Introduction
Mesenchymal stem cells (MSC) are a rare population
of bone marrow-derived non-hematopoietic fibroblast-like cells,
representing ∼0.001–0.01% of the nucleated cells in the marrow. The
accessibility, ease of handling, and enormous expansion potential
of MSC, together with their capacity for self-renewal (reviewed in
ref. 1) and amenability to
allogeneic transplantation (2),
provide a nearly unlimited supply of cells for therapeutic
applications. MSC are attractive candidates for manipulation as
they can easily be isolated from patients, cultured in
vitro, and autologously transplanted into patients, thus
overcoming the difficulties related to immune rejection of
transplanted cells (3). A
significant improvement in understanding MSC biology in recent
years has paved the way to their potential clinical use. In
particular, MSC are emerging as novel cell-based delivery agents.
Indeed, thanks to inherent tumor-trophic migratory properties, MSC
could vehicle effective targeted therapy to isolated tumors and
metastatic disease. However, defective migration is a major caveat
in cell-based therapy as systemic stem cell transplantation
requires efficient cell homing, and is therefore of utmost
importance to understand the underlying molecular mechanisms
driving MSC movement to specific target sites (reviewed in ref.
4).
Phosphoinositide-3 kinase (PI3K)/AKT signaling
circuit is a well established regulator of important functions in
cells, such as cell cycling, survival and cell growth (5). In addition, recent studies have
demonstrated that AKT plays a pivotal role in cell migration and
invasion (6–8). In particular, AKT enhances actin
remodeling by a number of possibly interconnected mechanisms, in
several cell systems. AKT modulates actin bundling by direct actin
binding, and generates membrane protrusions through downstream
activation of Rac1 and Cdc42 (9,10).
Besides, PI3K/AKT signals induce also the remodeling of actin
through the activation of p70S6K, leading to cell migration and
cell invasion (11). Furthermore,
AKT promotes actin organization and cell motility mediated by a
number of substrates and/or interactors, such as
Girdin/AKT-phosphorylation enhancer (APE) (12), the actin bundling protein Palladin
(13) and the mechano-protein and
AKT-substrate ANKRD2 (14).
Accordingly, PI3K/AKT signals are considered to be crucial in cell
migration by controlling actin dynamics.
Remarkably, it has been demonstrated that AKT
isoform 1 and AKT isoform 2 exert opposing functions in breast
cancer cell migration and metastasis (6,7,13,15).
There is little information, however, on their role in other cell
models. We therefore aimed to elucidate the contribution of AKT to
MSC migration by specific pharmacological inhibition.
Materials and methods
Mesenchymal stem cell culture
Bone marrow (BM) samples were obtained from patients
undergoing surgery at Rizzoli Orthopaedic Institute after obtaining
informed consent. Bone marrow derived MSC cultures were obtained as
previously described (16). Cells
were transferred to 150-cm2 culture flasks with
α-Modified minimum Essential medium (α-MEM; BioWhittaker, Lonza,
Verviers, Belgium) supplemented with 20% lot-selected fetal bovine
serum (FBS; Gibco, Invitrogen, Paisley, UK) and GlutaMAX™ 1%
(Invitrogen), incubated in a humidified atmosphere at 37°C with 5%
CO2 and medium changed every 3–4 days. When the cells
reached approximately 70–80% confluence, they were detached by mild
trypsinization (TripLe™ Select, Invitrogen) for 5 min at 37°C and
counted. Then, 1/3 of them were reseeded into a new 150
cm2 flask.
MSC were recognized by their ability to proliferate
in culture with adherent, spindle-shape morphology. The total
number of cells obtained at each passage was extrapolated from the
counted representative samples; the number was calculated by
multiplying the number of cells counted by the number of flasks
available at each passage. Cell number and cell viability were
assessed for each passage by NucleoCounter® (ChemoMetec,
Lillerød, Denmark), that detects non-viable cells by using
propidium iodide nuclear staining, and determines cell viability by
calculating the ratio of total and non-viable cell number. Further
characterization of MSC was performed by cytofluorimetric analysis
of cell surface markers at passage two. MSC resulted positive for
CD44, CD73, CD90, CD105, CD146 and negative for CD34 and CD45
(17). The multilineage potential
was evaluated by differentiation assays as previously described
(18). After the appropriate
stimulations, cells positively differentiated in osteogenic,
adipogenic and condrogenic lineages.
Drug treatments
Cells were seeded 1×104
cells/cm2 density in complete medium and starved
overnight with 0.2% FBS medium. On the third day, culture was
treated for 30 min with PI3K Inhibitor LY-294002 (#L9908,
Sigma-Aldrich, St. Louis, MO, USA) at 10 μM and AKT
Inhibitor IV (#B2311, Sigma-Aldrich) at 2.5 μM
concentration. Inhibitor VIII (#202048, Santa Cruz Biotechnology,
Santa Cruz, CA, USA) was used at 1 μM to inhibit AKT1/2 and
Inhibitor XII (#124029, Calbiochem, La Jolla, CA, USA) was used at
5 μM for AKT2 isoform. Specific Inhibitors for singular
isoforms were used for 1 h.
Immunofluorescence microscopy
Cells were seeded (2.5×104) onto glass
coverslips and fixed for 10 min in 4% formaldehyde at RT, as
described (19). After three
washes in PBS, cells were permeabilized for 10 min in PBT (PBS +
0,1% Triton X-100), then incubated in blocking solution (1X PBS +
5% BSA) for 30 min. Primary antibody anti-Vinculin (1:100,
#MAB3574, Chemicon, Temecula, CA, USA) diluted in blocking solution
was added and incubated for 1 h at RT. After three washes in 1X
PBS, secondary antibody anti-mouse Cy3 (1:100, #C2181,
Sigma-Aldrich) was added diluted in 1X PBS and incubated for 45 min
at RT. Cortical actin was stained using FITC-Phalloidin (1:500,
#P5282, Sigma-Aldrich) and added directly during secondary antibody
incubation. Nuclei were stained by incubating cells with Hoechst
33342 (1:2000, #H1399, Invitrogen) for 10 min at RT (20). Coverslips were mounted after washes
in PBS in Fluoromount-G (Southern Biotech, Birmingham, AL, USA)
solution and analyzed with a NikonTiE microscope equipped with the
fully automated A1 confocal laser that incorporates the resonant
scanner with a resonance frequency of 7.8 kHz that allows
high-speed imaging (A1R), and equipped with DS-QiMc-U2 12 bit
camera. Digital images were processed using Matlab and ImageJ
software for the analysis and Photoshop software for visualization
purposes without biased manipulations (21).
Preparation of cell extracts and
electrophoresis
Cell extracts were obtained as previously reported
(22). Briefly, sub-confluent
cells were directly extracted by addition of RIPA buffer (20 mM
Tris-Cl, pH 7.0, 1% NP-40, 150 mM NaCl, 10% glycerol, 10 mM EDTA,
20 mM NaF, 5 mM sodium pyrophosphate, 1 mM
Na3VO4) and freshly added Sigma-Aldrich
Protease Inhibitor Cocktail at 4°C for 20 min. Lysates were cleared
by centrifugation. Equal amounts of lysates were loaded on 7,5%
Bis-Tris-SDS-polyacrylamide gels (Bio-Rad, Hercules, CA, USA),
transferred to Immobilon-P membranes (Millipore Corporation,
Billerica, MA, USA), probed with the indicated antibodies by using
SNAP i.d.® 2.0 System (Millipore Corporation) and
detected by chemiluminescence with Supersignal kit (Pierce,
Rockford, IL, USA).
The primary antibodies used: anti-actin 1:1,000
(#sc-8432), anti-AKT 1:1,000 (#sc-1619), from Santa Cruz
Biotechnology and anti-pS473AKT 1:1,000 (#4051) from Cell
Signalling Technology (Denvers, MA, USA). Secondary antibodies were
anti-mouse (#sc-2031) and anti-rabbit (#2317) HRP-conjugated from
Santa Cruz Biotechnology and were used 1:5,000 dilution.
Scratch wound healing assay
Scratch wound tests were performed in a 6-well plate
in confluent culture. Cell suspensions (1,000 μl) containing
3×105 cells in 0.2% FBS medium were added to each well
and left overnight to allow cell adhesion. Immediately before
starting the assay, the scratch was performed by using 200
μl tips. Closing of the wound was monitored along 24 h with
a Nikon Eclipse TE2000-U inverted microscope, and calculated as
Migrated Cell Surface Area/Total Surface Area × 100 with a
web-based image automated analysis software developed by the
Koumoutsakos group (CSE Lab), at ETH Zürich (23).
Cytotoxicity and cell proliferation
assay
Cytotoxicity and cell proliferation were evaluated
using AlamarBlue® kit (AbD Serotec, Oxford, UK)
according to the manufacturer’s instructions. Briefly, the test was
performed in 96-well plate by seeding
3×103/cm2 in 0.2% FBS medium and treated or
not with inhibitors. The reagent was added to the sample for 4 h
incubation, at the end of which the indicator passes from the Redox
oxidized form (blue) to the reduced form (red) according to the
number of cells that are found in the sample. Absorbance was
monitored at 570 nm and 600 nm.
Cell cycle and apoptosis analysis
For detection and quantification of cell cycle
distribution samples containing 5×105 cells were
harvested by centrifugation, fixed in cold ethanol and subjected to
propidium iodide (PI) staining as previously described (24).
Migration assay
Migration assays were performed in transwell inserts
with 8-μm pore uncoated membrane filters (Corning
Incorporated, Corning, NY, USA). Cells were trypsinized and seeded
onto the upper chambers, in 0.2% FBS medium (4×104
cells/well in 100 μl). The bottom chambers were filled with
10% FBS medium (600 μl). Cells were allowed to migrate for 6
h at 37°C. Then, the upper side of the filters was carefully washed
with cold PBS and non-migrated cells at the top of the filter were
removed using a cotton swab. Cells that had migrated to the bottom
of the filters were fixed and stained with Hema 3®
Protocol Stain (Fisher Diagnostics, Middletown, CT, USA), and
imaged with a Nikon Eclipse TE2000-U inverted epifluorescence
microscope. Stained cells were counted in 10 pictures taken at x400
magnification. The average of cell number counted per field was
multiplied by the ratio between the surfaces (total/counted) to
determine the total number of migrated cells. Inhibition of
migration was calculated normalizing data to control cells.
In vivo imaging and motility
analysis
Cells were plated at a density of 2.5×104
cells in 35-mm Glass Bottom Dishes (MatTek Corporation, Ashland,
OR, USA) and let to adhere overnight. Drug treatments were
performed as described above. Images were recorded with a NikonTi
Eclipse microscope equipped with a CO2 temperature
controller atmosphere (Okolab, Ottaviano, NA, Italy) and a
DS-QiMc-U2 12 bit camera. A 10X Plan-Apochromatic Ph1 DL 0.25NA
objective was used, yielding 0.91 μm/pixel in the acquired
images. Total duration of recording is 15 h with a time frame
interval of 5 min. Three plates were acquired for each of the
control (CTRL) and treated (XII) MSC cultures for a total of six
sequences of 180 frames each. Among these, three representative
Regions of Interest (ROI) (labeled a,b,c) for each plate were
considered for the motility analysis. In order to assess cells
motility in treated and control MSC, we employed our tracking
algorithm designed for this purpose. Image processing methods were
devised to first detect the cell regions around the nuclei on each
frame of the considered sequences through non-linear and
morphological filters. Then, a combined feature-based and
morphology-based tracking method was employed to track each cell
center of mass (CM) along the sequence of frames to achieve the
cell trajectory. Since the image illumination may change during the
time-lapse sequence, phase contrast images could present artifacts
that mislead the tracking stage. Also, during the acquisition of
the six sequences, the repositioning error of the motorized stage
where the plates were placed yielded x/y displacement on the
imaging plane, this causing images not to be perfectly aligned.
While the latter problem was easily fixed by phase-correlation
registration methods, the former needed manual intervention.
Trajectories recovered from the sequence of images were therefore
analyzed by two experts to select only those with a high
confidence, while the others were discarded.
Statistical analysis
All the images shown in this paper are
representative of at least three independent experiments carried
out under the same conditions. Statistical analysis were performed
by ANOVA test followed by Bonferroni’s multiple comparison test.
P<0.05 were considered statistically significant. Data are
expressed as the mean ± standard deviation (SD).
Results
PI3K/AKT signaling pathway inhibition
triggers actin remodeling of MSC
MSC were exposed for 30 or 60 min to increasing
concentrations of well characterized PI3K/AKT inhibitors (25), in order to identify concentrations
that do not compromise cell viability (data not shown), and the
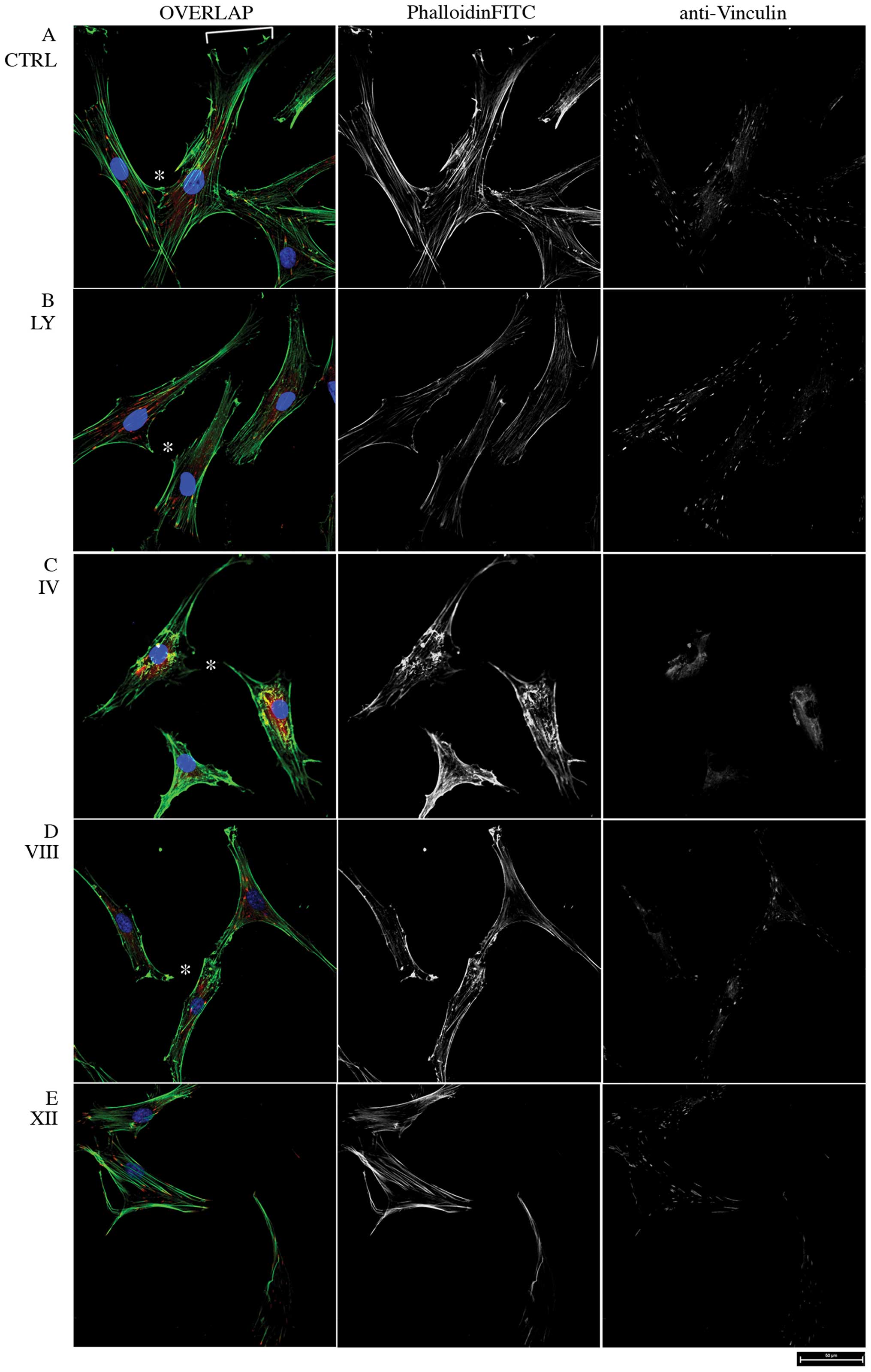
morphology was observed by confocal microscopy (Fig. 1). In control MSC, PhalloidinFITC
staining showed a dense network of F-actin bundles with tight,
parallel stress fibers and adhesion foci and a well shaped
migration front leading edge (Fig.
1A). Conversely, MSC treated with the PI3K Inhibitor LY 294002
were characterized by reduced actin bundling and partial loss of
cell-cell contacts and cell migration front (Fig. 1B). Moreover, the broad PI3K/Akt
Inhibitor IV leds to collapse of actin bundles and complete loss of
intercellular and focal adhesions, as visualized by vinculin
staining (Fig. 1C). The
AKT-dependent actin skeleton remodeling was monitored further in
response to PH domain-dependent, non-ATP-competitive and
isoform-specific Inhibitors VIII and XII (AKTi VIII and XII), that
potently inhibit Akt1/2 and Akt2 respectively, but not other AGC
kinases (26), showing that both
AKTi VIII and XII triggered partial loss of actin fibers and cell
rounding, whereas focal adhesions became restricted to discrete
protruding portions of the cell periphery (Fig. 1D and E). Thus, in good agreement
with previous reports in other cell types, alteration of cell
morphology and actin skeleton organization by pharmacological
inhibition of PI3K/AKT indicates a key role of this pathway in the
cytoskeletal remodeling of MSC.
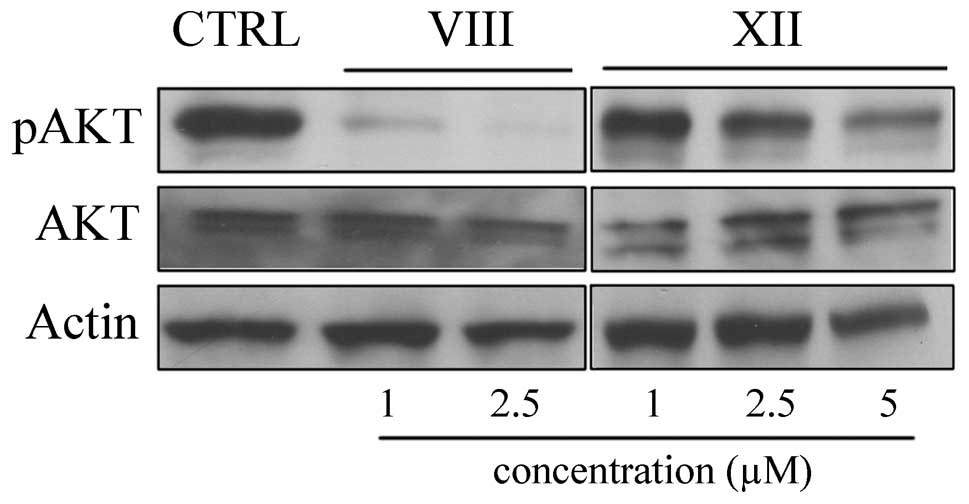
As a read-out of treatment efficacy, phosphorylation
of Akt S473 was monitored by western blot analysis. As expected, 60
min upon treatment AKT S473 phosphorylation was decreased in a
dose-dependent manner by both inhibitors (Fig. 2).
MSC migration and motility are affected
by AKT inhibition
Defective migration is a major limitation in
cell-based therapy. Therefore, in light of its well described role
in actin remodeling linked to cell migration in several cell
models, and based on the above observations, we aimed to
investigate deeper AKT functions in MSC. Since wound repair is a
homeostatic process in which cell migration features prominently,
we first examined the effect of AKT inhibition on the migration of
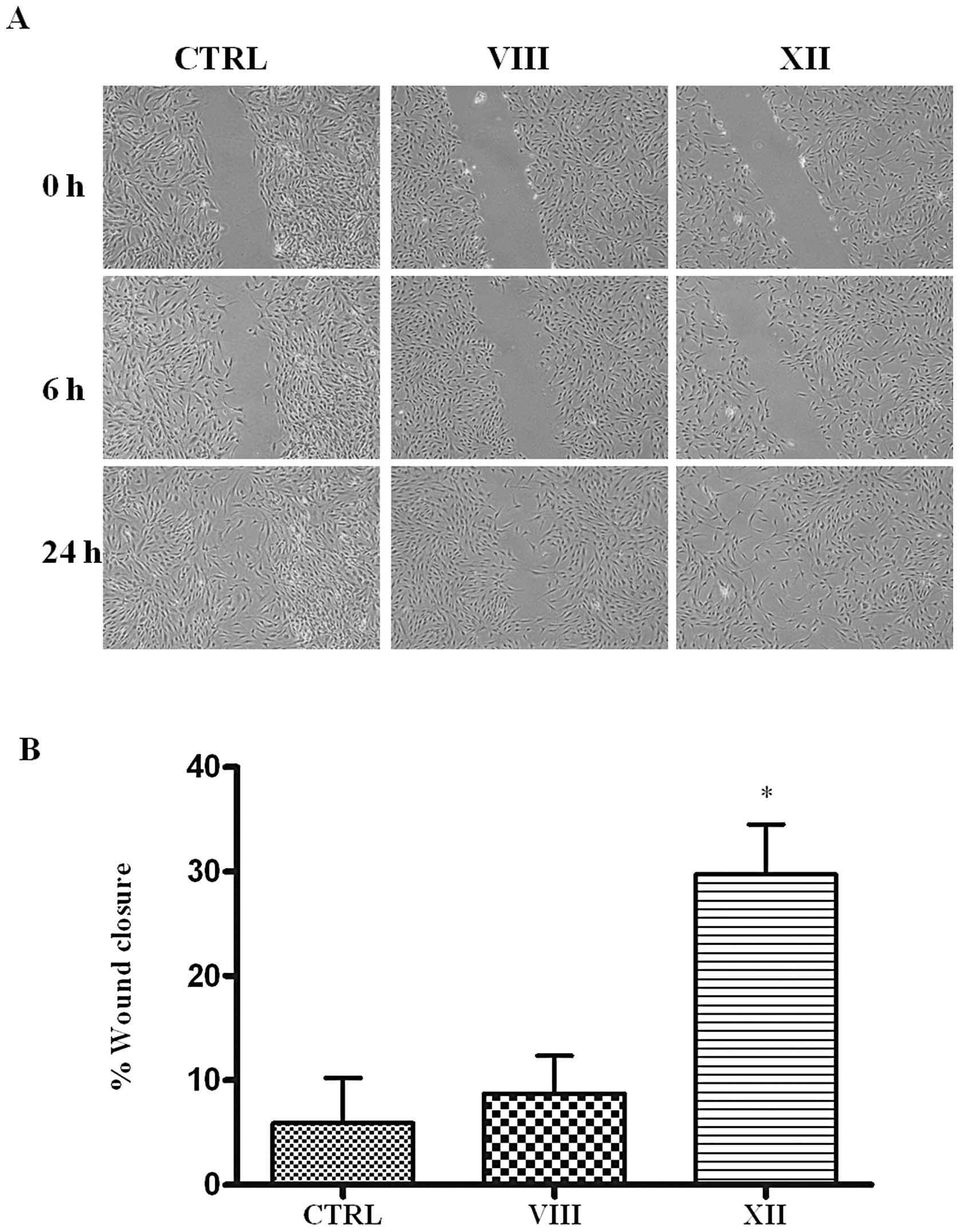
MSC by means of the monolayer scratch wound healing assay.
Remarkably, while vehicle-treated MSC were able to
close the scratch in 24 h, treatment with the AKTi XII, that
selectively inhibits isoform 2, markedly blunted migration and
after 24 h the wound closure was still incomplete (Fig. 3). In contrast, treatment with AKTi
VIII, that inactivates both isoform 1 and 2, reduced migration to a
lesser extent, resulting in an almost complete wound closure after
24 h.
Cell migration is not influenced by cell
proliferation and cytotoxicity
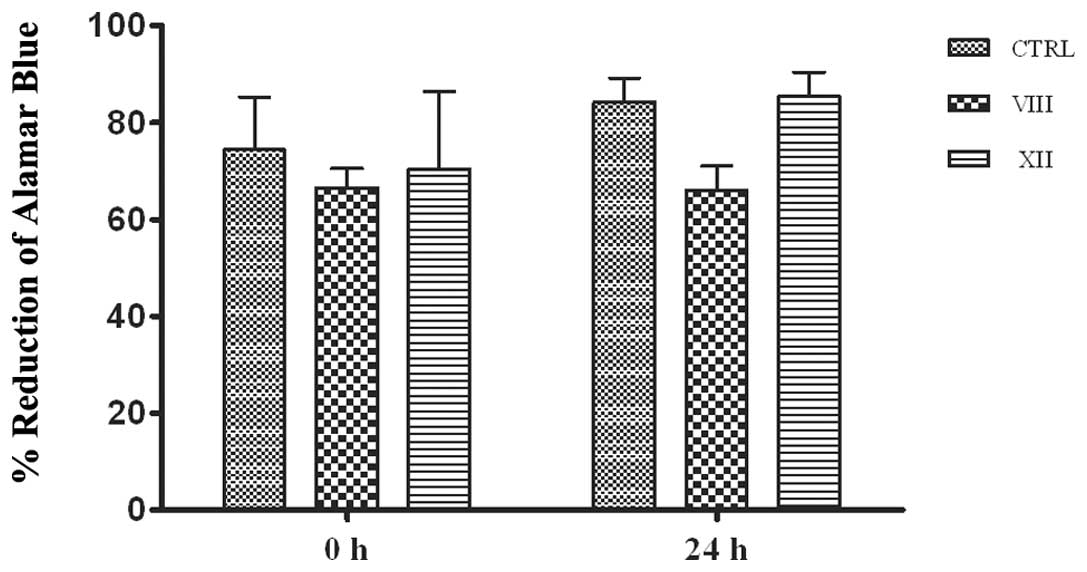
Cell proliferation itself can promote wound closure.
Besides, AKT inhibitors in turn might affect wound closure by
slowing proliferation. To exclude this bias, proliferation of
vehicle- or AKTi-treated MSC was calculated by Alamar Blue assay
(Fig. 4). The test is based on the
ability of metabolically active cells to convert the reagent from a
blue to red colorimetric indicator. Damaged and non-viable cells
have lower innate metabolic activity and thus generate a
proportionally lower signal. Total metabolic activity, checked at
time zero (0 h) and after 24 h, indicates no substantial variation
following AKT inhibition. Thus, the drugs did not alter cell growth
for the duration of the scratch wound test. Furthermore, cell cycle
progression measured by flow cytometry confirmed cell viability and
showed that cell cycle distribution was comparable between samples
(Table I). Moreover, no remarkable
apoptosis was detected, excluding cytotoxic effects of the
above-mentioned drugs in this context. We conclude that the effects
observed on MSC motility were independent of changes in MSC
proliferation as well as alteration of metabolic activity.
 | Table I.FACS analysis of cell cycle and
apoptosis.a |
Table I.
FACS analysis of cell cycle and
apoptosis.a
| CTRL (%) | VIII (%) | XII (%) |
|---|
| G0/G1 phase | | | |
| 0 h | 90 | 94 | 94 |
| 24 h | 93 | 90 | 87 |
| S phase | | | |
| 0 h | 0.7 | 0.8 | 0.7 |
| 24 h | 0.1 | 0.9 | 0.7 |
| G2/M phase | | | |
| 0 h | 1.3 | 2.3 | 2.5 |
| 24 h | 3.6 | 1.4 | 2.1 |
| h-apoptosis | | | |
| 0 h | 6.5 | 2.1 | 1.6 |
| 24 h | 1.4 | 6.6 | 7.3 |
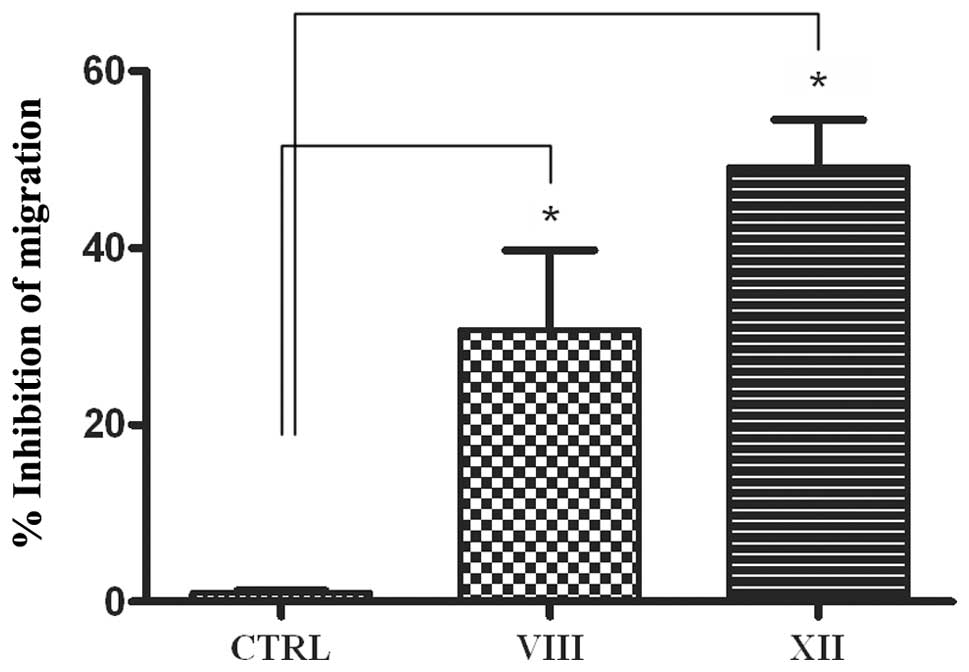
MSC migration is compromised by AKT2
isoform inhibition
The effect of AKT inhibitors on MSC chemotactic
mobility was assessed using 3D transwell migration assays.
Consistent with previous reports (27), MSC control cells migrate toward the
bottom chamber. Remarkably, treatment with the AKTi XII compound
inhibited MSC migration, indicative of a promigratory function of
AKT2 in human MSC (Fig. 5).
Conversely, combined inhibition of AKT1 and AKT2 by Inhibitor VIII
leads to a much weaker effect with respect to inactivation of AKT2
alone. Based on these results, we next tested AKTi XII on 2D-MSC
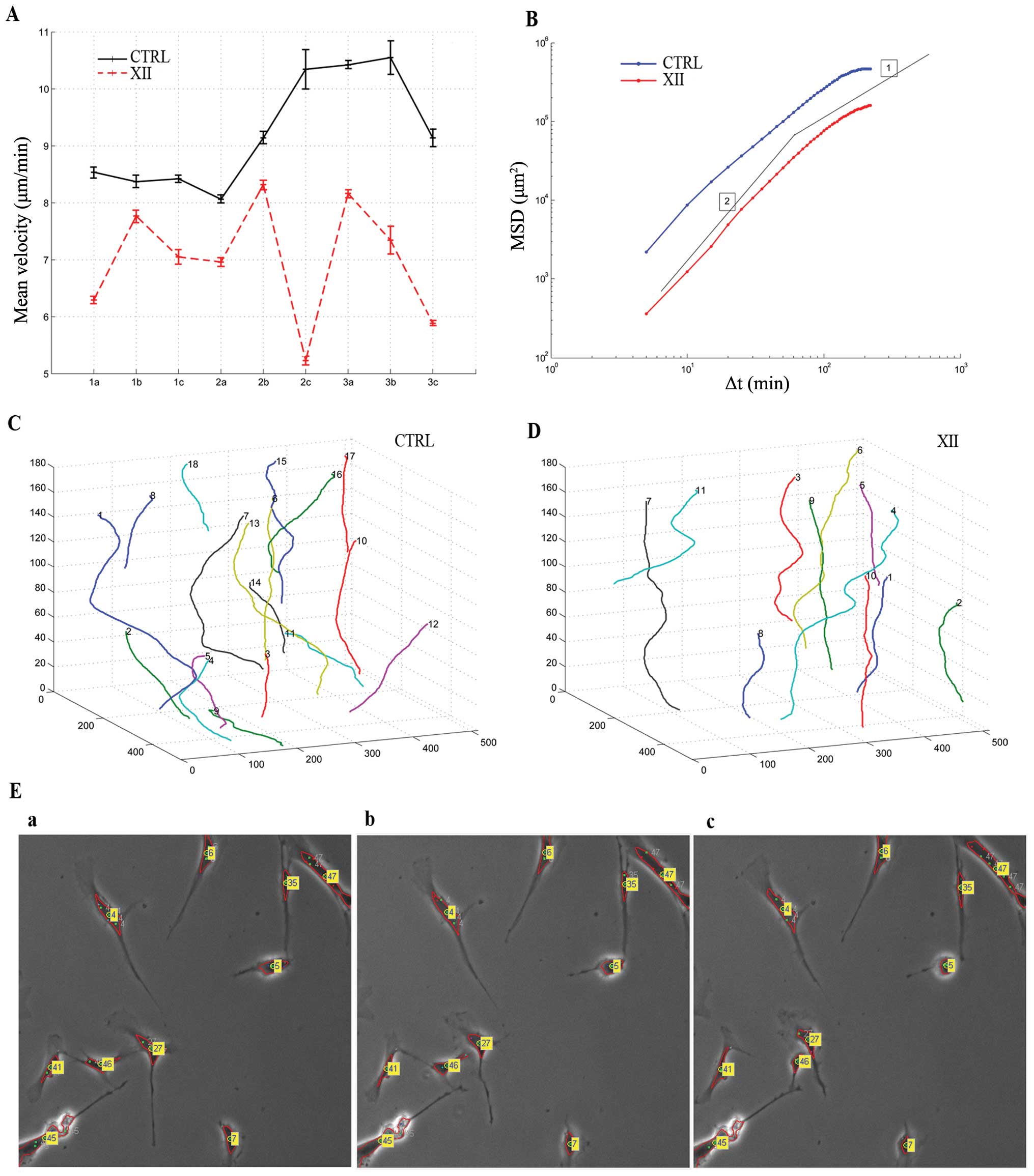
motility through time-lapse in vivo microscopy. Fig. 6A shows the plot of measured mean
velocity (μm/min) along the cell trajectories for the 18
analyzed sequences. Control cells show a higher velocity than
treated cells. The measured mean velocity among the whole sequences
is 9.22±0.98 μm/min for the control and 6.99±1.04
μm/min for treated cells. Cells treated with AKTi XII
exhibit a 24% reduction of mean velocity.
We calculated also the Mean Square Displacement
(MSD) <r2(Δt)> as a function of time lag Δt
(28). In Fig. 6B the logarithmic plot of MSD is
shown. Here, both control and treated cells exhibit super-diffusive
motion for small Δt (see for comparison the logarithmic slope of 2,
which represents a quasi ballistic motion, and slope of 1, which
corresponds to random walks in Brownian motion) and conversely,
approach sub-diffusive motion for large time lags. Besides, for
each time interval considered, untreated cells migrated further
than cells treated with AKTi XII. Overall, the movement and
directionality analyses show speed rate decrease and compromised
status of the treated cell culture. An example of trajectories
recovered from the sequence CTRL 3c and XII 3c is shown in Fig. 6C and D, respectively. These
trajectories depict in an explicatory manner the role of inhibitor
XII in cell motility. Control cell trajectories showed higher
displacements than those revealed on the treated MSC. Fig. 6E shows three subsequent frames
(acquired at time lapse Δt = 5 min) extracted from the same
sequence showing tracked and labeled cells. While the movement of
some cells is reduced (cell labels appear almost in the same
position over this limited period of time), cells labeled 46 and 47
show a more dynamic interaction in this time frame. In general,
considering the whole sequence, untreated cells tend to move faster
than those treated with inhibitor XII.
Discussion
MSC are released from the bone marrow and enter into
the peripheral circulation where, in homeostatic conditions, they
contribute to tissue repair. Moreover, inflammatory chemokines,
cytokines and growth factors released upon damage provide migratory
cues that drive their mobilization to tissue injury sites to
participate in immune modulation, tissue remodeling and wound
healing (29). Taking advantage of
their homing capacities, MSC targeted migration has been used as
delivery vehicle for treatments against cancer, graft versus host
disease, arthritis, multiple sclerosis, and many other diseases.
MSC migration toward the inflammatory signals produced by the
wounded environment has been widely studied, nevertheless the true
migratory mechanism has yet to be elucidated.
PI3K/AKT signaling regulates multiple biological
processes such as cell division, apoptosis, cell growth and cell
migration. A series of recent studies documents the
isoform-specific functions of AKT family members 1 and 2 in
regulation of cellular motility and migration by influencing
numerous cellular targets involved in organization of the actin
cytoskeleton, cellular interaction with the extracellular matrix,
expression of motility genes and establishment of cellular polarity
(30). In particular, AKT1 and
AKT2 opposing functions on migration of breast cancer epithelial
cell lines have been firmly established. AKT2 promotes mammary
epithelial cell migration and invasion, as shown by siRNA-mediated
depletion of AKT2, through upregulation of β1-integrin and
increased stability of palladin expression (6,31).
Conversely, AKT1 negatively affects cell migration by either
transcriptional regulation of motility genes, or actin organization
and formation of stress fibers and cellular interaction with the
extracellular matrix (6,31). The role of AKT isoforms in other
cell models is less clear. In particular, both AKT1 and 2 seem to
play an inhibitory role in prostate cancer cell migration and
invasion (32).
In this study, we examined human MSC migration and
wound closure potential in vitro. We also used selective
inhibitors of AKT isoforms 1 and 2 to dissect the contribution of
each isoform to migration, by selectively blocking either isoform 2
only or both isoforms. Our results showed that both isoforms are
expressed at high level in MSC obtained from three different
individual and cultured in vitro. Reactivity to anti-pSer473
further demonstrates their activity. Of note, only selective
inhibition of AKT2 effectively counteracted wound closure, whereas
dual inhibition of AKT1 and 2 was less effective. An explanation
for this result comes from previous reports, showing that AKT1
inhibition can lead to increased cell migration. Hence, in our cell
system blocking both isoforms weakens the efficacy of AKT2
inhibition. To our knowledge, this is the first study that provides
evidence for an isoform-specific positive role of AKT in regulating
migration of human mesenchymal stem cells.
Although we have a deep understanding of the
mechanisms by which the AKT kinase is regulated, what ultimately
qualifies a kinase are its substrates, as it is very well known
that the phosphorylation of specific substrates determines most of
protein kinase cellular functions (33). Recently, the mechano-sensor and
actin bundling protein Ankrd2 was demonstrated to be a specific
AKT2 substrate (14). It is
therefore tempting to speculate that AKT2 might promote MSC
migration through direct phosphorylation of Ankrd2. However, the
relative contribution, if any, of Ankrd2 to the phenotypic changes
associated with activation of AKT2 needs deeper investigation.
MSC are used with increasing frequency in
laboratories and clinics as potential treatments for injury and
tissue regeneration. The fundamental mechanisms through which MSC
migrate to sites of injury remains undefined. Our finding that AKT2
is required for directed chemotaxis by MSC offers a potential
target of clinical relevance for enhancing or blocking the effects
of migrating MSC.
Acknowledgements
The authors are grateful to Dr
Panagiota Dimopoulou for editorial assistance. Research was
supported in part by Progetto MIUR-FIRB (accordi di programma 2010
RBAP1044), MIUR-FIRB (Human Proteome Net), by Italian
MIUR-Programma di Ricerca di Rilevante Interesse Nazionale Prin
2008 and by Italian Ministry of Health (project IOR-2006-422755).
SM acknowledges the ‘International short term mobility program’
from Italian CNR.
References
|
1.
|
Charbord P: Bone marrow mesenchymal stem
cells: historical overview and concepts. Hum Gene Ther.
21:1045–1056. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
2.
|
Devine SM, Cobbs C, Jennings M,
Bartholomew A and Hoffman R: Mesenchymal stem cells distribute to a
wide range of tissues following systemic infusion into nonhuman
primates. Blood. 101:2999–3001. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
3.
|
Chamberlain G, Fox J, Ashton B and
Middleton J: Concise review: mesenchymal stem cells: their
phenotype, differentiation capacity, immunological features, and
potential for homing. Stem Cells. 25:2739–2749. 2007. View Article : Google Scholar
|
|
4.
|
Kang SK, Shin IS, Ko MS, Jo JY and Ra JC:
Journey of mesenchymal stem cells for homing: strategies to enhance
efficacy and safety of stem cell therapy. Stem Cells Int.
2012:3429682012.PubMed/NCBI
|
|
5.
|
Manning BD and Cantley LC: AKT/PKB
signaling: navigating downstream. Cell. 129:1261–1274. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
6.
|
Chin YR and Toker A: Akt isoform-specific
signaling in breast cancer: uncovering an anti-migratory role for
palladin. Cell Adh Migr. 5:211–214. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
7.
|
Worster DT, Schmelzle T, Solimini NL,
Lightcap ES, Millard B, Mills GB, Brugge JS and Albeck JG: Akt and
ERK control the proliferative response of mammary epithelial cells
to the growth factors IGF-1 and EGF through the cell cycle
inhibitor p57Kip2. Sci Signal. 5:ra192012.PubMed/NCBI
|
|
8.
|
Kakinuma N, Roy BC, Zhu Y, Wang Y and
Kiyama R: Kank regulates RhoA-dependent formation of actin stress
fibers and cell migration via 14-3-3 in PI3K-Akt signaling. J Cell
Biol. 181:537–549. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
9.
|
Cenni V, Sirri A, Riccio M, Lattanzi G,
Santi S, de Pol A, Maraldi NM and Marmiroli S: Targeting of the
Akt/PKB kinase to the actin skeleton. Cell Mol Life Sci.
60:2710–2720. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
10.
|
Scita G, Tenca P, Frittoli E, Tocchetti A,
Innocenti M, Giardina G and Di Fiore PP: Signaling from Ras to Rac
and beyond: not just a matter of GEFs. EMBO J. 19:2393–2398. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
11.
|
Qian Y, Corum L, Meng Q, Blenis J, Zheng
JZ, Shi X, Flynn DC and Jiang BH: PI3K induced actin filament
remodeling through Akt and p70S6K1: implication of essential role
in cell migration. Am J Physiol Cell Physiol. 286:C153–C163. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
12.
|
Enomoto A, Murakami H, Asai N, Morone N,
Watanabe T, Kawai K, Murakumo Y, Usukura J, Kaibuchi K and
Takahashi M: Akt/PKB regulates actin organization and cell motility
via Girdin/APE. Dev Cell. 9:389–402. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
13.
|
Yoeli-Lerner M, Yiu GK, Rabinovitz I,
Erhardt P, Jauliac S and Toker A: Akt blocks breast cancer cell
motility and invasion through the transcription factor NFAT. Mol
Cell. 20:539–550. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
14.
|
Cenni V, Bavelloni A, Beretti F,
Tagliavini F, Manzoli L, Lattanzi G, Maraldi NM, Cocco L and
Marmiroli S: Ankrd2/ARPP is a novel Akt2 specific substrate and
regulates myogenic differentiation upon cellular exposure to
H(2)O(2). Mol Biol Cell. 22:2946–2956. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
15.
|
Irie HY, Pearline RV, Grueneberg D, Hsia
M, Ravichandran P, Kothari N, Natesan S and Brugge JS: Distinct
roles of Akt1 and Akt2 in regulating cell migration and
epithelial-mesenchymal transition. J Cell Biol. 171:1023–1034.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
16.
|
Pierini M, Dozza B, Lucarelli E, Tazzari
PL, Ricci F, Remondini D, di Bella C, Giannini S and Donati D:
Efficient isolation and enrichment of mesenchymal stem cells from
bone marrow. Cytotherapy. 14:686–693. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
17.
|
Maraldi T, Riccio M, Resca E, Pisciotta A,
La Sala GB, Ferrari A, Bruzzesi G, Motta A, Migliaresi C, Marzona L
and De Pol A: Human amniotic fluid stem cells seeded in fibroin
scaffold produce in vivo mineralized matrix. Tissue Eng Part A.
17:2833–2843. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
18.
|
Dozza B, Gobbi G, Lucarelli E, Pierini M,
Bella CD, Frisoni T, Tazzari PL, Ricci F, Mirandola P, Carubbi C,
Giannini S, Donati D and Vitale M: A rapid method for obtaining
mesenchymal stem cells and platelets from bone marrow aspirate. J
Tissue Eng Regen Med. June 19–2012.(Epub ahead of print).
View Article : Google Scholar
|
|
19.
|
Maraldi T, Riccio M, Sena P, Marzona L,
Nicoli A, La Marca A, Marmiroli S, Bertacchini J, La Sala G and De
Pol A: MATER protein as substrate of PKCepsilon in human cumulus
cells. Mol Hum Reprod. 15:499–506. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
20.
|
Cenni V, Sabatelli P, Mattioli E,
Marmiroli S, Capanni C, Ognibene A, Squarzoni S, Maraldi NM, Bonne
G, Columbaro M, Merlini L and Lattanzi G: Lamin A N-terminal
phosphorylation is associated with myoblast activation: impairment
in Emery-Dreifuss muscular dystrophy. J Med Genet. 42:214–220.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
21.
|
Duca M, Dozza B, Lucarelli E, Santi S, Di
Giorgio A and Barbarella G: Fluorescent labeling of human
mesenchymal stem cells by thiophene fluorophores conjugated to a
lipophilic carrier. Chem Commun (Camb). 46:7948–7950. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
22.
|
Maraldi T, Bertacchini J, Benincasa M,
Guida M, De Pol A, Liotta LA, Petricoin E, Cocco L and Marmiroli S:
Reverse-phase protein microarrays (RPPA) as a diagnostic and
therapeutic guide in multidrug resistant leukemia. Int J Oncol.
38:427–435. 2011.PubMed/NCBI
|
|
23.
|
Gebäck T, Schulz MM, Koumoutsakos P and
Detmar M: TScratch: a novel and simple software tool for automated
analysis of monolayer wound healing assays. Biotechniques.
46:265–274. 2009.PubMed/NCBI
|
|
24.
|
Mirandola P, Sponzilli I, Gobbi G,
Marmiroli S, Rinaldi L, Binazzi R, Piccari GG, Ramazzotti G,
Gaboardi GC, Cocco L and Vitale M: Anticancer agents sensitize
osteosarcoma cells to TNF-related apoptosis-inducing ligand
downmodulating IAP family proteins. Int J Oncol. 28:127–133.
2006.PubMed/NCBI
|
|
25.
|
Martelli AM, Evangelisti C, Follo MY,
Ramazzotti G, Fini M, Giardino R, Manzoli L, McCubrey JA and Cocco
L: Targeting the phosphatidylinositol 3-kinase/Akt/mammalian target
of rapamycin signaling network in cancer stem cells. Curr Med Chem.
18:2715–2726. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
26.
|
Wu W-I, Voegtli WC, Sturgis HL, Dizon FP,
Vigers GPA and Brandhuber BJ: Crystal structure of human AKT1 with
an allosteric inhibitor reveals a new mode of kinase inhibition.
PLoS One. 5:e129132010. View Article : Google Scholar : PubMed/NCBI
|
|
27.
|
Chin YR and Toker A: The actin-bundling
protein palladin is an Akt1-specific substrate that regulates
breast cancer cell migration. Mol Cell. 38:333–344. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
28.
|
Nocentini S, Reginensi D, Garcia S,
Carulla P, Moreno-Flores MT, Wandosell F, Trepat X, Bribian A and
Del Río JA: Myelin-associated proteins block the migration of
olfactory ensheathing cells: an in vitro study using single-cell
tracking and traction force microscopy. Cell Mol Life Sci. 2012.
View Article : Google Scholar
|
|
29.
|
Fox JM, Chamberlain G, Ashton BA and
Middleton J: Recent advances into the understanding of mesenchymal
stem cell trafficking. Br J Haematol. 137:491–502. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
30.
|
Stambolic V and Woodgett JR: Functional
distinctions of protein kinase B/AKT isoforms defined by their
influence on cell migration. Trends Cell Biol. 16:461–466. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
31.
|
Toker A: Achieving specificity in Akt
signaling in cancer. Adv Enzyme Regul. 52:78–87. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
32.
|
Virtakoivu R, Pellinen T, Rantala JK,
Perälä M and Ivaska J: Distinct roles of AKT isoforms in regulating
β1-integrin activity, migration, and invasion in prostate cancer.
Mol Biol Cell. 23:3357–3369. 2012.
|
|
33.
|
Moritz A, Li Y, Guo A, Villén J, Wang Y,
MacNeill J, Kornhauser J, Sprott K, Zhou J, Possemato A, Ren JM,
Hornbeck P, Cantley LC, Gygi SP, Rush J and Comb MJ: AKT-RSK-S6
kinase signaling networks activated by oncogenic receptor tyrosine
kinases. Sci Signal. 3:ra642010.PubMed/NCBI
|