In HCV-related hepatocarcinogenesis, participation
of viral proteins: core (C), non-structural 3 (NS3) and NS5
proteins themselves used to be accentuated (1–3).
Several investigations point to a relationship between subcellular
localisation, concentration, specific molecular form of the
proteins and presence of specific domains and oncogenesis (4–7). HCV
proteins were found to be involved in control of cell cycle,
through their interaction with such cell cycle control proteins as
p53, p21, cyclins, proliferating cell nuclear antigen (PCNA),
transcription factors (mainly nuclear factor κB, NF-κB),
proto-oncogenes (c-fos, c-jun), growth
factors/cytokines, e.g. tumour necrosis factor (TNF)-α,
transforming growth factor (TGF)-β and their receptors and proteins
of apoptosis (8–13). A possible interactions between
oncogenic HCV proteins and components of IGF axis in human HCC
continue to be discussed (14).
Two key regulatory proteins of this axis are known:
IGF-1 and IGF-2, which manifest ∼50% sequence identical to that of
insulin (15). They are included
in the insulin-related family together with relaxin while genes of
the family members were located on distinct genomic fragments
(chromosome 2 and 11p-q13) (16).
To date, 6 types of the IGF BPs have been well characterised (IGF
BP1–6), plus two subsequent ones, less well recognised (IGF BP-7
and 8) (17–19). The basic functions of IGF BPs
include modulation of IGF-1 and IGF-2 bioactivity, mainly through
interactions with receptors of the factors and with insulin
receptor (IR). Moreover, IGF BPs extend half-life of IGFs in blood,
store them in selected tissue compartments and inhibit activity of
IGFs by lowering accessibility of their receptors. They may act
independently of the receptors, inducing mitogenesis and cell
migration (19). They participate
in the interactions with other growth factors (e.g. TGF-β)
(18). The most commonly
manifested circulating form of IGF BP, is IGF BP-3, which binds
over 95% of IGFs. The protein as a dimer forms a complex with the
acid-labile protein (ALS) subunit (18). IGF BP-2, -3 and -5 contain a
nuclear localisation signal and may influence on activity of
transcription. The IGF BP-3 itself may act also as an inhibitor of
cell growth (20,21). Free IGF-1 has a half-life of ∼8 min
in serum. This can be increased to ∼30 min if bound to IGF BP-3 and
to ∼15 h in the ternary complex with IGF BP-3 and ALS (22).
IGFs affect a cell specifically binding three
various surface receptors: IGF-1R, IGF-2R and IR. In most of
activities of IGF-1 and IGF-2, IGF-1R plays the main role of a
mediator. Apart from mediating in mitogenic and anti-apoptotic
activities of IGFs, the receptor is engaged in cell transformation
(23) and, therefore, this review
accentuates first of all the role of IGF-1R.
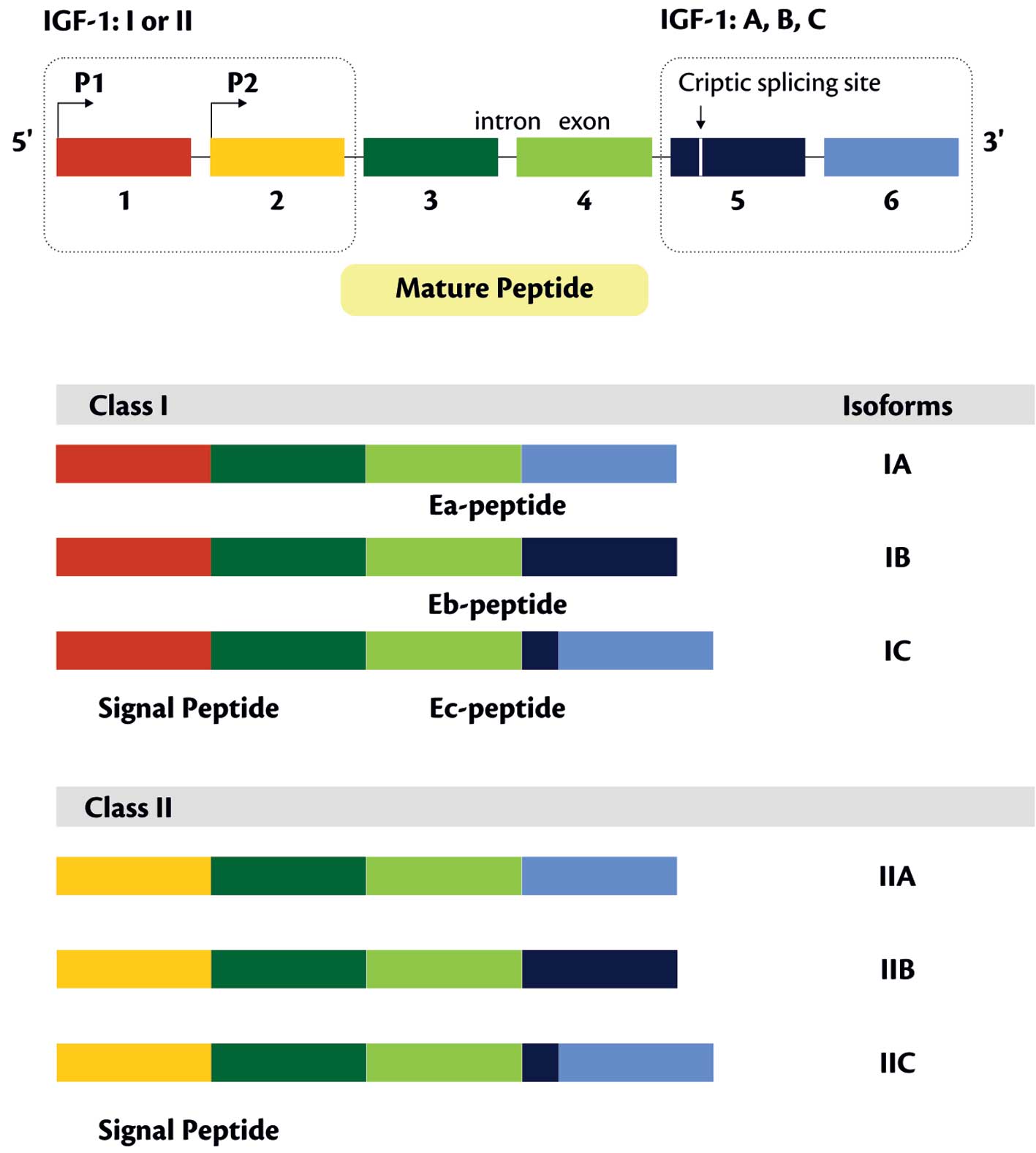
The nucleotide sequences of cDNAs predicting two
different IGF-1 protein precursors and defining the size of these
peptides (153 and 195 amino acids) were described for the first
time by Rotwein (34). In liver
cells IGF-1 isoform C (comprises 10% of IGF-1A isoform transcript)
was also detected. Its presence was confirmed also in cultured
HepG2 cells, in which its expression was increased following
supplementation of GH. The transcript codes for the prepro-IGF-1 of
158 amino acids (aa), with an E-peptide sequence of 24 aa (33).
IGF-1 is a secretory protein of 7649 Da molecular
mass, consisting of a single polypeptide chain with 70 aa, exerting
a variable effect on cells and tissues (17). In postnatal period liver remains to
be the main source of circulating IGF-1 and the protein is produced
mainly under effect of GH. Secretion of GH is affected also by age,
gender, diet and nutrition, insulin and sex hormones (17). Studies on RNA level demonstrated
that in adipose tissue amounts of the IGF-1 transcript are equal to
those in liver (17). IGF-1
produced in liver exerts mainly endocrine activity while IGF-1
synthetised by other tissues acts in a para- and/or autocrine way.
Interestingly, even if normal liver represents the organ with the
highest expression of IGF-1, it contains almost undetectable levels
of IGF-1R mRNA. In contrast to hepatocytes, presence of IGF-1R was
proven on Kupffer cells, myofibroblasts and hepatic stellate cells
(20).
Serum concentration of IGF-1 changes with patient’s
age and is dependent on gender. In childhood it grows
systematically, most rapidly before and during pubescence, when it
reaches the highest levels (53).
The gender-related difference in IGF-1 concentration (of ∼20 μg/l)
appears already in the first three years of age and it is most
pronounced (in girls higher by ∼70 μg/l) at the age of ∼11–13
years. Following 20th year of age a reverse change takes place and
higher IGF-1 concentrations are noted in men and the mean
inter-gender difference is 6–26 μg/l (54). Other investigators detected a
gradually decreasing with age concentrations of IGF-1, independent
of gender (55–57). Following a marked decrease in IGF-1
concentration following 25th year of life, a systematic slow
reduction in the level to 80th year of age is noted (53). Free IGF-1 accounted for ∼1% of the
total IGF-1 and its variation with age was similar to total IGF-1
(58).
Independently of GH level, serum concentration of
IGF-1 is affected by nutritional status. In childhood and during
pubescence no correlation was noted between IGF-1 concentration and
body mass index (BMI) (53). In
persons with BMI <21 and BMI >29 decreased levels of IGF-1
are noted (59). In cases of
obesity it achieves decreasingly lower values. Ther highest
standard deviation score (SDS) values for IGF-1 were noted in women
with BMI of 27.5 to 30, and in men with BMI ranging between 22.5
and 25 (60). In men of 40–75
years of age also race-dependent differences were noted in serum
concentration of IGF-1. Caucasians had the highest median IGF-1
level (224 ng/ml), followed by Asian (208 ng/ml) and African
Americans (205 ng/ml) (61).
In women an inverse correlation was documented
between IGF-1 and insulin levels (62). Low concentrations of IGF-1 were
proven to be strictly linked to growing risk of glucose intolerance
and development of type 2 diabetes mellitus (DM) (63). In DM IGF-1 levels decreases
(64) and manifests an inverse
correlation with concentration of glycosylated haemoglobin
(HbA1c) (65). Insulin
therapy of DM decreases portal concentrations of insulin. The
decreased concentrations of insulin may lead to an insufficient
production of IGF-1 in liver. In patients with DM concentration of
IGF BP-1 increases also (65,66).
In rats with induced diabetes, reductions in circulating IGF-1
levels and hepatic IGF-1 mRNA were noted as well as a significant
increase (>400%) in liver IGF BP-1 mRNA (67). IGF-1 stimulates glucose uptake by
peripheral tissues. This insulin-like effect may develop with
mediation of IGF-1R or IR (68).
Concentrations of IGF-1 and IGF BP-3 are stable
during a day even if the main factor which stimulates production
and secretion of the two proteins is GH. In children and youth
concentrations of IGF-1 and IGF BP-3 well correlate with 24 h
output of GH and reflect spontaneous secretion of GH in healthy
individuals. Levels of IGF-1 are more sensitive to GH control than
levels of IGF BP-3 (69).
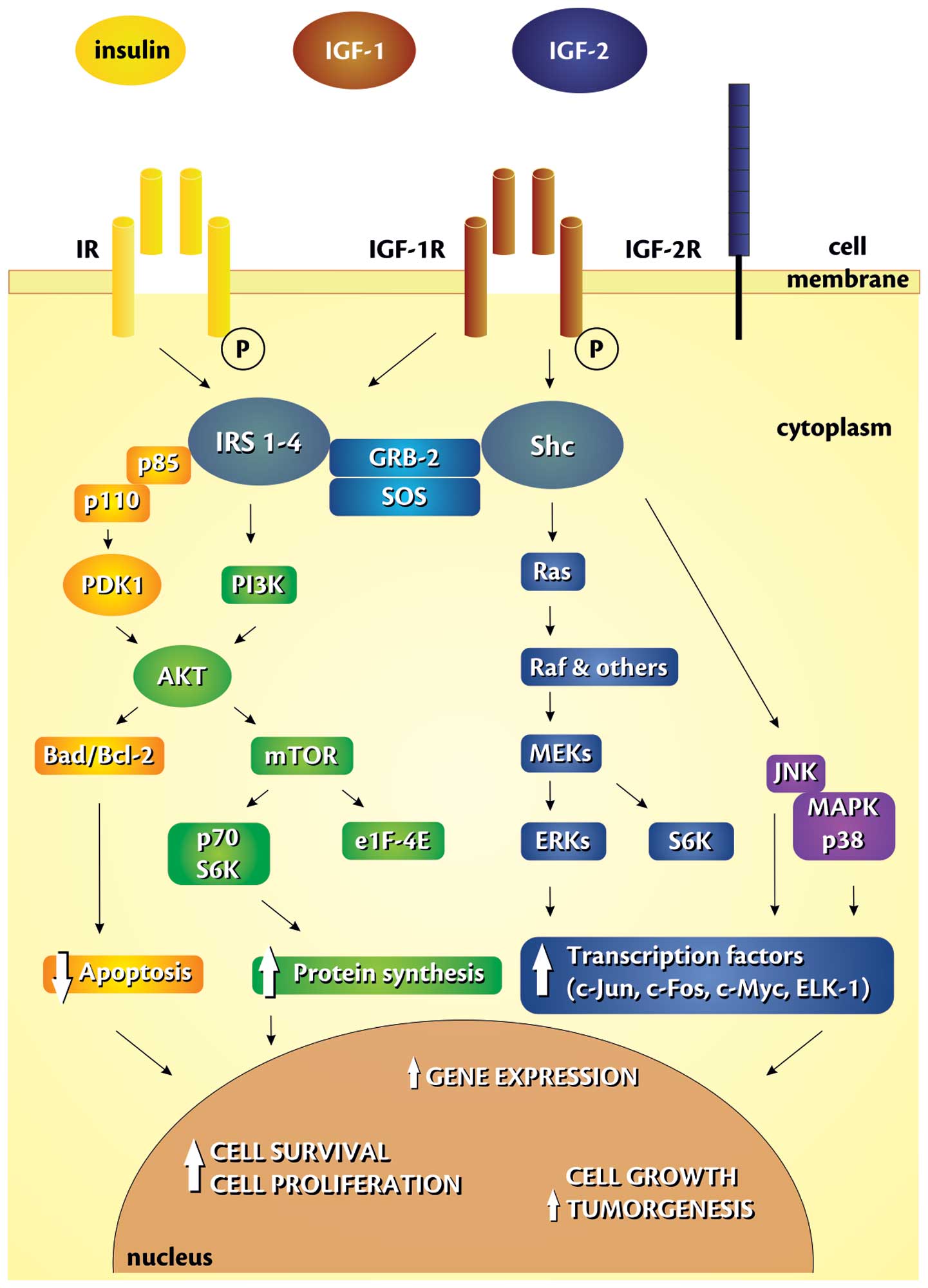
IGF-1 binds with IGF-1R to stimulate cellular
proliferation or inhibit apoptosis through different pathways,
increasing the risk of carcinogenesis (76–78)
(Fig. 2). IGF-1R/IGF axis may
positively control cell cycle progression in many phases, but the
major direct effect is probably exerted at the G1-S interface, and
this is mediated through the phosphatidylinositol
3-kinase/serine/threonine-specific protein kinase (PI-3K/Akt)
and/or extracellular signal-regulated kinase (ERK) pathways
(79). IGF-1 exerts a mitogenic
effect influencing stimulation of cyclin D1 and of some
proto-oncogenes (c-FOS, c-JUN) expression. Moreover, it acts
anti-apoptotically and modulates body immune response by control of
cytokine production (e.g. IL-3 and IL-14) (68). It was also demonstrated that short
peptides of IGF-1 precursors may promote growth of normal and
malignant cells in bronchial epithelium (17). IGF-1 controls expression of over 50
genes linked to mitogenesis and cell differentiation. However, it
exerts mitogenic activity mainly through stimulation of DNA
synthesis and of cyclin D1 expression (68).
Hepatocellluar carcinoma is the third leading cause
of cancer-related death worldwide (14,84).
The suggested roles of both IGFs and IGF-1R in development of HCC
reflect, first of all, observations indicating strong mitogenic
effects of the factors in in vitro conditions (21,85,86).
Different types of cultured cells produce IGF-1 mRNA (87) and insulin receptor substrate 1
(IRS-1) (86). IRS-1 undergoes
overexpression in human HCC (88,89).
The dominating pathways activated by IGF-1 in hepatocytes and
hepatoma cell lines involve the PI3K/Akt (90,91)
and signal transducer and activator family protein (STAT) signaling
pathways (92). IGF-1 has been
implicated in NF-κB-mediated transcriptional regulation of
inflammatory cytokines and endothelial cell adhesions receptors
such as intracellular adhesion molecule-1 (ICAM-1) (93). Studies on HepG2 and HuH-7 cells
demonstrated that the cells synthetised and secreted also IGF-2 and
inhibition of the protein production resulted in a reduced cellular
proliferation (87). Recent
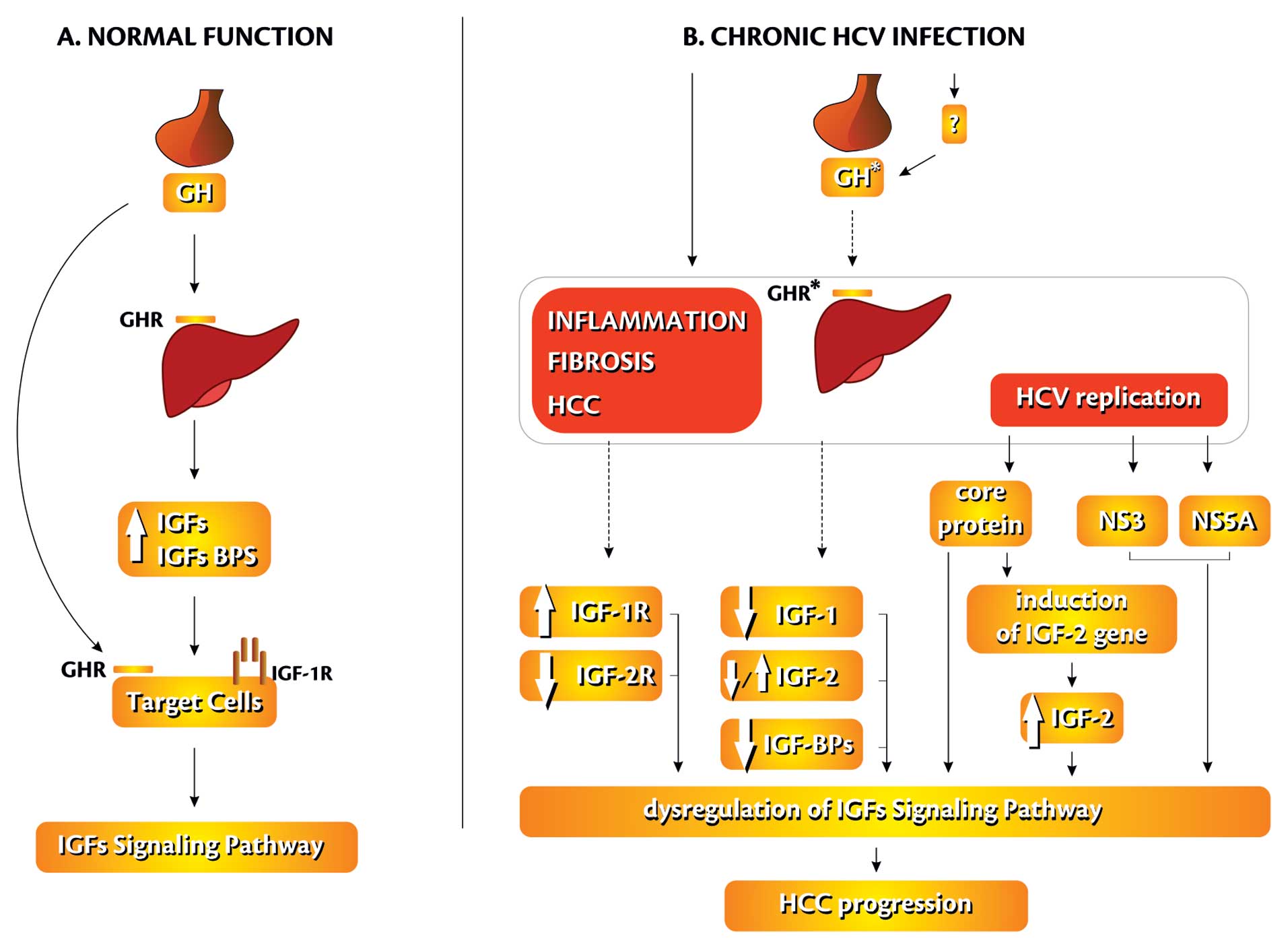
studies on HCV-related HCC demonstrated overexpression of
IGF-2 gene (resulting from reactivation of fetal promoters
P3 and P4), IGF BP3 down-regulation, decreased IGF-1 expression and
allelic losses of IGF-2R. Administration of IGF-1R-selective
inhibitors (A12) reduced IGF-1-induced effects and was associated
with a significant reduction of liver tumour growth (84) (Fig.
3).
A decreased serum levels of both IGFs were found to
parallel progression of liver diseases, independently of their
etiology (94,95). Besides, patients with HCC and
hypoglycaemia were found to carry higher IGF-2 concentrations than
patients without hypoglycaemia. The higher IGF-2 than those of
IGF-1 levels were accompanied by more advanced histological lesions
in the liver (94). Kratzsch et
al demonstrated a continuous decline in the concentrations of
IGF-1, IGF BP-3 and serum GH-binding activity during progression of
cirrhosis (95). Other
investigations documented decreased levels of both IGFs in chronic
liver diseases but the positive correlation with Child score was
confirmed only for IGF-2 (96). It
should be added that only in a small number of studied patients
(17/55) viral etiology of chronic lesions in liver was confirmed
(96). In chronic hepatitis B and
C were also demonstrated elevated levels of IGF-1 and lowered
levels of IGF BP-3 (97). Studies
on serum IGF-1 concentrations in HCC and in metastatic liver
cancers demonstrated markedly lowered concentrations of IGF-1 in
either type of hepatic tumour, as compared to the control (98). The decrease in IGF-1 was more
pronounced in cases with viral-associated than in virus-negative
HCCs (98). In liver cirrhosis
lower IGF-1 concentrations were observed in comparison with control
(99,100), as well as its increased
concentration following anti-viral therapy (100). In HCV-infected children serum
IGF-1 level was significantly lower compared with the control group
(101). In adult patients with
chronic hepatitis C the reduction of serum IGF-1 level was found to
precede by ∼9 months development of HCC (102).
Similarly to IGF-1, concentration of IGF-2 also
markedly decreases in patients with liver cirrhosis, as compared to
healthy individuals. Advancement of liver cirrhosis, is paralleled
by gradual decreases in IGF-2 levels. The lowest concentrations of
the factor are encountered in patients with incurable ascites and
extended activated partial tromboplastin time (APTT). During
half-a-year observation of the patients, those with very low
concentrations of IGF-2 (<200 ng/ml) died manifesting hepatic
insufficiency (103). A
successful liver transplantation was followed by a significant
increase in IGF-2 concentration in almost all of the patients
(104). In patients with HCC
levels of IGF-2 decrease (but the difference was not statistically
significant) and thus the protein cannot serve as a diagnostic
marker of the tumour (105). The
amount of IGF-2 mRNA was also lowered in patients with liver
cirrhosis as compared to the control (106). The studies on rats with
experimentally induced liver cancer, demonstrated a decreased
expression of IGF-1 mRNA, and an increased expression of IGF-2 mRNA
(107). Start of IGF-2 (fetal
type of IGF expression) production in HCC is supposed to represent
a late phenomenon in carcinogenesis of rats (107). Regarding human HCC activation of
IGF axis (e.g. IGF-2 overexpression, IGF-1R activation) was
observed in 21% of early liver tumours (83).
Role of IGFs receptors in liver carcinogenesis
remains to be fully recognised. On one hand there exist papers
pointing to an increased expression of IGF-1R in preneoplastic
focal lesions in liver, leading to HCC, in HCC itself and in human
hepatoma cells (20), on the other
attempts to stimulate mitogenesis with IGF-1 in cultured rat cell
lines of HCC proved to be unsuccessful (109). Hepatitis B virus (HBV) and HCV
are known to be capable of activating promoters of IGF-2 and
IGF-2R genes, thereby increasing the production of this
ligand and receptor by liver cancer cells (110–112). An increase in IGF-1R mRNA in
liver cirrhosis has been demonstrated, but no change in the amount
of IGF-2R mRNA was detected, as compared to the control (106). Activation of IGF-1R was
significantly associated with activation of IGF-2 mRNA levels in
∼20% of HCCs (84). Other authors
observed decreased levels of IGF-1R in 39% patients with cirrhosis,
as compared to the control (21).
Decreased levels of IGF BP-3 were shown in patients
with cirrhotic liver, as compared to the control (115). The lowered serum concentration of
IGF BP-3 was accompanied by absence of protease activity in
cirrhotic patients (115,119). It should be noted that
concentration of IGF BP-3 is also dependent on age (120,121). Serum concentrations increase with
progressing age to reach maximum levels at pubescence. Comparison
of IGF BP-3 and IGF-1 concentrations revealed that they did not
exibit the same developmental pattern at puberty. IGF-1 levels
increased to relatively higher levels than IGF BP-3 (120). Median IGF BP-3 concentration was
similar between Caucasian and Asian middle-aged men but was more
than 13% lower in African Americans. Median molar IGF-1:IGF BP-3
ratio was greatest in Caucasian and lowest in Asians (61). IGF BP-3 levels manifested no
differences before and after menopause (122). Also, concentration of IGF BP-3
failed to correlate with BMI (117). A relationship was detected
between lower levels of serum IGF-1, decreased IGF-1/IGF BP-3
ratio, higher serum concentrations of IGF BP-3 on one hand and
liver steatosis on the other (116).
IGF BP-6 level significantly increases with age
beginning at birth up to pubescence. In most age groups adult men
manifested higher concentrations of IGF BP-6 than those noted in
women (58).
In liver cirrhosis significantly lower serum level
of IGF BP-3 is noted, as compared to healthy individuals (103). Also, it becomes reduced with an
advancement in cirrhosis. Therefore, concentration of this protein
provides a real potential index of progression manifested by
hepatic lesions (103,117). A positive correlation was
detected between concentration of IGF BP-3 and of albumin as well
as a negative correlation with bilirubin concentration, size of
spleen and activity of aspartate aminotransferase (AST) (117,121). Concentration of IGF BP-3 return
to normal level following a successful liver transplantation
(104). In patients with HCC
concentrations of IGF BP-3 are decreased in cases with a disturbed
nutrition, markedly deteriorated liver function and reduced
secretion of GH (105). An
increase in IGF-1/IGF BP-3 ratio in patients with HCC was also
demonstrated as compared to patients with cirrhotic liver and a
similar extent of liver insufficiency (118). In >70% HCC patients, IGF BP-3
expression was lowered as compared to normal liver. A less
pronounced tissue expression of the protein was observed in tumour
cells as compared to control (21). Decreased IGF BP-3 mRNA levels
correlated with smaller tumour size, less vascular invasion, and a
lower incidence of early recurrence (84).
Analysis of gene expression using cDNA microarrays
performed in 20 primary liver tumours demonstrated 170 genes with a
decreased regulation in HCC, including IGF BP-3 and ALS (123). In turn, application of 75
antibodies to evaluate markers for early detection of
viral-associated HCC, identified 7 proteins which significantly
differentiated patients with HCC and those with chronic hepatitis,
including IGF BP-6 (83).
A lowered serum concentration of IGF BP-3 was noted
also in patients with variably advanced chronic hepatitis
(including 12 HCV-positive patients). No correlation could be
disclosed between grading/staging on one hand and activity of
transaminases on the other (97).
Other investigators failed to detect significant differences
between concentration of IGF BP-3 in patients with liver cirrhosis
which would be related to its aetiology (alcohol vs. HBV vs. HCV
(117). Other studies
demonstrated a significantly decreased concentration of IGF BP-3 in
patients with chronic hepatitis C as compared to healthy
individuals (121).
In all studied tissues of HCC (n=28) a decreased
expression of IGF-1 and IGF BP-3 and in 32% of them an increased
expression of IGF-2 was detected (21). Decreased amounts of IGF-1 mRNA
(84) or both IGF’s mRNA (124–126) in HCCs were demonstrated as
compared to the control. In case of IGF-1 transcript, the decrease
was more pronounced than that in IGF-2 (124–126). In hepatocarcinogenesis in rats
appearance of IGF-2 mRNA was demonstrated upon a decrease in IGF-1
mRNA (107). An increased
expression of IGF-2 (protein and mRNA) was demonstrated in
HBV-related HCC (127). IGF-1,
IGF-2 and IR mRNAs were detected at various stages of HCC
development and in cultured cells. The studies showed that isolated
hepatocytes synthetise IGF-2 mRNA with a switch between fetal and
adult mRNA profiles occurring 21 days after birth (128).
Other studies demonstrated presence of IGF-2 in 60%
HCC arising in HBV-associated cirrhosis, as compared to only 26%
tumours in HBV-negative patients (129). Studies employing DNA microarrays
showed that imbalances in levels of IGF-2 and H19 transcripts were
correlated with advanced tumour stage and poor outcome in HCC
patients (130). High focal
levels of IGF-2 mRNA were found in some hepatocytes of all HBV and
HCV-related cirrhotic livers (131). In subgroups of HCV-related HCC an
increased tissue expression of IGF-2 (protein, mRNA) was noted
(20,83,111). Compared with noncirrhotic liver,
all cirrhotic specimens showed reduced expression of IGF-2R/M6PR
protein. It was suggested that downregulation of hepatocellular
IGF-2R/M6PR and upregulation of IGF-2 might be early events in
hepatocarcinogenesis (131).
Tovar et al observed losses in the IGF2R locus in ∼25% of
HCCs (84).
The increased expression of IGF-1R was detected in
cirrhosis, in HCC and in human hepatoma cells, as compared to
normal liver (84,86,87,132,133). More pronounced membraneous
location of IGF-1R was documented in HepG2 cells, as compared to
Chang liver cell lines (134).
Overexpression of IGF-1R in the cell lines was supposed to indicate
a malignant phenotype of the cells. Administration of anti-IGF-1R
monoclonal antibody (αIR3) inhibited cell proliferation and induced
apoptosis in HepG2 cells (134).
It has been proven that mutations resulting in upregulation of
IGF-1R gene in certain HCC include p53mt249 (135). The same mutation enhances
transcription from fetal IGF-2 promoter P4 (136). Recent studies have drawn
attention to the role of polymorphism in IGF-1 gene promoter
in carcinogenesis. One of IGF-1 gene polymorphisms
recognised in greatest detail seems to involve the sequence
consisting of several CA repeats, present in 5′UTR of IGF-1
gene (41). The mutations of
IGF-2R/M6PR gene give rise to truncated receptor protein and
significant aa substitutions, and provide evidence that this gene
functions as a tumour suppressor in human liver carcinogenesis
(137).
Serum concentrations of IGF-1 were significantly
lower in HCV-associated HCC than in healthy subjects. Moreover, the
lower levels of IGF-1 were detected in all patients below 55th year
of age and with homeostasis model assessment of insulin resistance
(HOMA-IR) <2.53 as compared to the control (138). Moreover in >80% of
HCV-infected patients, severe GH insufficiency was documented and
about half of these patients had low IGF-1 level. Basal and
stimulated GH concentration increased significantly during therapy,
but IGF-1 remained low (139).
Only few investigations pertained hepatic
expression of IGFs and their receptors at different stages of
chronic hepatitis C in humans (144). The studies demonstrated an
increased IGF-1R mRNA, but not IGF-1 protein in patients with CHC
as compared to the control (144). HCV replication was associated
with the overexpression of IGF-2 in the cirrhotic livers (145). Studies on HCV-related HCCs
documented increased expression of IGF-2 in 50% of HCCs. The
increased synthesis of IGF-2 (mRNA and protein) was associated with
an increased cellular proliferation in HCCs (146). The results observed in tissue
samples suggest that IGF-2 might be responsible for IGF-1R
activation (83).
Data from examination of serum and tissue levels
involving IGF axis components in the patients with advanced liver
diseases (including HCV-associated HCC) are summarized in Tables I and II.
IGF-1 may play a role both in persistence of
chronic hepatic inflammation through control of signaling pathways
linked to proinflammatory cytokines and receptors for endothelial
adhesion molecules (e.g. ICAM-1) (93), and in induction of acute
inflammatory reaction, triggered by tumour cells during early
stages of liver metastasis (79,147,148). In recent years increased
attention is devoted also to complex interactions between HCV
proteins and IGF axis. HCV core protein was shown to increase
endogenous expression of IGF-2 in HepG2 cell line, regulating
positively its transcription, and it may promote cell divisions
(149).
Our own studies in chronic hepatitis C patients
demonstrated positive correlation between tissue expression of
IGF-1 and two HCV proteins: core and NS3 while studies on tissue
material originating from HCC detected no significant correlations
between tissue expression of IGF-1 and the histological malignancy
of the tumour (150).
Several components of the IGF signaling axis, such
as IGF-1, IGF-2 and IGF-1R, are dysregulated during HCV-related
human HCC. Only few investigations pertained hepatic expression of
IGFs and their receptors at different stages of chronic hepatitis
C. The studies demonstrated an increased IGF-1R synthesis, the
abberant IGF-2 expression (decreased/increased), and decreased
synthesis of IGF-1 as an events in human hepatocarcinogenesis.
Recognition of the role played by HCV in different splicing
profiles of IGF-1 gene in progression of chronic hepatitis C
will require further studies. Better understanding of the HCV
protein and IGF axis component interactions will facilitate
development of novel approaches to prognose and to treat the
virus-related HCC.
The work was supported by a grant
(no. NN401009437) from the Minister of Education and Science,
Warsaw, Poland. The authors thank Dr Marek Konkol for assistance
with the color figures.
|
1.
|
Anzola M: Hepatocellular carcinoma: role
of hepatitis B and hepatitis C viruses proteins in
hepatocarcinogenesis. J Viral Hepat. 11:383–393. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
2.
|
Kasprzak A and Adamek A: Role of hepatitis
C virus proteins (C, NS3, NS5A) in hepatic oncogenesis. Hepatol
Res. 38:1–26. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
3.
|
Wang F, Yoshida I, Takamatsu M, Fujita T,
Oka K and Hotta H: Complex formation between hepatitis C virus core
protein and p21Waf1/Cip1/Sdi1. Biochem Biophys Res Commun.
273:479–484. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
4.
|
Chang SC, Yen JH, Kang HY, Jang MH and
Chang MF: Nuclear localization signals in the core protein of
hepatitis C virus. Biochem Biophys Res Commun. 205:1284–1290. 1994.
View Article : Google Scholar : PubMed/NCBI
|
|
5.
|
Suzuki R, Sakamoto S, Tsutsumi T, et al:
Molecular determinants for subcellular localization of hepatitis C
virus core protein. J Virol. 79:1271–1281. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
6.
|
Yamanaka T, Uchida M and Doi T: Innate
form of HCV core protein plays an important role in the
localization and the function of HCV core protein. Biochem Biophys
Res Commun. 294:521–527. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
7.
|
Nielsen SU, Bassendine MF, Burt AD, Bevitt
DJ and Toms GL: Characterization of the genome and structural
proteins of hepatitis C virus resolved from infected human liver. J
Gen Virol. 85:1497–1507. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
8.
|
Cho JW, Baek WK, Suh SI, Yang SH, Chang J,
Sung YC and Suh MH: Hepatitis C virus core protein promotes cell
proliferation through the upregulation of cyclin E expression
levels. Liver. 21:137–142. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
9.
|
Jung EY, Lee MN, Yang HY, Yu D and Jang
KL: The repressive activity of hepatitis C virus core protein on
the transcription of p21(waf1) is regulated by protein kinase
A-mediated phosphorylation. Virus Res. 79:109–115. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
10.
|
Kwun HJ, Jung EY, Ahn JY, Lee MN and Jang
KL: p53-dependent transcriptional repression of p21 (waf1) by
hepatitis C virus NS3. J Gen Virol. 82:2235–2241. 2001.PubMed/NCBI
|
|
11.
|
Taniguchi H, Kato N, Otsuka M, Goto T,
Yoshida H, Shiratori Y and Omata M: Hepatitis C virus core protein
upregulates transforming growth factor-beta 1 transcription. J Med
Virol. 72:52–59. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
12.
|
Sato Y, Kato J, Takimoto R, et al:
Hepatitis C virus core protein promotes proliferation of human
hepatoma cells through enhancement of transforming growth factor
alpha expression via activation of nuclear factor-kappaB. Gut.
55:1801–1808. 2006. View Article : Google Scholar
|
|
13.
|
Kasprzak A, Adamek A, Biczysko W, et al:
Intracellular expression of the proliferative marker Ki-67 and
viral proteins (NS3, NS5A and C) in chronic, long lasting hepatitis
C virus (HCV) infection. Folia Histochem Cytobiol. 45:357–366.
2007.PubMed/NCBI
|
|
14.
|
Breuhahn K, Longerich T and Schirmacher P:
Dysregulation of growth factor signalling in human hepatocellular
carcinoma. Oncogene. 25:3787–3800. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
15.
|
Daughday WH and Rotweien P: Insulin-like
growth factors I and II. Peptide, messenger ribonucleic acid and
gene structures, serum and tissue concentrations. Endocr Rev.
10:68–91. 1989. View Article : Google Scholar : PubMed/NCBI
|
|
16.
|
Rotwein P, Naylor SL and Chirgwin JM:
Human insulin-related DNA sequences map to chromosomes 2 and 11.
Somat Cell Mol Genet. 12:625–631. 1986. View Article : Google Scholar : PubMed/NCBI
|
|
17.
|
Zarilli R, Bruni CB and Riccio A: Multiple
levels of control of insulin-like growth factor gene expression.
Mol Cell Endocrinol. 101:R1–R14. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
18.
|
Murphy LJ: Insulin-like growth
factor-binding proteins: functional diversity or redundancy? J Mol
Endocrinol. 21:97–107. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
19.
|
Kostecka Z and Blahovec J: Insulin-like
growth factor binding proteins and their functions (minireview).
Endocr Regul. 33:90–94. 1999.PubMed/NCBI
|
|
20.
|
Scharf JG, Dombrowski F and Ramadori G:
The IGF axis and hepatocarcinogenesis. Mol Pathol. 54:138–144.
2001. View Article : Google Scholar : PubMed/NCBI
|
|
21.
|
Huynh H, Chow PKH, Ooi LLP and Soo KC: A
possible role for insulin-like growth factor-binding protein-3
autocrine/paracrine loops in controlling hepatocellular carcinoma
cell proliferation. Cell Growth Differ. 13:115–122. 2002.
|
|
22.
|
Rechler MM and Clemmonds DR: Regulatory
actions of insulin-like growth factor-binding proteins. Trends
Endocrinol Metab. 9:176–183. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
23.
|
Baserga R: The insulin-like growth factor
I receptor: a key to tumor growth? Cancer Res. 55:249–255.
1995.PubMed/NCBI
|
|
24.
|
Höppener JW, de Pagter-Holthuizen P,
Geurts van Kessel AHM, et al: The human gene encoding insulin- like
growth factor I is located on chromosome 12. Hum Genet. 69:157–160.
1985.PubMed/NCBI
|
|
25.
|
Bell GI, Stempien MM, Fong NM and Rall LB:
Sequences of liver cDNAs encoding two different mouse insulin-like
growth factor 1 precursors. Nucleic Acids Res. 14:7873–7882. 1986.
View Article : Google Scholar : PubMed/NCBI
|
|
26.
|
Adamo ML: Regulation of insulin-like
growth factor I gene expression. Implications for normal and
pathological growth. Diabetes Rev. 3:2–27. 1995.
|
|
27.
|
Temmerman L, Slonimsky E and Rosenthal N:
Class 2 IGF-1 isoforms are dispensabile for viability, growth and
maintenance of IGF-1 serum levels. Growth Horm IGF Res. 20:255–263.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
28.
|
Matheny RW Jr, Nindl BC and Adamo ML:
Minireview: Mechano-Growth Factor: a putative product of IGF-I gene
expression involved in tissue repair and regeneration.
Endocrinology. 151:865–875. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
29.
|
Adamo ML, Ben-Hur H, LeRoith D and Roberts
CT Jr: Transcription initiation in the two leader exons of the rat
IGF-1 gene occurs from disperse versus localized sites. Biochem
Biophys Res Commun. 176:887–893. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
30.
|
Simmons JG, Van Wyk JJ, Hoyt EC and Lund
PK: Multiple transcription start sites in the rat insulin-like
growth factor-I gene give rise to IGF-I mRNAs that encode different
IGF-I precursors and are processed differently in vitro. Growth
Factors. 9:205–221. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
31.
|
Jansen M, van Schaik FM, Ricker AT, et al:
Sequence of cDNA encoding human insulin-like growth factor I
precursor. Nature. 306:609–611. 1983. View Article : Google Scholar : PubMed/NCBI
|
|
32.
|
Roberts CT Jr, Lasky SR, Lowe WI Jr,
Seaman WT and LeRoith D: Molecular cloning of rat insulin-like
growth factor I complementary deoxyribonucleic acids: differential
messenger ribonucleic acid processing and regulation by growth
hormone in extrahepatic tissues. Mol Endocrinol. 1:243–248. 1987.
View Article : Google Scholar
|
|
33.
|
Chew SL, Lavender P, Clark AJ and Ross RJ:
An alternatively spliced human insulin-like growth factor-1
transcript with hepatic tissue expression that diverts away from
mitogenic IBE1 peptide. Endocrinology. 136:1939–1944.
1995.PubMed/NCBI
|
|
34.
|
Rotwein P: Two insulin-like growth factor
I messenger RNAs are expressed in human liver. Proc Natl Acad Sci
USA. 83:77–81. 1986. View Article : Google Scholar : PubMed/NCBI
|
|
35.
|
Shavlakadze T, Winn N, Rosenthal N and
Grounds MD: Reconciling data from transgenic mice that overexpress
IGF-I specifically in skeletal muscle. Growth Horm IGF Res.
15:4–18. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
36.
|
Winn N, Paul A, Musaro A and Rosenthal N:
Insulin-like growth factor isoforms in skeletal muscle aging,
regeneration, and disease. Cold Spring Harb Symp Quant Biol.
67:507–518. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
37.
|
Rotwein P: Molecular biology of IGF-1 and
IGF-2. The IGF System. Rosenfeld RG and Roberts CT Jr: Humana
Press; Totowa, NJ: pp. 19–35. 1999, View Article : Google Scholar
|
|
38.
|
Siegfried JM, Kasprzyk PG, Treston AM,
Mulshine JL, Quinn KA and Cuttitta F: A mitogenic peptide amide
encoded within the E peptide domain of the insulin-like growth
factor IB prohormone. Proc Natl Acad Sci USA. 89:8107–8111. 1992.
View Article : Google Scholar : PubMed/NCBI
|
|
39.
|
Barton ER: The ABCs of IGF-I isoforms:
impact on muscle hypertrophy and implications for repair. Appl
Physiol Nutr Metab. 31:791–797. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
40.
|
Barton ER: Viral expression of
insulin-like growth factor-1 isoforms promotes different responses
in skeletal muscle. J Appl Physiol. 100:1778–1784. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
41.
|
Rotwein P, Pollock KM, Didier DK and Krivi
GG: Organization and sequence of the human insulin-like growth
factor I gene. Alternative RNA processing produces two insulin-like
growth factor I precursor peptides. J Biol Chem. 261:4828–4832.
1986.PubMed/NCBI
|
|
42.
|
Bach MA, Roberts CT Jr, Smith EP and
LeRoith D: Alternative splicing produces messenger RNAs encoding
insulin-like growth factor-I prohormones that are differentially
glycosylated in vitro. Mol Endocrinol. 4:899–904. 1990. View Article : Google Scholar : PubMed/NCBI
|
|
43.
|
Hameed M, Lange KH, Andersen JL,
Schjerling P, Kjaer M, Harridge SD and Goldspink G: The effect of
recombinant human growth hormone and resistance training of IGF-I
mRNA expression in the muscles of elderly men. J Physiol.
555:231–240. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
44.
|
Tan DS, Cook A and Chew SL: Nucleolar
localization of an isoform of the IGF-I precursor. BMC Cell Biol.
3:172003. View Article : Google Scholar
|
|
45.
|
Adamo ML, Neuenschwander S, LeRoith D and
Roberts CT Jr: Structure, expression, and regulation of the IGF-I
gene. Adv Exp Med Biol. 343:1–11. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
46.
|
Wang X, Yang Y and Adamo ML:
Characterization of the rat insulin-like growth factor I gene
promoters and identification of a minimal exon 2 promoter.
Endocrionology. 138:1528–1536. 1997.PubMed/NCBI
|
|
47.
|
Mittanck DW, Kim SW and Rotweien P:
Essential promoter elements are located within the 5′ untranslated
region of human insulin-like growth factor-I exon I. Mol Cell
Endocrinol. 126:153–163. 1997.
|
|
48.
|
Nolten LA, van Schaik FM, Steenbergh PH
and Sussenbach JS: Expression of the insulin-like growth factor I
gene is stimulated by the liver-enriched transcription factors
C/EBP alpha and LAP. Mol Endocrinol. 8:1636–1645. 1994.PubMed/NCBI
|
|
49.
|
Nolten LA, Steenbergh PH and Sussenbach
JS: Hepatocyte nuclear factor 1 alpha activates promoter 1 of the
human insulin-like growth factor I gene via two distinct binding
sites. Mol Endocrinol. 9:1488–1499. 1995.PubMed/NCBI
|
|
50.
|
Armakolas A, Philippou A, Panteleakou Z,
Nezos A, Sourla A, Petraki C and Koutsilieris M: Preferential
expression of IGF-IEc (MGF) transcript in cancerous tissues of
human prostate: evidence for a novel and autonomous growth factor
activity of MGF E peptide in human prostate cancer cells. Prostate.
70:1233–1242. 2010. View Article : Google Scholar
|
|
51.
|
Koczorowska MM, Kwasniewska A and
Gozdzicka-Jozefiak A: IGF1 mRNA isoform expression in the cervix of
HPV-positive woman with pre-cancerous and cancer lesions. Exp Ther
Med. 2:149–156. 2011.PubMed/NCBI
|
|
52.
|
Górecki DC, Beresewicz M and Zabłocka B:
Neuroprotective effects of short peptides derived from the
Insulin-like growth factor 1. Neurochem Int. 51:451–458.
2007.PubMed/NCBI
|
|
53.
|
Juul A, Bang P, Hertel NT, et al: Serum
insulin-like growth factor-I in 1030 healthy children, adolescents,
and adults: relation to age, stage of puberty, testicular size, and
body mass index. J Clin Endocrinol Metabol. 78:744–752.
1994.PubMed/NCBI
|
|
54.
|
Brabant G, von zur Muhlen A, Wurster Ch,
et al: Serum Insulin-like Growth factor I reference values for an
automated chemiluminescence immunoassay system: results from a
multi-center study. Horm Res. 60:53–60. 2003. View Article : Google Scholar
|
|
55.
|
Aimaretti G, Boschetti M, Corneli G, et
al: Normal age-dependent values of serum insulin growth factor-I:
results from a healthy Italian population. J Endocrinol Invest.
31:445–449. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
56.
|
Rosario P: Normal values of serum IGF-1 in
adults: results from a Brazilian population. Arq Bras Endocrinol
Metab. 54:477–481. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
57.
|
Andreassen M, Nielsen K, Raymond I,
Kristensen LØ and Faber J: Characteristics and reference ranges of
Insulin-Like Growth factor-I measured with a commercially avaible
immunoassay in 724 healthy adult Caucasians. Scand J Clin Lab
Invest. 69:880–885. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
58.
|
Yu H, Mistry J, Nicar MJ, Khosravi MJ,
Diamandis A, van Doorn J and Juul A: Insulin-like growth factors
(IGF-I, free IGF-I, and IGF-II) and insulin-like growth factor
binding proteins (IGFBP-2, IGFBP-3, IGFBP-6, and ALS) in blood
circulation. J Clin Lab Anal. 13:166–172. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
59.
|
Holmes MD, Pollak MN and Hankinson SE:
Lifestyle correlates of plasma insulin-like growth factor I and
insulin-like growth factor binding protein 3 concentrations. Cancer
Epidemiol Biomarkers Prev. 11:862–867. 2002.
|
|
60.
|
Schneider HJ, Saller B, Klotsche J, März
W, Erwa W, Wittchen HU and Stalla GK: Oposite associations of
age-dependent insulin-like growth factor-I standard deviation
scores with nutritional state in normal weight and obese subject.
Eur J Endocrinol. 154:699–706. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
61.
|
Platz EA, Pollak MN, Rimm EB, Majeed N,
Tao Y, Willett WC and Giovannucci E: Racial variation in
insulin-like growth factor-1 and binding protein-3 concentrations
in middlle-aged men. Cancer Epidemiol Biomarkers Prevent.
8:1107–1110. 1999.PubMed/NCBI
|
|
62.
|
Lecomte P, Lecureuil N, Lecureuil M,
Lemonnier Y, Mariotte N, Valat C and Garrigue MA: Sex differences
in the control of sex-hormone-binding globulin in the elderly: role
of insulin-like growth factor-I and insulin. Eur J Endocrinol.
139:178–183. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
63.
|
Sandhu MS, Heald AH, Gibson JM,
Cruickshank JK, Dunger DB and Wareham NJ: Circulating
concentrations of insulin like growth factor-I and development of
glucose intolerance: a prospective observational study. Lancet.
359:1740–1745. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
64.
|
Sesti G, Sciacqua A, Cardellini M, et al:
Plasma concentration of IGF-I is independently associated with
insulin sensitivity in subjects with different degrees of glucose
tolerance. Diabetes Care. 28:120–125. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
65.
|
Livingstone C and Ferns G: Insulin-like
growth factor-related proteins and diabetic complications. Br J
Diabetes Vasc Dis. 3:326–331. 2003. View Article : Google Scholar
|
|
66.
|
Froesch E, Hussain M, Schmid Ch and Zapf
J: Insulin-like growth factor I: physiology, metabolic effects and
clinical uses. Diabetes Metab Rev. 12:195–215. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
67.
|
Pao CI, Farmer PK, Begovic S, Goldstein S,
Wu GJ and Philips LS: Expression of hepatic insulin-like growth
factor-I and insulin-like growth factor-binding protein-1 genes is
transcriptionally regulated in streptozotocin-diabetic rats. Mol
Endocrinol. 6:969–977. 1992.PubMed/NCBI
|
|
68.
|
LeRoith D, Bondy C, Yakar S, Liu JL and
Butler A: The somatomedin hypothesis: 2001. Endocr Rev. 22:53–74.
2001. View Article : Google Scholar
|
|
69.
|
Blum W, Albertsson-Wikland K, Rosberg S
and Ranke M: Serum levels of insulin-like growth factor (IGF-I) and
IGF binding protein 3 reflect spontaneous growth hormone secretion.
J Clin Endocrinol Metabol. 76:1610–1616. 1993.PubMed/NCBI
|
|
70.
|
Khandwala HM, McCutcheon IE, Flyvbjerg A
and Friend KE: The effects of insulin-like growth factors on
tumorigenesis and neoplastic growth. Endocr Rev. 21:215–244. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
71.
|
O’Dell SD and Day IN: Insulin-like growth
factor II (IGF-II). Int J Biochem Cell Biol. 30:767–771. 1998.
|
|
72.
|
Szebenyi G and Rotweien P: The mouse
insulin-like growth factor II/cation-independent mannose
6-phosphate (IGF-II/MPR) receptor gene: molecular cloning and
genomic organization. Genomics. 19:120–129. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
73.
|
Frasca F, Pandini G, Scalia P, et al:
Insulin receptor isoform A, a newly recognized, high-affinity
insulin-like growth factor II receptor in fetal and cancer cells.
Mol Cell Biol. 19:3278–3288. 1999.PubMed/NCBI
|
|
74.
|
Stewart CEH and Rotwein P: Growth,
differentiation, and survival: multiple physiological functions for
insulin-like growth factors. Physiol Rev. 76:1005–1026.
1996.PubMed/NCBI
|
|
75.
|
Baserga R: The IGF-1 receptor in cancer
biology. Int J Cancer. 107:873–877. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
76.
|
Grimberg A: Mechanisms by which IGF-I may
promote cancer. Cancer Biol Ther. 2:630–635. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
77.
|
Delafontaine P, Song YH and Li Y:
Expression, regulation, and function of IGF-1, IGF-1R, and IGF-1
binding proteins in blood vessels. Arterioscler Thromb Vasc Biol.
24:435–444. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
78.
|
Yu H and Rohan T: Role of the insulin-like
growth factor family in cancer development and progression. J Natl
Cancer Inst. 92:1472–1489. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
79.
|
Samani AA, Yakar S, LeRoith D and Brodt P:
The role of the IGF system in cancer growth and metastasis:
overview and recent insights. Endocr Rev. 28:20–47. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
80.
|
Laureys G, Barton DE, Ullrich A and
Francke U: Chromosomal mapping of the gene for the type II
insulin-like growth factor receptor/cation-independent mannose
6-phosphate receptor in man and mouse. Genomics. 3:224–229. 1988.
View Article : Google Scholar : PubMed/NCBI
|
|
81.
|
Morgan DO, Edman JC, Standring DN, Fried
VA, Smith MC, Roth RA and Rutter WJ: Insulin-like growth factor II
receptor as a multifunctional binding protein. Nature. 329:301–307.
1987. View Article : Google Scholar : PubMed/NCBI
|
|
82.
|
Lau MM, Stewart CE, Liu Z, Bhatt H,
Rotwein P and Stewart CL: Loss of the imprinted
IGF2/cation-independent mannose 6-phosphate receptor results in
fetal overgrowth and perinatal lethality. Genes Dev. 8:2953–2963.
1994. View Article : Google Scholar
|
|
83.
|
Sun H, Chua M-S, Yang D, Tsalenko A, Peter
B and So S: Antibody arrays identify potential diagnostic markers
of hepatocellular carcinoma. Biomark Insights. 3:1–18.
2008.PubMed/NCBI
|
|
84.
|
Tovar V, Alsinet C, Villanueva A, et al:
IGF activation in a molecular subclass of hepatocellular carcinoma
and pre-clinical efficacy of IGF-1R blockage. J Hepatol.
52:550–559. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
85.
|
Weng CJ, Hsieh YH, Tsai CM, et al:
Relationship of insulin-like growth factors system gene
polymorphisms with the susceptibility and pathological development
of hepatocellular carcinoma. Ann Surg Oncol. 17:1808–1815. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
86.
|
Scharf JG, Schmidt-Sandte W, Pahernik SA,
Ramadori G, Braulke T and Hartmann H: Characterization of the
insulin-like growth factor axis in a human hepatoma cell line
(PLC). Carcinogenesis. 19:2121–2128. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
87.
|
Tsai TF, Yauk YK, Chou CK, et al: Evidence
of autocrine regulation in human hepatoma cell lines. Biochem
Biophys Res Commun. 153:39–45. 1988. View Article : Google Scholar : PubMed/NCBI
|
|
88.
|
Tanaka S, Mohr L, Schmidt EV, Sugimachi K
and Wands JR: Biological effects of human insulin receptor
substrate-1 over-expression in hepatocytes. Hepatology. 26:598–604.
1997. View Article : Google Scholar : PubMed/NCBI
|
|
89.
|
Nishiyama M and Wands JR: Cloning and
increased expression of an insulin receptor substrate-1-like gene
in human hepatocellular carcinoma. Biochem Biophys Res Commun.
183:280–285. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
90.
|
Coutant A, Rescan C, Gilot D, Loyer P,
Guguen-Guillouzo C and Baffet G: PI3K- FRAP/mTOR pathway is
critical for hepatocyte proliferation whereas MEK/ERK supports both
proliferation and survival. Hepatology. 36:1079–1088. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
91.
|
Alexia C, Fallot G, Lasfer M,
Schweizer-Groyer G and Groyer A: An evaluation of the role of
insulin-like growth factors (IGF) and of type-I IGF receptor
signaling in hepatocarcinogenesis and in the resistance of
hepatocellular cells against drug-induced apoptosis. Biochem
Pharmacol. 86:1003–1015. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
92.
|
LeRoith D: Insulin-like growth factor I
receptor signaling-overlapping or redundant pathways.
Endocrinology. 141:1287–1288. 2000.PubMed/NCBI
|
|
93.
|
Knittel T, Dinter C, Kobold D, Neubauer K,
Mehde M, Eichhorst S and Ramadori G: Expression and regulation of
cell adhesion molecules by hepatic stellate cells (HSC) of rat
liver: involvement of HSC in recruitment of inflammatory cells
during hepatic tissue repair. Am J Pathol. 154:153–167. 1999.
View Article : Google Scholar
|
|
94.
|
Wu JC, Daughaday WH, Lee SD, et al:
Radioimmunoassay of serum IGF-I and IGF-II in patients with chronic
liver diseases and hepatocellular carcinoma with or without
hypoglycemia. J Lab Clin Med. 112:589–594. 1988.PubMed/NCBI
|
|
95.
|
Kratzsch J, Blum WF, Schenker E and Keller
E: Regulation of growth hormone (GH), insulin-like growth factor
(IGF) I, IGF binding proteins -1, -2, -3 and GH binding protein
during progression of liver cirrhosis. Exp Clin Endocrinol
Diabetes. 103:285–291. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
96.
|
Nikolić JA, Todorović V, Bozić M, et al:
Serum insulin-like growth factor (IGF)-II is more closely
associated with liver disfunction than is IGF-I in patients with
cirrhosis. Clin Chim Acta. 294:169–177. 2000.PubMed/NCBI
|
|
97.
|
Okan A, Comlekci A, Akpinar H, Okan I,
Yesil S, Tankurt E and Simsek I: Serum concentration of
insulin-like growth factor-I and insulin-like growth factor binding
protein-3 in patients with chronic hepatitis. Scand J
Gastroenterol. 35:1212–1215. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
98.
|
Stuver SO, Kuper H, Tzonou A, Lagiou P,
Spanos E and Hsieh CC: Insulin-like growth factor 1 in
hepatocellular carcinoma and metastatic liver cancer in men. Int J
Cancer. 87:118–121. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
99.
|
Vyzantiadis T, Theodoridou S, Giouleme O,
Harsoulis P, Evgenidis N and Vyzantiadis A: Serum concentrations of
insulin-like growth factor-I (IGF-I) in patients with liver
cirrhosis. Hepatogastroenterology. 50:814–816. 2003.PubMed/NCBI
|
|
100.
|
Lorenzo-Zúñiga V, Bartoli R, Masnou H,
Montoliu S, Morillas RM and Planas R: Serum concentration of
insulin-like growth factor-I (IGF-I) as a marker of liver fibrosis
in patients with chronic hepatitis C. Dig Dis Sci. 52:3245–3250.
2007.PubMed/NCBI
|
|
101.
|
Mahdy KA, Ahmed HH, Mannaa F and
Abdel-Shaheed A: Clinical benefits of biochemical markers of bone
turnover in Egyptian children with chronic liver diseases. World J
Gastroenterol. 13:785–790. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
102.
|
Mazziotti G, Sorvillo F, Morisco F, et al:
Serum insulin-growth factor I evaluation as a useful tool for
predicting the risk of developing hepatocellular carcinoma in
patients with hepatitis C virus-related cirrhosis: a prospective
study. Cancer. 95:2539–2545. 2002. View Article : Google Scholar
|
|
103.
|
Wu YL, Ye J, Zhang S, Zhong J and Xi RP:
Clinical significance of serum IGF-I, IGF-II and IGFBP-3 in liver
cirrhosis. World J Gastroenterol. 10:2740–2743. 2004.PubMed/NCBI
|
|
104.
|
Weber MM, Auernhammer CJ, Lee PD,
Engelhardt D and Zachoval R: Insulin-like growth factors and
insulin-like growth factor binding proteins in adult patients with
severe liver disease before and after orthotropic liver
transplantation. Horm Res. 57:105–112. 2002. View Article : Google Scholar
|
|
105.
|
Ranke MB, Maier KP, Schweizer R, Stadler
B, Schleicher S, Elmlinger MW and Flehmig B: Pilot study of
elevated levels of insulin-like growth factor-binding protein-2 as
indicators of hepatocellular carcinoma. Horm Res. 60:174–180. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
106.
|
Morali G, Shitrit AB, Eran M, Freier S,
Reinus C and Braverman D: Hepatic production of insulin-like growth
factors in normal and diseased liver. Hepatogastroenterology.
52:1511–1515. 2005.PubMed/NCBI
|
|
107.
|
Nordstedt G, Levinovitz A, Möller C,
Eriksson LC and Anderson G: Expression of insulin-like growth
factor I (IGF-I) and IGF-II mRNA during hepatic development,
proliferation and carcinogenesis in the rat. Carcinogenesis.
9:209–213. 1988. View Article : Google Scholar : PubMed/NCBI
|
|
108.
|
Couvert P, Carrie A, Paries J, et al:
Liver insulin-like growth factor 2 methylation in hepatitis C virus
cirrhosis and further occurrence of hepatocellular carcinoma. World
J Gastroenterol. 14:5419–5427. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
109.
|
Price JA, Kovach SJ, Johnson T, Koniaris
LG, Cahill PA, Sitzmann JV and McKillop IH: Insulin-like growth
factor I is a comitogen for hepatocytes growth factor in a rat
model of hepatocellular carcinoma. Hepatology. 36:1089–1097. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
110.
|
Kim SO, Park JG and Lee YI: Increased
expression of the insulin-like growth factor I (IGF-I) receptor
gene in hepatocellular carcinoma cell lines: implications of the
IGF-I receptor gene activation by hepatitis B virus X gene product.
Cancer Res. 56:3831–3836. 1996.PubMed/NCBI
|
|
111.
|
Nardone G, Romano M, Calabro A, et al:
Activation of fetal promoters of insulin-like growth factors II
gene in hepatitis C virus-related chronic hepatitis, cirrhosis, and
hepatocellular carcinoma. Hepatology. 23:1304–1312. 1996.
View Article : Google Scholar : PubMed/NCBI
|
|
112.
|
Sohda T, Kamimura S, Iwata K, Shijo H and
Okumura M: Immunohistochemical evidence of insulin-like growth
factor II in human small hepatocellular carcinoma with hepatitis C
virus infection: relationship to fatty change in carcinoma cells. J
Gastroenterol Hepatol. 12:224–228. 1997. View Article : Google Scholar
|
|
113.
|
Chin E, Zhou J, Dai J, Baxter RC and Bondy
CA: Cellular localization and regulation of gene expression for
components of the insulin-like growth factor ternary binding
protein complex. Endocrinology. 134:2498–2504. 1994.PubMed/NCBI
|
|
114.
|
Arany E, Afford S, Strain AJ, Winwood PJ,
Arthur MJ and Hill DJ: Differential cellular synthesis of
insulin-like growth factor binding protein-1 (IGFBP-1) and IGFBP-3
within human liver. J Clin Endocrinol Metabol. 79:1871–1876.
1994.PubMed/NCBI
|
|
115.
|
Ross RJM, Chew SL, D’Souza Li L, et al:
Expression of IGF-I and IGF-binding protein genes in cirrhotic
liver. J Endocrinol. 149:209–216. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
116.
|
Völzke H, Nauck M, Rettig R, Dörr M,
Higham C, Brabant G and Wallaschofski H: Association between
hepatic steatosis and serum IGF1 and IGFBP-3 levels in a
population-based sample. Eur J Endocrinol. 161:705–713.
2009.PubMed/NCBI
|
|
117.
|
Colakoğlu O, Taşkiran B, Colakoğlu G,
Kizildağ S, Ari Ozkan F and Unsal B: Serum insulin like growth
factor-1 (IGF-1) and insulin growth factor binding protein-3
(IGFBP-3) levels in liver cirrhosis. Turk J Gastroenterol.
18:245–249. 2007.
|
|
118.
|
Mattera D, Capuano G, Colao A, Pivonello
R, Manguso F, Puzziello A and D’Agostino L: Increased IGF-I :
IGFBP-3 ratio in patients with hepatocellular carcinoma. Clin
Endocrinol. 59:699–706. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
119.
|
Moller S, Juul A, Becker U, Flyvbjerg A,
Skakkebaek NE and Henriksen JH: Concentrations, release, and
disposal of insulin-like growth factor (IGF)-binding proteins
(IGFBP), IGF-I, and growth hormone in different vascular beds in
patients with cirrhosis. J Clin Endocrinol Metab. 80:1148–1157.
1995.PubMed/NCBI
|
|
120.
|
Juul A, Dalgaard P, Blum WF, et al: Serum
levels of insulin-like growth factor (IGF)-binding protein-3
(IGFBP-3) in healthy infants, children, and adolescents: the
relation to IGF-I, IGF-II, IGFBP-1, IGFBP-2, age, sex, body mass
index, and pubertal maturation. J Clin Endocrin Metabol.
80:2534–2542. 1995.PubMed/NCBI
|
|
121.
|
Raslan HM, Ezzat WM, Ahmed MM and Rasheed
EA: Insulin growth factor-1 and insulin growth factor binding
protein-3 in Egyptian patients with chronic hepatitis C. Arch Med
Sci. 3:46–51. 2007.
|
|
122.
|
Shin SY, Lee JR, Noh GW, Kim HJ, Kang WJ,
Kim SH and Chung JK: Analysis of serum levels of anti-Mullerian
hormone, inhibin B, insulin-like growth factor-I, insulin-like
growth factor binding protein-3, and follicle-stimulating hormone
with respect to age and menopausal status. J Korean Med Sci.
23:104–110. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
123.
|
Okabe H, Satoh S, Kato T, et al:
Genome-wide analysis of gene expression in human hepatocellular
carcionamas using cDNA microarray: identification of genes involved
in viral carcinogenesis and tumor progression. Cancer Res.
61:2129–2137. 2001.PubMed/NCBI
|
|
124.
|
Su TS, Liu WY, Han SH, Jansen M, Yang-Fen
TL, P’eng FK and Chou CK: Transcripts of the insulin-like growth
factors I and II in human hepatoma. Cancer Res. 49:1773–1777.
1989.PubMed/NCBI
|
|
125.
|
Ng IO, Lee J MF, Srivastava G and Ng M:
Expression of insulin-like growth factor II mRNA in hepatocellular
carcinoma. J Gastroenterol Hepatol. 13:152–157. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
126.
|
Cariani E, Lasserre C, Seurin D, et al:
Differential expression of insulin-like growth factor II mRNA in
human primary liver cancers, benign liver tumors, and liver
cirrhosis. Cancer Res. 48:6844–6849. 1988.PubMed/NCBI
|
|
127.
|
Lamas E, Le Bail B, Housset C, Boucher O
and Brechot C: Localization of insulin-like growth factor-II and
hepatitis B virus mRNAs and proteins in human hepatocellular
carcinomas. Lab Invest. 64:98–104. 1991.PubMed/NCBI
|
|
128.
|
Lamas E, Zindy F, Seurin D,
Guguen-Guillouzo and Brechot C: Expression of insulin-like growth
factor II and receptors for insulin-like growth factor II,
insulin-like growth factor I and insulin in isolated and cultured
rat hepatocytes. Hepatology. 13:936–940. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
129.
|
D’Errico A, Grignioni WF, Fiorentino M, et
al: Expression of insulin-like growth factor II (IGF-II) in human
hepatocellular carcinoma: an immunohistochemical study. Pathol Int.
44:131–137. 1994.
|
|
130.
|
Iizuka N, Oka M, Tamesa T, Hamamoto Y and
Yamada-Okabe H: Imbalance in expression levels of insulin-like
growth factor 2 and H19 transcripts linked to progression of
hepatocellular carcinoma. Anticancer Res. 24:4085–4089.
2004.PubMed/NCBI
|
|
131.
|
Sedlaczek N, Hasilik A, Neuhaus P,
Schuppan D and Herbst H: Focal overexpression of insulin-like
growth factor 2 by hepatocytes and cholangiocytes in viral liver
cirrhosis. Br J Cancer. 88:733–739. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
132.
|
Caro JF, Poulos J, Ittoop O, Pories WJ,
Flickinger EG and Sinha MK: Insulin-like growth factor binding in
hepatocytes from human liver, human hepatoma, and normal,
regenerating, and fetal rat liver. J Clin Invest. 81:976–981. 1988.
View Article : Google Scholar : PubMed/NCBI
|
|
133.
|
Verspohl EJ, Maddux BA and Goldfine ID:
Insulin and insulin-like growth factor I regulate the same
biological functions in HEP-G2 cells via their own specific
receptors. J Clin Endocrinol Metab. 67:169–174. 1988. View Article : Google Scholar : PubMed/NCBI
|
|
134.
|
Zhang YC, Wang XP, Zhang LY, Song AL, Kou
ZM and Li XS: Effect of blocking IGF-I receptor on growth of human
hepatocellular carcinoma cells. World J Gastroenterol.
12:3977–3982. 2006.PubMed/NCBI
|
|
135.
|
Lee YI, Han YJ, Lee SY, et al: Activation
of insulin-like growth factor II signaling by mutant type p53.
Physiological implications for potentiation of IGF-II signaling by
p53 mutant 249. Mol Cell Endocrinol. 203:51–63. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
136.
|
Lee YI, Lee S, Das GC, Park US and Park
SM: Activation of the insulin-like growth factor II transcription
by aflatoxin B1 induced p53 mutant 249 is caused by activation of
transcription complexes; implications for a gain-of-function during
formation of hepatocellular carcinoma. Oncogene. 19:3717–3726.
2000. View Article : Google Scholar
|
|
137.
|
De Souza AT, Hankins GR, Washington MK,
Orton TC and Jirtle RL: M6P/IGF2R gene is mutated in human
hepatocellular carcinomas with loss of heterozygosity. Nat Genet.
11:447–449. 1995.PubMed/NCBI
|
|
138.
|
Su WW, Lee KT, Yeh YT, Soon MS, Wang CL,
Yu ML and Wang SN: Association of circulating insulin-like growth
factor 1 with hepatocellular carcinoma: one cross-sectional
correlation study. J Clin Lab Anal. 24:195–200. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
139.
|
Plöckinger U, Krüger D, Bergk A,
Wiedenmann B and Berg T: Hepatitis-C patients have reduced growth
hormone (GH) secretion which improves during long-term therapy with
pegylated interferon-alpha. Am J Gastroenterol. 102:2724–2731.
2007.PubMed/NCBI
|
|
140.
|
Helaly GF, Hussein NG, Refai W and Ibrahim
M: Relation of serum insulin-like growth factor-1 (IGF-1) levels
with hepatitis C virus infection and insulin resistance. Transl
Res. 158:155–162. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
141.
|
Adamek A, Kasprzak A, Seraszek A, Mikos H,
Bura A and Mozer-Lisewska I: Alterations of serum levels of
insulin-like growth factor I (IGF-I) and estradiol in chronic
hepatitis C. Onkol Wspol. 16:234–239. 2012.PubMed/NCBI
|
|
142.
|
Conchillo M, Prieto J and Quiroga J:
Insulin like growth factor I (IGF-I) and liver cirrhosis. Rev Esp
Enferm Dig. 99:156–164. 2007.(In Spanish).
|
|
143.
|
Hung CH, Wang JH, Hu TH, et al: Insulin
resistance is associated with hepatocellular carcinoma in chronic
hepatitis C infection. World J Gastroenterol. 16:2265–2271. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
144.
|
Stefano JT, Correa-Giannella ML, Ribeiro
CMF, Alves VAF, Massarollo PCB, Machado MCC and Giannella-Neto D:
Increased hepatic expression of insulin-like growth factor-I
receptor in chronic hepatitis C. World J Gastroenterol.
28:3821–3828. 2006.PubMed/NCBI
|
|
145.
|
Tanaka S, Takenaka K, Matsumata T, Mori R
and Sugimachi K: Hepatitis C virus replication is associated with
expression of transforming growth factor-alpha and insulin-like
growth factor-II in cirrhotic livers. Dig Dis Sci. 41:208–215.
1996. View Article : Google Scholar : PubMed/NCBI
|
|
146.
|
Sohda T, Oka Y, Iwata K, et al:
Co-localisation of insulin-like growth factor II and the
proliferation marker MIB1 in hepatocellular carcinoma cells. J Clin
Pathol. 50:135–137. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
147.
|
Yakar S, LeRoith D and Brodt P: The role
of the growth hormone/insulin-like growth factor axis in tumor
growth and progression: lessons from animal models. Cytokine Growth
Factor Rev. 16:407–420. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
148.
|
Wu Y, Brodt P, Sun H, et al: Insulin-like
growth factor-I regulates the liver microenvironment in obese mice
and promotes liver metastasis. Cancer Res. 70:57–67. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
149.
|
Lee S, Park U and Lee YI: Hepatitis C
virus core protein transactivates insulin-like growth factor II
gene transcription through acting concurrently on Egr1 and Sp1
sites. Virology. 283:167–177. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
150.
|
Kasprzak A, Adamek A, Przybyszewska W, et
al: Expression of IGF-I and viral proteins (C, NS3, NS5A) in the
livers of patients with chronic HCV infection. Adv Clin Exp Med.
20:263–273. 2011.
|