Introduction
Angiogenesis is an essential step in new blood
vessel formation for tumor growth and metastasis (1,2), and
it is regulated by the balance between angiogenic activators and
inhibitors (3,4). In addition, complex and diverse
cellular actions are implicated in angiogenesis, such as
degradation of the extracellular matrix (ECM), proliferation and
migration of endothelial cells, and morphological differentiation
of endothelial cells into tubes (5,6).
A critical role for inflammation in tumorigenesis
was postulated by Rudolf Virchow in 1863; generally, chronic
inflammation is linked to various steps involved in tumorigenesis,
including cellular transformation, promotion, survival,
proliferation, invasion, angiogenesis and metastasis (7–10).
Various inflammatory mediators, chemokines and growth factors have
been shown to enhance the proliferation and spread of tumor cells
with necessary nutrients and oxygen (11–13).
Both angiogenesis and inflammation are important
processes in tumor growth and expansion (7,14),
and these processes share common signal pathways in NF-κB and Akt
(15,16). Thus, we speculate that some
compounds with anti-inflammatory properties could also have
anti-angiogenic activities.
Recently, scientists have become interested in the
potential antitumor effects of nutrients due to their safety and
general acceptance. Many publications have demonstrated that a
large class of natural compounds, including pharmaconutrients,
exhibits antitumoral activities against selected cancer types
(17,18). We focused on the natural compound
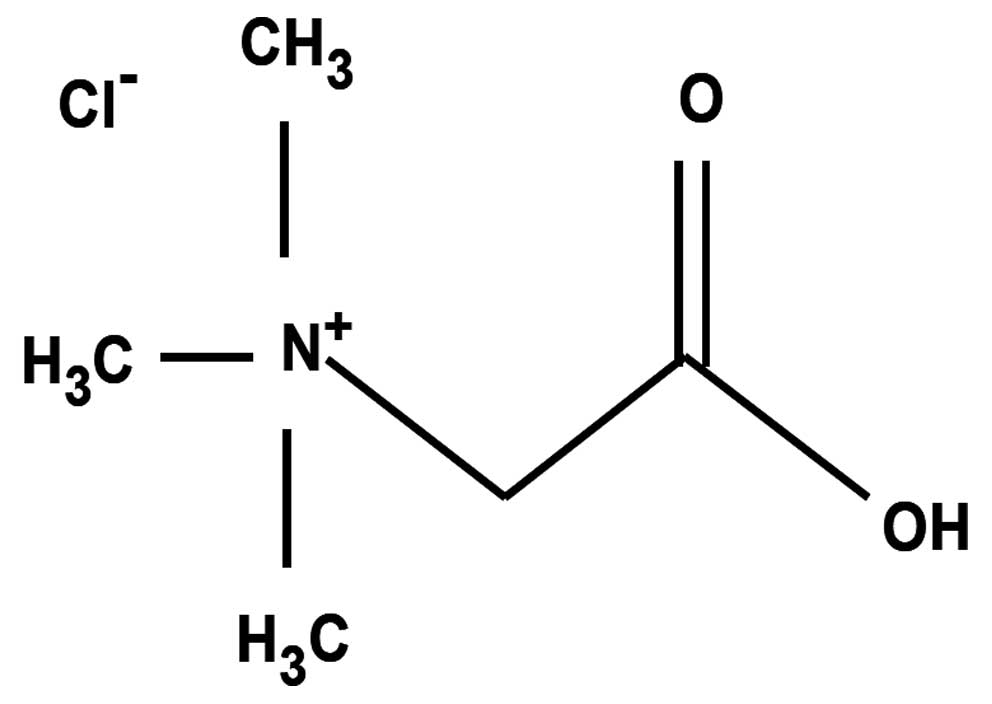
betaine (Fig. 1). Betaine is an
essential biochemical molecule of the methionine/homeocystein cycle
and is synthesized by conversion of choline. It was first
discovered in the juice of sugar beets (Beta vulgaris) in
the 19th century (19), and since
then has been found in various microorganisms, plants and animals
(20,21). In addition to being used as a food
source, water extracts of betaine from Lyciumchinensis have
been used in traditional oriental medicine for the treatment of
hepatic disorders in Southeast Asia (22). The biological function of betaine
was proposed to act as an osmolyte to protect cells under stress as
a catabolic source of methyl through trans-methylation in many
biochemical pathways (23–25).
Recent studies have shown that betaine suppresses
inflammation through NF-κB activation during aging (26). Since NF-κB signalling is common to
inflammation and angiogenesis, we investigated the anti-angiogenic
activities of betaine using in vitro and in vivo
angiogenesis assays. As a result, we found that betaine suppressed
tube formation, migration, and invasion of human umbilical vein
endothelial cells (HUVECs). Betaine also inhibited in vivo
angiogenesis in the mouse Matrigel plug assay. In addition, betaine
downregulated the mRNA expression levels of basic fibroblast growth
factor (bFGF), matrix metalloproteinase-2 (MMP-2) and matrix
metalloproteinase-9 (MMP-9) in HUVECs. We also found that betaine
inhibited NF-κB and Akt activation in HUVECs.
Materials and methods
Materials and reagents
HUVECs were purchased from Innopharma Screen
(Chungnam, Korea). Matrigel was from Collaborative Biomedical
Products (Bedford, MA, USA) for the mouse Matrigel plug assay.
Basic fibroblast growth factor (bFGF) and heparin were from
Peprotech (Gaithersburg, MD, USA). FBS, penicillin and streptomycin
were purchased from JBI (Daegu, Korea). Drabkin reagent kit 525 was
purchased from Sigma (St. Louis, MO, USA). Trans-well filter
chambers (8-μm pores) were purchased from Corning-Costar
(Cambridge, MA, USA). Antibodies for NF-κB, phospho-NF-κB, Akt and
phospho-Akt were purchased from Cell Signaling Technology Inc.
(Danvers, MA, USA). Betaine and gelatin were purchased from
Sigma.
Cell culture
HUVECs were grown in M199, supplemented with
heat-inactivated 20% fetal bovine serum (JBI), 20 ng/Ml of bFGF,
100 U/Ml of penicillin and 100 μg/Ml of streptomycin in a
37°C incubator with a humidified atmosphere containing 5%
CO2.
Animals
Seven-week-old, specific pathogen-free (SPF) male
C57BL/6 mice were supplied from Hyochang Science and Samtako
(Daegu, Korea). They were provided with autoclaved tap water and
lab chow ad libitum and were housed at 23±0.5°C, 10%
humidity under a 12 h light-dark cycle. The animal protocol used in
this study was reviewed on the ethical procedures and scientific
care, and approved by the Pusan National University-Institutional
Animal Care and Use Committee (PNU-IACUC).
In vitro tube formation assay
HUVECs (2x104 cells) were seeded on a
layer of previously polymerized matrigel and treated with or
without betaine. Matrigel culture was incubated at 37°C. After 18
h, changes in cell morphology were captured through a phase
contrast microscope and photographed at x40 magnification. Each
sample was assayed in duplicate, and independent experiments were
repeated three times.
In vitro wounding migration assay
HUVECs were seeded onto 24-well culture plates until
confluence and left overnight. Media were aspirated the next day,
and cells were scratched with a 200 μl pipette tip along the
diameter of the well. Cells were washed twice with PBS and
incubated at 37°C and 5% CO2. After wounding, the
cultures were washed with serum-free medium and further incubated
in M199 with 1% serum, 1 mM thymidine, and/or betiane. These
culture conditions minimized proliferation of HUVECs. Wound
diameters were photographed at 18 to 24 h. Wound closure was
determined by measurement with optical microscopy at x40
magnification. Migration was quantitated by counting the number of
cells that moved beyond the reference line. Each sample was assayed
in duplicate, and independent experiments were repeated three
times.
In vitro invasion assay
Invasiveness of HUVECs was performed in vitro
using a trans-well chambers system (Corning-Costar) with
8.0-μm pore polycarbonate filter inserts. The upper side was
coated with 10 μl of matrigel (0.5 mg/Ml) at room
temperature for 1 h. Cells (2x104 cells) and betaine in
serum-free media were placed in the upper part of the filters, and
full media was treated in the lower parts. Cells were incubated at
37°C for 24 h, fixed with methanol, and then stained with H&E.
Cells on the upper surface of the membrane were removed by wiping
with a cotton swab. Cell invasion was determined by counting whole
cell numbers in a single filter by optical microscopy at x40
magnification. Each sample was assayed in duplicate, and
independent experiments were repeated three times.
In vivo mouse Matrigel plug assay
C57BL/6 mice (7 weeks of age) were injected
subcutaneously into 500 μl of matrigel (Collaborative
Biomedical Products) containing bFGF (100 ng/Ml) and heparin (50
U/Ml) without or with betaine. After injection, the matrigel
rapidly formed a plug. After 7 days, skin of the mouse was pulled
back to expose the matrigel plug, which remained intact. After
quantitative differences were noted and photographed, hemoglobin
content was measured using the Drabkin reagent kit 525 (Sigma) for
quantification of blood vessel formation. The amount of hemoglobin
was calculated from a known amount of hemoglobin assayed in
parallel. Independent experiments were repeated twice and at least
five mice in each experiment were used.
Reverse transcription-polymerase chain
reaction (RT-PCR)
Total-RNA from HUVECs was isolated using TRIzol
reagent (Invitrogen Corporation, Carlsbad, CA, USA) according to
the manufacturer’s instructions. First-stranded cDNA was
synthesized by M-MLV reverse transcriptase (Promega Corporation,
Madison, WI, USA) with 2 μg each of DNA-free total-RNA
sample and oligo (dT)15 (Life Technologies, Grand Island, NY, USA)
according to the manufacturer’s instructions. Equal amounts of cDNA
were subsequently amplified by PCR in a 20 μl reaction
volume containing 1X PCR buffer, dNTP mixture, 10 μM of each
specific primer and i-Taq™ DNA polymerase (iNtRON Biotechnology,
Korea). Amplification products were electrophoresed on 2% agarose
gels and visualized by GelRed (Biotium Inc, Hayward, CA, USA)
staining under ultraviolet trans-illumination.
Western blot analysis
HUVECs were treated with or without betaine for 24 h
in medium. Total cell lysates were prepared by addition of PRO-PREP
Protein Extraction Solution (iNtRON Biotechnology), including 1 mM
sodium orthovanadate. Equal amounts (30 μg) of samples were
resolved by electrophoresis on a 10% SDS-polyacrylamide gel,
transferred to a membrane, and sequentially probed with antibody.
The following primary antibodies were used at the indicated
dilutions: total NF-κB and anti-phospho-NF-κB, total Akt and
anti-phospho-Akt, 1:1,000 in 5% BSA in TBS-T.
Data analysis and statistics
Data are presented as means ± SD or as percentage of
control. Statistical comparisons between groups were performed
using Student’s t-test. *p<0.05 was considered
statistically significant.
Results
Betaine inhibits vascular network
formation of HUVECs
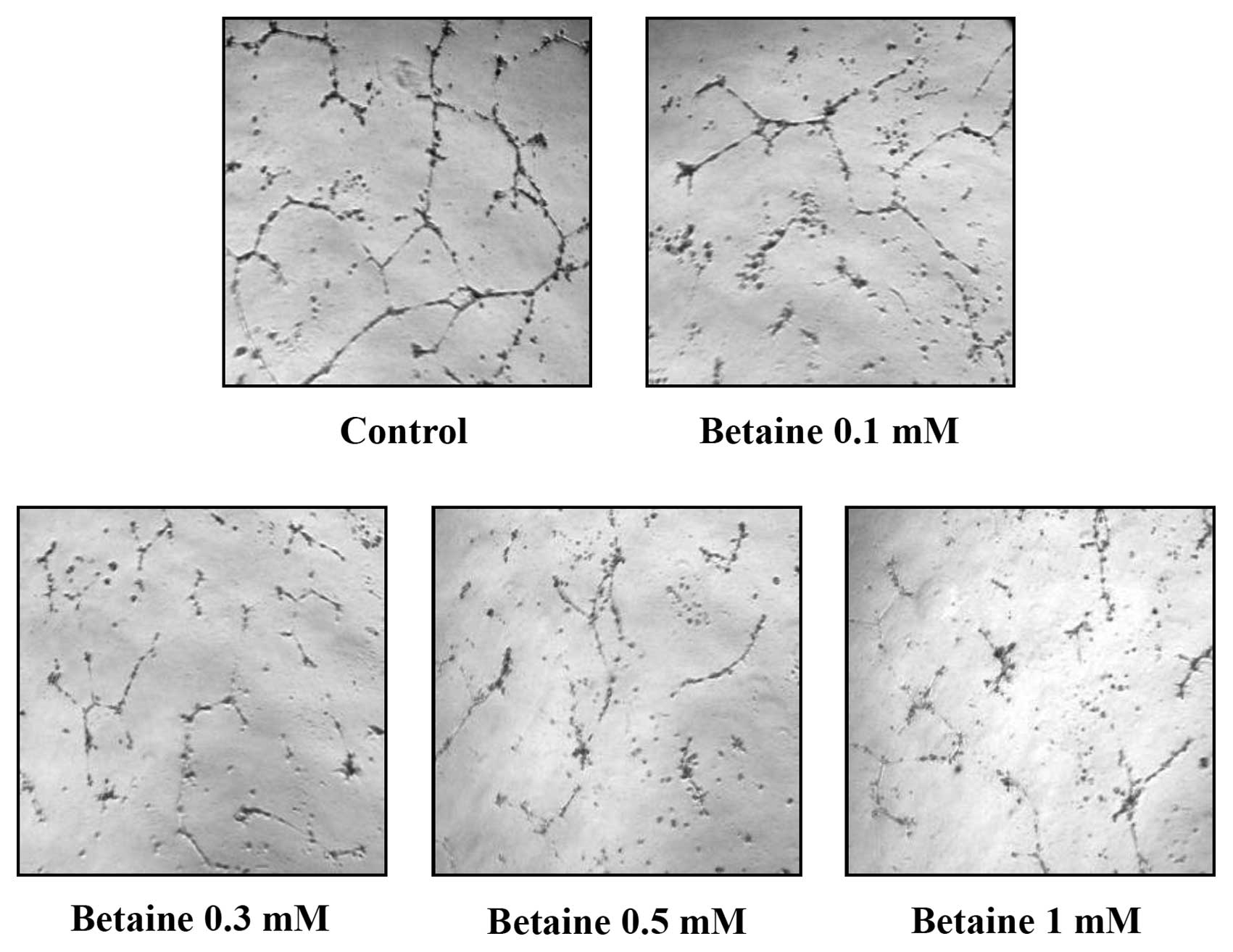
We carried out tube formation assay to examine the
effects of betaine on the morphological differentiation of
endothelial cells into a capillary-like structure, an important
step in the process of angiogenesis. HUVECs were placed on a
matrigel-coated plate and incubated. Endothelial cells formed
hollow capillaries on matrigel beds, and these tubes became
stronger and more robust with elongated networks between 6–24 h. As
shown in Fig. 2, HUVECs on
matrigel formed a typical blood vessel network in the absence of
betaine. However, treatment with betaine for 18 h resulted in
broken, shortened and much narrower tube networks. This finding
demonstrates that betaine inhibits tube formation of HUVECs.
Betaine inhibits migration and invasion
of HUVECs
The migration and invasion of endothelial cells are
critical features in the formation of new blood vessels and in the
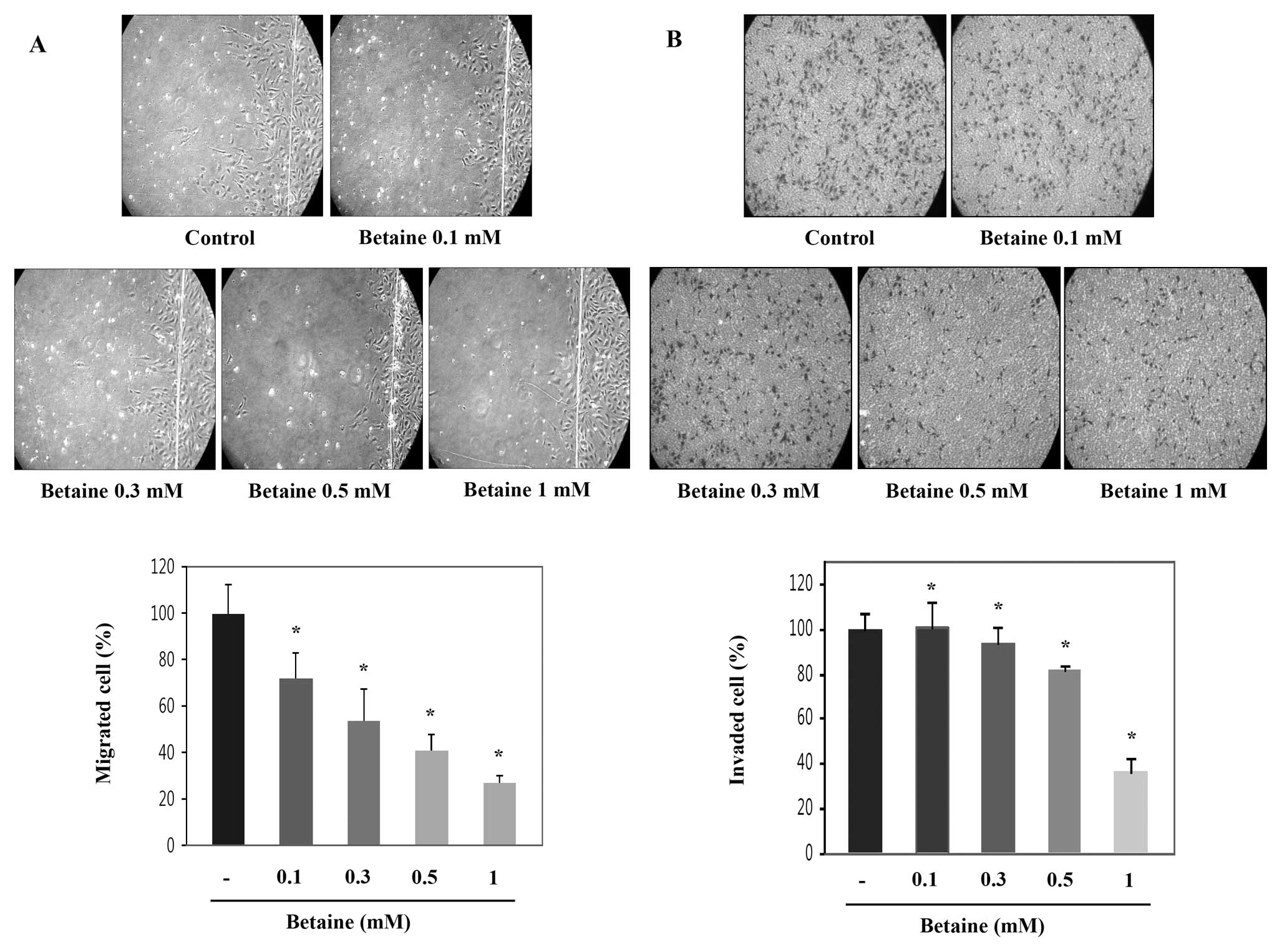
repair of injured vessels. Therefore, we investigated the effects
of betaine on the movement of HUVECs from a wounded edge to an open
area using wounding migration assay (Fig. 3). Treatment with betaine for 20 h
profoundly decreased the migration of HUVECs compared with that of
control in a dose-dependent manner (Fig. 3A). Migration of endothelial cells
was decreased by about 70% by betaine treatment compared with that
of control.
To examine the effects of betaine on the
invasiveness of HUVECs, we performed invasion assay with a
trans-well system. Trans-wells were prepared such that the upper
sides of the filter were coated with matrigel and used for invasion
assay. Betaine suppressed the invasiveness of HUVECs compared with
that of control in a dose-dependent manner after 24 h of incubation
(Fig. 3B). The invasiveness of
endothelial cells was inhibited by about 60% by betaine treatment
compared with that of control. Taken together, betaine strongly
suppressed migration and invasion of HUVECs, which are crucial
steps of angiogenesis.
Betaine inhibits angiogenesis in
vivo
We demonstrated the in vitro anti-angiogenic
activities of betaine as above. We then performed the mouse
Matrigel plug assay to examine the effects of betaine on
angiogenesis in vivo. As shown in Fig. 4A, matrigel plugs containing bFGF
were abundantly filled with intact red blood cells, which indicates
the formation of a functional vasculature inside the matrigel,
whereas vessels were not observed in matrigel alone (blank).
Matrigel plug containing betaine produced fewer vessels compared
with plug containing bFGF, indicating that betiane inhibited the
formation of bFGF-induced neomicrovessels. In addition, we measured
the hemoglobin contents inside the matrigel plugs in order to
quantify the anti-angiogenic effects of betaine. The hemoglobin
content of bFGF-treated plug was 32 g/dl, whereas that of 1 mM
betaine-treated plugs was profoundly diminished to about 2 g/dl
(Fig. 4B). These results indicate
that betaine has strong anti-angiogenic activity in
vivo.
Betaine downregulates mRNA expression
levels of bFGF, MMP-2 and MMP-9
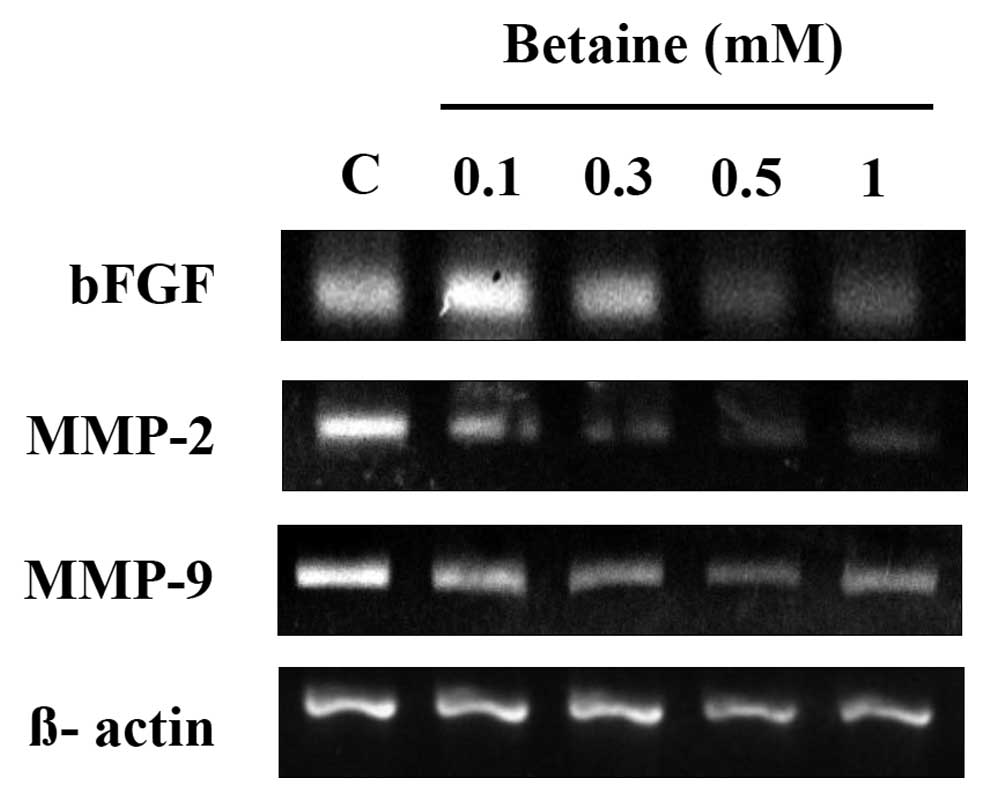
To determine which factors are involved in the
anti-angiogenic activity of betaine, we examined the mRNA
expression levels of angiogenic factors (VEGF and bFGF) in HUVECs
treated with betaine using RT-PCR. As shown in Fig. 5, treatment with betaine decreased
the mRNA expression of bFGF in a dose-dependent manner. However,
treatment with betaine did not produce any significant change in
mRNA levels of VEGF (date not shown).
Regulation of extracellular proteolytic activity is
important in the process of endothelial cell migration and invasion
through the basement membrane and capillary morphogenesis. Thus,
the mRNA expression levels of both MMP-2 and MMP-9, which are major
functional molecules in the degradation of the extracellular
membrane, were examined. The mRNA expression levels of both MMP-2
and MMP-9 were markedly downregulated by treatment with betaine in
a dose-dependent manner (Fig. 5).
These data suggest that the downregulated expression of bFGF, MMP-2
and MMP-9 mRNA may be responsible for the anti-angiogenic activity
observed in HUVECs treated with betaine.
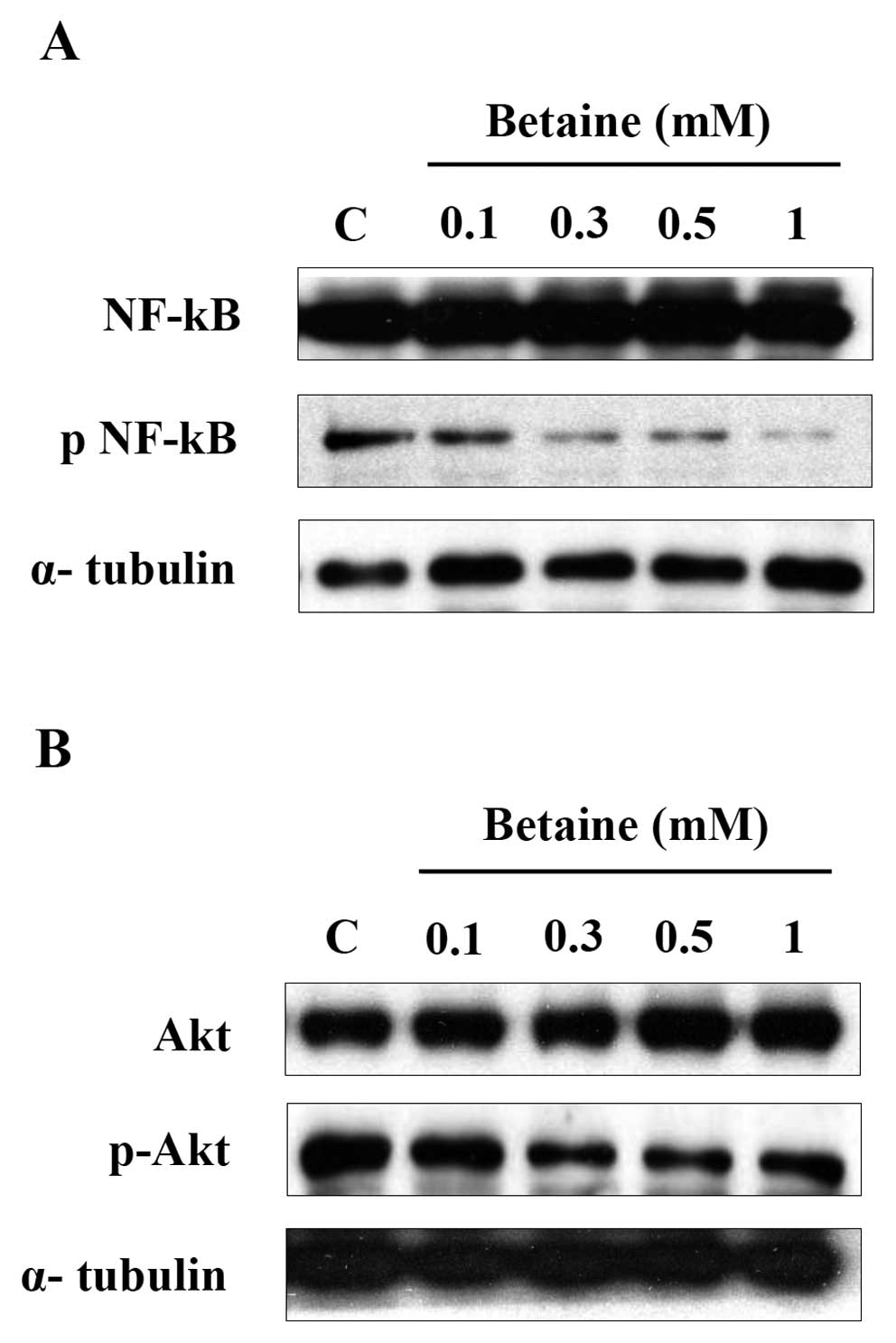
Betaine suppresses NF-κB and Akt
activation in HUVECs
Since NF-κB and Akt activation are two of the main
signaling pathways for tumor progression and angiogenesis, we
examined the effects of betaine on NF-κB and Akt activation in
HUVECs. In this experiment, total proteins were prepared and
analyzed using western blotting for detection of NF-κB and Akt
phosphorylation. As shown in Fig.
6A, NF-κB phosphorylation was suppressed by betaine in a
dose-dependent manner. As NF-κB activation is mediated by the Akt
signaling pathway, we next examined Akt activation. Phosphorylation
of Akt was inhibited by the presence of betaine in a dose-dependent
manner (Fig. 6B). These results
indicate that betaine interferes with NF-κB phosphorylation and Akt
phosphorylation in HUVECs.
Discussion
Angiogenesis, the formation of new blood vessels
from previously existing vessels, is required for tumor growth and
metastasis. Angiogenic processes involve several steps: i)
enzymatic degradation of vascular basement membranes by endothelial
cells, ii) directed migration and proliferation of endothelial
cells to provide cells for new vessels, iii) invasion of
endothelial cells and iv) formation of capillary-like networks by
endothelial cells (27).
Inflammation is also known to be a key process in
many cancers during tumorigenesis (28). Both angiogenesis and inflammation
are initiated by secreted chemokines/cytokines, growth factors and
other mediators (13). These
processes often share a common signal pathway. Thus,
anti-inflammation in the form of angiogenesis inhibitors has been
investigated as an anticancer therapy (29,30).
For example, anti-inflammatory drugs such as nonsteroidal
anti-inflammatory drugs (NAIDs) lead to the inhibition of
angiogenesis (31).
One of the approaches to anti-angiogenesis therapy
is using natural compounds from various organisms. Paclitaxel
(Taxol), doxorubicine and citarabine are examples of anticancer
agents originating from plants, microbes and a marine source,
respectively (17,32). Natural compounds from medicinal
plants display diverse pharmacological activities and have
advantages over synthetic drugs, such as smoother action and better
tolerance (33,34). Thus, the search for novel
anticancer drugs from natural sources has continued through the
collaboration of scientists looking for new bio-active compounds
(17,35).
Recently, betaine was demonstrated as having
anti-inflammatory effects by modulating NF-κB activation in the
kidney tissue of aged rats (26).
Since the NF-κB signal pathway participates both in inflammation
and angiogenesis, we examined the anti-angiogenic activity of
betaine by performing in vitro and in vivo
angiogenesis assays. Firstly, we observed the anti-angiogenic
effects of betaine in each step of angiogenesis in vitro.
Treatment with betaine clearly inhibited tube formation (Fig. 2), migration (Fig. 3A) and invasion (Fig. 3B) of HUVECs. To demonstrate the
in vivo anti-angiogenic effects of betaine, in vivo
mouse Matrigel plug assay was performed. Betaine excellently
inhibited the formation of neomicrovessels in matrigel (Fig. 4A), and we noted that these newly
synthesized vessels in matrigel plug containing betaine were not
abundantly filled with intact red blood cells (Fig. 4B). Taken together, these results
show that betaine suppressed angiogenesis in vitro and in
vivo.
In addition, tumors produce a wide array of
angiogenic molecules during angiogenesis (4). VEGF and bFGF are well known as key
stimuli for angiogenesis (36,37).
Therefore, we determined the involvement of betaine in the
expression of major angiogenic factors. As shown in Fig. 5, mRNA expression of bFGF was
remarkably reduced by treatment with betaine. However, we could not
observe any change in VEGF mRNA expression (date not shown).
Matrix metalloproteinases (MMPs) are secreted as
proenzymes and act as key regulators of extracellular matrix
turnover through degradation of a wide variety of extracellular
matrix proteins (38). The
regulation of extracellular proteolytic activity is important in
the process of endothelial cell migration and invasion, both of
which occur during angiogenesis, and is an essential step in tumor
invasion and metastasis. Based on our finding that motility of
HUVECs was profoundly suppressed by betaine, we examined the
effects of betaine on the expression of MMP-2 and MMP-9 using
RT-PCR. As shown in Fig. 5,
betaine downregulated the expression of both MMP-2 and MMP-9.
NF-κB and Akt are important mediators in tumor cell
invasion and metastasis (15,39).
Since betaine was recently reported to modulate the NF-κB signaling
pathway during aging (26), we
examined the activation of NF-κB and Akt by treatment with betaine
using western blot analysis. In the presence of betaine, activation
of NF-κB and Akt was suppressed in a dose-dependent manner
(Fig. 6).
In summary, betaine inhibited angiogenesis in
vitro and in vivo. These effects of betaine are linked
with decreased mRNA expression of bFGF, MMP-2 and MMP-9.
Furthermore, betaine suppressed angiogenesis through the NF-κB and
Akt signal pathways. Therefore, we conclude that betaine possesses
strong anti-angiogenic activity and could be developed as a useful
inhibitor of angiogenesis.
Acknowledgements
This study was supported in part by
National Research Foundation of Korea Grant funded by the Korean
Government (Ministry of Education, Science and Technology.
NRF-2010-355-C00044), and by a 2-Year Research Grant of Pusan
National University.
References
|
1.
|
Folkman J and Shing Y: Angiogenesis. J
Biol Chem. 267:10931–10934. 1992.
|
|
2.
|
Siddiqui FA, Desai H, Siddiqui TF and
Francis JL: Hemoglobin induces the expression and secretion of
vascular endothelial growth factor from human malignant cells.
Hematol J. 3:264–270. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
3.
|
Carmeliet P and Jain RK: Angiogenesis in
cancer and other diseases. Nature. 407:249–257. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
4.
|
Ferrara N and Kerbel RS: Angiogenesis as
therapeutic target. Nature. 438:967–974. 2005. View Article : Google Scholar
|
|
5.
|
Isner JM and Asahara T: Angiogenesis and
vasculogenesis as therapeutics strategies for postnatal
neovascularization. J Clin Invest. 103:1231–1236. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
6.
|
Folkman J: Tumor angiogenesis: therapeutic
implications. N Engl J Med. 285:1182–1186. 1971. View Article : Google Scholar : PubMed/NCBI
|
|
7.
|
Balkwill F and Mantovani A: Inflammation
and cancer: back to Virchow? Lancet. 357:539–545. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
8.
|
Albini A, Tosetti F, Benelli R and Noonan
DM: Tumor inflammatory angiogenesis and its chemoprevention. Cancer
Res. 65:10637–10641. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
9.
|
Aggarwal BB, Shishodia S, Sandur SK,
Pandey MK and Sethi G: Inflammation and cancer: how hot is the
link? Biochem Pharmacol. 72:1605–1621. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
10.
|
Sgambato A and Cittadini A: Inflammation
and cancer: a multifaceted link. Eur Rev Med Pharmacol Sci.
14:263–268. 2010.
|
|
11.
|
Raman D, Sobolik-Delmaire T and Richmond
A: Chemokines in health and disease. Exper Cell Res. 317:575–589.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
12.
|
Rock GA and Dalgleish A: Infection,
immunoregulation, and cancer. Immunol Rev. 240:141–159. 2011.
View Article : Google Scholar
|
|
13.
|
Grivennikov SI and Karin M: Inflammation
and oncogenesis: a vicious connection. Curr Opin Genet Dev.
20:65–71. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
14.
|
Yoshimura A: Signal transduction of
inflammatory cytokines and tumor development. Cancer Sci.
97:439–447. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
15.
|
Karin M and Greteo FR: NF-kappaB: linking
inflammation and immunity to cancer development and progression.
Nat Rev Immunol. 5:749–759. 2005. View
Article : Google Scholar : PubMed/NCBI
|
|
16.
|
Naldini A and Carraro F: Role of
inflammatory mediators in angiogenesis. Curr Drug Targets Inflam
Allergy. 4:3–8. 2005. View Article : Google Scholar
|
|
17.
|
da Rocha AB, Lopes RM and Schwartsmann G:
Natural products in anticancer therapy. Curr Opin Pharmacol.
1:364–369. 2001.PubMed/NCBI
|
|
18.
|
Granci V, Dupertuis YM and Pichard C:
Agiogenesis as a potential target of pharmaconutients in cancer
therapy. Curr Opin Clin Nutr Metab Care. 13:417–422. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
19.
|
Craig SA: Betaine in human nutrition. Am J
Clin Nutr. 80:539–549. 2004.PubMed/NCBI
|
|
20.
|
Schrama JW, Heetkamp MJ, Simmins PH and
Gerrits WJ: Dietary betaine supplementation affects energy
metabolism of pigs. J Anim Sci. 81:1202–1209. 2003.PubMed/NCBI
|
|
21.
|
Holm PI, Ueland PM, Vollset SE, et al:
Betaine and folate status as cooperative determinants of plasma
homocysteine in humans. Arterioscler Thromb Vasc Biol. 25:379–385.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
22.
|
Kim SK, Choi KH and Kim YC: Effect of
acute betaine administration on hepatic metabolism of S-amino acids
in rats and mice. Biochem Pharmacol. 65:1565–1574. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
23.
|
Garcia-Perez A and Burg MB: Renal
medullary organic osmolytes. Physiol Rev. 71:1081–1115. 1991.
|
|
24.
|
Sakamoto A and Murata N: Genetic
engineering of glycine betaine synthesis in plants: current status
and implications for enhancement of stress tolerance. J Exp Bot.
51:81–88. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
25.
|
Steenge GR, Verhoef P and Katan MB:
Betaine supplementation lowers plasma homocysteine in healthy men
and women. J Nutr. 133:1291–1295. 2003.PubMed/NCBI
|
|
26.
|
Go EK, Jung KJ, Kim JY, Yu BP and Chung
HY: Betaine suppresses proinflammatory signaling during aging: the
involvement of nuclear factor-kappaB via nuclear factor-inducing
kinase/IkappaB kinase and mitogen-activated protein kinases. J
Gerontol A Biol Sci Med Sci. 60:1252–1264. 2005. View Article : Google Scholar
|
|
27.
|
Dobrzanski P, Hunter K, Jones-Bolin S, et
al: Antiangiogenic and antitumor efficacy of EphA2 receptor
antagonist. Cancer Res. 64:910–919. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
28.
|
Lin YG, Kunnumarkkara AB, Nair A, et al:
Curcumin inhibits growth and angiogenesis in ovarian carcinoma by
targeting the nuclear factor-kappaB pathway. Clin Cancer Res.
13:3423–3430. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
29.
|
Szekanecz Z and Koch AE: Endothelial cells
in inflammation and angiogenesis. Curr Drug Targets Inflamm
Allergy. 4:319–323. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
30.
|
Ono M: Molecular links between tumor
angiogenesis and inflammation: inflammatory stimuli of macrophages
and cancer cells as targets for therapeutic strategy. Cancer Sci.
99:1501–1506. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
31.
|
Jones KM, Wang H, Peskar BM, et al:
Inhibition of angiogenesis by nonsteroidal anti-inflammatory drugs:
insight into mechanism and implications for cancer growth ulcer
healing. Nat Med. 5:1418–1423. 1999. View
Article : Google Scholar : PubMed/NCBI
|
|
32.
|
Cragg G and Suffness M: Metabolism of
plant-derived anticancer agents. Pharmacol Ther. 37:425–461. 1988.
View Article : Google Scholar : PubMed/NCBI
|
|
33.
|
Wani MC, Taylor HL, Wall ME, Coggon P and
McPhail AT: Plant antitumor agents. VI. The isolation and structure
of taxol, a novel antileukemic and antitumor agent from Taxus
brevifolia. J Am Chem Soc. 93:2325–2327. 1971. View Article : Google Scholar : PubMed/NCBI
|
|
34.
|
Nagy JA, Dvorak AM and Dvorak HF:
EGF-A(164/165) and PIGF: roles in angiogenesis and arteriogenesis.
Trends Cardiovasc Med. 13:169–175. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
35.
|
Tsuda H, Ohshima Y, Nomoto H, et al:
Cancer prevention by natural compounds. Drug Metab Pharmacokinet.
19:245–263. 2004. View Article : Google Scholar
|
|
36.
|
Parikh AA and Ellis LM: The vascular
endothelial growth factor family and its receptors. Hematol Oncol
Clin North Am. 18:951–971. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
37.
|
Munoz-Chapuli R, Quesada AR and Angel
Medina M: Angiogenesis and signal transduction in endothelial
cells. Cell Mol Life Sci. 61:2224–2243. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
38.
|
Blavier L, Lazaryev A, Dorey F,
Shackleford GM and DeClerck TA: Matrix metalloproteinases play an
active role in Wnt1-induced mammary tumorigenesis. Cancer Res.
66:2691–2699. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
39.
|
Bisacchi D, Benelli R, Venzetto C, Ferrari
N, Tosetti F and Albini A: Anti-angiognesis and angioprevention:
mechanisms, problems and perspectives. Cancer Detect Prev.
27:229–238. 2003. View Article : Google Scholar : PubMed/NCBI
|