Introduction
Breast cancer is the most prevalent form of cancer
diagnosed in women, and there continues to be limited drug
treatment options for the ∼30% of patients whose cancer lacks the
estrogen receptor (ER) (1,2). ER-negative breast cancer can either
express the human epidermal growth factor receptor 2 (Her2, erbb2)
or lack this receptor. Triple negative breast cancer (TNBC) is
defined by a lack of the expression of the ER, progesterone
receptor and HER2. They account for 10–17% of all breast cancers
and represent 85% of the basal-like subtype (3), one of the five subgroups of breast
cancer categorized in recent years (4). TNBC has an aggressive clinical
behavior and is generally associated with poor prognosis, thus
chemotherapy remains the only systemic treatment option available
for these patients (5).
In the search for effective drugs for ER-negative
breast cancer, several lead compounds from natural products such as
curcumin (diferuloylmethane), the primary bioactive compound
isolated from the rhizome of turmeric (Curcuma longa Linn.),
have emerged. Curcumin has numerous pharmacological,
chemopreventative and chemotherapeutic actions, and in vivo
studies have demonstrated decreased tumorigenesis of many organs,
including the mammary gland (6–11).
In vitro studies have also demonstrated that curcumin
exhibits potent cytoxicity toward numerous cell lines including
ER-negative human breast cancer cells (12–18).
However, curcumin has shown limited clinical efficacy, due to its
low bioavailability and low stability in physiological media
(7). Therefore, research groups
have concentrated on the synthesis and characterization of curcumin
analogs. Cyclohexanone analogs of curcumin as a group have shown
enhanced activity and stability in biological medium compared to
curcumin (19). Specifically, the
cyclohexanone derivative
2,6-bis((3-methoxy-4-hydroxyphenyl)methylene)-cyclohexanone (BMHPC)
was cytotoxic towards ER-negative breast cancer cells
(IC50 of 5.0 μM) (20), although bioavailability and in
vivo efficacy were still problematic. More recently fluorinated
cyclohexanone derivatives have not only shown potent cytotoxicity
toward MDA-MB-231 cells (IC50 value of 0.8 μM)
(21,22), but also induced cell cycle arrest
and apoptosis in both breast and prostate cancer cells (22).
Our laboratory has been involved in the search for
new drug treatments for ER-negative breast cancer and uses the
aggressive triple negative MDA-MB-231 human breast cancer cells,
amongst others, to identify new potent drugs. Recently we have
performed structure activity relationships to determine the
cytotoxic potency of 2nd generation heterocyclic cyclohexanone
curcumin analogs. This study demonstrated that
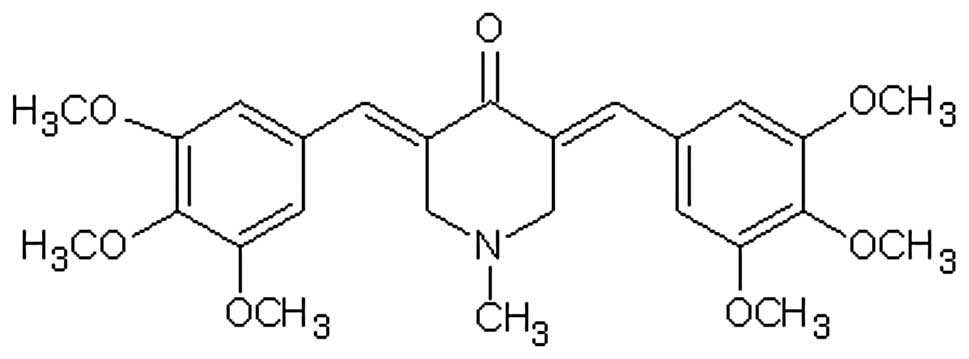
3,5-bis(3,4,5-trimethoxybenzylidene)-1-methylpiperidine-4-one
(RL71)(Fig. 1) showed the lowest
IC50 value (0.3 μM) toward MDA-MB-231 and
MDA-MB-468 breast cancer cells (23). It also elicited the strongest
apoptotic response, as ∼40% of MDA-MB-231 cells underwent apoptosis
18 h following RL71 treatment (1 μM) (23). Therefore, this study was designed
to comprehensively investigate the in vitro activity of this
lead compound in various ER-negative breast cancer cell lines in
order to determine its potential to be developed into a drug for
aggressive breast cancer that lacks the ER.
Materials and methods
Materials
HUVEC, MDA-MB-231, MDA-MB-468 and SKBr3 cells were
purchased from American Type Culture Collection (Manassas, VA).
Primary antibodies to p38, pp38, NF-κB, JNK, pJNK, cleaved
caspase-3, 4EBP1, p4EBP1, p27, mTOR, pmTOR, HER2, pHER2 and β-actin
were purchased from Cell Signaling Technology (Danvers, MA). Akt
and pAkt primary antibodies were purchased from BD Biosciences
(Auckland, NZ). Dulbecco’s modified Eagle’s medium (DMEM) nutrient
mixture Ham’s F-12, sulforhodamine B salt, propidium iodide (PI),
ammonium persulfate, horseradish peroxidase were purchased from
Sigma Aldrich (Auckland, NZ). Acrylamide, bisacrylamide, sodium
dodecylsulfate and PVDF membrane were purchased from Bio-Rad
Laboratories (Hercules, CA). Complete mini EDTA-free protease
inhibitor cocktail and Annexin-V-FLUOS were purchased from Roche
Diagnostics Corporation (Mannheim, Germany). RL71 was prepared as
described previously (18). All
other chemicals were of the highest purity commercially
available.
Cell maintenance
MDA-MB-231, MDA-MB-468 and SkBr3 cells were
maintained in complete growth media composed of DMEM/Ham’s F12
supplemented with 5% fetal bovine serum, 2 mM L-glutamine, 100
units/ml streptomycin, 250 ng/ml amphotericin B, and 100 units/ml
penicillin and 2.2 g/l NaHCO3.
Cytotoxicity
MDA-MB-231, MDA-MB-468 and SKBr3 cells
(95×104 cells/well) were seeded in 12-well plates in 1
ml DMEM/HamF12 supplemented with 5% FBS, 100 U/ml penicillin, 100
μg/ml streptomycin, 25 ng/ml amphotericin B and 2.2 g/l
NaHCO3 and incubated for 24 h at 37°C. For time course
study, cells were treated with RL71 (1 μM) for 6, 12, 24,
36, 48 and 72 h. Vehicle control cells were treated with DMSO
(0.1%). Cell number in each well was determined using the
sulforhodamine B (SRB) assay (24).
Cell cycle analysis
Flow cytometry was used to analyze DNA content in
order to determine cell cycle distribution. MDA-MB-231, MDA-MB-468
and SKBr3 cells were plated and treated with RL71 (1 μM) or
0.1% DMSO as control for 6–48 h in 6-well plates. The cells were
harvested, washed with PBS and then fixed in 70% ethanol. Following
rehydration with PBS, the cells were stained with PI in the dark at
4°C as described (25). The
samples were analyzed via flow cytometry using a FACScaliber flow
cytometer (Becton-Dickinson). The percentage of cells in each phase
of cell cycle was determined using Cell Quest Pro software. Results
are expressed as percent of cells in each phase of the cell
cycle.
Induction of apoptosis
MDA-MB-468, and SKBr3 cells were seeded in 6-well
culture plate in 2 ml of DMEM/HamF12 supplemented with 5% FBS, 100
U/ml penicillin, 100 μl/ml streptomycin, 25 ng/ml
amphotericin B and 2.2 g/l NaHCO3. The cells were
treated with RL71 (1 μM) or vehicle control for 12–48 h.
Apoptosis was assessed using Annexin-V-FLUCOS/PI staining, as
described (26). The samples were
analyzed using a FACScaliber flow cytometer (Becton-Dickinson) and
the proportion of apoptotic cells was determined using CellQuest
Pro software.
Preparation of cell lysates
MDA-MB-231, MDA-MB-468 and SKBr3 cells were seeded
in 10 cm culture dishes at 2.5×106 cells per well in 10
ml of DMEM/HamF12 supplemented with 5% FBS, 100 U/ml penicillin,
100 μg/ml streptomycin, 25 ng/ml amphotericin B and 2.2 g/l
NaHCO3. Cells were treated with RL71 (1 μM) or
vehicle control for 0–36 h. At the end of treatment, whole cell
lysates were prepared and protein concentration of the lysates was
determined using the bicinchoninic acid (BCA) method (25).
Western blot analysis
Cell lysates were resolved by SDS-PAGE (40 μg
protein per well) and then the proteins were transferred to a PVDF
membrane. Protein levels were analyzed with the desired primary
antibodies, followed by horseradish peroxidase-conjugated secondary
antibodies (Bio-Rad). The digital chemiluminescence images were
taken by a Versadoc densitometer (Bio-Rad).
Transwell migration
Transwell migration was performed using 24 well
plates containing BioCoat™ Matrigel™ Invasion Chamber inserts (BD
Biosciences, Bedford, MA). HUVEC cells (50,000/well) were plated on
rehydrated Matrigel coated culture inserts. The bottom chamber
contained 500 μl of EGM serum free media. The cells were
treated with 0.1% DMSO or RL71 (1 μM) and incubated for 18 h
at 37°C in a humidified 5% CO2 incubator. After
incubation, all contents from well inserts were aspirated and
non-migrated cells were removed with a cotton swab. Migrated cells
on the bottom of the filters were stained with DiffQuick solution
for 1 min and excess stain was washed with water and dried. Cells
on the filters were counted using a Zeiss Axioplan camera and
compared to the control well insert that contained no Matrigel.
Results are expressed as migrated cells as a percent of total cell
population.
Endothelial tube formation
The day before performing the tube formation assay,
a Matrigel matrix was incubated on ice overnight. On the day of the
assay, 125 μl Geltrex Matrigel was transferred into wells of
a 24-well plate. The plate was incubated at 37°C, 5% CO2
for 30 min. HUVEC cells (5×104/well) were then loaded
into each well, followed by addition of DMSO (0.1%) or RL71 (1
μM). The plate was incubated at 37°C, 5% CO2 for
18 h and photographs (200×) were taken by an individual blinded to
the treatment groups.
Animals and housing
Female CD-1 mice (6-weeks old) were purchased from
the Hercus Taieri Resource Unit (Dunedin, NZ). All procedures were
approved by the University of Otago (AEC# 91/07). Mice were housed
in pathogen-free conditions with woodchip bedding with access to
food (Reliance rodent diet, Dunedin, NZ) and water ad
libitum. Mice were housed in a 21–24°C environment on a
scheduled 12 h light/dark cycle and acclimatized for 3 days prior
to experimentation.
Oral bioavailability of RL71
Mice (3/group) were orally gavaged with RL71 and
blood samples were collected at the following time points 0, 5 min,
10 min, 15 min, 30 min, 1 h, 1.5 h and 2 h). Plasma was separated
and stored at −20°C. The samples for analysis were prepared by
addition of methanol to precipitate the proteins, followed by
sonication and filtration. The samples were analyzed by HPLC with
UV-DAD detection. HPLC analysis was performed using an Agilent
HP1100 system at 25°C on a C18 column (Phenomenex Gemini-NX) 3
μ (110A, 150×2 mm) with a 2×4 mm C18 guard column. RL71 was
monitored 390 nm. The mobile phase was acetonitrile in water with
0.1% formic acid: t0=30%, t10=70%,
t15=100%, t17=30%, t20=30%. The
flow rate was 0.3 ml/min, with an injection volume of 5
μl.
Statistical analysis
When time was a factor data were analyzed using a
two-way ANOVA coupled with a Bonferroni post-hoc test. Data that
did not have time as a facor were analysed using a one-way ANOVA
coupled with a Bonferroni post-hoc test. Transwell migration data
were analyzed by a Student’s t-test. p<0.05 was the minimal
requirement for a statistically significant difference.
Results
Previously we have shown that treatment for 5 days
with RL71 elicited sub micromolar IC50 values in three
different ER-negative breast cancer cell lines (20). Therefore, the first aim of this
study was to examine the cytotoxicity of RL71 toward MDA-MB-231,
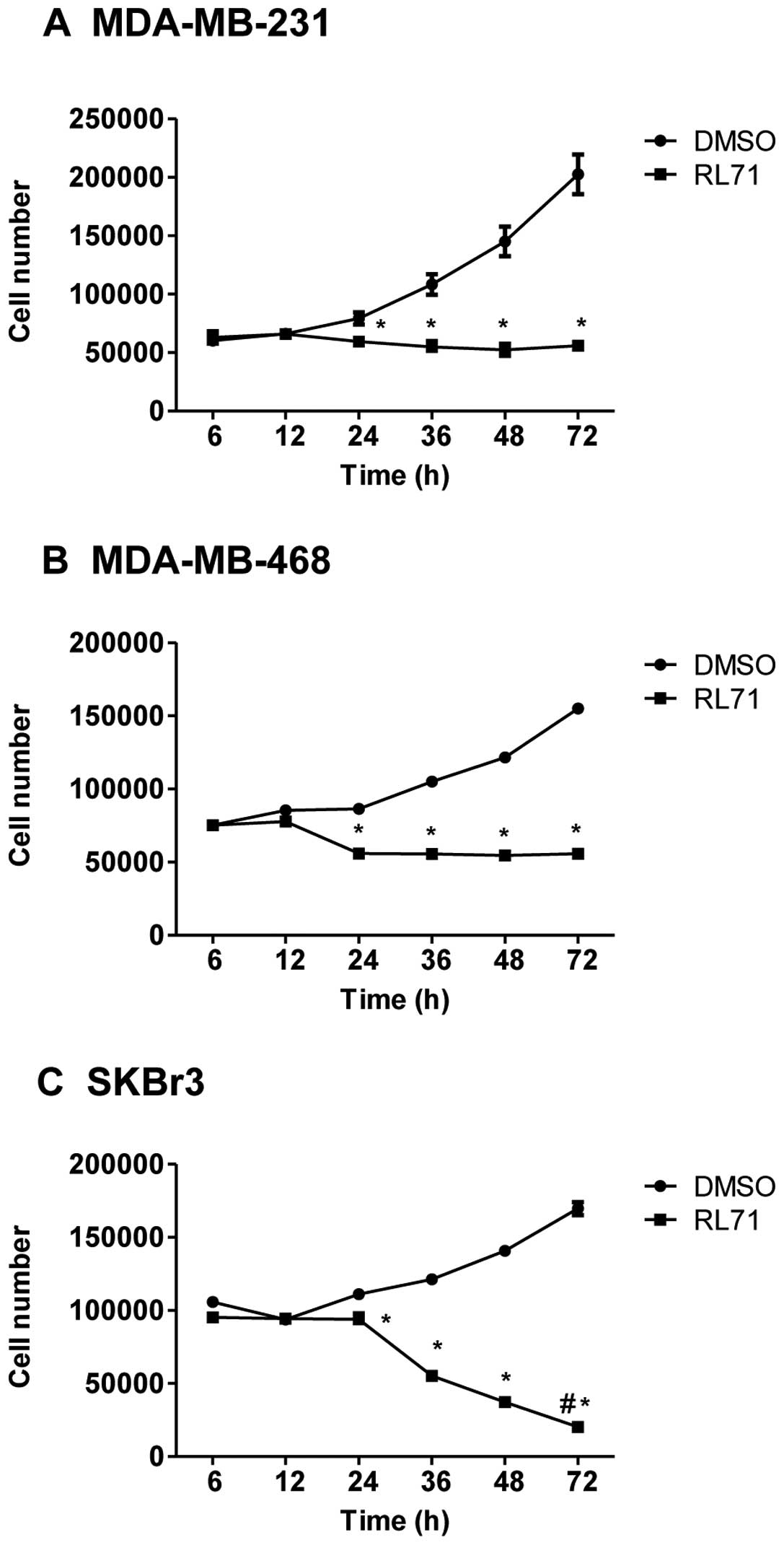
MDA-MB-468 and SKBr3 cells over a time-course. The results showed
that RL71 (1 μM) elicited time-dependent and cell
line-dependent cytotoxicity. Specifically, time-dependent
cytotoxicity was elicited in SKBr3 cells with significantly
increased cytotoxicity at 72 h compared with all other time points
(Fig. 2). However, in the two
triple negative breast cancer (TNBC) cell lines no further
cytotoxicity was elicited after 24 h. Thus, RL71 showed potent
cytotoxicity toward SKBr3 cells compared to a cytostatic effect in
TNBC cells.
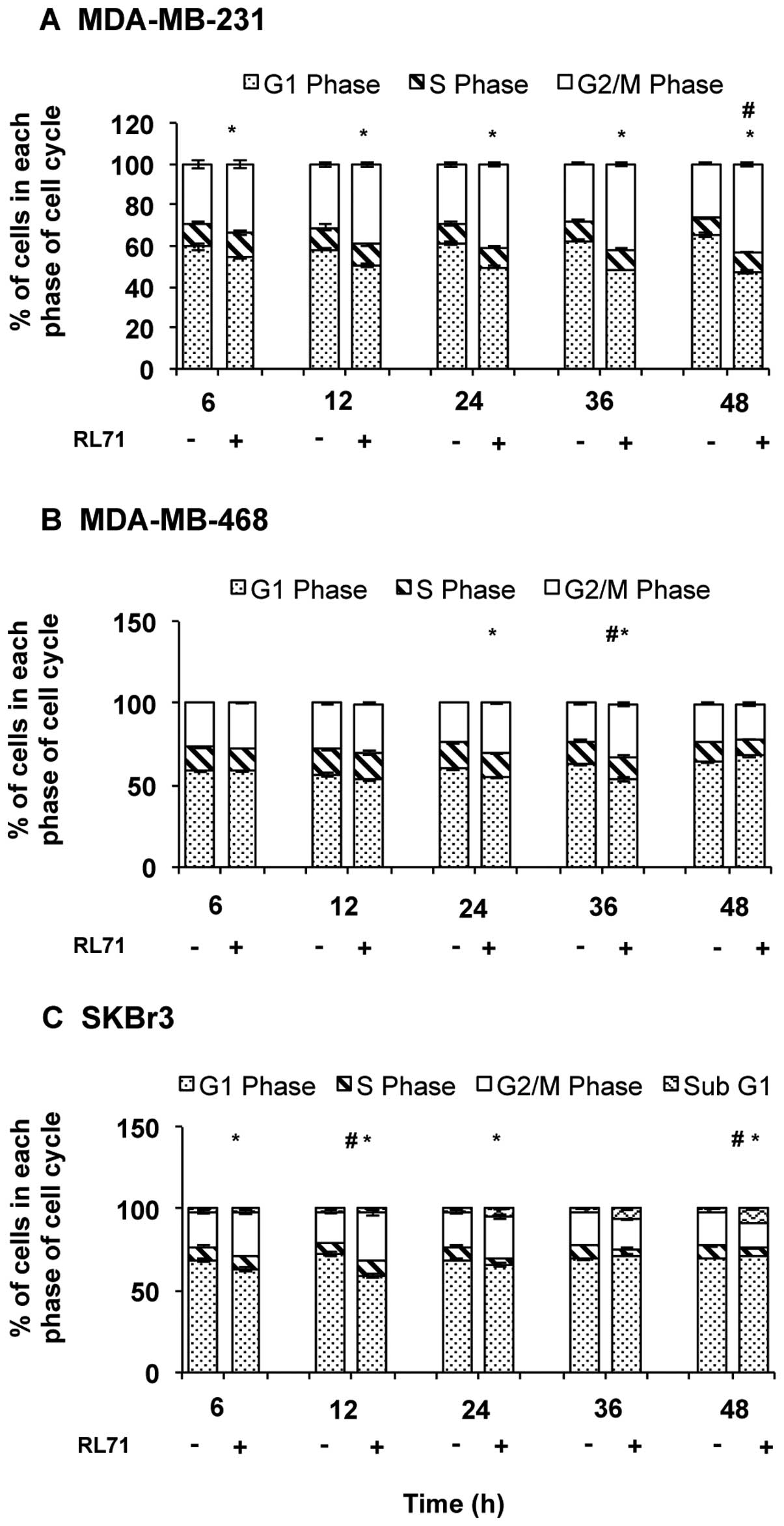
We next examined whether the cytotoxicity of RL71
was due to G2/M phase cell cycle arrest. Treatment of MDA-MB-231,
MDA-MB-468, and SKBr3 cells with RL71 (1 μM) produced G2/M
phase arrest in all three cell lines. Specifically, at 48 h, RL71
caused an 162% increase in the proportion of MDA-MB-231 cells in
G2/M phase over control (Fig. 3A),
while in MDA-MB-468 cells, the proportion of cells in G2/M phase
increased by 140% over control at 36 h (Fig. 3B). In SKBr3 cells, after 12 h, the
proportion of cells undergoing G2/M phase was increased by 153%
over control (Fig. 3C). Moreover,
there was a significant reduction in the proportion of cells in S
phase at 24, 36 and 48 h. SKBr3 cells were the only cell type to
show an increase in subG1 cells. The effect in MDA-MB-231 cells was
time-dependent as the number of cells undergoing G2/M phase arrest
was signficantly increased at 48 h compared to all other time
points.
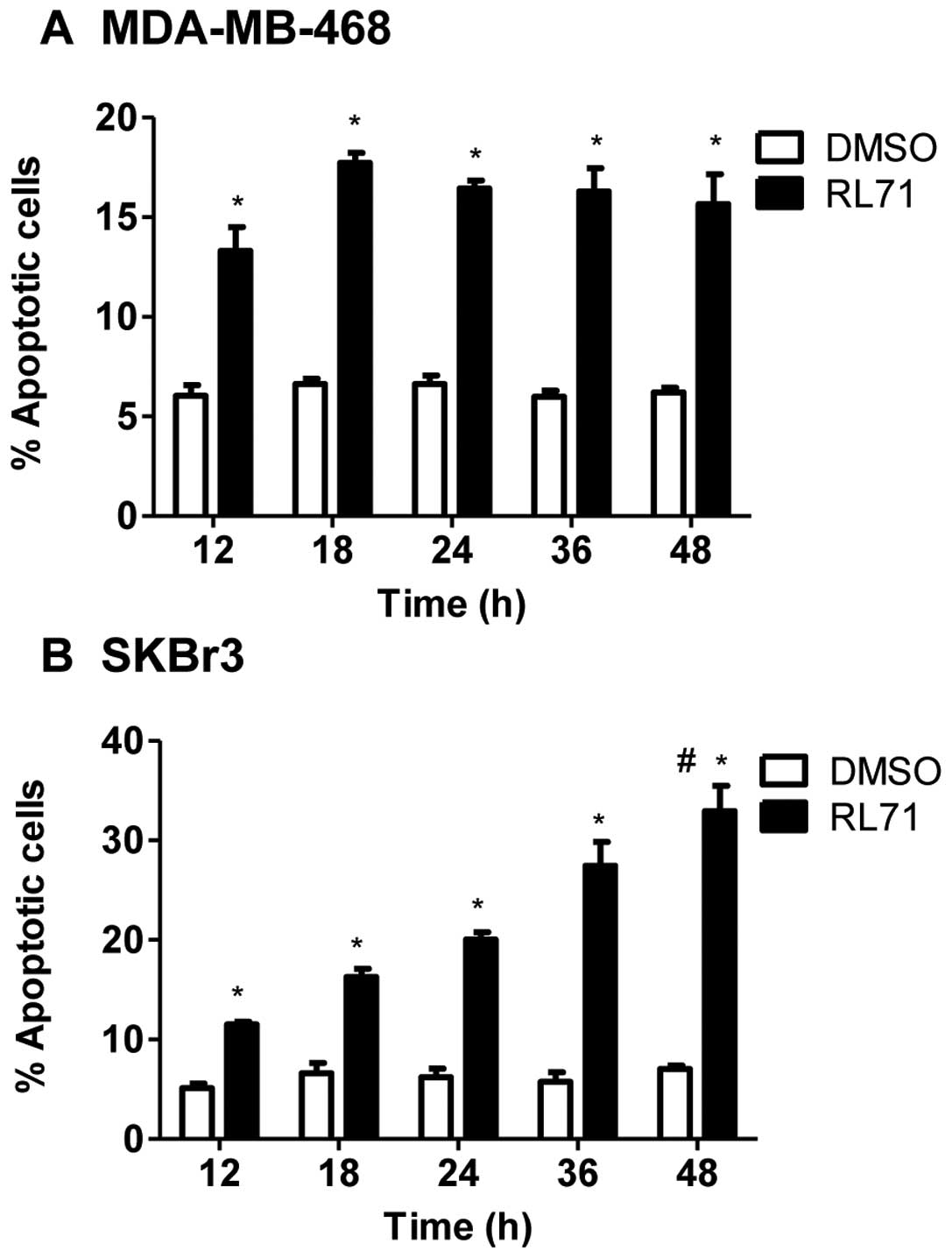
To determine if cell cycle arrest drives apoptosis,
time-dependent changes in apoptosis were examined. RL71 (1
μM) induced apoptosis in MDA-MB-468 and SKBr3 cells
(Fig. 4). The effect was
time-dependent in SkBr3 cells, as 35% of cells were apoptotic after
48 h and this was significantly elevated compared to all other time
points (Fig. 4B). In contrast
14–18% of MDA-MB-468 cells underwent apoptosis and this effect was
maintained from 12–48 h (Fig. 4A)
indicating the lack of a time-dependent effect. G2/M arrest did not
drive apoptosis in MDA-MB-468 cells, as apoptosis was increased at
12 h, which was prior to the increase in G2/M phase arrest.
However, the early appearance of G2/M phase arrest at 12 h in SKBr3
cells is a likely reason why these cells show a strong apoptotic
response over time. Additionally, our previous work with RL71 in
MDA-MB-231 cells indicated that the induction of apoptosis was
strongest in this cell line, as 43% of cells underwent apoptosis
and this effect was maintained from 18–36 h (23). Thus, the time-dependent increase in
G2/M phase arrest leads to the sustained apoptotic effect. It is
clear that overall RL71 displayed a more potent cytotoxic effect in
SKBr3 cells. To determine if this was due to the inhibition of
HER2/neu expression, changes in cell signaling proteins were
determined via western blotting.
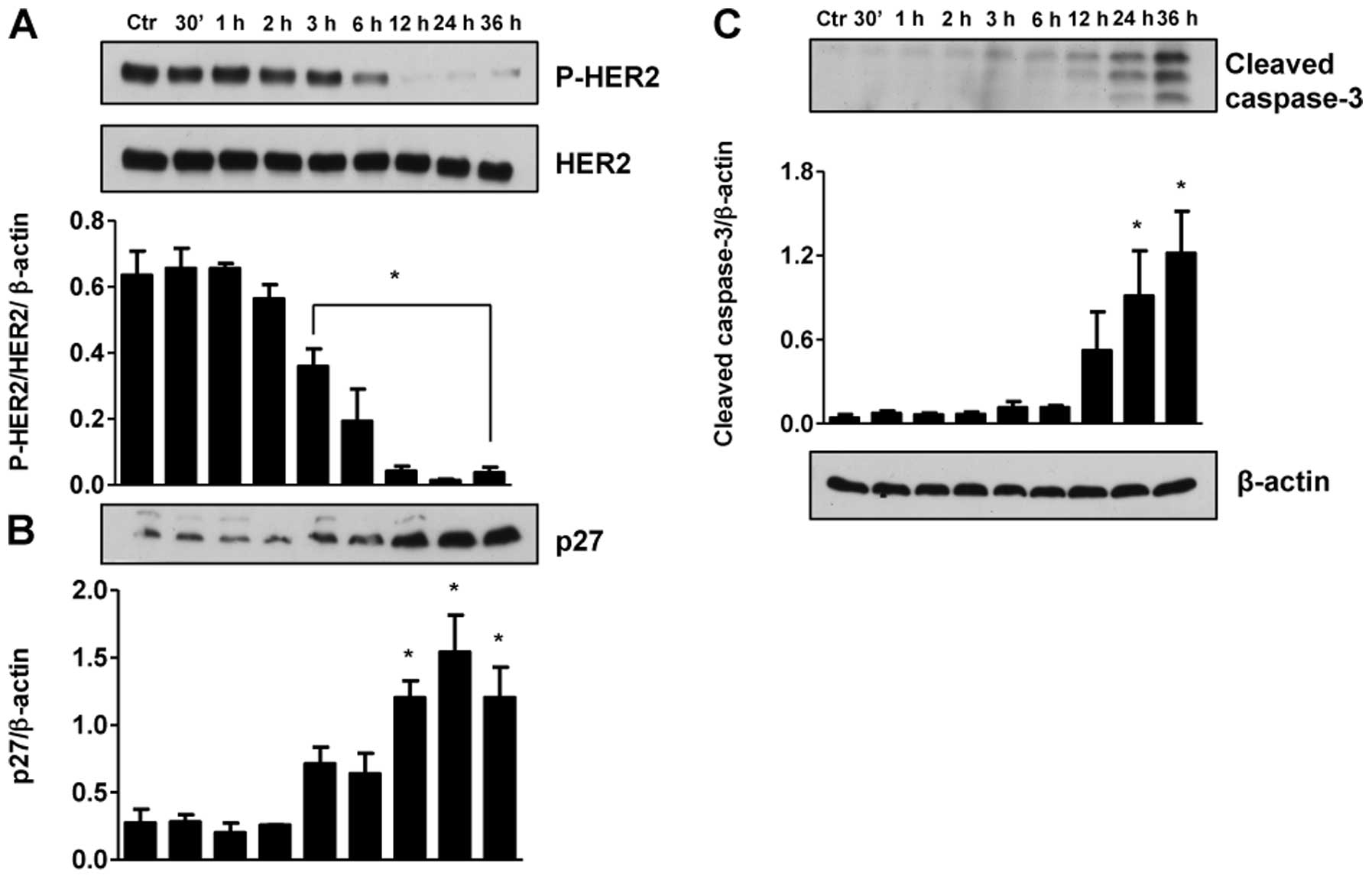
HER2/neu expression was assessed in SKBr3 cells to
determine the role of this protein in the cytotoxic actions of
RL71. Treatment of SKBr3 cells with RL71 (1 μM) decreased
the ratio of pHER2/HER2 in a time-dependent manner, with an almost
complete inhibition following 12 h (Fig. 5A). To link the changes in HER2/neu
with cell cycle progression protein changes in the cyclin dependent
kinase inhibitor, p27 were determined. The results showed that that
decrease in HER2/neu correlated with a significant increase in the
expression of p27 (Fig. 5B). Thus
the decrease in HER2/neu leads to an increase in p27 leading to the
observed G2/M arrest and apoptosis. The presence of apoptosis was
also confirmed in SKBr3 cells by the signficant increase in cleaved
caspase-3 (Fig. 5C).
To determine the molecular mechanisms responsible
for apoptosis and cell cycle arrest in TNBC cells we first examined
other isoforms of the EGFR in these cells. RL71 (1 μM)
failed to alter the ratio of pEGFR/EGFR protein levels (data not
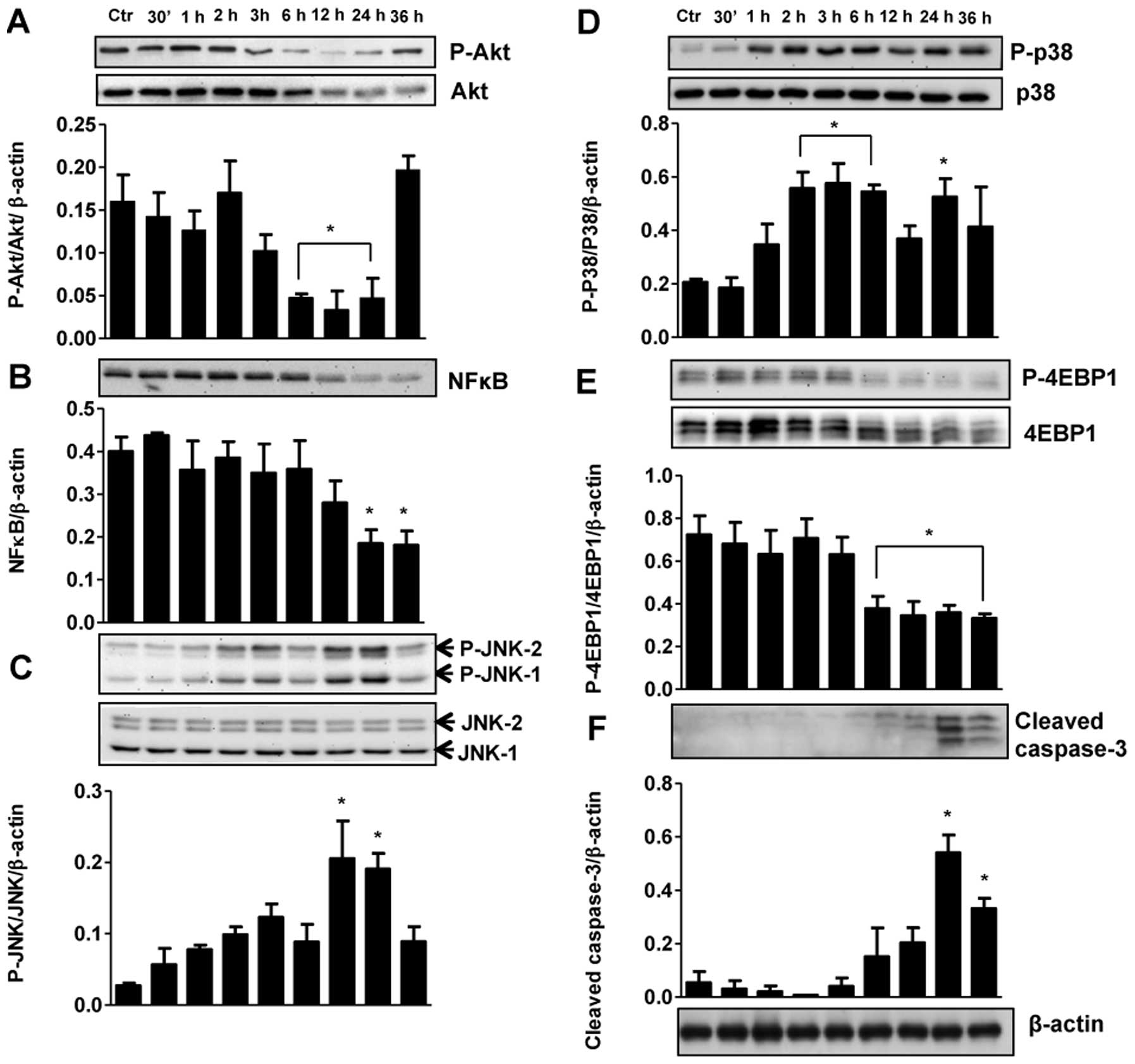
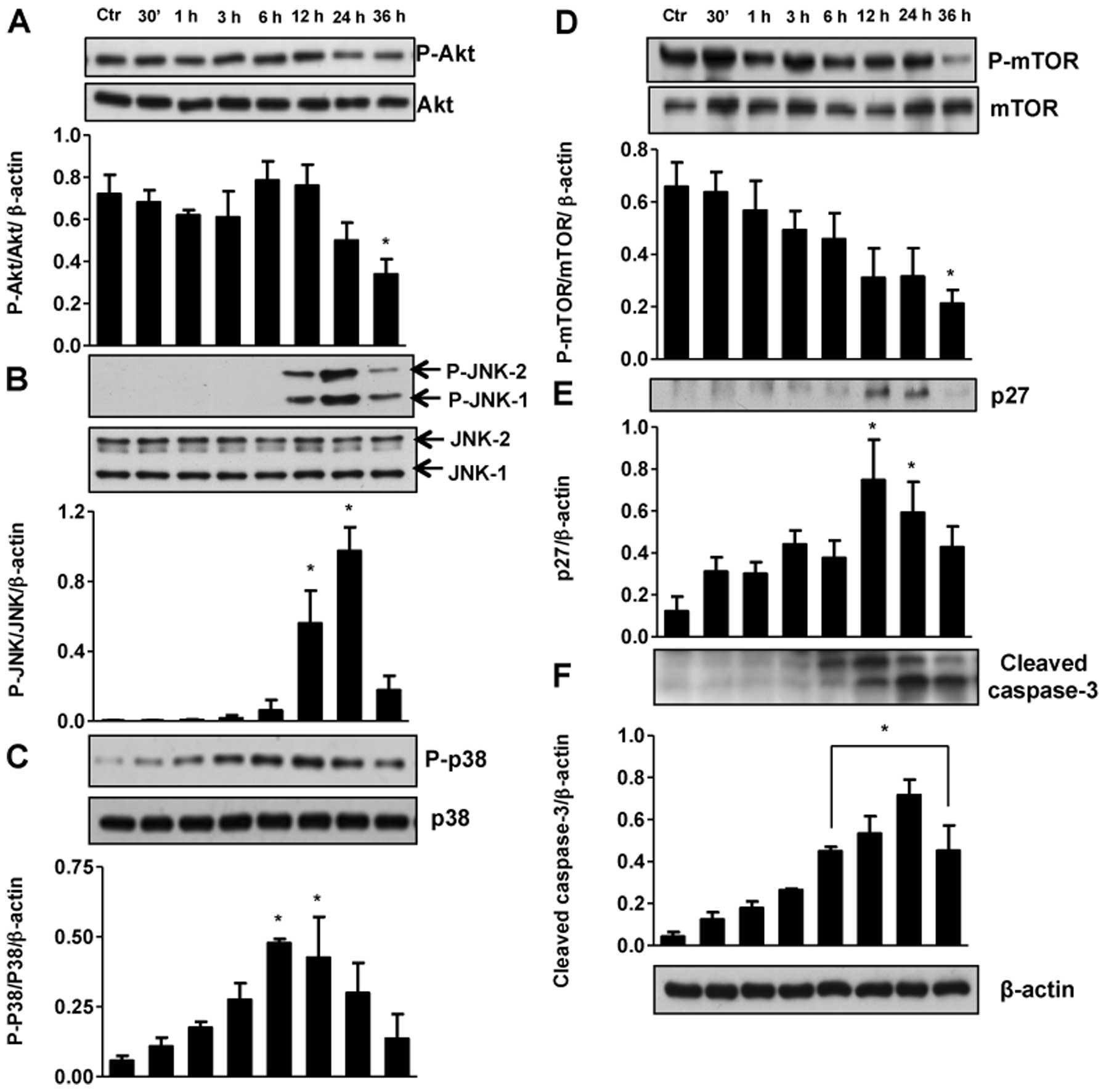
shown). However, RL71 did modulate the expression of Akt, JNK1/2,
p38 and caspase-3 in MDA-MB-231 and MDA-MB-468 cells and
differentially expressed NF-κB, mTOR, 4EBP1 and p27. Specifically,
RL71 significantly decreased the ratio of pAkt/Akt from 6–24 h in
MDA-MB-231 cells and at 36 h in MDA-MB-468 cells (Fig. 6A and 7A). The stress initiated by the treatment
of MDA-MB-231 and MDA-MB-468 cells resulted in a transient increase
in both JNK1/2 and p38 MAPK phosphorylation (Fig. 6C and D and Fig. 7B and C). Furthermore, RL71
increased levels of cleaved caspase-3 in a time-dependent manner
and this effect was strongest in MDA-MD-468 cells with a
significant increase occuring as early as 6 h (Fig. 6F and 7F). MDA-MB-231 cells were the only cell
line to show a signficant decrease in NF-κB and the ratio of
p4EBP1/4EBP1 following RL71 (Fig. 6B
and E), while the ratio of pmTOR/mTOR was signficantly
decreased and p27 was signficantly increased by RL71 in MDA-MB-468
cells (Fig. 7D and E).
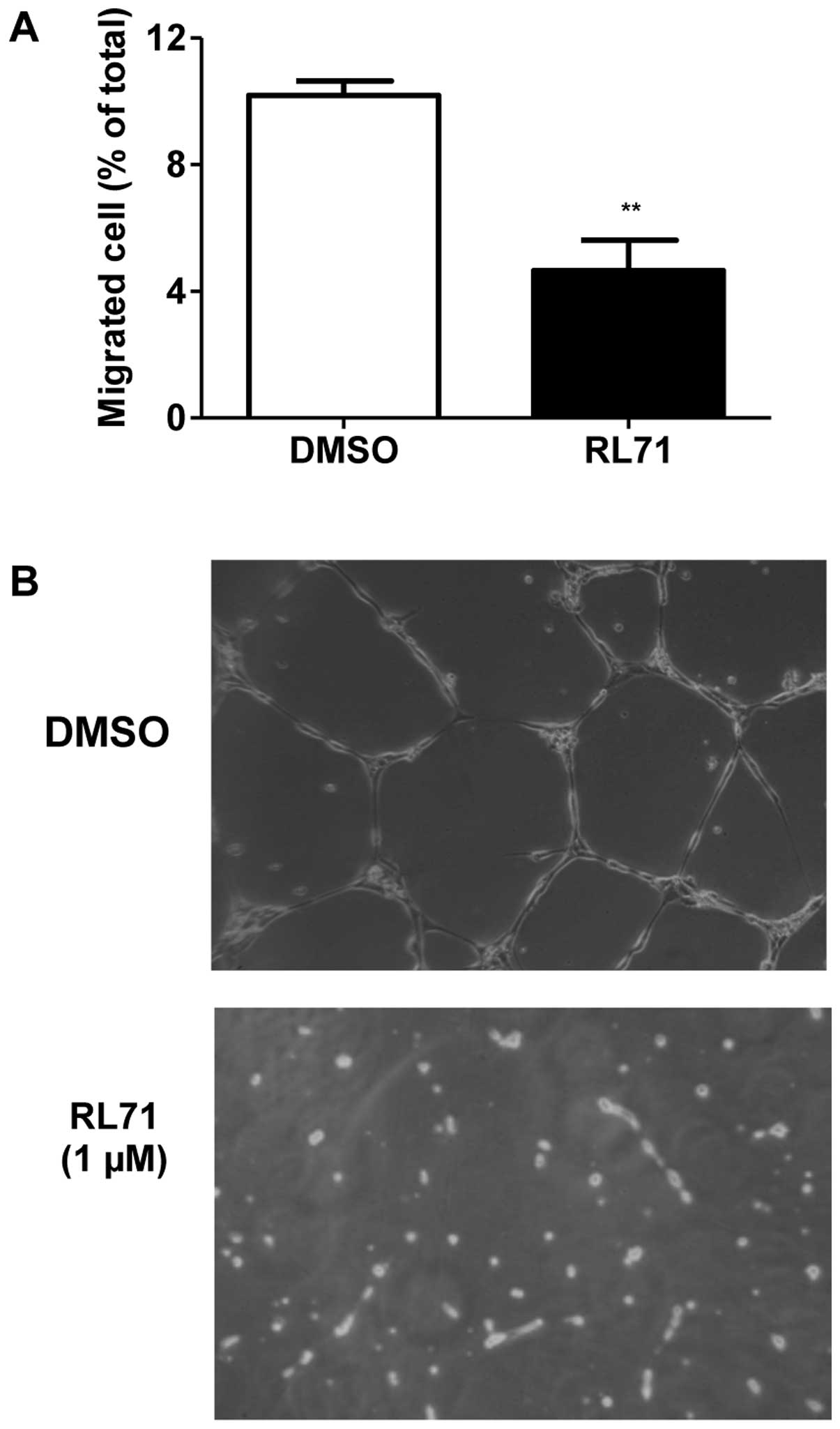
To determine if RL71 could modulate angiogenesis,
in vitro assays using HUVEC cells were performed, as the
ability of these cells to migrate through Matrigel and form
tube-like networks are hallmarks of angiogenesis. We used both
quantifiable and visual assys to form a more complete in
vitro picture. The results showed that RL71 (1 μM)
significantly reduced HUVEC cell migration by 46% compared to
vehicle control (Fig. 8A) and
completely inhibited endothelial tube formation after 18 h
(Fig. 8B).
To confirm that RL71 was a new analog that showed
improved oral bioavailability, female CD-1 mice were orally gavaged
with a single dose of RL71 (8.5 mg/kg) and the resulting plasma
concentration was determine by HPLC analysis. The results showed a
peak plasma concentration of 0.405 μg/ml, 5 min after oral
drug administration. The plasma concentration of RL71 decreased in
a time-dependent manner, as 0.15 μg/ml was present after 1 h
and by 2 h the plasma concentration was at the limit of
detection.
Discussion
We have previously shown that RL71 elicited the most
potent cytotoxicity towards ER-negative breast cancer cells
compared to all of our previously examined second generation
curcumin derivatives (23).
Therefore, this study was designed to further characterize this
cytotoxic effect in vitro in a range of ER-negative breast
cancer cell lines. The data presented show that RL71 promoted G2/M
cell cycle arrest, induced apoptosis and modulated the
Akt-dependent signaling pathway and stress response MAPK pathway.
RL71 also downregulated the expression of HER2/neu in SKBr3 cells.
In addition, RL71 exhibited anti-angiogenic effects in vitro
by inhibiting the invasion of HUVEC cells and their ability to form
endothelial tube like network. Importantly, RL71 is also orally
bioavailable.
Breast cancer patients whose tumors overexpress
HER2/neu have a poor prognosis, shorter relapse time and shorter
survival time (27). In this study
we showed that RL71 inhibited HER2/neu expression in SKBr3 cells
and this correlated with a concominant increase in p27 and cleaved
caspase-3. Since p27 is a key regulator of G2/M phase arrest and
apoptosis (28,29), inhibition of HER2/neu is a key
initial mechanism for the apoptotic effect elicited by RL71 in
SKBr3 cells. RL71 was more potent than other curcumin analogs at
downregulating the expression of HER2/neu, as 4 μM
concentrations of RL90 and RL91 and 10 μM concentrations of
and FLLL11 and FLLL12, were required to elicit a similar effect
(25). This confirms that RL71 is
a lead drug candidate for ER-negative/HER2-positive breast
cancer.
MAPK signaling which includes activation of JNK and
p38 has been shown to be involved in the regulation of the cell
cycle and induction of apoptosis in breast cancer cells (30). Various cytotoxic agents induce
apoptotic cell death via activation of MAPK signaling and induction
of caspase-3 (31–33). Our studies showed that RL71
treatment induced JNK1/2 and p38 MAPK in MDA-MB-231 and MDA-MB-468
cells. Anticancer agents such as curcumin, which causes activation
of p38, JNK1/2 and caspase-3, also induce similar apoptotic events
(34,35). The MAPK pathway may also upregulate
cell cycle regulatory protein, p27 in breast cancer cells (36). Our results demonstrated that in
MDA-MB-468 RL71 enhanced the expression of p27 which would
contribute to the observed G2/M cell cycle arrest.
We further studied the effect of RL71 on the
PI3K/Akt/mTOR pathway. Akt is an important oncoprotein which is
constitutively active in breast cancer cells and has been
implicated in a myriad of regulatory mechanisms involving protein
synthesis, cell cycle progression and inhibition of apoptosis
(37,38). Our results showed that RL71
decreased the phosphorylation of Akt on Ser-473 in a cell line and
time-dependent manner. Specifically, in MDA-MB-468 cells, RL71
decreased Akt phosphorylation fully whereas in MDA-MB-231 cells the
phosphorylation of Akt was partially decreased. However, RL71 was
more potent than the analogs RL90 and RL91, which did not decrease
the ratio of pAkt/Akt at concentrations of 4 μM (25). The decreased activity of Akt led to
decreased activation of its substrate mTOR in MDA-MB-468 cells. In
contrast, in MDA-MB-231 cells there was no change in the expression
of mTOR but the expression of 4-EBP1 which is downstream of mTOR
was fully downregulated. RL71 failed to alter the expression of
PI3K (data not shown) and this suggests that RL71 directly targets
downstream events of PI3K/Akt signaling in MDA-MB-231 cells and
increased phosphorylation of Akt could be due to a feedback loop
mechanism.
Akt contributes to the activity of NF-κB by
controling its translocation to the nucleus (39) and a decrease in Akt activity may
affect the stability and level of NF-κB (40). NF-κB belongs to a family of
transcription factors which has been associated with inhibition of
apoptosis by promoting the expression of anti-apoptotic proteins
such as Bcl-xL, c-Myb and caspase inhibitors (41,42).
RL71 downregulated the expression of NF-κB in MDA-MB-231 cells.
However, higher concentrations were required to downregulate NF-κB
in MDA-MB-468 cells (data not shown). Curcumin has also been shown
to interfere with the functions of Akt and MAPKs and further
inhibit its downstream target NF-κB (43,44)
and thus RL71 retains many of the same actions as curcumin.
To determine if RL71 exhibited anti-angiogenic
properties, we examined the effect of RL71 in multiple in
vitro angiogenesis assays. Angiogenesis is essential for tumor
growth and metastasis and inhibition of metastasis is critical as
it is cancer spread that leads to mortality. Importantly, RL71 had
an inhibitory effect on endothelial cell migration and tube
formation demonstrating an anti-angiogenic effect from this
compound. The mechanism of this effect will be studied further
using in vivo live imaging.
RL71 showed potent cytotoxicity in ER-negative
breast cancer cells compared to other cyclohexanone curcumin
analogs (21,23). Moreover, it had superior
cytotoxicity compared with other curcumin analogs such as
3,5-bis(flurobenzylidene) piperidin-4-one (EF24) (45) 5-bis
(4-hydroxy-3-methoxybenzylidnen)-N-methyl-4-piperidone (PAC)
(46) and GO-Y030 (47) in MDA-MB-231 cells. Specifically
IC50 values of 1.2, 1 and 0.3 μM were reported
for EF24, GO-Y030 and RL71, respectively (23,45,47).
While EF24 induced G2/M phase arrest and apoptosis in MDA-MB-231
cells (22) and inhibited the
NF-κB pathway in a TNFα-dependent manner (45), it has not been examined in other
breast cancer cells. Additionally, RL71 has a stronger ability to
induce apoptosis compared to the analog 4-hydroxy-3-methoxybenzoic
acid methyl ester (HM-BME), where 25 μM was required to
cause 37% of LNCaP prostate cancer cells to undergo apoptosis after
24 h (48). The curcumin analogs
FLLL11 and FLLL12 were equally potent as RL71 in MDA-MB-468 cells
with similar IC50 values (0.3 μM). However, this
did not translate to other breast cancer cell types as these
analogs had IC50s of 2–5 μM in MDA-MB-231 and
SkBr3 cells (42). These analogs
also down-regulated Akt phosphorylation and HER2/neu expression in
SKBr3 breast cancer cells but at concentrations of 10 μM,
10-fold greater than RL71 (49).
While other curcumin analogs appear to have a similar mechanism of
action as RL71, RL71 is more potent as all of its anti-cancer
actions were elicited at concentrations of 1 μM or less.
Importantly RL71 is orally available following a single oral dose
of 8.5 mg/kg and this is a significant improvement on both curcumin
(50) and other first generation
analogs (19).
In summary, we showed that RL71 causes cell cycle
arrest and induces apoptosis in ER-negative breast cancer cells and
also modulates a variety of signaling pathways that culminate in
potent cytotoxicity. Specifically, inhibition of Akt pathway and
the activation of p38/JNK pathway may contribute to the anti-cancer
activity of RL71 in TNBC cells, while inhibition of HER2/neu and
induction of p27 are key mechanisms in SKBr3 cells. Thus, RL71
shows potential as a new drug therapy for
ER-negative/Her-2-positive breast cancer. Therefore, RL71 is a
novel 2nd generation curcumin derivative that warrants further
investigation as a potential therapy for ER-negative breast
cancer.
Acknowledgements
This study was supported by a grant
from the Breast Cancer Research Trust (RJR) and a University of
Otago postgraduate scholarship (B.Y.).
References
|
1.
|
Parl FF, Schmidt BP, Dupont WD and Wagner
RK: Prognostic significance of estrogen receptor status in breast
cancer in relation to tumor stage, axillary node metastasis, and
histopathologic grading. Cancer. 54:2237–2242. 1984. View Article : Google Scholar : PubMed/NCBI
|
|
2.
|
Doane AS, Danso M, Lal P, Donaton M, Zhang
L, Hudis C and Gerald WL: An estrogen receptor-negative breast
cancer subset characterized by a hormonally regulated
transcriptional program and response to androgen. Oncogene.
25:3994–4008. 2006. View Article : Google Scholar
|
|
3.
|
Carey LA, Dees EC, Sawyer L, et al: The
triple negative paradox: primary tumor chemosensitivity of breast
cancer subtypes. Clin Cancer Res. 13:2329–2334. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
4.
|
Sorlie T, Perou CM, Tibshirani R, et al:
Gene expression patterns of breast carcinomas distinguish tumor
subclasses with clinical implications. Proc Natl Acad Sci USA.
98:10869–10874. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
5.
|
Kaplan HG and Malmgren JA: Impact of
triple negative phenotype on breast cancer prognosis. Breast J.
14:456–463. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
6.
|
Anand P, Thomas Sherin G, Kunnumakkara
Ajaikumar B, et al: Biological activities of curcumin and its
analogues (Congeners) made by man and Mother Nature. Biochem
Pharmacol. 76:1590–1611. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
7.
|
Cheng A-L, Hsu C-H, Lin J-K, et al: Phase
I clinical trial of curcumin, a chemopreventive agent, in patients
with high-risk or pre-malignant lesions. Anticancer Res.
21:2895–2900. 2001.PubMed/NCBI
|
|
8.
|
Inano H, Onoda M, Inafuku N, et al:
Chemoprevention by curcumin during the promotion stage of
tumorigenesis of mammary gland in rats irradiated with gamma-rays.
Carcinogenesis. 20:1011–1018. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
9.
|
Pereira MA, Grubbs CJ, Barnes LH, et al:
Effects of the phytochemicals, curcumin and quercetin, upon
azoxymethane-induced colon cancer and
7,12-dimethylbenz[a]anthracene-induced mammary cancer in rats.
Carcinogenesis. 17:1305–1311. 1996.PubMed/NCBI
|
|
10.
|
Schaaf C, Shan B, Buchfelder M, et al:
Curcumin acts as anti-tumorigenic and hormone-suppressive agent in
murine and human pituitary tumour cells in vitro and in vivo.
Endocr Relat Cancer. 16:1339–1350. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
11.
|
Singletary K, MacDonald C, Wallig M and
Fisher C: Inhibition of 7,12-dimethylbenz[a]anthracene
(DMBA)-induced mammary tumorigenesis and DMBA-DNA adduct formation
by curcumin. Cancer Lett. 103:137–141. 1996.
|
|
12.
|
Chiu T-L and Su C-C: Curcumin inhibits
proliferation and migration by increasing the Bax to Bcl-2 ratio
and decreasing NF-κBp65 expression in breast cancer MDA-MB-231
cells. Int J Mol Med. 23:469–475. 2009.PubMed/NCBI
|
|
13.
|
Kang HJ, Lee SH, Price JE and Kim LS:
Curcumin suppresses the paclitaxel-induced nuclear factor-κB in
breast cancer cells and potentiates the growth inhibitory effect of
paclitaxel in a breast cancer nude mice model. Breast J.
15:223–229. 2009.
|
|
14.
|
Liu Q, Loo WTY, Sze SCW and Tong Y:
Curcumin inhibits cell proliferation of MDA-MB-231 and BT-483
breast cancer cells mediated by down-regulation of NFκB, cyclinD
and MMP-1 transcription. Phytomed. 16:916–922. 2009.PubMed/NCBI
|
|
15.
|
Prasad CP, Rath G, Mathur S, Bhatnagar D
and Ralhan R: Potent growth suppressive activity of curcumin in
human breast cancer cells: Modulation of Wnt/beta-catenin
signaling. Chem Biol Interact. 181:263–271. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
16.
|
Rowe DL, Ozbay T, O’Regan RM and Nahta R:
Modulation of the BRCA1 protein and induction of apoptosis in
triple negative breast cancer cell lines by the polyphenolic
compound curcumin. Breast Cancer Basic Clin Res. 3:61–75.
2009.PubMed/NCBI
|
|
17.
|
Somers-Edgar TJ, Scandlyn MJ, Stuart EC,
Le Nedelec MJ, Valentine SP and Rosengren RJ: The combination of
epigallocatechin gallate and curcumin suppresses ERalpha-breast
cancer cell growth in vitro and in vivo. Int J Cancer.
122:1966–1971. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
18.
|
Wu X and Wu K: Antiproliferative effect of
curcumin on human breast cancer of MCF-7 cells. Di-San Junyi Daxue
Xuebao. 28:1870–1872. 2006.
|
|
19.
|
Liang G, Shao L, Wang Y, et al:
Exploration and synthesis of curcumin analogues with improved
structural stability both in vitro and in vivo as cytotoxic agents.
Bioorg Med Chem. 17:2623–2631. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
20.
|
Markaverich BM, Schauweker TH, Gregory RR,
Varma M, Kittrell FS, Medina D and Varma RS: Nuclear type II sites
and malignant cell proliferation: inhibition by
2,6-bis-benzylidenecyclohexanones. Cancer Res. 52:2482–2488.
1992.PubMed/NCBI
|
|
21.
|
Adams BK, Ferstl EM, Davis MC, et al:
Synthesis and biological evaluation of novel curcumin analogs as
anti-cancer and anti-angiogenesis agents. Bioorg Med Chem.
12:3871–3883. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
22.
|
Adams Brian K, Cai J, Armstrong J, et al:
EF24, a novel synthetic curcumin analog, induces apoptosis in
cancer cells via a redox-dependent mechanism. Anticancer Drugs.
16:263–275. 2005.PubMed/NCBI
|
|
23.
|
Yadav B, Taurin S, Rosengren RJ,
Schumacher M, Diederich M, Somers-Edgar TJ and Larsen L: Synthesis
and cytotoxic potential of heterocyclic cyclohexanone analogues of
curcumin. Bioorg Med Chem. 18:6701–6707. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
24.
|
Skehan P, Storeng R, Scudiero D, et al:
New colorimetric cytotoxicity assay for anti-cancer drug screening.
J Natl Cancer Inst. 82:1107–1112. 1990. View Article : Google Scholar : PubMed/NCBI
|
|
25.
|
Somers-Edgar TJ, Taurin S, Larsen L,
Chandramouli A, Nelson MA and Rosengren RJ: Mechanisms for the
activity of heterocyclic cyclohexanone curcumin derivatives in
estrogen receptor negative human breast cancer cell lines. Invest
New Drugs. 29:87–97. 2011. View Article : Google Scholar
|
|
26.
|
Stuart EC and Rosengren RJ: The
combination of raloxifene and epigallocatechin gallate suppresses
growth and induces apoptosis in MDA-MB-231 cells. Life Sci.
82:943–948. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
27.
|
Wang SC and Hung MC: HER2 overexpression
and cancer targeting. Semin Oncol. 28:115–124. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
28.
|
Hsieh WT, Huang KY, Lin HY and Chung JG:
Physalis angulata induced G2/M phase arrest in human breast cancer
cells. Food Chem Toxicol. 44:974–983. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
29.
|
Hsu JD, Kao SH, Ou TT, Chen YJ, Li YJ and
Wang CJ: Gallic acid induces G2/M phase arrest of breast cancer
cell MCF-7 through stabilization of p27(Kip1) attributed to
disruption of p27(Kip1)/Skp2 complex. J Agric Food Chem.
59:1996–2003. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
30.
|
Santen RJ, Song RX, McPherson R, Kumar R,
Adam L, Jeng MH and Yue W: The role of mitogen-activated protein
(MAP) kinase in breast cancer. J Steroid Biochem Mol Biol.
80:239–256. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
31.
|
Wada T and Penninger JM: Mitogen-activated
protein kinases in apoptosis regulation. Oncogene. 23:2838–2849.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
32.
|
Liu B, Han M, Sun RH, Wang JJ, Zhang YP,
Zhang DQ and Wen JK: ABL-N-induced apoptosis in human breast cancer
cells is partially mediated by c-Jun NH2-terminal kinase
activation. Breast Cancer Res. 12:R92010. View Article : Google Scholar : PubMed/NCBI
|
|
33.
|
Kuo PL, Chen CY and Hsu YL:
Isoobtusilactone a induces cell cycle arrest and apoptosis through
reactive oxygen species/apoptosis signal-regulating kinase 1
signaling pathway in human breast cancer cells. Cancer Res.
67:7406–7420. 2007. View Article : Google Scholar
|
|
34.
|
Collett GP and Campbell FC: Curcumin
induces c-jun N-terminal kinase-dependent apoptosis in HCT116 human
colon cancer cells. Carcinogenesis. 25:2183–2189. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
35.
|
Weir NM, Selvendiran K, Kutala VK, et al:
Curcumin induces G2/M arrest and apoptosis in cisplatin-resistant
human ovarian cancer cells by modulating Akt and p38 MAPK. Cancer
Biol Ther. 6:178–184. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
36.
|
Eto I: Nutritional and chemopreventive
anti-cancer agents up-regulate expression of p27Kip1, a
cyclin-dependent kinase inhibitor, in mouse JB6 epidermal and human
MCF7, MDA-MB-321 and AU565 breast cancer cells. Cancer Cell Int.
6:202006. View Article : Google Scholar : PubMed/NCBI
|
|
37.
|
Dillon RL, White DE and Muller WJ: The
phosphatidyl inositol 3-kinase signaling network: implications for
human breast cancer. Oncogene. 26:1338–1345. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
38.
|
Vivanco I and Sawyers CL: The
phosphatidylinositol 3-kinase AKT pathway in human cancer. Nat Rev
Cancer. 2:489–501. 2002. View
Article : Google Scholar : PubMed/NCBI
|
|
39.
|
Burow ME, Weldon CB, Melnik LI, Duong BN,
Collins-Burow BM, Beckman BS and McLachlan JA: PI3-K/AKT regulation
of NF-kappaB signaling events in suppression of TNF-induced
apoptosis. Biochem Biophys Res Commun. 271:342–345. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
40.
|
Gong L, Li Y, Nedeljkovic-Kurepa A and
Sarkar FH: Inactivation of NF-kappaB by genistein is mediated via
Akt signaling pathway in breast cancer cells. Oncogene.
22:4702–4709. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
41.
|
Barkett M and Gilmore TD: Control of
apoptosis by Rel/NF-kappaB transcription factors. Oncogene.
18:6910–6924. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
42.
|
Lauder A, Castellanos A and Weston K:
c-Myb transcription is activated by protein kinase B (PKB)
following interleukin 2 stimulation of T cells and is required for
PKB-mediated protection from apoptosis. Mol Cell Biol.
21:5797–5805. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
43.
|
Shehzad A, Wahid F and Lee YS: Curcumin in
cancer chemoprevention: molecular targets, pharmacokinetics,
bioavailability, and clinical trials. Arch Pharm. 343:489–499.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
44.
|
Dhandapani KM, Mahesh VB and Brann DW:
Curcumin suppresses growth and chemoresistance of human
glioblastoma cells via AP-1 and NFkappaB transcription factors. J
Neurochem. 102:522–538. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
45.
|
Kasinski AL, Du Y, Thomas SL, et al:
Inhibition of IkappaB kinase-nuclear factor-kappaB signaling
pathway by 3,5-bis(2-flurobenzylidene)piperidin-4-one (EF24), a
novel monoketone analog of curcumin. Mol Pharmacol. 74:654–661.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
46.
|
Al-Hujaily EM, Mohamed AG, Al-Sharif I, et
al: PAC, a novel curcumin analogue, has anti-breast cancer
properties with higher efficiency on Er-negative cells. Breast
Cancer Res Treat. 128:97–107. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
47.
|
Hutzen B, Friedman L, Sobo M, et al:
Curcumin analogue GO-Y030 inhibits STAT3 activity and cell growth
in breast and pancreatic carcinomas. Int J Oncol. 35:867–872.
2009.PubMed/NCBI
|
|
48.
|
Kumar AP, Garcia GE, Ghosh R, Rajnarayanan
RV, Alworth WL and Slaga TJ: 4-Hydroxy-3-methoxybenzoic acid methyl
ester: a curcumin derivative targets Akt/NF kappa B cell survival
signaling pathway: potential for prostate cancer management.
Neoplasia. 5:255–266. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
49.
|
Lin L, Hutzen B, Ball S, et al: New
curcumin analogues exhibit enhanced growth-suppressive activity and
inhibit AKT and signal transducer and activator of transcription 3
phosphorylation in breast and prostate cancer cells. Cancer Sci.
100:1719–1727. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
50.
|
Anand P, Kunnumakkara AB, Newman RA and
Aggarwal BB: Bioavailability of curcumin: problems and promises.
Mol Pharm. 4:807–818. 2007. View Article : Google Scholar : PubMed/NCBI
|