Introduction
Rho family proteins function as molecular switches
in various cellular processes, including actin cytoskeletal
organization, microtubule dynamics, vesicle trafficking, cell cycle
progression and cell polarity (1).
More than 22 Rho family proteins have been identified in humans
(2). There are three classes of
regulators of Rho proteins, namely, Rho guanine nucleotide exchange
factors (RhoGEFs), Rho GTPase-activating proteins (RhoGAPs) and
RhoGDIs. In humans, over 69 RhoGEFs and 59 RhoGAPs have been
characterized (3,4), while only three types of RhoGDIs
(RhoGDIα/RhoGDI1, RhoGDIβ/RhoGDI2/LyGDI/D4GDI, and RhoGDIγ/RhoGDI3)
have been identified (5). The
existence of a great many types of RhoGEFs and RhoGAPs enables
assignment of individual regulators to specific cellular processes.
On the other hand, because there are fewer RhoGDIs, each type must
regulate a wide range of cellular processes. Thus, RhoGDIs are
considered multifunctional central regulatory molecules for Rho
family proteins (5–8). This multifunctional nature of RhoGDIs
makes it difficult to clarify their specific roles in various
cellular events. RhoGDIα is a major RhoGDI and is universally
expressed. RhoGDIγ is expressed in brain, lung and pancreas
(9,10). RhoGDIβ was originally isolated as a
RhoGDI that is abundantly expressed in hematopoietic cells
(11,12), however, it is also expressed in
several other cell types, including keratinocytes, fibroblasts,
amnion cells (13),
non-hematopoietic tumors (14–16)
and in various normal human tissues (17). Therefore, RhoGDIβ is expected to
have a more general cellular role, not specific to the
hematopoietic cell lineage.
RhoGDIβ is implicated in cancer progression,
however, reports have presented contradictory evidence as to the
nature of the correlation between cancer progression and RhoGDIβ
expression level. RhoGDIβ was found to be upregulated in ovarian
cancer (18), breast cancer
(19), gastric cancer (20), and in pancreatic cancer cells that
show high perineural invasion (21,22).
The full-length RhoGDIβ promotes cancer cell invasion (19) and survival (23) in human breast cancer. In our
previous studies, RhoGDIβ lacking C-terminal region was identified
to induce metastasis by activating the Rac1 signaling pathway in
c-Ha-Ras-transformed fibroblasts (15,24).
In other studies, RhoGDIβ has been reported to suppress invasion
(14) and its expression is
inversely correlated with invasive capacity in human bladder cancer
cells (16). Our experiments
showed that RhoGDIβ lacking the N-terminal regulatory domain
suppresses metastasis by promoting anoikis in v-Src-transformed
fibroblasts (25). Metastasis
suppression by RhoGDIβ is enhanced by Src-induced RhoGDIβ
phosphorylation (26) and
correlates with increased Rac1 activity (27). Thus, indicating yet-undetermined
roles in cancer cells, there are inconsistent results regarding
RhoGDIβ expression and cancer progression.
To clarify the role of RhoGDIβ in cancer
progression, in the present study, we examined the subcellular
localization of RhoGDIβ and the effects of overexpression and RNAi
knockdown of RhoGDIβ in cancer cells. We found that RhoGDIβ
localized to centrosomes in human cancer cells. In HeLa cells,
exogenous GFP-tagged RhoGDIβ localized to centrosomes and its
expression resulted in prolonged mitosis and aberrant cytokinesis.
Knockdown of RhoGDIβ increased the incidence of monopolar spindle
mitosis and polyploid cells in HeLa cells. The resulting polyploid
cells were possibly caused by perturbation of centrosomal function
with a lack of RhoGDIβ. Our presented results give new insights
about the role of RhoGDIβ in cancer progression.
Materials and methods
Cells and cell culture
The human cervical cancer cell line HeLa was
provided by the late Professor Masakatsu Horikawa, Faculty of
Pharmaceutical Sciences, Kanazawa University (Kanazawa, Japan)
(28). The human colon cancer cell
lines HT-29, HCT116 and SW48 were purchased from ATCC (Rockville,
MD). The human colon cancer cell lines DLD-1 and LoVo were
purchased from the Human Science Research Resources Bank (Osaka,
Japan). The human colon cancer cell lines SW480 and SW620 (29) were provided by Dr Ryuichi Yatani,
Mie University School of Medicine (Mie, Japan). These cells were
cultured in Dulbecco’s modified Eagle’s medium (DMEM) containing
10% fetal bovine serum (FBS), penicillin (50 U/ml), and
streptomycin (50 μg/ml) at 37°C in a humidified atmosphere
of 5% CO2 and 95% air. Human microvascular endothelial
cells (HMVEC) (Kurabo Industries Ltd., Osaka, Japan) were
maintained in HuMedia-MvG in accordance with the supplier’s
instructions. Immortal OKF keratinocytes (OKF6/TERT-2) were kindly
provided by Dr J.G. Rheinwald (Harvard Medical School, Boston, MA)
and were cultured in keratinocyte serum-free medium (K-sfm, BD
Biosciences, San Diego, CA) with 30 μg/ml bovine pituitary
extract, 0.1 ng/ml EGF and 0.4 mM Ca2+(30). This culture condition permits cells
to form cadherin- and desmosome-mediated junctions, and for some
cells to stratify.
Antibodies
Anti-RhoGDIα antibody (sc-360), anti-RhoGDIβ
antibody (sc-6047) and the blocking peptide (sc-6047P) for the
anti-RhoGDIβ antibody (sc-6047) were purchased from Santa Cruz
Biotechnology Inc. (Santa Cruz, CA). Anti-GFP (ab290),
anti-γ-tubulin (clone GTU-88) and anti-RhoGDIβ (556498) antibodies
were purchased from Abcam (Cambridge, UK), Sigma-Aldrich (St.
Louis, MO) and BD Biosciences (San Jose, CA), respectively. The
antigen peptides for the anti-RhoGDIβ (sc-6047) and anti-RhoGDIβ
(556498) antibodies were amino acid residues 175–194 and 4–21,
respectively, of human RhoGDIβ. Anti-RhoGDIα/RhoGDIβ antibody
(66586E) was purchased from Pharmingen (San Diego, CA).
Peroxidase-conjugated anti-mouse and anti-rabbit IgG antibodies
were purchased from DakoCytomation (Glostrup, Denmark).
Peroxidase-conjugated anti-goat IgG antibody was purchased from
Nichirei Corporation (Tokyo, Japan).
Preparation of mouse organs for
immunoblotting
Six-week-old female ICR mice were obtained from
Japan SLC Inc. (Shizuoka, Japan) and maintained under pathogen-free
conditions. Seven-week-old mice were anesthetized with
pento-barbital, then blood was gently drained from the inferior
vena cava using a heparinized syringe equipped with a 22-gauge
needle. Leukocytes and erythrocytes were isolated from blood using
Lympholyte®-Mammal (Cedarlane Laboratories Ltd.,
Burlington, Canada) according to the manufacturer’s instructions
and were lysed in Laemmli buffer (31). After blood collection, organs were
quickly removed, washed with ice-cold phosphate-buffered saline
(PBS), minced into small pieces and lysed in Laemmli buffer. All
experiments using mice were approved by the Committee on
Experimental Animals at Kanazawa Medical University and conducted
in accordance with their guidelines.
Immunoblotting
Protein concentrations of lysed cells and organs
were measured using the Bradford ULTRA kit (Novexin Ltd.,
Cambridge, UK). Proteins were resolved by sodium dodecyl
sulfate-polyacrylamide gel electrophoresis and transferred to
Immobilon-P membranes (Millipore, Billerica, MA). The membranes
were then probed with a primary antibody, followed by incubation
with a peroxidase-conjugated secondary antibody. Immunoreactive
proteins were visualized using ECL Plus reagents (GE Healthcare UK
Ltd., Little Chalfont, UK). In the immunoblot experiments, we used
RhoGDIα as a loading control because of its universal expression.
We applied the same amount of protein for immunoblot experiments.
The samples from organs contain several kinds of extracellular
matrices at various levels and the cell numbers contained in each
samples was not constant, therefore the expression levels of
RhoGDIα among various organs were not as constant as those among
cultured cell lines.
Immunofluorescence staining
For immunofluorescence staining, cells were grown on
35-mm culture dishes (BD Biosciences, San Jose, CA) or Lab-Tek II
chamber slides (Nalge Nunc International, Naperville, IL). Cells
were fixed with freshly prepared 4% paraformaldehyde for 2 min,
permeabilized with 0.5% Triton X-100 for 10 min and fixed again
with 4% paraformaldehyde for 10 min at room temperature. In some
experiments, cells were simply fixed with 99.8% methanol for 30
min. After washing with PBS, cells were incubated with 0.5% bovine
serum albumin (BSA) in PBS for 60 min at room temperature and then
incubated overnight at 4°C with primary antibodies diluted in 0.5%
BSA in PBS (1:2,000 for anti-γ-tubulin antibody or 1:200 for all
other primary antibodies). After three washes with PBS, cells were
incubated for 60 min at room temperature with secondary antibodies
(Invitrogen, Carlsbad, CA) diluted 1:400 with 0.5% BSA in PBS
containing 0.1 μg/ml 4′,6-diamidino-2-phenylindole (DAPI).
After washing with PBS, cells were mounted with Prolong Gold
(Invitrogen). For the blocking experiment, anti-RhoGDIβ antibody
(sc-6047) was incubated with 10-fold concentration of the blocking
peptide for 60 min at room temperature before use. Images were
obtained using an Axiovert 200 inverted fluorescence microscope or
LSM710 confocal microscope (Carl Zeiss, Jena, Germany).
Plasmids and transfection
The entire sequence was amplified by PCR using
pcDNA3.1-LyGDI, an expression vector for wild-type human RhoGDIβ
(15). The product was then
subcloned into pAcGFP1-C3 and used as pAcGFP-RhoGDIβ. Cells were
transfected with the expression plasmids using Lipofectamine 2000
(Invitrogen). To obtain cell lines that stably expressed the
introduced genes, G418-resistant cells were isolated in medium
containing 800 μg/ml G418 (Nacalai Tesque Inc., Kyoto,
Japan).
Observation of GFP-RhoGDIβ in living
cells and time-lapse analysis
HeLa cells that stably expressed GFP-RhoGDIβ were
cultured in 35-mm glass-bottomed dishes (Asahi Glass Co. Ltd.,
Tokyo, Japan). The cells were maintained at 37°C in a humidified
atmosphere of 5% CO2 and 95% air in an enclosed stage
incubator (Incubator-XL, Carl Zeiss) built on top of an Axiovert
200 M inverted microscope. Time-lapse images of green fluorescence
and differential interference contrast (DIC) were acquired on an
Axiovert 200 M controlled by AxioVision image processing and
analysis system 4.4 (Carl Zeiss). The onset of mitosis was
considered to be the beginning of cell rounding, the onset of
anaphase was defined as the beginning of chromosome segregation and
the end of cytokinesis was recognized when the cleavage furrow
maximally contracted. We analyzed the progression of mitosis
temporally in cells in which these mitotic processes could be
clearly recognized. The entire duration of mitosis was measured as
the time from the beginning of cell rounding to the maximal
contraction of the cleavage furrow.
Small interfering RNA (siRNA)
Three different 25-mer Stealth RNAi duplexes
targeting human RhoGDIβ and negative control Stealth RNAi were
purchased from Invitrogen. The sequences of the three Stealth RNAi
are as follows: no. 153, sense 5′-GAGCUGGACAGCAAGCUCAAUUAUA-3′,
anti-sense 5′-UAUAAUUGAGCUUGCUGUCCAGCUC-3′; no. 469, sense
5′-GCCUGAAAUACGUUCAGCACACCUA-3′, anti-sense
5′-UAGGUGUGCUGAACGUAUUUCAGGC-3′; and no. 800, sense
5′-GGUCCCUCUUCAACACUGCCACAUU-3′, anti-sense
5′-AAUGUGGCAGUGUUGAAGAGGGACC-3′. Stealth RNAi duplexes were
transfected into HeLa cells using Lipofectamine 2000 according to
the manufacturer’s protocol.
DNA histograms
Cells were fixed with 20% ethanol and incubated with
0.1% RNase (Type II-A, Sigma-Aldrich, St. Louis, MO) for 30 min at
37°C. The cells were stained with propidium iodide (50
μg/ml) and analyzed using a FACSort flow cytometer (BD
Biosciences, San Jose, CA).
Statistical analysis
Differences between values were analyzed by the
two-tailed Mann-Whitney U-test using the statistics function of
KaleidaGraph (Version 4.1). P<0.05 was considered
significant.
Results
Protein expression levels of RhoGDIβ
To confirm the previous observations showing the
expression of RhoGDIβ in cells other than those in the
hematopoietic cell lineage (13–17),
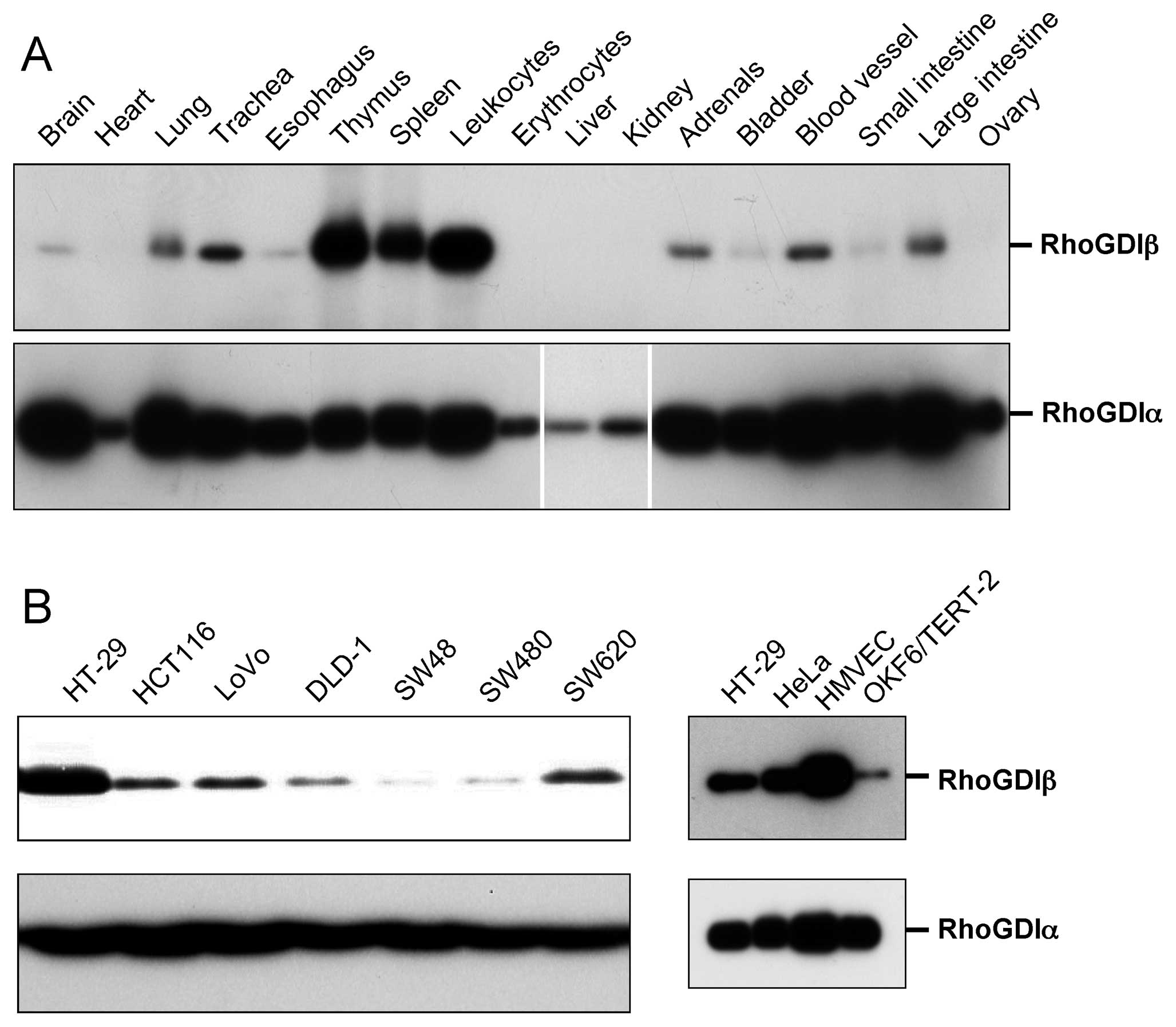
we examined the levels of RhoGDIβ protein in various mouse organs
(Fig. 1A, upper panel). RhoGDIβ
was expressed in mouse brain, lung, trachea, esophagus, adrenals,
bladder, blood vessels, small and large intestine, although the
expression levels were much lower than in hematopoietic cells such
as those from the thymus, spleen and leukocytes. Blood was
thoroughly drained before the isolation of organs, but slight
contamination of hematopoietic cells was unavoidable. Therefore, it
could not be ruled out that very low expression of RhoGDIβ reflects
contamination of hematopoietic cells. In contrast, RhoGDIα was
expressed at various levels in all examined tissues (Fig. 1A, lower panel). Consistent with the
expression of RhoGDIβ in the large intestine and in blood vessels,
RhoGDIβ was expressed in cultured human colon cancer cells and
microvascular endothelial cells (HMVEC) (Fig. 1B). RhoGDIβ was also expressed in
other types of cultured epithelial cells, such as HeLa and
OKF6/TERT-2 human keratinocytes (Fig.
1B).
Subcellular localization of RhoGDIβ
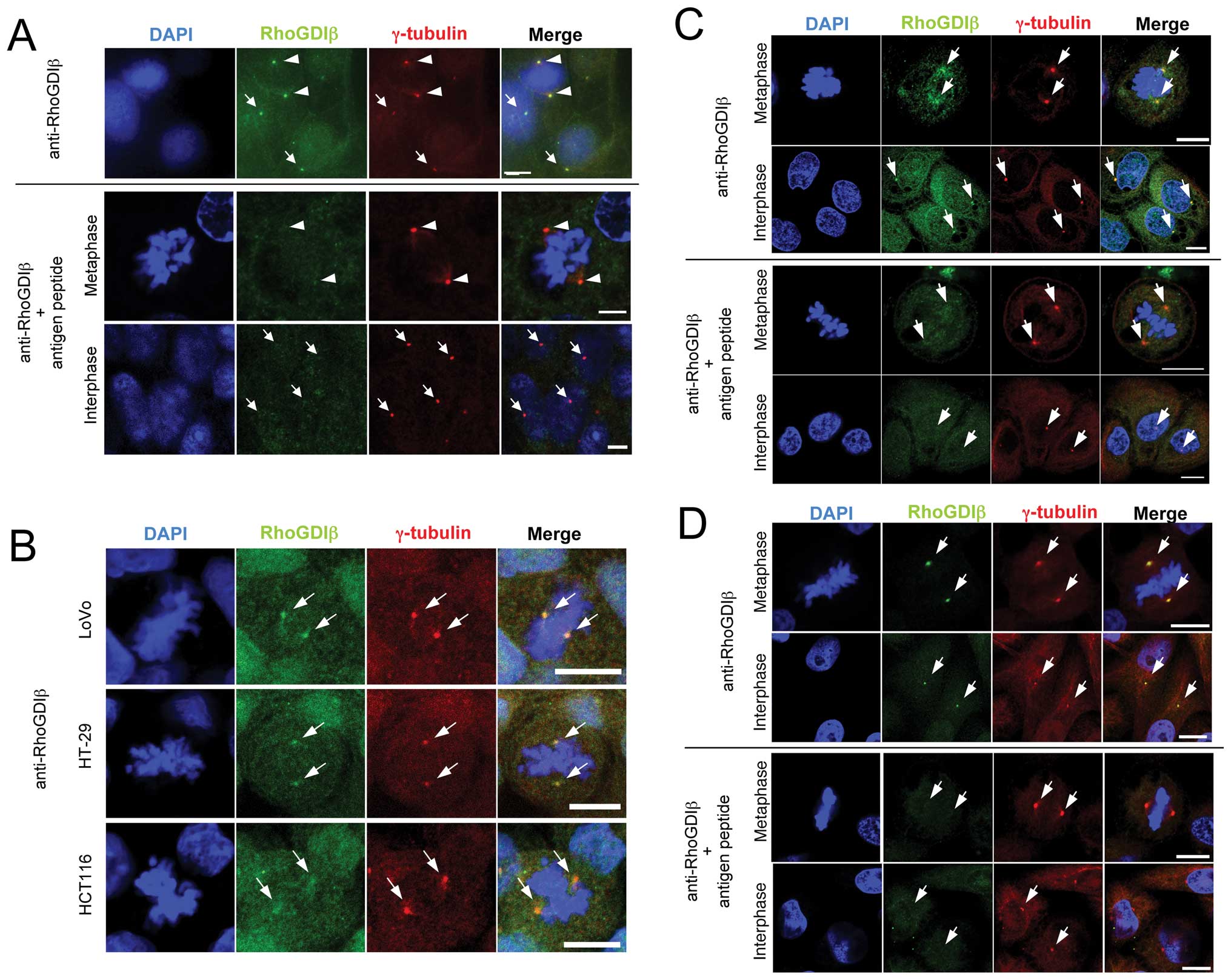
We examined the subcellular localization of RhoGDIβ
in cultured colon cancer cells by immunofluorescence staining.
Simultaneous staining with anti-RhoGDIβ and anti-γ-tubulin
antibodies showed the colocalization of them in DLD-1 human colon
cancer cells (Fig. 2A, upper
panels). Similar results were also obtained in HT-29, HCT116, LoVo,
SW48, SW480 and SW620 human colon cancer cell lines. Representative
images of simultaneous staining with anti-RhoGDIβ and
anti-γ-tubulin antibodies in LoVo, HT-29 and HCT116 human colon
cancer cells are shown in Fig. 2B.
RhoGDIβ colocalized with γ-tubulin also in OKF6/TERT-2 human
keratinocyte (Fig. 2C, upper
panels) and HeLa cells (Fig. 2D,
upper panels). During interphase, the colocalization of RhoGDIβ
with γ-tubulin was not as clear as during metaphase. Such
centrosome staining pattern is likely to be associated with
centrosome maturation. The staining with anti-RhoGDIβ antibody was
abolished by pretreatment of the antibody with antigen peptide
(Fig. 2A, C and D, lower panels).
Unlike RhoGDIβ, RhoGDIα did not colocalize with γ-tubulin in colon
cancer cells and HeLa cells (data not shown).
Subcellular localization of GFP-RhoGDIβ
in HeLa cells
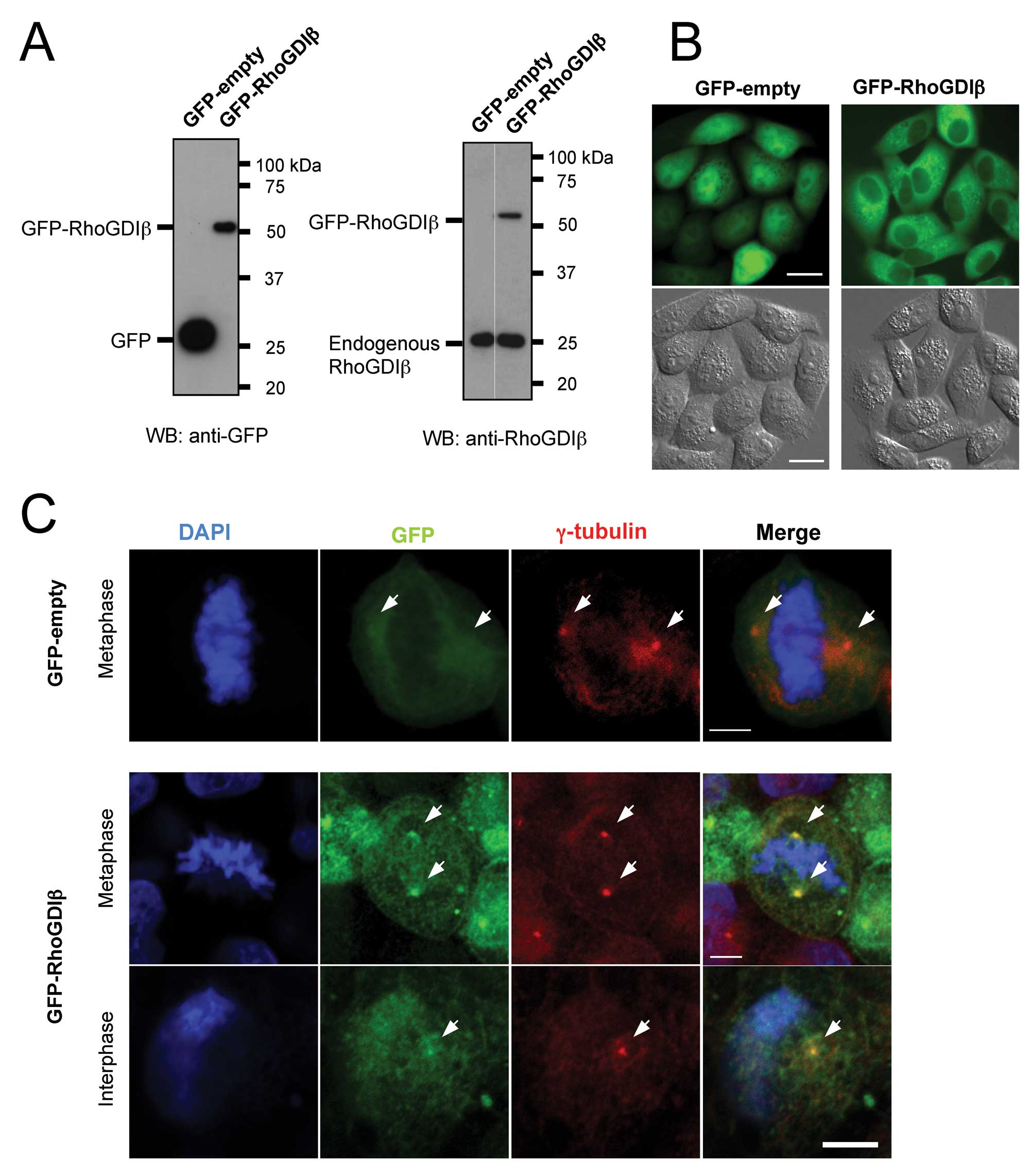
We used GFP-RhoGDIβ to confirm the localization of
RhoGDIβ to centrosomes. pAcGFP-RhoGDIβ was transfected into HeLa
cells and the cells stably expressing the introduced genes were
selected (Fig. 3A). Expression of
GFP-RhoGDIβ did not affect the levels of endogenous RhoGDIβ protein
(Fig. 3A, right panel). In living
cells, localization of GFP-RhoGDIβ was distinct from localization
of GFP only. Specifically, GFP-empty localized all over the cell,
although it was brighter in the nuclei than in cell cytoplasm,
while GFP-RhoGDIβ localized predominantly to the cytoplasm
(Fig. 3B). It was difficult to
observe centrosomal localization of GFP-RhoGDIβ in living cells
because of its granular localization throughout the cytoplasm. To
confirm the centrosomal localization of GFP-RhoGDIβ, cells were
fixed and stained with both anti-GFP and anti-γ-tubulin antibodies.
GFP-RhoGDIβ colocalized with γ-tubulin in both metaphase and
interphase cells, while GFP-empty did not (Fig. 3C).
Time-lapse observation of mitosis in HeLa
cells stably expressing GFP-RhoGDIβ
Our observations suggested that RhoGDIβ had a role
related to centrosome function. To investigate this, we used
time-lapse microscopy to observe the mitotic progression in HeLa
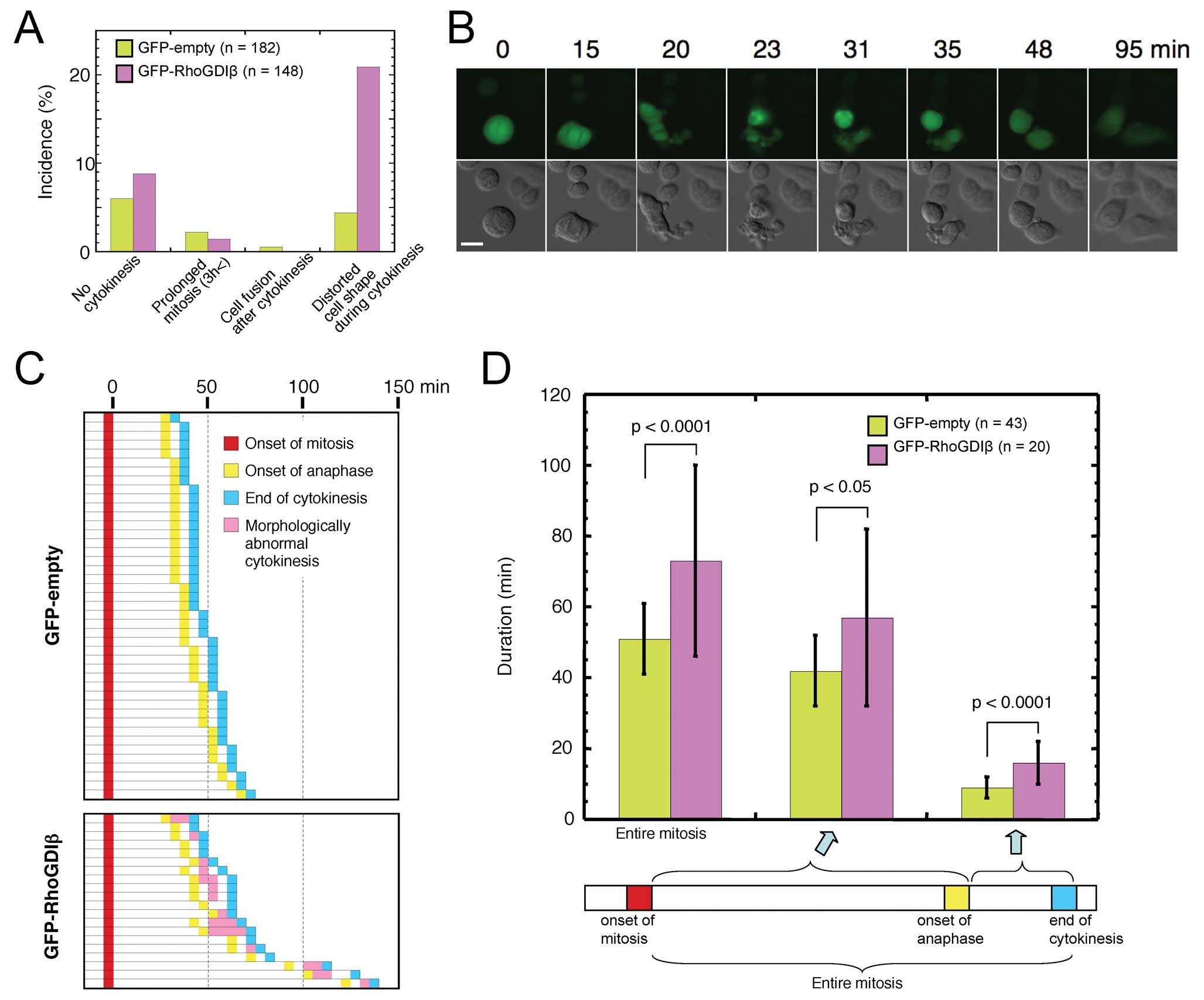
cells stably expressing GFP-RhoGDIβ. The incidence of
morphologically aberrant cytokinesis, in which the cell shape was
distorted, increased about 4-fold in GFP-RhoGDIβ expressing cells
(Fig. 4A). Representative images
of aberrant cytokinesis of GFP-RhoGDIβ-expressing cell are shown
(Fig. 4B). To examine the defects
in cytokinesis in detail we analyzed the time-lapse images of the
cells, in which the onset of mitosis, the onset of anaphase, and
the end of cytokinesis could be clearly distinguished (Fig. 4C). In these cells, morphologically
aberrant cytokinesis was observed in 70% (14/20) of cells
expressing GFP-RhoGDIβ, but was not observed in cells expressing
GFP-empty. Duration from the onset of anaphase and from the onset
of anaphase to the end of cytokinesis was significantly increased
in cells expressing GFP-RhoGDIβ compared with those of cells
expressing GFP-empty (Fig. 4D).
These observations indicated that GFP-RhoGDIβ affected the mitotic
processes including anaphase and cytokinesis in HeLa cells,
suggesting that RhoGDIβ plays a role in these mitotic
processes.
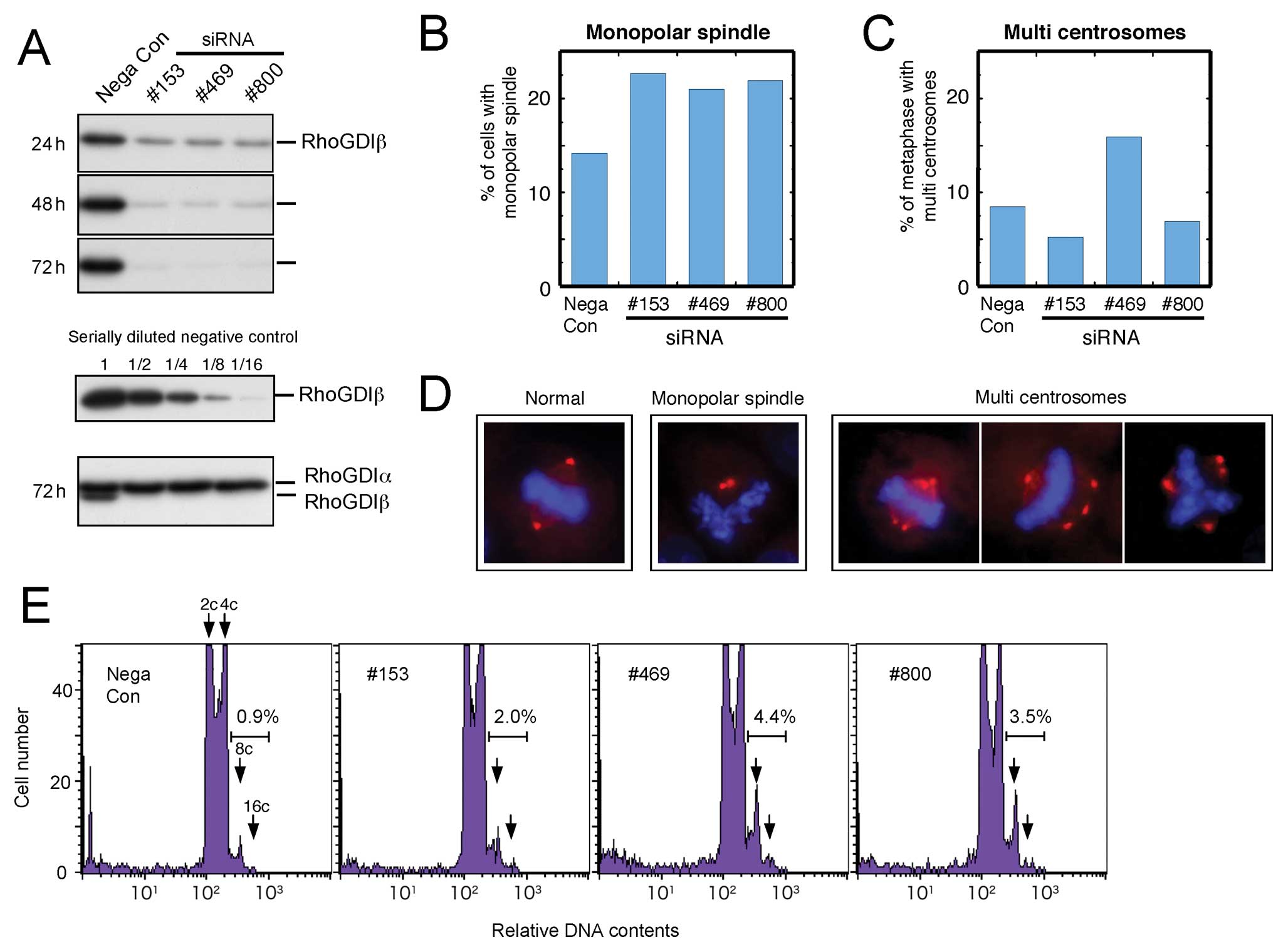
Effect of knockdown of RhoGDIβ by
siRNA
We examined the effect of RhoGDIβ knockdown in HeLa
cells using three different siRNAs. All siRNAs decreased the
expression of RhoGDIβ by about 90% and did not decrease the
expression of RhoGDIα 72 h after transfection (Fig. 5A). Knockdown of RhoGDIβ by all
three siRNAs increased the incidence of monopolar spindle mitosis
(Fig. 5B). In contrast with the
appearance of monopolar mitotic cells, a slightly higher frequency
of multiple centrosomes was observed only in cells that had RhoGDIβ
knocked down by no. 469 siRNA (Fig.
5C). Representative images of monopolar spindle mitosis and
multiple centrosomes mitosis are shown (Fig. 5D). Therefore, we concluded that
RhoGDIβ knockdown induced inhibition of centrosome separation
rather than multipolar mitosis. The increase of polyploid (8, 16c
or more) cells after RhoGDIβ knockdown as shown in Fig. 5E were thought to mainly result from
monopolar mitosis. Overall, our data show that both upregulation
and downregulation of RhoGDIβ constitute key events leading to
perturbed mitotic processes in cancer cells.
Discussion
RhoGDIβ is abundantly expressed in hematopoietic
cells (11), but is also expressed
in various non-hematopoietic cells (13–17).
Using immunoblotting, we confirmed that RhoGDIβ protein is
expressed in many human epithelial cell lines as well as in many
mouse organs, suggesting a general role for RhoGDIβ that is not
specific to hematopoietic cells. We showed that RhoGDIβ, but not
RhoGDIα, localized to centrosomes in human colon cancer cells,
human keratinocytes and HeLa cells by immunofluorescence staining.
Furthermore, we showed that exogenously introduced GFP-RhoGDIβ also
localized to centrosomes in HeLa cells. Previously, RhoGDIβ was
identified in the purified centrosome fraction from a human
lymphoblastic cell line by the proteomic analysis, however, it was
not confirmed as a genuine centrosomal protein (32). Since RhoGDIβ functions as a
chaperone (8), its association
with the centrosome would be expected to be transient and less
stable than those of the scaffold proteins of centrosomes.
Furthermore, RhoGDIβ is abundant in the cytosol. These properties
of RhoGDIβ make it difficult to clarify its localization to the
centrosome. In the present study, we confirmed that RhoGDIβ
localized to centrosomes and suggested that RhoGDIβ had a role
related to centrosome function. Actually, we showed that the
expression of GFP-RhoGDIβ significantly prolonged anaphase and
cytokinesis and increased the morphologically aberrant cytokinesis
in HeLa cells. Furthermore, RhoGDIβ knockdown caused defects in
centrosome separation. Supporting a previous proteomics study
(32), collectively, our present
data regarding localization and functional analyses strongly
suggest that RhoGDIβ functions in centrosomes during mitosis.
Rho family proteins are important regulators of both
cytokinesis and centrosome positioning (33). RhoA is the central regulator of
cytokinesis in animal cells (34).
Cdc42 and MgcRacGAP are reported to contribute to the correct
formation of mitotic spindles during metaphase in HeLa cells
(35). Rac and Tiam1 are localized
to the centrosomal regions during early mitosis, and bipolar
spindle formation is regulated by Tiam1-Rac signaling in MDCK II
cells (36). Therefore, our
results collectively suggest that RhoGDIβ, that is a regulator for
Rho proteins, plays a role in the regulation of cytokinesis and the
formation of bipolar spindles. To our knowledge this is the first
report suggesting that RhoGDIβ participates in the regulation of
these mitotic processes.
In normal cells, the mitotic checkpoint prevents the
transition to anaphase when the monopolar spindle is formed;
however, in cancer cells, this checkpoint is compromised and some
cells become polyploid through aberrant mitosis (37). Therefore, an increase of polyploid
cells by knockdown of RhoGDIβ could be due to the increased
incidence of monopolar spindle. The monopolar spindles can result
from defects in many different molecules, such as specific motor
proteins, centrosome proteins, and mitotic kinases (38). Which of these molecules are
associated with the observed phenotype by RhoGDIβ knockdown is
unknown, but RhoGDIβ should be involved in the formation of normal
bipolar spindles when required.
Correct centriole and centrosome positioning is
important for many biological processes (39) and centrosomes play important roles
in maintaining the polarity axis during asymmetrical cell division,
and disruption of polarity is implicated in cancer development and
progression (40–42). Cell motility, invasion, and anoikis
in cancer progression are regulated by RhoGDIβ in cancer
progression, irrespective of the direction of correlation with
RhoGDIβ expression (14,15,19,21,23,25,27)
and are closely associated with cell polarity (43,44).
There is crosstalk between Rho family proteins and polarity
proteins (44). The roles of
RhoGDIβ in cancer progression may be related, at least in part, to
its role in the regulation of cell polarity. RhoGDIs are conserved
among eukaryotes and are suggested to have a universal role in the
regulation of cell polarity in a wide range of eukaryotes (45–51).
Many unicellular eukaryotes and lower metazoa have a single RhoGDI
(52), while vertebrates, except
for bony fish, have three kinds of RhoGDIs. Our results suggest
that among RhoGDIs at least RhoGDIβ plays a role in the regulation
of cell polarity in mammalian cells.
Acknowledgements
This study was supported in part by a
Grant-in-Aid for Scientific Research from the Japan Society for the
Promotion of Science (20591596).
References
|
1.
|
Etienne-Manneville S and Hall A: Rho
GTPases in cell biology. Nature. 420:629–635. 2002. View Article : Google Scholar
|
|
2.
|
Wennerberg K and Der CJ: Rho-family
GTPases: it’s not only Rac and Rho (and I like it). J Cell Sci.
117:1301–1312. 2004.
|
|
3.
|
Rossman KL, Der CJ and Sondek J: GEF means
go: turning on RHO GTPases with guanine nucleotide-exchange
factors. Nat Rev Mol Cell Biol. 6:167–180. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
4.
|
Tcherkezian J and Lamarche-Vane N: Current
knowledge of the large RhoGAP family of proteins. Biol Cell.
99:67–86. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
5.
|
Dovas A and Couchman JR: RhoGDI: multiple
functions in the regulation of Rho family GTPase activities.
Biochem J. 390:1–9. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
6.
|
DerMardirossian C and Bokoch GM: GDIs:
central regulatory molecules in Rho GTPase activation. Trends Cell
Biol. 15:356–363. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
7.
|
Dransart E, Olofsson B and Cherfils J:
RhoGDIs revisited: novel roles in Rho regulation. Traffic.
6:957–966. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
8.
|
Garcia-Mata R, Boulter E and Burridge K:
The ‘invisible hand’: regulation of RHO GTPases by RHOGDIs. Nat Rev
Mol Cell Biol. 12:493–504. 2011.
|
|
9.
|
Adra CN, Manor D, Ko JL, et al: RhoGDIγ: a
GDP-dissociation inhibitor for Rho proteins with preferential
expression in brain and pancreas. Proc Natl Acad Sci USA.
94:4279–4284. 1997.
|
|
10.
|
Zalcman G, Closson V, Camonis J, et al:
RhoGDI-3 is a new GDP dissociation inhibitor (GDI). Identification
of a non-cytosolic GDI protein interacting with the small
GTP-binding proteins RhoB and RhoG. J Biol Chem. 271:30366–30374.
1996. View Article : Google Scholar : PubMed/NCBI
|
|
11.
|
Lelias JM, Adra CN, Wulf GM, et al: cDNA
cloning of a human mRNA preferentially expressed in hematopoietic
cells and with homology to a GDP-dissociation inhibitor for the rho
GTP-binding proteins. Proc Natl Acad Sci USA. 90:1479–1483. 1993.
View Article : Google Scholar : PubMed/NCBI
|
|
12.
|
Scherle P, Behrens T and Staudt LM:
Ly-GDI, a GDP-dissociation inhibitor of the RhoA GTP-binding
protein, is expressed preferentially in lymphocytes. Proc Natl Acad
Sci USA. 90:7568–7572. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
13.
|
Leffers H, Nielsen MS, Andersen AH, et al:
Identification of two human Rho GDP dissociation inhibitor proteins
whose overexpression leads to disruption of the actin cytoskeleton.
Exp Cell Res. 209:165–174. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
14.
|
Gildea JJ, Seraj MJ, Oxford G, et al:
RhoGDI2 is an invasion and metastasis suppressor gene in human
cancer. Cancer Res. 62:6418–6423. 2002.PubMed/NCBI
|
|
15.
|
Ota T, Maeda M, Suto S and Tatsuka M:
LyGDI functions in cancer metastasis by anchoring Rho proteins to
the cell membrane. Mol Carcinog. 39:206–220. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
16.
|
Seraj MJ, Harding MA, Gildea JJ, Welch DR
and Theodorescu D: The relationship of BRMS1 and RhoGDI2 gene
expression to metastatic potential in lineage related human bladder
cancer cell lines. Clin Exp Metastasis. 18:519–525. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
17.
|
Theodorescu D, Sapinoso LM, Conaway MR,
Oxford G, Hampton GM and Frierson HF Jr: Reduced expression of
metastasis suppressor RhoGDI2 is associated with decreased survival
for patients with bladder cancer. Clin Cancer Res. 10:3800–3806.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
18.
|
Tapper J, Kettunen E, El-Rifai W, Seppala
M, Andersson LC and Knuutila S: Changes in gene expression during
progression of ovarian carcinoma. Cancer Genet Cytogenet. 128:1–6.
2001. View Article : Google Scholar : PubMed/NCBI
|
|
19.
|
Zhang Y and Zhang B: D4-GDI, a Rho GTPase
regulator, promotes breast cancer cell invasiveness. Cancer Res.
66:5592–5598. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
20.
|
Cho HJ, Baek KE, Park SM, et al: RhoGDI2
expression is associated with tumor growth and malignant
progression of gastric cancer. Clin Cancer Res. 15:2612–2619. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
21.
|
Abiatari I, DeOliveira T, Kerkadze V, et
al: Consensus transcriptome signature of perineural invasion in
pancreatic carcinoma. Mol Cancer Ther. 8:1494–1504. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
22.
|
Koide N, Yamada T, Shibata R, et al:
Establishment of peri-neural invasion models and analysis of gene
expression revealed an invariant chain (CD74) as a possible
molecule involved in perineural invasion in pancreatic cancer. Clin
Cancer Res. 12:2419–2426. 2006. View Article : Google Scholar
|
|
23.
|
Zhang Y, Rivera Rosado LA, Moon SY and
Zhang B: Silencing of D4-GDI inhibits growth and invasive behavior
in MDA-MB-231 cells by activation of Rac-dependent p38 and JNK
signaling. J Biol Chem. 284:12956–12965. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
24.
|
Ota T, Maeda M, Murakami M, Takegami T,
Suto S and Tatsuka M: Activation of Rac1 by Rho-guanine nucleotide
dissociation inhibitor-β with defective isoprenyl-binding pocket.
Cell Biol Int. 31:92–96. 2007.
|
|
25.
|
Ota T, Maeda M, Sakita-Suto S, et al:
RhoGDIβ lacking the N-terminal regulatory domain suppresses
metastasis by promoting anoikis in v-src-transformed cells. Clin
Exp Metastasis. 23:323–334. 2006.
|
|
26.
|
Wu Y, Moissoglu K, Wang H, et al: Src
phosphorylation of RhoGDI2 regulates its metastasis suppressor
function. Proc Natl Acad Sci USA. 106:5807–5812. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
27.
|
Moissoglu K, McRoberts KS, Meier JA,
Theodorescu D and Schwartz MA: Rho GDP dissociation inhibitor 2
suppresses metastasis via unconventional regulation of RhoGTPases.
Cancer Res. 69:2838–2844. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
28.
|
Scherer WF, Syverton JT and Gey GO:
Studies on the propagation in vitro of poliomyelitis viruses. IV.
Viral multiplication in a stable strain of human malignant
epithelial cells (strain HeLa) derived from an epidermoid carcinoma
of the cervix. J Exp Med. 97:695–710. 1953. View Article : Google Scholar
|
|
29.
|
Leibovitz A, Stinson JC, McCombs WB III,
McCoy CE, Mazur KC and Mabry ND: Classification of human colorectal
adenocarcinoma cell lines. Cancer Res. 36:4562–4569.
1976.PubMed/NCBI
|
|
30.
|
Dickson MA, Hahn WC, Ino Y, et al: Human
keratinocytes that express hTERT and also bypass a
p16INK4a-enforced mechanism that limits life span become immortal
yet retain normal growth and differentiation characteristics. Mol
Cell Biol. 20:1436–1447. 2000. View Article : Google Scholar
|
|
31.
|
Laemmli UK: Cleavage of structural
proteins during the assembly of the head of bacteriophage T4.
Nature. 227:680–685. 1970. View
Article : Google Scholar : PubMed/NCBI
|
|
32.
|
Andersen JS, Wilkinson CJ, Mayor T,
Mortensen P, Nigg EA and Mann M: Proteomic characterization of the
human centrosome by protein correlation profiling. Nature.
426:570–574. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
33.
|
Narumiya S and Yasuda S: Rho GTPases in
animal cell mitosis. Curr Opin Cell Biol. 18:199–205. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
34.
|
Piekny A, Werner M and Glotzer M:
Cytokinesis: welcome to the Rho zone. Trends Cell Biol. 15:651–658.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
35.
|
Ban R, Irino Y, Fukami K and Tanaka H:
Human mitotic spindle-associated protein PRC1 inhibits MgcRacGAP
activity towards Cdc42 during the metaphase. J Biol Chem.
279:16394–16402. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
36.
|
Woodcock SA, Rushton HJ, Castaneda-Saucedo
E, et al: Tiam1-Rac signaling counteracts Eg5 during bipolar
spindle assembly to facilitate chromosome congression. Curr Biol.
20:669–675. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
37.
|
Weaver BA and Cleveland DW: Does
aneuploidy cause cancer? Curr Opin Cell Biol. 18:658–667. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
38.
|
Tillement V, Remy MH, Raynaud-Messina B,
Mazzolini L, Haren L and Merdes A: Spindle assembly defects leading
to the formation of a monopolar mitotic apparatus. Biol Cell.
101:1–11. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
39.
|
Vaughan S and Dawe HR: Common themes in
centriole and centrosome movements. Trends Cell Biol. 21:57–66.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
40.
|
Feigin ME and Muthuswamy SK: Polarity
proteins regulate mammalian cell-cell junctions and cancer
pathogenesis. Curr Opin Cell Biol. 21:694–700. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
41.
|
Knoblich JA: Asymmetric cell division:
recent developments and their implications for tumour biology. Nat
Rev Mol Cell Biol. 11:849–860. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
42.
|
Lee M and Vasioukhin V: Cell polarity and
cancer - cell and tissue polarity as a non-canonical tumor
suppressor. J Cell Sci. 121:1141–1150. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
43.
|
Fischer AH, Zhao C, Li QK, et al: The
cytologic criteria of malignancy. J Cell Biochem. 110:795–811.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
44.
|
Iden S and Collard JG: Crosstalk between
small GTPases and polarity proteins in cell polarization. Nat Rev
Mol Cell Biol. 9:846–859. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
45.
|
Carol RJ, Takeda S, Linstead P, et al: A
RhoGDP dissociation inhibitor spatially regulates growth in root
hair cells. Nature. 438:1013–1016. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
46.
|
Court H and Sudbery P: Regulation of Cdc42
GTPase activity in the formation of hyphae in Candida
albicans. Mol Biol Cell. 18:265–281. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
47.
|
Imai K, Kijima T, Noda Y, Sutoh K, Yoda K
and Adachi H: A Rho GDP-dissociation inhibitor is involved in
cytokinesis of Dictyostelium. Biochem Biophys Res Commun.
296:305–312. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
48.
|
Price MS, Nichols CB and Alspaugh JA: The
Cryptococcus neoformans Rho-GDP dissociation inhibitor
mediates intra-cellular survival and virulence. Infect Immun.
76:5729–5737. 2008.
|
|
49.
|
Richman TJ, Toenjes KA, Morales SE, et al:
Analysis of cell-cycle specific localization of the Rdi1p RhoGDI
and the structural determinants required for Cdc42p membrane
localization and clustering at sites of polarized growth. Curr
Genet. 45:339–349. 2004. View Article : Google Scholar
|
|
50.
|
Rivero F, Illenberger D, Somesh BP,
Dislich H, Adam N and Meyer AK: Defects in cytokinesis, actin
reorganization and the contractile vacuole in cells deficient in
RhoGDI. EMBO J. 21:4539–4549. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
51.
|
Yap SF, Chen W and Lim L: Molecular
characterization of the Caenorhabditis elegans Rho
GDP-dissociation inhibitor. Eur J Biochem. 266:1090–1100. 1999.
|
|
52.
|
Menotta M, Amicucci A, Basili G, Polidori
E, Stocchi V and Rivero F: Molecular and functional
characterization of a Rho GDP dissociation inhibitor in the
filamentous fungus Tuber borchii. BMC Microbiol. 8:572008.
View Article : Google Scholar : PubMed/NCBI
|