Introduction
Colorectal cancer (CRC) is one of the most common
forms of cancer and a leading cause of cancer-related deaths around
the world (1). To date,
chemotherapy is one of the main therapeutic approaches for patients
with advanced CRC (2,3). However, both drug resistance and
toxicity limit the effectiveness of current CRC chemotherapy, thus
there is a need for the development of new therapeutic agents.
Recently, natural products received great interest since they have
relatively fewer side-effects and have been used clinically to
treat various kinds of diseases including cancer for thousands of
years. Numerous plants and their constituents have been shown to
possess beneficial therapeutic effects for cancer (4,5).
Therefore, discovering naturally occurring agents is a promising
approach for cancer chemotherapy.
Angiogenesis plays a critical role in the growth and
development of cancer, suggesting that anti-angiogenic therapy is
effective in inhibiting tumor progression (6–8).
Tumor angiogenesis is regulated by multiple cellular signaling
pathways including hedgehog (HH) transduction cascade. HH pathway
is important for embryonic development (9) and its aberrant activation is
implicated as an initiating or maintaining factor in the
progression of various human cancers (9–17).
Mammals have three hedgehog homologues (Sonic hedgehog, Indian
hedgehog and Desert hedgehog), of which Sonic hedgehog (SHH) is the
best studied. Activation of HH signaling is initiated by binding of
HH to the transmembrane receptor Patched (Ptch), resulting in the
release of Ptch-mediated suppression of Smoothened (Smo), a
G-protein-coupled receptor. Released Smo subsequently activates the
Gli family of transcription factors that regulate the expression of
HH target genes (9,17–19),
including vascular endothelial growth factor-A (VEGF-A). VEGF-A is
considered as one of the most potent angiogenic factors
overexpressed in many human cancers (20–24).
VEGF-A secreted from tumor cells primarily binds to specific
receptors such as VEGFR-2 that are located on vascular endothelial
cells (EC) (17,25–27).
Binding of VEGF-A to VEGFR-2 in turn triggers a tyrosine kinase
signaling cascade that induces EC proliferation, migration,
survival, sprouting and tube formation (17,25,28–30).
Thus, inhibition of angiogenesis via modulation of SHH signaling
could be a major focus for anticancer drug development.
Hedyotis diffusa Willd (HDW), belonging to
the Rubiaceae family, is a medicinal herb widely distributed in
Northeast Asia. As a well known traditional Chinese folk-medicine,
HDW has long been used as a major component in many traditional
Chinese medicine (TCM) formulas for the clinical treatment of
various kinds of cancer including CRC (31–33).
Recently, we reported that HDW can inhibit cancer growth in
vivo and in vitro via suppression of STAT3 pathway
(3,34). In addition, we proposed that HDW
displays anti-angiogenic activity in vitro(25). However, the precise mechanism of
its anticancer activity remains largely unclear. To further
elucidate the mechanism of the tumoricidal activity of Hedyotis
diffusa Willd, here we evaluated its in vivo effect on
colorectal cancer growth and tumor angiogenesis and investigated
the underlying molecular mechanisms.
Materials and methods
Materials and reagents
Dulbecco’s modified Eagle’s medium (DMEM), fetal
bovine serum (FBS), penicillin/streptomycin, Trypsin-EDTA and
TRIzol Reagent were purchased from Invitrogen (Carlsbad, CA, USA).
SuperScript II reverse transcriptase was obtained from Promega
(Madison, WI, USA). CD31, SHH, PTCH-1, SMO, Gli-1, VEGF-A and
VEGFR2 antibodies, secondary antibodies were obtained from Santa
Cruz Biotechnology (Santa Cruz, CA, USA). All other chemicals,
unless otherwise stated, were obtained from Sigma Chemicals (St.
Louis, MO, USA).
Preparation of ethanol extract from
Hedyotis diffusa Willd (EEHDW)
Authentic plant material was purchased from Guo Yi
Tang Chinese Herbal medicine store, Fujian, China. The original
herb was identified as Hedyotis diffusa Willd (HDW) by Dr
Wei Xu at Department of Pharmacology, Fujian University of
Traditional Chinese Medicine, China. Ethanol extract of HDW (EEHDW)
was prepared as previously described (3). Briefly, 500 g of HDW was extracted
with 5,000 ml of 85% ethanol using a refluxing method and filtered.
The ethanol solvent was then evaporated on a rotary evaporator
(Yarong, Model RE-2000, Shanghai, China). The resultant solution
was concentrated to a relative density of 1.05 and the dried powder
of EEHDW was obtained by a spraying desiccation method using a
spray dryer (Buchi, Model B-290, Flawil, Switzerland). The working
concentrations of EEHDW were made by dissolving the extract in
saline to a concentration of 0.6 g/ml.
Cell culture and animals
Human colon carcinoma cell line HT-29 was obtained
from the American Type Culture Collection (ATCC, Manassas, VA, USA)
and grown in DMEM medium supplemented with 10% serum (Gibco-BRL),
100 U/ml penicillin and 100 μg/ml streptomycin in a
humidified environment with 5% CO2 at 37°C. Male BALB/C
athymic nude mice (with an initial body weight of 20–22 g) were
purchased from Shanghai Si-Lai-Ke Experimental Animal Ltd
(Shanghai, China). The mice were housed five per cage in specific
pathogen-free room in an environment with controlled temperature
(22°C), humidity and a 12 h light/dark cycle with free access to
water and standard laboratory food. All animal treatments were
strictly in accordance with international ethical guidelines and
the National Institutes of Health Guide concerning the Care and Use
of Laboratory Animals, and the experiments were approved by the
Institutional Animal Care and Use committee of Fujian University of
Traditional Chinese Medicine.
In vivo nude mouse xenograft study
HT-29 cells (5×106) mixed with Matrigel
(1:1) were subcutaneously injected in the right flank area of
athymic nude mice to initiate tumor growth. After 5 days of
xenograft implantation, mice were randomly divided into two groups
(n=10) and given intra-gastric administration with 3 g/kg of EEHDW
or saline daily, 6 days a week for 16 days. Tumor diameters were
measured at regular intervals with digital calipers and the tumor
volume (V) was calculated using the formula: V =
(width)2 × length × π/6. At the end of experiment, the
animals were anaesthetized and the tumor issue was removed. Blood
was obtained aseptically from orbit. Blood-containing tubes were
allowed to stand at room temperature for 5 h and serums were
obtained by centrifugation at 3, 000 × g for 20 min.
RNA extraction and RT-PCR analysis
Total RNA was isolated from fresh tumor tissues with
TRIzol Reagent. Oligo(dT)-primed RNA (1 μg) was
reverse-transcribed with SuperScript II reverse transcriptase
(Promega) according to the manufacturer’s instructions. The
obtained cDNA was used to determine the mRNA amount of SHH, PTCH,
SMO, Gli-1, VEGF-A and VEGFR2 by PCR with Taq DNA polymerase
(Fermentas). GAPDH was used as an internal control. Samples were
analyzed by gel electrophoresis (1.5% agarose). The DNA bands were
examined using a Gel Documentation System (Bio-Rad, Model Gel Doc
2000, Hercules, CA, USA).
Immunohistochemical staining
After fixing with 10% formaldehyde for 12 h, tumor
samples were processed conventionally for 5-μm-thick
paraffin-embedded tumor slides. The slides were subjected to
antigen retrieval and the endogenous peroxidase activity was
quenched with hydrogen peroxide. After blocking non-specific
proteins with normal serum in PBS, slides were incubated with
rabbit polyclonal antibodies against CD31, SHH, PTCH-1, SMO, Gli-1,
VEGF-A and VEGFR2 (all in 1:100 dilution, Santa Cruz
Biotechnology). After washing with PBS, slides were incubated with
biotinylated secondary antibody followed by conjugated HRP-labelled
streptavidin (Dako) and then washed with PBS. The slides were then
incubated with diaminobenzidine (DAB, Sigma Chemicals) as the
chromogen, followed by counterstaining with diluted Harris
hematoxylin (Sigma Chemicals). After staining, five high-power
fields (×400) were randomly selected in each slide, and the average
proportion of positive cells in each field were counted using the
true color multi-functional cell image analysis management system
(Image-Pro Plus, Media Cybernetics, Bethesda, MD, USA). To rule out
any non-specific staining, PBS was used to replace the primary
antibody as a negative control.
Statistical analysis
All data are the means of three determinations and
data were analyzed using the SPSS package for Windows (Version
11.5). Statistical analysis of the data was performed with
Student’s t-test and ANOVA. Differences with P<0.05 were
considered statistically significant.
Results
EEHDW inhibits tumor growth in colorectal
cancer (CRC) xenograft mice
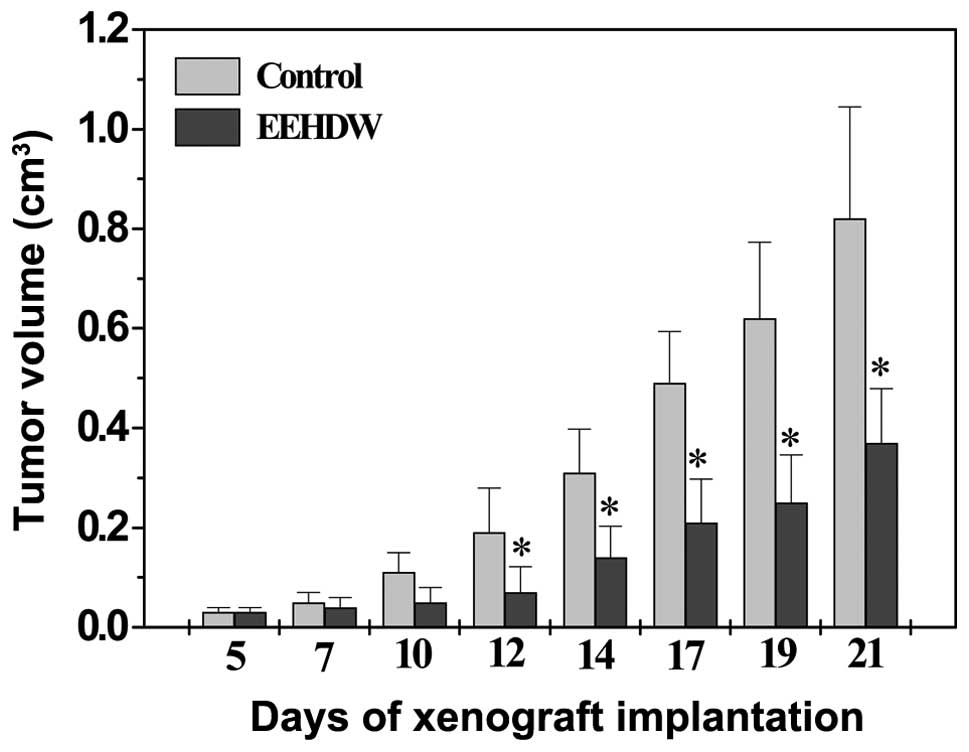
The inhibitory effect of EEHDW on cancer growth was
evaluated by measuring tumor volume. We found that administration
with EEHDW significantly inhibited tumor growth throughout the
study, as compared with the control group (Fig. 1). At the end of experiments, tumor
volume per mouse in EEHDW treatment group was 0.37±0.11
cm3, whereas that in control group was 0.82±0.23
cm3 (Fig. 1,
P<0.01), demonstrating the therapeutic efficacy of EEHDW against
CRC in vivo.
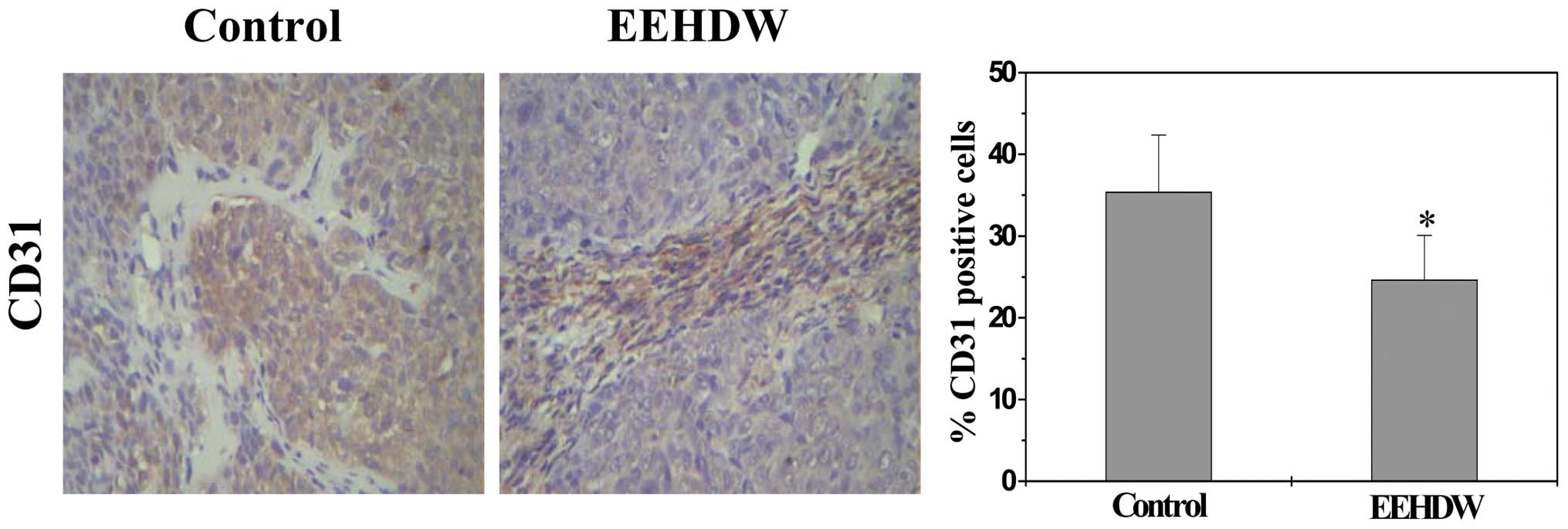
EEHDW displays anti-angiogenic activity
in CRC xenograft mice
We performed immunohistochemical staining (IHS) for
endothelial cell-specific marker CD31, to determine the effect of
EEHDW on intratumoral microvessel density (MVD) that is considered
as an indicator of tumor angiogenesis and is associated with cancer
growth and progression (17). As
shown in Fig. 2, the percentage of
CD31-positive cells in tumors from EEHDW-treated or control mice
was 24.67±5.43 or 35.33±2.88%, respectively (P<0.01), suggesting
that inhibition of tumor angiogenesis by EEHDW could have
contributed to the inhibition of colorectal tumor growth.
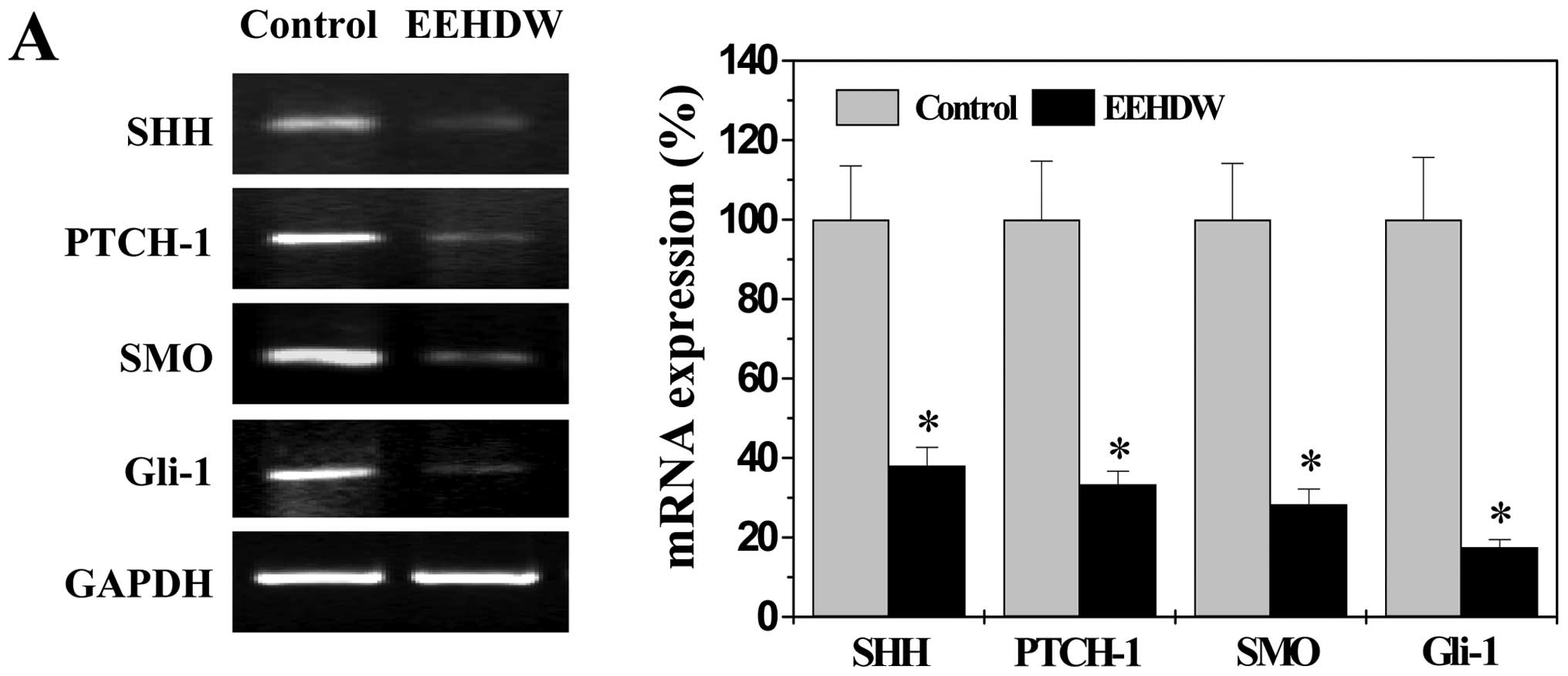
EEHDW suppresses SHH pathway in CRC
xenograft mice
To determine the effect of EEHDW on SHH pathway, we
evaluated mRNA and protein expression levels of the key mediators
of this pathway in CRC xenograft tumors using RT-PCR and IHS assay.
Data from RT-PCR showed that EEHDW profoundly reduced the mRNA
expression of SHH, Ptch-1, Smo and Gli-1 in CRC tumor tissues
(Fig. 3A, P<0.01). In a
consistent manner, the protein expression of these factors was
significantly downregulated by EEHDW treatment. The percentage of
cells in the CRC xenograft tumors expressing SHH, Ptch-1, Smo or
Gli-1 in the EEHDW-treated mice was 25.67±7.81, 18.00±3.96,
26.00±4.98 or 25.17±5.12%, whereas that in control group was
34.83±5.31, 35.33±5.24, 37.33±5.99 or 35.83±7.30%, respectively
(Fig. 3B, P<0.01).
Collectively, these data suggest that EEHDW inhibits the activation
of SHH signaling in colorectal cancer.
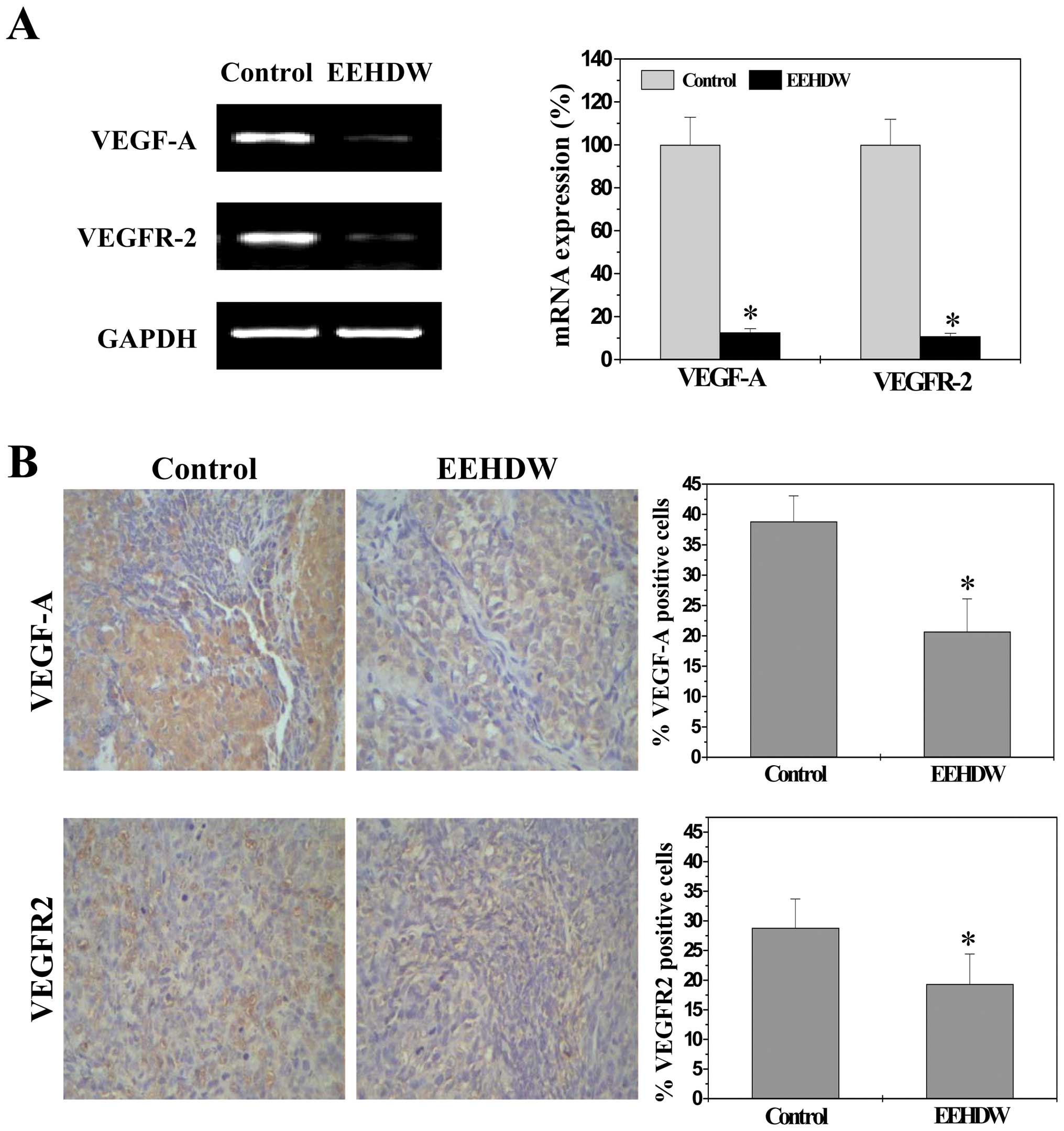
EEHDW decreases VEGF and VEGFR2
expression in tumors
To further investigate the mechanism of
anti-angiogenic activity of EEHDW, we examined the expression of
VEGF-A, a critical target gene of SHH pathway, as well as the
expression of its specific receptor VEGFR2. As shown in Fig. 4, EEHDW treatment obviously
decreased the mRNA and protein expression levels of VEGF-A and
VEGFR2 in CRC tumor tissues (P<0.01).
Discussion
Angiogenesis has become an attractive target for
anticancer chemotherapy due to its critical role for the
progression and metastasis of solid tumors. A number of
anti-angiogenic agents are currently in preclinical development,
with some of them now entering the clinic trials. However, due to
the intrinsic cytotoxicity against non-tumor associated endothelial
cells, long-term use of angiogenesis inhibitors usually causes
cardiovascular complications such as impaired wound healing,
bleeding, hypertension and thrombosis (35–37).
Therefore, discovering novel antitumor agents with minimal adverse
effects is needed. Natural products, including traditional Chinese
medicine (TCM), have relatively fewer side-effects and have been
used clinically to treat various kinds of diseases including cancer
for thousands of years. Hedyotis diffusa Willd (HDW), a well
known traditional Chinese folk-medicine, has long been used as a
major component in several TCM formulas for the clinical treatment
of various kinds of cancer including colorectal cancer (CRC)
(3). Recently, we reported that
HDW can inhibit cancer growth in vivo and in vitro
via suppression of STAT3 pathway (3,34)
and displays anti-angiogenic activity in vitro(25). However, the mode of action for its
antitumor effect is still largely unknown.
In the present study we confirmed that the ethanol
extract of Hedyotis diffusa Willd (EEHDW) could inhibit
cancer growth in vivo, consistent with our previous study
(34). More importantly, by using
immunohistochemical (IHC) staining for the endothelial
cell-specific marker CD31 we found that EEHDW significantly reduced
the intratumoral microvessel density (MVD), demonstrating its in
vivo anti-angiogenic activity. Tumor angiogenesis is tightly
regulated by Sonic hedgehog (SHH) pathway. Activation of HH
signaling upregulates the expression of VEGF-A which is the
strongest angiogenesis stimulator (20,21,29,30,38).
VEGF-A exerts its pro-angiogenic function via binding to its
specific receptors including VEGFR2 which is located on vascular
endothelial cells (EC) (25–29),
leading to series of angiogenic processes (29,30).
In this study we found that EEHDW treatment inhibited the
activation of SHH pathway in tumor tissues since EEHDW
significantly suppressed the mRNA and protein expression of several
key mediators of SHH signaling, including SHH, PTCH-1, SMO and
GLi-1. Consistently, administration of EEHDW profoundly reduced the
expression of VEGF-A and VEGFR2 in vivo, at both
transcriptional and translational levels.
In conclusion, here for the first time we
demonstrate that Hedyotis diffusa Willd inhibits colorectal
cancer growth in vivo via inhibition of SHH-mediated tumor
angiogenesis, which may in part explain its anticancer
activity.
Abbreviations:
|
EEHDW
|
ethanol extract of Hedyotis
diffusa Willd;
|
|
CRC
|
colorectal cancer;
|
|
IHS
|
immunohistochemical staining;
|
|
VEGF-A
|
vascular endothelial growth
factor-A;
|
|
MVD
|
microvessel density; SHH, sonic
hedgehog signal pathway
|
Acknowledgements
This study was sponsored by the
Natural Science Foundation of Fujian Province of China
(2010J01195), the Research Foundation of Education Bureau of Fujian
Province of China (JA10162) and the National Natural Science
Foundation of China (81073097).
References
|
1.
|
Jemal A, Bray F, Center MM, Ferlay J, Ward
E and Forman D: Global cancer statistics. CA Cancer J Clin.
61:69–90. 2011. View Article : Google Scholar
|
|
2.
|
Gustin DM and Brenner DE: Chemoprevention
of colon cancer: current status and future prospects. Cancer Metast
Rev. 21:323–348. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
3.
|
Lin J, Chen Y, Wei L, Chen X, Xu W, Hong
Z, Sferra TJ and Peng J: Hedyotis diffusa Willd extract
induces apoptosis via activation of the mitochondrion-dependent
pathway in human colon carcinoma cells. Int J Oncol. 37:1331–1338.
2010.
|
|
4.
|
Gordaliza M: Natural products as leads to
anticancer drugs. Clin Transl Oncol. 9:767–776. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
5.
|
Ji HF, Li XJ and Zhang HY: Natural
products and drug discovery. Can thousands of years of ancient
medical knowledge lead us to new and powerful drug combinations in
the fight against cancer and dementia? EMBO Rep. 10:194–200.
2009.PubMed/NCBI
|
|
6.
|
Folkman J: Tumor angiogenesis: therapeutic
implications. N Engl J Med. 285:1182–1186. 1971. View Article : Google Scholar : PubMed/NCBI
|
|
7.
|
Folkman J: Angiogenesis. Annu Rev Med.
57:1–18. 2006. View Article : Google Scholar
|
|
8.
|
Cook KM and Figg WD: Angiogenesis
inhibitors: current strategies and future prospects. CA Cancer J
Clin. 60:222–243. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
9.
|
Ingham PW, Nakano Y and Seger C:
Mechanisms and functions of Hedgehog signalling across the metazoa.
Nat Rev Genet. 12:393–406. 2011. View
Article : Google Scholar : PubMed/NCBI
|
|
10.
|
Zhang S, Wang Y, Mao JH, Hsieh D, Kim IJ,
Hu LM, Xu Z, Long H, Jablons DM and You L: Inhibition of CK2α
down-regulates Hedgehog/Gli signaling leading to a reduction of a
stem-like side population in human lung cancer cells. PLoS One.
7:e389962012.
|
|
11.
|
Theunissen JW and de Sauvage FJ: Paracrine
Hedgehog signaling in cancer. Cancer Res. 69:6007–6010. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
12.
|
Mazumdar T, DeVecchio J, Shi T, Jones J,
Agyeman A and Houghton JA: Hedgehog signaling drives cellular
survival in human colon carcinoma cells. Cancer Res. 71:1092–1102.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
13.
|
Yoshikawa K, Shimada M, Miyamoto H,
Higashijima J, Miyatani T, Nishioka M, Kurita N, Iwata T and Uehara
H: Sonic hedgehog relates to colorectal carcinogenesis. J
Gastroenterol. 44:1113–1117. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
14.
|
Varnat F, Duquet A, Malerba M, Zbinden M,
Mas C, Gervaz P and Ruiz i Altaba A: Human colon cancer epithelial
cells harbour active HEDGEHOG-GLI signalling that is essential for
tumour growth, recurrence, metastasis and stem cell survival and
expansion. EMBO Mol Med. 1:338–351. 2009. View Article : Google Scholar
|
|
15.
|
Das S, Tucker JA, Khullar S, Samant RS and
Shevde LA: Hedgehog signaling in tumor cells facilitates
osteoblast-enhanced osteolytic metastases. PLoS One. 7:e343742012.
View Article : Google Scholar : PubMed/NCBI
|
|
16.
|
Sahebjam S, Siu LL and Razak AA: The
utility of Hedgehog signaling pathway inhibition for cancer.
Oncologist. 17:1090–1099. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
17.
|
Wei L, Lin J, Xu W, Cai Q, Shen A, Hong Z
and Peng J: Scutellaria barbata D. Don inhibits tumor
angiogenesis via suppression of Hedgehog pathway in a mouse model
of colorectal cancer. Int J Mol Sci. 13:9419–9430. 2012. View Article : Google Scholar
|
|
18.
|
Lum L and Beachy PA: The Hedgehog response
network: sensors, switches, and routers. Science. 304:1755–1759.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
19.
|
Varjosalo M and Taipale J: Hedgehog:
functions and mechanisms. Genes Dev. 22:2454–2472. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
20.
|
Pola R, Ling LE, Silver M, Corbley MJ,
Kearney M, Pepinsky RB, Shapiro R, Taylor FR, Baker DP and Asahara
T: The morphogen Sonic hedgehog is an indirect angiogenic agent
upregulating two families of angiogenic growth factors. Nat Med.
7:706–711. 2001. View
Article : Google Scholar : PubMed/NCBI
|
|
21.
|
Claret S, Sanial M and Plessis A: Evidence
for a novel feedback loop in the Hedgehog pathway involving
smoothened and fused. Curr Biol. 17:1326–1333. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
22.
|
Kerbel RS: Tumor angiogenesis. N Engl J
Med. 358:2039–2049. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
23.
|
Prasad S, Yadav VR, Sung B, Reuter S,
Kannappan R, Deorukhkar A, Diagaradjane P, Wei C,
Baladandayuthapani V, Krishnan S, Guha S and Aggarwal BB: Ursolic
acid inhibits growth and metastasis of human colorectal cancer in
an orthotopic nude mouse model by targeting multiple cell signaling
pathways: chemosensitization with capecitabine. Clin Cancer Res.
18:4942–4953. 2012. View Article : Google Scholar
|
|
24.
|
Dai J, Wan S, Zhou F, Myers RE, Guo X, Li
B, Fu X, Palazzo JP, Dou K, Yang H and Xing J: Genetic polymorphism
in a VEGF-independent angiogenesis gene ANGPT1 and overall survival
of colorectal cancer patients after surgical resection. PLoS One.
7:e347582012. View Article : Google Scholar
|
|
25.
|
Lin J, Wei L, Xu W, Hong Z, Liu X and Peng
J: Effect of Hedyotis diffusa Willd extract on tumor
angiogenesis. Mol Med Rep. 4:1283–1288. 2011.
|
|
26.
|
Ferrara N, Gerber HP and LeCouter J: The
biology of VEGF and its receptors. Nat Med. 9:669–676. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
27.
|
Shigami SI, Arii S, Furutani M, Niwano M,
Harada T, Mizumoto M, Mori A, Onodera H and Imamura M: Predictive
value of vascular endothelial growth factor (VEGF) in metastasis
and prognosis of human colorectal cancer. Br J Cancer.
78:1379–1384. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
28.
|
Gille H, Kowalski J, Li B, LeCouter J,
Moffat B, Zioncheck TF, Pelletier N and Ferrara N: Analysis of
biological effects and signaling properties of Flt-1 (VEGFR-1) and
KDR (VEGFR-2). A reassessment using novel receptor-specific
vascular endothelial growth factor mutants. J Biol Chem.
276:3222–3230. 2001. View Article : Google Scholar
|
|
29.
|
Ferrara N: Role of vascular endothelial
growth factor in physiologic and pathologic angiogenesis:
therapeutic implications. Semin Oncol. 29:10–14. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
30.
|
Risau W: Mechanisms of angiogenesis.
Nature. 386:671–674. 1997. View
Article : Google Scholar : PubMed/NCBI
|
|
31.
|
Ren F, Liu G, Zhang L and Niu G: Studies
on chemical constituents of Hedyotis diffusa Willd. Chin
Pharm J. 40:502–504. 2005.
|
|
32.
|
Ahmad R, Shaari K, Lajis NH, Hamzah AS,
Ismail NH and Kitajima M: Anthraquinones from Hedyotis
capitellata. Phytochemistry. 66:1141–1147. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
33.
|
Li C, Xue X, Zhou D, Zhang F, Xu Q, Ren L
and Liang X: Analysis of iridoid glucosides in Hedyotis
diffusa by high-performance liquid chromatography/electrospray
ionization tandem mass spectrometry. J Pharm Biomed Anal.
48:205–211. 2008.
|
|
34.
|
Cai Q, Lin J, Wei L, Zhang L, Wang L, Zhan
Y, Zeng J, Xu W, Shen A, Hong Z and Peng J: Hedyotis diffusa
Willd inhibits colorectal cancer growth in vivo via inhibition of
STAT3 signaling pathway. Int J Mol Sci. 13:6117–6128. 2012.
View Article : Google Scholar
|
|
35.
|
Chen HX and Cleck JN: Adverse effects of
anticancer agents that target the VEGF pathway. Nat Rev Clin Oncol.
6:465–477. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
36.
|
Zangari M, Fink LM, Elice F, Zhan F,
Adcock DM and Tricot GJ: Thrombotic events in patients with cancer
receiving antiangiogenesis agents. J Clin Oncol. 27:4865–4873.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
37.
|
Higa GM and Abraham J: Biological
mechanisms of bevacizumab-associated adverse events. Expert Rev
Anticancer Ther. 9:999–1007. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
38.
|
Jain RK: Tumor angiogenesis and
accessibility: role of vascular endothelial growth factor. Semin
Oncol. 29:3–9. 2002. View Article : Google Scholar : PubMed/NCBI
|