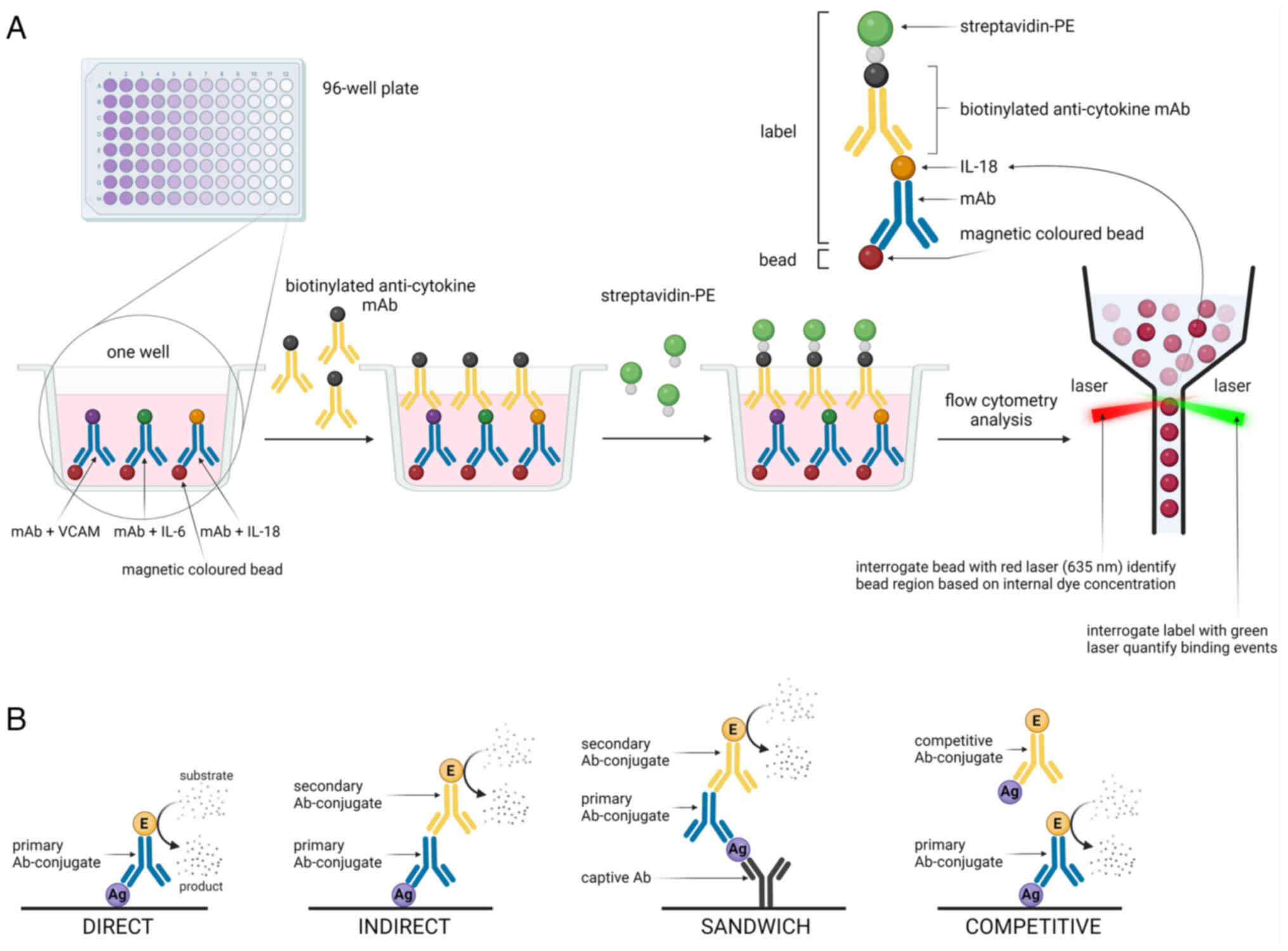
molecules (138,140). Perfusate analysis using a
method that is a bridge between classical ELISA and microarrays,
such as a multiplex immunoassay on a single platform, was performed
by Baboudjian et al (141) in a prospective study of 74
kidneys from deceased donors (both ECDs and SCDs). VCAM obtained at
the end of HMP was found to be a non-invasive predictor of early
graft dysfunction at both 1 week (OR=3.57; 95% CI, 1.06-12.03) and
3 months (OR=4.039; 95% CI, 1.11-14.73) after transplantation
(141). Table IV lists all the techniques
utilized for proteomic analysis of renal perfusate. Fig. 5 shows a representative multiplex
immunoassay on one platform and different types of ELISA.
| Table IVAnalytical methods for protein
detection in perfusate. |
Table IV
Analytical methods for protein
detection in perfusate.
| Method | Mechanism of
action | Advantages | Disadvantages |
|---|
| LC and
high-performance LC | Separation and
identification of proteins by mass transfer of a sample through a
polar mobile phase and a non-polar stationary phase. | i) High resolution
and speed of analysis; ii) precise flow rate control; iii) senstive
detection; and iv) low sample requirement | i) Expensive
solvents and columns; ii) regular maintenance and calibration is
needed; and iii) sophisticated software is required |
| LC-mass
spectrometry | Mixtures are
separated according to their physical and chemical properties, then
the components within each peak are identified and detected based
on their mass spectrum | i) The capability
to quantify multiple analytes simultaneously; and ii) the method
does not necessarily require immunological reagents | i) Regular
maintenance and calibration is needed; ii) sophisticated software
is required; iii) the instrument requires skilled personnel; and
iv) expensive |
| 2D-PAGE | Separation of
proteins in SDS gel by PAGE first by their charge then by their
size, identifying each protein after staining for example within
Coomassie Blue | i) Enables the
detection of hundreds of proteins on a single gel plate; and ii)
enhanced resolution, detection, quantitation and
reproducibility | i) Inability to
resolve proteins that are too basic or too acidic, too large or too
small; and ii) restricted to soluble and/or high abundant
proteins |
| Enzymatic activity
assay | Carry out a
reaction catalyzed by the protein under investigation and measure
the concentration of the enzyme product using colorimetric,
chemiluminescent or fluorescent method | i) High sensitivity
and specificity; ii) rapid response; iii) easy to perform | i) Difficulties in
assay standardization; ii) instability of the enzymesduring
storage |
| ELISA | Protein
identification by binding a specific antibody to an antigen present
on the desired molecule anddetecting a signal from staining agent
conjugated with an antibody. It could be performed in 4
manners:Direct, indirect, Sandwich, competitive | i high sensitivity
and specificity; ii) high throughput; iii) easy to perform; and iv)
it can determine the concentration of antigen in a sample | i) Information
limited to the amount or presence of the antigen in the sample; ii)
detection is based on enzyme/substrate reactions |
| Western
blotting | Separation of
proteins by gel electrophoresis to resolve them by size and
transfer to a membrane to immobilize the proteins. The proteins
immobilized on the membrane are then identified in an immunoassay
using colorimetric, chemiluminescent or fluorescent detection | i) The ability to
identify specific target proteins; and ii) high sensitivity | i) Time-consuming
procedure; ii) problems in reproducibility; and iii) false-positive
results when an antibody reacts with a non-intended protein |
| Multiplex
immunoassay on one platform | Simultaneous
binding of proteins of interest to specific antibody conjugated
beads, complex protein bead-antibody is conjugated to the
fluorescent antibody, the specific signals of protein of interest
are emitted by the fluorescent antibody and analyzed using a laser
detector | i) Good
reproducibility; ii) rapid processing speed; iii) low sample
volume; and iv) high sensitivity | i)
Cross-reactivity; ii) assay development time; iii) commitment to
vendor-supplied assays; and iv) dynamic range |
Other potential protein biomarkers
In whole-proteome studies, other non-specific
proteins associated with kidney injury have been detected in HMP
perfusates. Proteins found at statistically significant levels
include immunoglobulins κ, γ1, γ2 and γ3 chains (17). α1-Anti-trypsin with peroxidoxin-2
may discriminate between renal damage in DBDs, DCDs and LKDs
(34).
Another group of proteins considered to be
biomarkers and detected by whole-proteome analysis methods include
proteins involved in transport (transthyretin, albumin and vitamin
D-binding protein), blood coagulation (fibrinogen α-chain,
fibrinogen γ-chain, annexin A5 and plasminogen activator
inhibitor-1), energy metabolism (alcohol dehydrogenase subunit β,
GAPDH, α-enolase, aldose reductase and aminoacylase), cellular iron
homeostasis (transferrin, hemopexin, haptoglobin and oxyhemoglobin)
and high-density lipoproteins (17,31). However, further studies are
required to confirm their role in predicting short- and long-term
outcomes in recipients.
A previous targeted proteomic analysis revealed the
presence of Ala-AP (41,142), leucyl- and pyroglutamyl
aminopeptidase (143),
alanyl-arginyl- and dipeptidyl IV-aminopeptidase (143), aspartate aminotransferase
(33) and
N-acetyl-β-d-gluconamidase (33), glutathione peroxidase, catalase
and superoxide dismutase (144), TNF-α, IFN-γ (141), IL-1β, -2, -4, -6, -8 and -10
(141,145,146), IL-1Rα, macrophage inflammatory
protein-1, monocyte chemoattractant protein-1 (145), IL-6R and chemokine C-X-C motif
ligand 1 (141). However, these
proteins have been previously investigated, but none was found to
be a reliable predictor of short- or long-term outcomes in
recipients.
Conclusions and future perspectives
The number of patients with end-stage renal disease
awaiting kidney transplantation is much higher compared with the
number of transplanted organs (147). Therefore, an increasing number
of kidneys have been retrieved from ECDs, which increases the risk
of poor outcomes (148-150). However, organs in good
condition, namely those not developing DGF or not requiring
continuous dialysis therapy after KTx, are typically selected from
this donor group.
Proteomic analysis of the perfusate remains to be a
promising tool for the evaluation of kidneys before
transplantation. This is especially true for the ECDs, DCDs and
SCDs. Despite previous studies on this topic, protein markers that
are sufficiently accurate for predicting the state of the kidney
before KTx have remained elusive. However, the problem lies in the
methodologies used in such studies. Searching for only one or a
small group of proteins may not be the optimal approach. Changes in
kidney metabolism during the entire transplantation procedure are
extensive and must be monitored constantly. Therefore, biomarkers
should include a large panel of proteins. Modern techniques of
protein analysis will allow for such studies, whereas fast
diagnostic methods (such as microarrays) will allow for the
up-to-date supervision of kidney state during perfusion. To
identify a reliable group of markers, profiling research must show
the correlations of altered protein levels in the kidney and
metabolic pathways activated during organ transplantation. Such
pathways include ischemia-reperfusion, endothelial reticulum
stress, oxidative stress and autophagy pathways. The optimal panel
of markers should establish connections with all the proteins.
Another consideration should be the type of
materials used for proteomic analysis. The present study has
described proteins detected during HMP, which is a recognized
method for kidney preservation, particularly for organs from ECDs
(151,152). Continuous or pulsatile
perfusion allows for the release and secretion of proteins into the
perfusate. As a result, the dynamics of the processes during the
HMP can be monitored. In addition, the quantity of proteins
detected is likely to be more reliable compared with that detected
during SCS. HMP ensures the removal of blood residues that may
interfere with the final conclusions regarding the proteome
profile.
Proteomic studies should be performed in different
donor groups if the study aims to find an association between the
detected protein and the selected endpoint. Various factors, such
as CIT (153), diabetes and
hypertension (154), cause of
mortality (34), age (155,156) and terminal sCr level (157,158), can all affect the condition of
the donated kidneys. Mixing organs from ECDs and SCDs will not lead
to correct conclusions in the context of the selected endpoint due
to the aforementioned factors. Another issue is defining the
endpoint. In the majority of studies, DGF was defined as the need
for dialysis within the first week after KTx (159-162). However, dialysis can be
performed due to post-operative hyperkalemia or fluid overload that
does not result from renal injury (163,164). Further studies should consider
a more appropriate definition of DGF and other endpoints.
The quantification method is crucial for certain
proteins. This applies to biomarkers that have isoforms, such as
GSTs or form complexes, such as NGAL. Once a biomarker has been
selected from the entire proteome, the next step should involve
finding a suitable laboratory test or developing a novel method to
detect the entire quantity of the desired molecule.
In summary, although numerous studies have been
conducted with conflicting results, perfusate remains to be a
promising source of protein biomarkers. Novel studies concerning
the aforementioned issues may unravel other properties of
well-known proteins, such as NGAL, KIM-1 or MMPs. Whole-proteome
techniques will likely provide insights into non-specific kidney
processes during preservation and complex protein profiles of
numerous biological samples. Methods such as immunofixation,
western blotting and ELISA enable the precise detection and
quantification of specific proteins based on existing data.
Immunofixation separates proteins electrophoretically and
identifies them using specific antibodies, whilst western blotting
employs primary and secondary antibodies for protein detection and
ELISA quantifies proteins through spectrophotometric analysis.
Current advancements for studying proteins include
microarrays and protein chips, which will allow for the
simultaneous analysis of multiple targets. These techniques are
particularly beneficial for studying specific protein fractions,
cancer markers or viral antigens. Light scattering methods, such as
dynamic light scattering and circular dichroism spectrometry,
provide additional details on protein size, charge and
interactions. Untargeted techniques, such as 2D-PAGE and LC-MS,
enable comprehensive proteome profiling. 2D-PAGE separates proteins
by isoelectric point and molecular mass, whereas LC-MS identifies
proteins through peptide mass fingerprints and enzymatic digestion.
Despite their complexity and cost, these methods will prove crucial
for identifying novel proteins and understanding proteomic changes.
Collectively, these techniques compose a robust framework for
proteomic analysis, which is expected to advance the current
understanding of protein function, interactions and roles in renal
health and disease.
Recent studies on the role of the complement system
and cytoskeletal elements show the association with short- and
long-term outcomes (116,117,129). However, randomized controlled
trials are required to establish their role in the assessment of
kidney quality before transplantation. In addition, targeted
methods for a protein of interest should be developed for further
analysis. Despite novel storage protocols, such as NMP and SNMP,
hypothermic methods remain in use in numerous countries and
proteomic analysis of these perfusate is justified. SCS is the
current storage standard for kidneys procured from DCDs in the UK
(165). In the Netherlands, HMP
is the standard method for kidney preservation in all donor types
(166). To date, one randomized
controlled trial has been performed in which SCS alone was compared
with SCS plus a 1-h period of NMP at the end of storage (165). This previous study showed that
SCS plus 1-h of NMP is not superior in reducing the DGF rate
compared with kidney preservation by SCS alone, since this method
did not improve the long-term results (135). However, additional studies
comparing longer periods of NMP with HMP (165,167) are essential to confirm the
safety and superiority of normothermic techniques over HMP or SCS.
Nevertheless, proteomic analysis of the solutions used for
normothermic perfusion should be performed as a source of novel
biomarkers.
Availability of data and materials
Not applicable.
Authors' contributions
MB contributed to the design of the review. MB and
MiS wrote the manuscript. MB and RD saw and verified all articles
included in this review from the following four databases: Embase,
PubMed (Medline), Web of Sscience and Scopus. MB designed the
figures and tables. MiS, RD, MK and MaS revised the manuscript. All
authors read and approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Acknowledgements
The authors would like to thank Ms Anna Palińska
(Off Course School, Warsaw) for creating the figures with
BioRender® Science Suite Inc.
Funding
No funding was received.
References
|
1
|
Querard AH, Le Borgne F, Dion A, Giral M,
Mourad G, Garrigue V, Rostaing L, Kamar N, Loupy A, Legendre C, et
al: Propensity score-based comparison of the graft failure risk
between kidney transplant recipients of standard and expanded
criteria donor grafts: Toward increasing the pool of marginal
donors. Am J Transplant. 18:1151–1157. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Querard AH, Foucher Y, Combescure C,
Dantan E, Larmet D, Lorent M, Pouteau LM, Giral M and Gillaizeau F:
Comparison of survival outcomes between expanded criteria donor and
standard criteria donor kidney transplant recipients: A systematic
review and meta-analysis. Transpl Int. 29:403–415. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Barba J, Zudaire JJ, Robles JE, Rosell D,
Berian JM and Pascual I: Complications of kidney transplantation
with grafts from expanded criteria donors. World J Urol.
31:893–900. 2013. View Article : Google Scholar
|
|
4
|
Coupel S, Giral-Classe M, Karam G, Morcet
JF, Dantal J, Cantarovich D, Blancho G, Bignon JD, Daguin P,
Soulillou JP and Hourmant M: Ten-year survival of second kidney
transplants: Impact of immunologic factors and renal function at 12
months. Kidney Int. 64:674–680. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Kousoulas L, Vondran FWR, Syryca P,
Klempnauer J, Schrem H and Lehner F: Risk-adjusted analysis of
relevant outcome drivers for patients after more than two kidney
transplants. J Transplant. 2015:7120492015. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Zádori G, Kovács DÁ, Fedor R, Kanyári Z,
Zsom L, Asztalos L and Nemes B: Results of expanded-criteria donor
kidneys: A single-center experience in hungary. Transplant Proc.
47:2189–2191. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Hwang JK, Park SC, Kwon KH, Choi BS, Kim
JI, Yang CW, Kim YS and Moon IS: Long-term outcomes of kidney
transplantation from expanded criteria deceased donors at a single
center: Comparison with standard criteria deceased donors.
Transplant Proc. 46:431–436. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
De Beule J and Jochmans I: Kidney
perfusion as an organ quality assessment tool-are we counting our
chickens before they have hatched? J Clin Med. 9:8792020.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Mourão TB, Mine KL, Campos EF,
Medina-Pestana JO, Tedesco-Silva H and Gerbase-DeLima M: Predicting
delayed kidney graft function with gene expression in
preimplantation biopsies and first-day posttransplant blood. Hum
Immunol. 77:353–357. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Hall IE, Reese PP, Weng FL, Schröppel B,
Doshi MD, Hasz RD, Reitsma W, Goldstein MJ, Hong K and Parikh CR:
Preimplant histologic acute tubular necrosis and allograft
outcomes. Clin J Am Soc Nephrol. 9:573–582. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Bachmann Q, Haberfellner F, Büttner-Herold
M, Torrez C, Haller B, Assfalg V, Renders L, Amann K, Heemann U,
Schmaderer C and Kemmner S: The kidney donor profile index (KDPI)
correlates with histopathologic findings in post-reperfusion
baseline biopsies and predicts kidney transplant outcome. Front Med
(Lausanne). 9:8752062022. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Rege A, Irish B, Castleberry A, Vikraman
D, Sanoff S, Ravindra K, Collins B and Sudan D: Trends in usage and
outcomes for expanded criteria donor kidney transplantation in the
United States characterized by kidney donor profile index. Cureus.
8:e8872016.PubMed/NCBI
|
|
13
|
Rao PS, Schaubel DE, Guidinger MK,
Andreoni KA, Wolfe RA, Merion RM, Port FK and Sung RS: A
comprehensive risk quantification score for deceased donor kidneys:
The kidney donor risk index. Transplantation. 88:231–236. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Nyberg SL, Baskin-Bey ES, Kremers W,
Prieto M, Henry ML and Stegall MD: Improving the prediction of
donor kidney quality: Deceased donor score and resistive indices.
Transplantation. 80:925–929. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Guzzi F, Knight SR, Ploeg RJ and Hunter
JP: A systematic review to identify whether perfusate biomarkers
produced during hypothermic machine perfusion can predict graft
outcomes in kidney transplantation. Transpl Int. 33:590–602. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Bhangoo RS, Hall IE, Reese PP and Parikh
CR: Deceased-donor kidney perfusate and urine biomarkers for kidney
allograft outcomes: A systematic review. Nephrol Dial Transplant.
27:3305–3314. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Snoeijs MG, Pulinx B, van Dieijen-Visser
MP, Buurman WA, van Heurn LW and Wodzig WK: Characterization of the
perfusate proteome of human donor kidneys. Ann Clin Biochem.
50:140–146. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Khan AA, Allemailem KS, Alhumaydhi FA,
Gowder SJT and Rahmani AH: The biochemical and clinical
perspectives of lactate dehydrogenase: An enzyme of active
metabolism. Endocr Metab Immune Disord Drug Targets. 20:855–868.
2020. View Article : Google Scholar
|
|
19
|
Mårtensson J and Bellomo R: The rise and
fall of NGAL in acute kidney injury. Blood Purif. 37:304–310. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Markert CL: Lactate dehydrogenase.
Biochemistry and function of lactate dehydrogenase. Cell Biochem
Funct. 2:131–134. 1984. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Holmes RS and Goldberg E: Computational
analyses of mammalian lactate dehydrogenases: Human, mouse, opossum
and platypus LDHs. Comput Biol Chem. 33:379–385. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Dubach UC: On the origin of lactic
dehydrogenase isoenzymes in urine. Helv Med Acta. 33:139–150.
1966.PubMed/NCBI
|
|
23
|
Osis G, Traylor AM, Black LM, Spangler D,
George JF, Zarjou A, Verlander JW and Agarwal A: Expression of
lactate dehydrogenase A and B isoforms in the mouse kidney. Am J
Physiol Renal Physiol. 320:F706–F718. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Kootstra G and Daemen JH: The
non-heart-beating donor. Transplant Proc. 28:161996.PubMed/NCBI
|
|
25
|
Daemen JW, Oomen AP, Janssen MA, van de
Schoot L, van Kreel BK, Heineman E and Kootstra G: Glutathione
S-transferase as predictor of functional outcome in transplantation
of machine-preserved non-heart-beating donor kidneys.
Transplantation. 63:89–93. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Modgill VK, Wiggins PA, Rosenberg IL,
Humphrey CS and Giles GR: An evaluation of viability tests of human
cadaveric kidneys. Br J Surg. 64:548–553. 1977. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Skillen AW: Clinical biochemistry of
lactate dehydrogenase. Cell Biochem Funct. 2:140–144. 1984.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Huijgen HJ, Sanders GT, Koster RW, Vreeken
J and Bossuyt PM: The clinical value of lactate dehydrogenase in
serum: A quantitative review. Eur J Clin Chem Clin Biochem.
35:569–579. 1997.PubMed/NCBI
|
|
29
|
Moser MA, Arcand S, Lin HB, Wojnarowicz C,
Sawicka J, Banerjee T, Luo Y, Beck GR, Luke PP and Sawicki G:
Protection of the transplant kidney from preservation injury by
inhibition of matrix metalloproteinases. PLoS One. 11:e01575082016.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Nagelschmidt M, Minor T, Gallinat A, Moers
C, Jochmans I, Pirenne J, Ploeg RJ, Paul A and Treckmann J: Lipid
peroxidation products in machine perfusion of older donor kidneys.
J Surg Res. 180:337–342. 2013. View Article : Google Scholar
|
|
31
|
de Vries B, Snoeijs MGJ, von Bonsdorff L,
Ernest van Heurn LW, Parkkinen J and Buurman WA: Redox-active iron
released during machine perfusion predicts viability of
ischemically injured deceased donor kidneys. Am J Transplant.
6:2686–2693. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Hoogland ER, de Vries EE, Christiaans MH,
Winkens B, Snoeijs MG and van Heurn LW: The value of machine
perfusion biomarker concentration in DCD kidney transplantations.
Transplantation. 95:603–610. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Moers C, Varnav OC, van Heurn E, Jochmans
I, Kirste GR, Rahmel A, Leuvenink HG, Squifflet JP, Paul A, Pirenne
J, et al: The value of machine perfusion perfusate biomarkers for
predicting kidney transplant outcome. Transplantation. 90:966–973.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Moser MAJ, Sawicka K, Arcand S, O'Brien P,
Luke P, Beck G, Sawicka J, Cohen A and Sawicki G: Proteomic
analysis of perfusate from machine cold perfusion of transplant
kidneys: Insights into protection from injury. Ann Transplant.
22:730–739. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Udomsinprasert R, Pongjaroenkit S,
Wongsantichon J, Oakley AJ, Prapanthadara LA, Wilce MC and
Ketterman AJ: Identification, characterization and structure of a
new Delta class glutathione transferase isoenzyme. Biochem J.
388:763–771. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Di Ilio C, Aceto A, Bucciarelli T,
Angelucci S, Felaco M, Grilli A, Zezza A, Tenaglia R and Federici
G: Glutathione transferase isoenzymes in normal and neoplastic
human kidney tissue. Carcinogenesis. 12:1471–1475. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Harrison DJ, Kharbanda R, Cunningham DS,
McLellan LI and Hayes JD: Distribution of glutathione S-transferase
isoenzymes in human kidney: Basis for possible markers of renal
injury. J Clin Pathol. 42:624–628. 1989. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Laborde E: Glutathione transferases as
mediators of signaling pathways involved in cell proliferation and
cell death. Cell Death Differ. 17:1373–1380. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Hall IE, Bhangoo RS, Reese PP, Doshi MD,
Weng FL, Hong K, Lin H, Han G, Hasz RD, Goldstein MJ, et al:
Glutathione S-transferase iso-enzymes in perfusate from pumped
kidneys are associated with delayed graft function. Am J
Transplant. 14:886–896. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Qiao Y, Ding C, Li Y, Tian X, Tian P, Ding
X, Xiang H, Zheng J and Xue W: Predictive value of hypothermic
machine perfusion parameters combined perfusate biomarkers in
deceased donor kidney transplantation. Chin Med J (Engl).
135:181–186. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Gok MA, Pelsers M, Glatz JFC, Bhatti AA,
Shenton BK, Peaston R, Cornell C, Mantle D and Talbot D: Comparison
of perfusate activities of glutathione S-transferase, alanine
aminopeptidase and fatty acid binding protein in the assessment of
non-heart-beating donor kidneys. Ann Clin Biochem. 40:252–258.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
van Smaalen TC, Beurskens DMH, Hoogland
ERP, Winkens B, Christiaans MHL, Reutelingsperger CP, van Heurn LWE
and Nicolaes GAF: Presence of cytotoxic extracellular histones in
machine perfusate of donation after circulatory death kidneys.
Transplantation. 101:e93–e101. 2017. View Article : Google Scholar
|
|
43
|
Felsenfeld G and Groudine M: Controlling
the double helix. Nature. 421:448–453. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Silk E, Zhao H, Weng H and Ma D: The role
of extracellular histone in organ injury. Cell Death Dis.
8:e28122017. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Wickman GR, Julian L, Mardilovich K,
Schumacher S, Munro J, Rath N, Zander SA, Mleczak A, Sumpton D,
Morrice N, et al: Blebs produced by actin-myosin contraction during
apoptosis release damage-associated molecular pattern proteins
before secondary necrosis occurs. Cell Death Differ. 20:1293–1305.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
van Smaalen TC, Beurskens DMH, Kox JJHFM,
Polonia R, Vos R, Duimel H, van de Wetering WJ, López-Iglesias C,
Reutelingsperger CP, Ernest van Heurn LW, et al: Extracellular
histone release by renal cells after warm and cold ischemic kidney
injury: Studies in an ex-vivo porcine kidney perfusion model. PLoS
One. 18:e02799442023. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Campos EI and Reinberg D: Histones:
Annotating chromatin. Annu Rev Genet. 43:559–599. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Strahl BD and Allis CD: The language of
covalent histone modifications. Nature. 403:41–45. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Kono H and Rock KL: How dying cells alert
the immune system to danger. Nat Rev Immunol. 8:279–289. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Li B, Hao J, Zeng J and Sauter ER:
SnapShot: FABP functions. Cell. 182:1066–1066.e1. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Zager RA, Johnson ACM and Hanson SY: Renal
tubular triglyercide accumulation following endotoxic, toxic, and
ischemic injury. Kidney Int. 67:111–121. 2005. View Article : Google Scholar
|
|
52
|
Bobulescu IA: Renal lipid metabolism and
lipotoxicity. Curr Opin Nephrol Hypertens. 19:393–402. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Storch J and Thumser AE: The fatty acid
transport function of fatty acid-binding proteins. Biochim Biophys
Acta. 1486:28–44. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Pelsers MMAL: Fatty acid-binding protein
as marker for renal injury. Scand J Clin Lab Invest Suppl.
241:73–77. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Yamamoto T, Noiri E, Ono Y, Doi K, Negishi
K, Kamijo A, Kimura K, Fujita T, Kinukawa T, Taniguchi H, et al:
Renal L-type fatty acid-binding protein in acute ischemic injury. J
Am Soc Nephrol. 18:2894–2902. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Parikh CR, Hall IE, Bhangoo RS, Ficek J,
Abt PL, Thiessen-Philbrook H, Lin H, Bimali M, Murray PT, Rao V, et
al: Associations of perfusate biomarkers and pump parameters with
delayed graft function and deceased donor kidney allograft
function. Am J Transplant. 16:1526–1539. 2016. View Article : Google Scholar :
|
|
57
|
Sun Z, Gao Z, Li X, Zheng X, Wang W and
Qiao P: Perfusate neutrophil gelatinase-associated lipocalin,
kidney injury molecular-1, liver-type fatty acid binding protein,
and interleukin-18 as potential biomarkers to predict delayed graft
function and long-term prognosis in kidney transplant recipients: A
single-center retrospective study. Med Sci Monit. 29:e9387582023.
View Article : Google Scholar
|
|
58
|
Borregaard N and Cowland JB: Granules of
the human neutrophilic polymorphonuclear leukocyte. Blood.
89:3503–3521. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Flo TH, Smith KD, Sato S, Rodriguez DJ,
Holmes MA, Strong RK, Akira S and Aderem A: Lipocalin 2 mediates an
innate immune response to bacterial infection by sequestrating
iron. Nature. 432:917–921. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Yang J, Goetz D, Li JY, Wang W, Mori K,
Setlik D, Du T, Erdjument-Bromage H, Tempst P, Strong R and Barasch
J: An iron delivery pathway mediated by a lipocalin. Mol Cell.
10:1045–1056. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Cowland JB, Sørensen OE, Sehested M and
Borregaard N: Neutrophil gelatinase-associated lipocalin is
up-regulated in human epithelial cells by IL-1 beta, but not by
TNF-alpha. J Immunol. 171:6630–6639. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Mishra J, Ma Q, Prada A, Mitsnefes M,
Zahedi K, Yang J, Barasch J and Devarajan P: Identification of
neutrophil gelatinase-associated lipocalin as a novel early urinary
biomarker for ischemic renal injury. J Am Soc Nephrol.
14:2534–2543. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Mishra J, Mori K, Ma Q, Kelly C, Barasch J
and Devarajan P: Neutrophil gelatinase-associated lipocalin: A
novel early urinary biomarker for cisplatin nephrotoxicity. Am J
Nephrol. 24:307–315. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Weissenbacher A, Stone JP, Lo Faro ML,
Hunter JP, Ploeg RJ, Coussios CC, Fildes JE and Friend PJ:
Hemodynamics and metabolic parameters in normothermic kidney
preservation are linked with donor factors, perfusate cells, and
cytokines. Front Med (Lausanne). 8:8010982022. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Cai L, Rubin J, Han W, Venge P and Xu S:
The origin of multiple molecular forms in urine of HNL/NGAL. Clin J
Am Soc Nephrol. 5:2229–2235. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Rosell A and Lo EH: Multiphasic roles for
matrix metalloproteinases after stroke. Curr Opin Pharmacol.
8:82–89. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Soccal PM, Gasche Y, Miniati DN, Hoyt G,
Berry GJ, Doyle RL, Theodore J and Robbins RC: Matrix
metalloproteinase inhibition decreases ischemia-reperfusion injury
after lung transplantation. Am J Transplant. 4:41–50. 2004.
View Article : Google Scholar
|
|
68
|
Viappiani S, Sariahmetoglu M and Schulz R:
The role of matrix metalloproteinase inhibitors in
ischemia-reperfusion injury in the liver. Curr Pharm Des.
12:2923–2934. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Nagase H, Visse R and Murphy G: Structure
and function of matrix metalloproteinases and TIMPs. Cardiovasc
Res. 69:562–573. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Chow AK, Cena J and Schulz R: Acute
actions and novel targets of matrix metalloproteinases in the heart
and vasculature. Br J Pharmacol. 152:189–205. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Roach DM, Fitridge RA, Laws PE, Millard
SH, Varelias A and Cowled PA: Up-regulation of MMP-2 and MMP-9
leads to degradation of type IV collagen during skeletal muscle
reperfusion injury; protection by the MMP inhibitor, doxycycline.
Eur J Vasc Endovasc Surg. 23:260–269. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Mathalone N, Lahat N, Rahat MA,
Bahar-Shany K, Oron Y and Geyer O: The involvement of matrix
metalloproteinases 2 and 9 in rat retinal ischemia. Graefes Arch
Clin Exp Ophthalmol. 245:725–732. 2007. View Article : Google Scholar
|
|
73
|
Cavdar Z, Ural C, Celik A, Arslan S,
Terzioglu G, Ozbal S, Yildiz S, Ergur UB, Guneli E, Camsari T, et
al: Protective effects of taurine against renal
ischemia/reperfusion injury in rats by inhibition of gelatinases,
MMP-2 and MMP-9, and p38 mitogen-activated protein kinase
signaling. Biotech Histochem. 92:524–535. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Zhao H, Dong Y, Tian X, Tan TK, Liu Z,
Zhao Y, Zhang Y, Harris DCh and Zheng G: Matrix metalloproteinases
contribute to kidney fibrosis in chronic kidney diseases. World J
Nephrol. 2:84–89. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Cavdar Z, Ozbal S, Celik A, Ergur BU,
Guneli E, Ural C, Camsari T and Guner GA: The effects of
alpha-lipoic acid on MMP-2 and MMP-9 activities in a rat renal
ischemia and re-perfusion model. Biotech Histochem. 89:304–314.
2014. View Article : Google Scholar
|
|
76
|
Kunugi S, Shimizu A, Kuwahara N, Du X,
Takahashi M, Terasaki Y, Fujita E, Mii A, Nagasaka S, Akimoto T, et
al: Inhibition of matrix metalloproteinases reduces
ischemia-reperfusion acute kidney injury. Lab Invest. 91:170–180.
2011. View Article : Google Scholar
|
|
77
|
Han WK, Waikar SS, Johnson A, Betensky RA,
Dent CL, Devarajan P and Bonventre JV: Urinary biomarkers in the
early diagnosis of acute kidney injury. Kidney Int. 73:863–869.
2008. View Article : Google Scholar
|
|
78
|
Nagase H: Activation mechanisms of matrix
metalloproteinases. Biol Chem. 378:151–160. 1997.PubMed/NCBI
|
|
79
|
Visse R and Nagase H: Matrix
metalloproteinases and tissue inhibitors of metalloproteinases:
Structure, function, and biochemistry. Circ Res. 92:827–839. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
80
|
Nagase H and Woessner JF Jr: Matrix
metalloproteinases. J Biol Chem. 274:21491–21494. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
81
|
Carmeliet P, Moons L, Lijnen R, Baes M,
Lemaître V, Tipping P, Drew A, Eeckhout Y, Shapiro S, Lupu F and
Collen D: Urokinase-generated plasmin activates matrix
metalloproteinases during aneurysm formation. Nat Genet.
17:439–444. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
82
|
Mazzieri R, Masiero L, Zanetta L, Monea S,
Onisto M, Garbisa S and Mignatti P: Control of type IV collagenase
activity by components of the urokinase-plasmin system: A
regulatory mechanism with cell-bound reactants. EMBO J.
16:2319–2332. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
83
|
Emonard HP, Remacle AG, Noël AC, Grimaud
JA, Stetler-Stevenson WG and Foidart JM: Tumor cell
surface-associated binding site for the M(r) 72,000 type IV
collagenase. Cancer Res. 52:5845–5848. 1992.PubMed/NCBI
|
|
84
|
Monsky WL, Kelly T, Lin CY, Yeh Y,
Stetler-Stevenson WG, Mueller SC and Chen WT: Binding and
localization of M(r) 72,000 matrix metalloproteinase at cell
surface invadopodia. Cancer Res. 53:3159–3164. 1993.PubMed/NCBI
|
|
85
|
Aimes RT and Quigley JP: Matrix
metalloproteinase-2 is an interstitial collagenase. Inhibitor-free
enzyme catalyzes the cleavage of collagen fibrils and soluble
native type I collagen generating the specific 3/4- and 1/4-length
fragments. J Biol Chem. 270:5872–5876. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
86
|
Yabluchanskiy A, Ma Y, Iyer RP, Hall ME
and Lindsey ML: Matrix metalloproteinase-9: Many shades of function
in cardiovascular disease. Physiology (Bethesda). 28:391–403.
2013.PubMed/NCBI
|
|
87
|
Fu Z, Ye Q, Zhang Y, Zhong Z, Xiong Y,
Wang Y, Hu L, Wang W, Huang W and Ko DS: Hypothermic machine
perfusion reduced inflammatory reaction by downregulating the
expression of matrix metalloproteinase 9 in a reperfusion model of
donation after cardiac death. Artif Organs. 40:E102–E111. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
88
|
Sulikowski T, Domanski L, Zietek Z, Adler
G, Pawlik A, Ciechanowicz A, Ciechanowski K and Ostrowski M: Effect
of preservation solutions UW and EC on the expression of matrix
metalloproteinase II and tissue inhibitor of metalloproteinase II
genes in rat kidney. Postepy Hig Med Dosw (Online). 66:45–50.
2012.PubMed/NCBI
|
|
89
|
Ichimura T, Bonventre JV, Bailly V, Wei H,
Hession CA, Cate RL and Sanicola M: Kidney injury molecule-1
(KIM-1), a putative epithelial cell adhesion molecule containing a
novel immunoglobulin domain, is up-regulated in renal cells after
injury. J Biol Chem. 273:4135–4142. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
90
|
Khan KNM, Hard GC and Alden CL: Chapter
47-Kidney. Haschek and Rousseaux's Handbook of Toxicologic
Pathology. 3rd. Haschek WM, Rousseaux CG and Wallig MA: Academic
Press; Boston: pp. 1667–1773. 2013, View Article : Google Scholar
|
|
91
|
van Timmeren MM, van den Heuvel MC, Bailly
V, Bakker SJL, van Goor H and Stegeman CA: Tubular kidney injury
molecule-1 (KIM-1) in human renal disease. J Pathol. 212:209–217.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
92
|
Amin RP, Vickers AE, Sistare F, Thompson
KL, Roman RJ, Lawton M, Kramer J, Hamadeh HK, Collins J, Grissom S,
et al: Identification of putative gene based markers of renal
toxicity. Environ Health Perspect. 112:465–479. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
93
|
Griffin BR, You Z, Noureddine L, Gitomer
B, Perrenoud L, Wang W, Chonchol M and Jalal D; HALT Investigators:
KIM-1 and kidney disease progression in autosomal dominant
polycystic kidney disease: HALT-PKD results. Am J Nephrol.
51:473–479. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
94
|
Han WK, Alinani A, Wu CL, Michaelson D,
Loda M, McGovern FJ, Thadhani R and Bonventre JV: Human kidney
injury molecule-1 is a tissue and urinary tumor marker of renal
cell carcinoma. J Am Soc Nephrol. 16:1126–1134. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
95
|
Bonventre JV: Kidney injury molecule-1
(KIM-1): A urinary biomarker and much more. Nephrol Dial
Transplant. 24:3265–3268. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
96
|
Ichimura T, Asseldonk EJPV, Humphreys BD,
Gunaratnam L, Duffield JS and Bonventre JV: Kidney injury
molecule-1 is a phosphatidylserine receptor that confers a
phagocytic phenotype on epithelial cells. J Clin Invest.
118:1657–1668. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
97
|
Kaplanski G: Interleukin-18: Biological
properties and role in disease pathogenesis. Immunol Rev.
281:138–153. 2018. View Article : Google Scholar
|
|
98
|
Ghayur T, Banerjee S, Hugunin M, Butler D,
Herzog L, Carter A, Quintal L, Sekut L, Talanian R, Paskind M, et
al: Caspase-1 processes IFN-gamma-inducing factor and regulates
LPS-induced IFN-gamma production. Nature. 386:619–623. 1997.
View Article : Google Scholar : PubMed/NCBI
|
|
99
|
Sugawara S, Uehara A, Nochi T, Yamaguchi
T, Ueda H, Sugiyama A, Hanzawa K, Kumagai K, Okamura H and Takada
H: Neutrophil proteinase 3-mediated induction of bioactive IL-18
secretion by human oral epithelial cells. J Immunol. 167:6568–6575.
2001. View Article : Google Scholar : PubMed/NCBI
|
|
100
|
Ihim SA, Abubakar SD, Zian Z, Sasaki T,
Saffarioun M, Maleknia S and Azizi G: Interleukin-18 cytokine in
immunity, inflammation, and autoimmunity: Biological role in
induction, regulation, and treatment. Front Immunol. 13:9199732022.
View Article : Google Scholar : PubMed/NCBI
|
|
101
|
Banerjee S and Bond JS: Prointerleukin-18
is activated by meprin beta in vitro and in vivo in intestinal
inflammation. J Biol Chem. 283:31371–31377. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
102
|
Okamura H, Tsutsui H, Kashiwamura S,
Yoshimoto T and Nakanishi K: Interleukin-18: A novel cytokine that
augments both innate and acquired immunity. Adv Immunol.
70:281–312. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
103
|
Tsutsui H, Nakanishi K, Matsui K,
Higashino K, Okamura H, Miyazawa Y and Kaneda K: IFN-gamma-inducing
factor up-regulates Fas ligand-mediated cytotoxic activity of
murine natural killer cell clones. J Immunol. 157:3967–3973. 1996.
View Article : Google Scholar : PubMed/NCBI
|
|
104
|
Li P, Li YL, Li ZY, Wu YN, Zhang CC, A X,
Wang CX, Shi HT, Hui MZ, Xie B, et al: Cross talk between vascular
smooth muscle cells and monocytes through
interleukin-1β/interleukin-18 signaling promotes vein graft
thickening. Arterioscler Thromb Vasc Biol. 34:2001–2011. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
105
|
Hoshino T, Wiltrout RH and Young HA: IL-18
is a potent coinducer of IL-13 in NK and T cells: A new potential
role for IL-18 in modulating the immune response. J Immunol.
162:5070–5077. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
106
|
Yoshimoto T, Mizutani H, Tsutsui H,
Noben-Trauth N, Yamanaka K, Tanaka M, Izumi S, Okamura H, Paul WE
and Nakanishi K: IL-18 induction of IgE: Dependence on CD4+ T
cells, IL-4 and STAT6. Nat Immunol. 1:132–137. 2000. View Article : Google Scholar
|
|
107
|
Yoshimoto T, Tsutsui H, Tominaga K,
Hoshino K, Okamura H, Akira S, Paul WE and Nakanishi K: IL-18,
although antiallergic when administered with IL-12, stimulates IL-4
and histamine release by basophils. Proc Natl Acad Sci USA.
96:13962–13966. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
108
|
Doshi MD, Reese PP, Hall IE, Schröppel B,
Ficek J, Formica RN, Weng FL, Hasz RD, Thiessen-Philbrook H and
Parikh CR: Utility of applying quality assessment tools for kidneys
with KDPI ≥80. Transplantation. 101:1125–1133. 2017. View Article : Google Scholar
|
|
109
|
Issaq H and Veenstra T: Two-dimensional
polyacrylamide gel electrophoresis (2D-PAGE): Advances and
perspectives. Biotechniques. 44:697–698. 7002008. View Article : Google Scholar : PubMed/NCBI
|
|
110
|
Smith BJ: SDS polyacrylamide gel
electrophoresis of proteins. Methods Mol Biol. 32:23–34.
1994.PubMed/NCBI
|
|
111
|
Kielkopf CL, Bauer W and Urbatsch IL:
Sodium dodecyl sulfate-polyacrylamide gel electrophoresis of
proteins. Cold Spring Harb Protoc. 20212021.
|
|
112
|
Pitt JJ: Principles and applications of
liquid chromatography-mass spectrometry in clinical biochemistry.
Clin Biochem Rev. 30:19–34. 2009.PubMed/NCBI
|
|
113
|
Aslam B, Basit M, Nisar MA, Khurshid M and
Rasool MH: Proteomics: Technologies and their applications. J
Chromatogr Sci. 55:182–196. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
114
|
Li C, Chu S, Tan S, Yin X, Jiang Y, Dai X,
Gong X, Fang X and Tian D: Towards higher sensitivity of mass
spectrometry: A perspective from the mass analyzers. Front Chem.
9:8133592021. View Article : Google Scholar
|
|
115
|
Wieser A, Schneider L, Jung J and Schubert
S: MALDI-TOF MS in microbiological diagnostics-identification of
microorganisms and beyond (mini review). Appl Microbiol Biotechnol.
93:965–974. 2012. View Article : Google Scholar
|
|
116
|
van Leeuwen LL, Spraakman NA, Brat A,
Huang H, Thorne AM, Bonham S, van Balkom BWM, Ploeg RJ, Kessler BM
and Leuvenink HGD: Proteomic analysis of machine perfusion solution
from brain dead donor kidneys reveals that elevated complement,
cytoskeleton and lipid metabolism proteins are associated with
1-year outcome. Transpl Int. 34:1618–1629. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
117
|
Mulvey JF, Ul Shaheed S, Charles PD,
Snashall C, Lo Faro ML, Sutton CW, Jochmans I, Pirenne J, van
Kooten C, Leuvenink HGD, et al: Perfusate proteomes provide
biological insight into oxygenated versus standard hypothermic
machine perfusion in kidney transplantation. Ann Surg. 278:676–682.
2023.PubMed/NCBI
|
|
118
|
Karpman D, Bekassy Z, Grunenwald A and
Roumenina LT: A role for complement blockade in kidney
transplantation. Cell Mol Immunol. 19:755–757. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
119
|
Yamanaka K, Kakuta Y, Miyagawa S, Nakazawa
S, Kato T, Abe T, Imamura R, Okumi M, Maeda A, Okuyama H, et al:
Depression of complement regulatory factors in rat and human renal
grafts is associated with the progress of acute T-cell mediated
rejection. PLoS One. 11:e01488812016. View Article : Google Scholar : PubMed/NCBI
|
|
120
|
De Vries B, Matthijsen RA, Wolfs TGAM, Van
Bijnen AAJHM, Heeringa P and Buurman WA: Inhibition of complement
factor C5 protects against renal ischemia-reperfusion injury:
Inhibition of late apoptosis and inflammation. Transplantation.
75:375–382. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
121
|
Biglarnia AR, Huber-Lang M, Mohlin C,
Ekdahl KN and Nilsson B: The multifaceted role of complement in
kidney transplantation. Nat Rev Nephrol. 14:767–781. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
122
|
Zhang R: Donor-specific antibodies in
kidney transplant recipients. Clin J Am Soc Nephrol. 13:182–192.
2018. View Article : Google Scholar :
|
|
123
|
Nauser CL, Farrar CA and Sacks SH:
Complement recognition pathways in renal transplantation. J Am Soc
Nephrol. 28:2571–2578. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
124
|
Sato T, Van Dixhoorn MG, Prins FA, Mooney
A, Verhagen N, Muizert Y, Savill J, Van Es LA and Daha MR: The
terminal sequence of complement plays an essential role in
antibody-mediated renal cell apoptosis. J Am Soc Nephrol.
10:1242–1252. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
125
|
Shimizu A, Masuda Y, Kitamura H, Ishizaki
M, Ohashi R, Sugisaki Y and Yamanaka N: Complement-mediated killing
of mesangial cells in experimental glomerulonephritis: Cell death
by a combination of apoptosis and necrosis. Nephron. 86:152–160.
2000. View Article : Google Scholar : PubMed/NCBI
|
|
126
|
Zhou W, Farrar CA, Abe K, Pratt JR, Marsh
JE, Wang Y, Stahl GL and Sacks SH: Predominant role for C5b-9 in
renal ischemia/reperfusion injury. J Clin Invest. 105:1363–1371.
2000. View Article : Google Scholar : PubMed/NCBI
|
|
127
|
Jane-wit D, Surovtseva YV, Qin L, Li G,
Liu R, Clark P, Manes TD, Wang C, Kashgarian M, Kirkiles-Smith NC,
et al: Complement membrane attack complexes activate noncanonical
NF-κB by forming an Akt+ NIK+ signalosome on Rab5+ endosomes. Proc
Natl Acad Sci USA. 112:9686–9691. 2015. View Article : Google Scholar
|
|
128
|
Jager NM, Venema LH, Arykbaeva AS,
Meter-Arkema AH, Ottens PJ, van Kooten C, Mollnes TE, Alwayn IPJ,
Leuvenink HGD and Pischke SE; PROPER study consortium: Complement
is activated during normothermic machine perfusion of porcine and
human discarded kidneys. Front Immunol. 13:8313712022. View Article : Google Scholar : PubMed/NCBI
|
|
129
|
Coskun A, Baykal AT, Kazan D, Akgoz M,
Senal MO, Berber I, Titiz I, Bilsel G, Kilercik H, Karaosmanoglu K,
et al: Proteomic analysis of kidney preservation solutions prior to
renal transplantation. PLoS One. 11:e01687552016. View Article : Google Scholar : PubMed/NCBI
|
|
130
|
Nieuwenhuijs-Moeke GJ, Pischke SE, Berger
SP, Sanders JSF, Pol RA, Struys MMRF, Ploeg RJ and Leuvenink HGD:
Ischemia and reperfusion injury in kidney transplantation: Relevant
mechanisms in injury and repair. J Clin Med. 9:2532020. View Article : Google Scholar : PubMed/NCBI
|
|
131
|
Kako K, Kato M, Matsuoka T and Mustapha A:
Depression of membrane-bound Na+-K+-ATPase activity induced by free
radicals and by ischemia of kidney. Am J Physiol. 254:C330–C337.
1988. View Article : Google Scholar : PubMed/NCBI
|
|
132
|
Salvadori M, Rosso G and Bertoni E: Update
on ischemia-reperfusion injury in kidney transplantation:
Pathogenesis and treatment. World J Transplant. 5:52–67. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
133
|
Molitoris BA, Dahl R and Geerdes A:
Cytoskeleton disruption and apical redistribution of proximal
tubule Na(+)-K(+)-ATPase during ischemia. Am J Physiol.
263:F488–F495. 1992.PubMed/NCBI
|
|
134
|
Caron A, Desrosiers RR and Béliveau R:
Kidney ischemia-reperfusion regulates expression and distribution
of tubulin subunits, beta-actin and rho GTPases in proximal
tubules. Arch Biochem Biophys. 431:31–46. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
135
|
Genescà M, Sola A and Hotter G: Actin
cytoskeleton derangement induces apoptosis in renal
ischemia/reperfusion. Apoptosis. 11:563–571. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
136
|
Sanz AB, Sanchez-Niño MD, Ramos AM and
Ortiz A: Regulated cell death pathways in kidney disease. Nat Rev
Nephrol. 19:281–299. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
137
|
Garrod D and Chidgey M: Desmosome
structure, composition and function. Biochim Biophys Acta.
1778:572–587. 2008. View Article : Google Scholar
|
|
138
|
Chen CS and Zhu H: Protein microarrays.
Biotechniques. 40:423–425, 427 passim. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
139
|
Uttamchandani M, Neo JL, Ong BNZ and
Moochhala S: Applications of microarrays in pathogen detection and
biodefence. Trends Biotechnol. 27:53–61. 2009. View Article : Google Scholar
|
|
140
|
Shome M and Labaer J: Protein microarrays
and their fabrication. Methods Mol Biol. 2597:131–142. 2023.
View Article : Google Scholar
|
|
141
|
Baboudjian M, Gondran-Tellier B, Boissier
R, Ancel P, Marjollet J, Lyonnet L, François P, Sabatier F,
Lechevallier E, Dutour A and Paul P: An enhanced level of VCAM in
transplant preservation fluid is an independent predictor of early
kidney allograft dysfunction. Front Immunol. 13:9669512022.
View Article : Google Scholar : PubMed/NCBI
|
|
142
|
Gok MA, Pelzers M, Glatz JFC, Shenton BK,
Buckley PE, Peaston R, Cornell C, Mantle D, Soomro N, Jaques BC, et
al: Do tissue damage biomarkers used to assess machine-perfused
NHBD kidneys predict long-term renal function post-transplant? Clin
Chim Acta. 338:33–43. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
143
|
Balupuri S, Talbot D, El-Sheikh M, Snowden
C, Manas DM, Kirby J and Mantle D: Comparison of proteolytic
enzymes and glutathione S-transferase levels in non-heart-beating
donors' (NHBD) kidney perfusates. Clin Chem Lab Med. 38:1099–1102.
2000. View Article : Google Scholar
|
|
144
|
Tejchman K, Sierocka A, Kotfis K, Kotowski
M, Dolegowska B, Ostrowski M and Sienko J: Assessment of oxidative
stress markers in hypothermic preservation of transplanted kidneys.
Antioxidants (Basel). 10:12632021. View Article : Google Scholar : PubMed/NCBI
|
|
145
|
Cohen J, Ratigan E, Shigeoka A, Steiner R,
Stocks L and McKay D: Inflammatory profiling of hypothermic machine
pumped kidney allografts. Am J Transplant. 15(Suppl 3):
C2732015.
|
|
146
|
Maritan E, Franchin M, Amico F, Ietto G,
Villa F, Tozzi M, Ferraro S, Negri S, Alberio MG and Carcano G:
Ischemia and reperfusion injury markers in kidney transplant:
Mechanical perfusion vs cold storage. Preliminary experience: 536.
Transplantation. 94(10S): S11592012. View Article : Google Scholar
|
|
147
|
Boenink R, Astley ME, Huijben JA, Stel VS,
Kerschbaum J, Ots-Rosenberg M, Åsberg AA, Lopot F, Golan E, Castro
de la Nuez P, et al: The ERA registry annual report 2019: Summary
and age comparisons. Clin Kidney J. 15:452–472. 2021. View Article : Google Scholar
|
|
148
|
Aubert O, Kamar N, Vernerey D, Viglietti
D, Martinez F, Duong-Van-Huyen JP, Eladari D, Empana JP, Rabant M,
Verine J, et al: Long term outcomes of transplantation using
kidneys from expanded criteria donors: Prospective, population
based cohort study. BMJ. 351:h35572015. View Article : Google Scholar : PubMed/NCBI
|
|
149
|
Tomita Y, Tojimbara T, Iwadoh K, Nakajima
I and Fuchinoue S: Long-term outcomes in kidney transplantation
from expanded-criteria donors after circulatory death. Transplant
Proc. 49:45–48. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
150
|
Borda B, Németh T, Ottlakan A, Keresztes
C, Kemény É and Lázár G: Post-transplantation morphological and
functional changes in kidneys from expanded criteria donors.
Physiol Int. 104:329–333. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
151
|
Tingle SJ, Figueiredo RS, Moir JA,
Goodfellow M, Talbot D and Wilson CH: Machine perfusion
preservation versus static cold storage for deceased donor kidney
transplantation. Cochrane Database Syst Rev.
3:CD0116712019.PubMed/NCBI
|
|
152
|
Tingle SJ, Thompson ER, Figueiredo RS,
Moir JA, Goodfellow M, Talbot D and Wilson CH: Normothermic and
hypothermic machine perfusion preservation versus static cold
storage for deceased donor kidney transplantation. Cochrane
Database Syst Rev. 7:CD0116712024.PubMed/NCBI
|
|
153
|
Helanterä I, Ibrahim HN, Lempinen M and
Finne P: Donor age, cold ischemia time, and delayed graft function.
Clin J Am Soc Nephrol. 15:813–821. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
154
|
Heilman RL, Mathur A, Smith ML, Kaplan B
and Reddy KS: Increasing the use of kidneys from unconventional and
high-risk deceased donors. Am J Transplant. 16:3086–3092. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
155
|
Tasaki M, Saito K, Nakagawa Y, Ikeda M,
Imai N, Narita I and Takahashi K: Effect of donor-recipient age
difference on long-term graft survival in living kidney
transplantation. Int Urol Nephrol. 46:1441–1446. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
156
|
Lim K, Lee YJ, Gwon JG, Jung CW, Yang J,
Oh SW, Jo SK, Cho WY and Kim MG: Impact of donor age on the
outcomes of kidney transplantation from deceased donors with
histologic acute kidney injury. Transplant Proc. 51:2593–2597.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
157
|
Scurt FG, Ernst A, Hammoud B, Wassermann
T, Mertens PR, Schwarz A, Becker JU and Chatzikyrkou C: Effect of
creatinine metrics on outcome after transplantation of marginal
donor kidneys. Nephrology (Carlton). 27:973–982. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
158
|
Irish GL, Coates PT and Clayton PA:
Association of admission, nadir, and terminal donor creatinine with
kidney transplantation outcomes. Kidney Int Rep. 6:2075–2083. 2021.
View Article : Google Scholar : PubMed/NCBI
|
|
159
|
Perico N, Cattaneo D, Sayegh MH and
Remuzzi G: Delayed graft function in kidney transplantation.
Lancet. 364:1814–1827. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
160
|
Lai C, Yee SY, Ying T and Chadban S:
Biomarkers as diagnostic tests for delayed graft function in kidney
transplantation. Transpl Int. 34:2431–2441. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
161
|
Siedlecki A, Irish W and Brennan DC:
Delayed graft function in the kidney transplant. Am J Transplant.
11:2279–2296. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
162
|
Schrezenmeier E, Müller M, Friedersdorff
F, Khadzhynov D, Halleck F, Staeck O, Dürr M, Zhang K, Eckardt KU,
Budde K and Lehner LJ: Evaluation of severity of delayed graft
function in kidney transplant recipients. Nephrol Dial Transplant.
37:973–981. 2022. View Article : Google Scholar
|
|
163
|
Parsons FM: Haemodialysis; indications and
results. Postgrad Med J. 35:625–630, passim. 1959. View Article : Google Scholar : PubMed/NCBI
|
|
164
|
Clark JE and Soricelli RR: Indications for
dialysis. Med Clin North Am. 49:1213–1239. 1965. View Article : Google Scholar : PubMed/NCBI
|
|
165
|
Hosgood SA, Callaghan CJ, Wilson CH, Smith
L, Mullings J, Mehew J, Oniscu GC, Phillips BL, Bates L and
Nicholson ML: Normothermic machine perfusion versus static cold
storage in donation after circulatory death kidney transplantation:
A randomized controlled trial. Nat Med. 29:1511–1519. 2023.
View Article : Google Scholar : PubMed/NCBI
|
|
166
|
Brat A, de Vries KM, van Heurn EWE,
Huurman VAL, de Jongh W, Leuvenink HGD, van Zuilen AD,
Haase-Kromwijk BJJM, de Jonge J, Berger SP and Hofker SH:
Hypothermic machine perfusion as a national standard preservation
method for deceased donor kidneys. Transplantation. 106:1043–1050.
2022. View Article : Google Scholar :
|
|
167
|
Rijkse E, Bouari S, Kimenai HJAN, de Jonge
J, de Bruin RWF, Slagter JS, van den Hoogen MWF, Ijzermans JNM,
Hoogduijn MJ and Minnee RC: Additional Normothermic machine
perfusion versus hypothermic machine perfusion in suboptimal donor
kidney transplantation: Protocol of a randomized, controlled,
open-label trial. Int J Surg Protoc. 25:227–237. 2021. View Article : Google Scholar : PubMed/NCBI
|