Introduction
The delta-like homolog 1 (DLK1) and gene trap
locus 2 (GTL2, also known as MEG3) gene cluster is
located on human chromosome 14. The importance of the
DLK1-GTL2 locus in skeletal muscle development is supported
by the fact that a single-nucleotide polymorphism at this locus
deregulates its expression and leads to DLK1-mediated skeletal
muscle hypertrophy, as seen in callipyge sheep (1).
Rhabdomyosarcoma (RMS), a family of cancers related
to the skeletal muscle lineage, is the most common soft-tissue
sarcoma in children. From a histological and clinical perspective,
there are two major histological subtypes of RMS: embryonal
rhabdomyosarcoma (ERMS) and alveolar rhabdomyosarcoma (ARMS)
(2). Clinical evidence indicates
that ARMS is more aggressive, shows more enhanced metastatic
potential, and has a significantly worse outcome than ERMS. It has
been postulated that there are different cellular origins for these
tumors (3) but the molecular
mechanisms responsible for the development of RMS are not well
understood.
Evidence has accumulated that imprinted genes play a
role in RMS pathogenesis (4,5). In
the human genome, there are ∼80 genes that are imprinted and
expressed only from the maternally or paternally derived
chromosomes. This mechanism regulates the appropriate dosage and
expression level of developmentally important genes in mammalian
cells (6). The expression of
imprinted genes is regulated by the imposition of epigenetic marks
(DNA methylation) within differentially methylated regions (DMRs),
which are regulatory CpG-rich regions in the gene locus (7–9).
Most imprinted genes are methylated on maternally derived
chromosomes and only a few (e.g., IGF2 and DLK1) are
methylated within DMRs on paternally derived chromosomes.
According to the parent-offspring conflict theory,
while paternally expressed imprinted genes enhance embryo growth
and size of the offspring, maternally expressed genes inhibit cell
proliferation and negatively affect cell size (7–9).
Based on this theory, during pregnancy, the paternal allele
promotes increased body size, including muscle mass of the
developing fetus, through expression of paternally imprinted genes.
By contrast, the maternal allele conserves resources by epigenetic
modulation of genes bearing maternal imprinting marks.
Comparison of the IGF2-H19 and
DLK1-GTL2 tandem loci reveals significant similarities. The
molecular structure of these imprinted loci is very similar. The
DMRs for these tandem genes are both located between the two genes
within each locus (between IGF2 and H19 and between
DLK1 and GTL2, respectively) (10,11),
and these elements are thus called intergenic DMRs (IG-DMR).
IGF2 encodes insulin-like growth factor-2 (IGF-2) and
DLK1 encodes transmembrane delta-like 1 protein, which
contains six epidermal growth factor (EGF) repeat motifs and is
involved in cell differentiation in a juxtacrine/paracrine manner.
Both DLK1 and IGF2 are transcribed from paternal
chromosomes and are involved in skeletal muscle development as
stimulators of skeletal muscle growth (12,13).
By contrast, the two other genes in these tandem loci -H19 and
GTL2- transcribe non-coding RNAs (ncRNAs) that are precursors of
miRNAs that negatively regulate cell proliferation.
Loss of imprinting (LOI) of the IG-DMR at the
IGF2-H19 locus located at the 11p15.5 chromosome region is
observed both in ERMS and ARMS. Furthermore, this LOI have been
reported in both sporadic tumors and in RMS originating in
Beckwith-Wiedemann syndrome patients (5). The result of this LOI is
overexpression of IGF2, which acts as an autocrine growth factor in
this tumor. Since the IGF2-H19 locus is structurally similar
to DLK1-GTL2, we investigated whether LOI of the DMR at the
DLK1-GTL2 locus also occurs in RMS. We observed that, while
ERMS cells consistently showed LOI of the DMR at the
DLK1-GTL2 locus, ARMS cells displayed erasure of imprinting
(EOI) in this regulatory region. Despite EOI of the IG-DMR within
the DLK1-GTL2 locus, ARMS cells highly express DLK1
mRNA. Therefore, regulation of DLK1 seems to be more
complicated in RMS cells than initially thought. These findings
have biological implications and may also have diagnostic
utility.
Materials and methods
Patient samples
For 35 of the samples analyzed, frozen tumor samples
were provided by the Pediatric Division of the Cooperative Human
Tissue Network without patient identifiers. The only information
provided was histologic diagnosis and fraction of tumor cells in a
frozen section from the block from which the sample was taken. In
the samples, the fraction of tumor cells varied from 65 to 90%. In
17 of the samples, the histologic subtype was embryonal
rhabdomyosarcoma and in 18 of the samples the histologic subtype
was alveolar rhabdomyosarcoma. In the latter 18 cases, the presence
of the characteristic gene fusions was ascertained by RT-PCR
revealing 14 cases with a PAX3-FOXO1 fusion and 4 cases with a
PAX7-FOXO1 fusion (14).
Combined bisulfite-restriction analysis
(COBRA) and bisulfite-sequencing
DNA was isolated from frozen samples using the
QIAamp system (Qiagen Inc., Valencia, CA). The DNA methylation
status of IG-DMRs at the DLK1-GLT2, IGF2-H19,
LIT1, SNRPN and PEG1 loci were examined by
COBRA analysis. In brief, 500 ng of genomic DNA was used for
bisulfite modification using the EpiTect Bisulfite Kit (Qiagen
Inc.) or the EZ DNA Methylation Kit (Zymo Research, Irvine, CA)
according to the manufacturer’s instructions. IG-DMR regions were
amplified by nested PCR (except for LIT1, for which only one
round of PCR was performed) using bisulfite-treated gDNA as
template and specific primers (Table
I). For DLK1-GLT2, IGF2-H19 and PEG1, both
first-and second-round PCR were performed with 2 cycles of 2 min at
95°C, 1 min at 55°C and 1 min at 72°C; 35 cycles of 30 sec at 95°C,
1 min at 55°C and 1 min at 72°C; and 1 cycle of 10 min at 72°C. For
LIT1 and SNRPC, PCR was performed as previously
described (16,17). The COBRA assay was performed by
incubation of the final PCR products with BstUI restriction enzyme
for 2 h and subsequent agarose gel electrophoresis to assess the
cutting pattern.
 | Table I.Sequences of primers employed for
bisulfite sequencing and methylation-specific PCR. |
Table I.
Sequences of primers employed for
bisulfite sequencing and methylation-specific PCR.
| Region | | Sequence | (Refs.) |
|---|
| Bisulfite
sequencing | | | |
|
DLK1-GTL2 | Outer | F: TGG GAA TTG GGG
TAT TGT TTA T
R: AAA CAA TTT AAC AAC AAC TTT CCT C | (19) |
| Inner | F: GTT AAG AGT TTG
TGG ATT TGT GAG AAA TG
R: CTA AAA ATC ACC AAA ACC CAT AAA ATC AC | ✓ |
|
IGF2-H19 | Outer | F: AGG TGT TTT AGT
TTT ATG GAT GAT GG
R: TCC TAT AAA TAT CCT ATT CCC AAA TAA CC | (15) |
| Inner | F: TGT ATA GTA TAT
GGG TAT TTT TGG AGG TTT
R: H19-OR was used | (15) |
| LIT1 | (One PCR) | F: GTG TTA IGG IGG
TGG AGA TTT TGT
R: AAC TIA AAACACIAACCAATTCTCTA | (16) |
| PEG1 | Outer | F: TTG TTG TTG GTT
AGT TTT GTA TGG TT
R: AAA AAT AAC ACC CCC TCC TCA AAT | (15) |
| Inner | F: PEG1-OF was
used
R: CCC AAA AAC AAC CCC AAC TC | (15) |
| SNRPN | Outer | F: GTG TTG TGG GGT
TTT AGG GGT TTA G
R: CTC CCC AAA CTA TCT CTT AAA AAA AAC C | (17) |
| Inner | F: AGG GAG TTG GGA
TTT TTG TAT TG
R: SNRPN-OR was used | (17) |
|
Methylation-specific PCR | | | |
|
DLK1-GTL2 | Methylated | F: TAT TTT AAG ATT
GTT AGT TTT TTC GC
R: AAA ACC CAA CCC AAT AAA CG | ✓ |
| Unmethylated | F: GTTT TA TTT TAA
GAT TGT TAG TTT TTT
R: CAA AAC CCA ACC CAA TAA ACA | ✓ |
For sequencing, amplicons from the above-described
PCR reactions were eluted after agarose gel electrophoresis using
the QIAquick Gel Extraction Kit (Qiagen Inc.). Eluted amplicons
were ligated into the pCR®2.1-TOPO® vector
and transformed into TOP10 bacteria using a TOPO TA Cloning Kit
(Invitrogen, Carlsbad, CA). The plasmids were prepared using a
QIAprep Spin Miniprep Kit (Qiagen Inc.) and sequenced with M13
forward and reverse primers. The methylation pattern in DMRs was
analyzed using CpG viewer software (18).
Methylation-specific PCR of the IG-DMR at
the DLK1-GTL2 locus
Methylation-specific PCR was performed on the
bisulfite-treated genomic DNA with two different sets of primers
recognizing either methylated or unmethylated alleles (Table I). PCR was performed at 2 cycles of
2 min at 95°C, 1 min at 55°C and 1 min at 72°C; 35 cycles of 30 sec
at 95°C, 1 min at 55°C and 1 min at 72°C; and 1 cycle of 10 min at
72°C.
Quantitative RT-PCR (qRT-PCR)
Total RNA from RMS patient samples was isolated
using RNA STAT-60 (Tel-Test, Friendswood, TX) in accordance with
manufacturer’s instructions. RNA was screened for expression of
DLK1 and GTL2 using premade qRT-PCR assays from Applied Biosystems
(Foster City, CA). All qRT-PCR assays were run on 384-well plates
in the ABI Prism 7900 Sequence Detection System. The control 18S
RNA was assayed in separate wells of the same run using a premade
qRT-PCR assay (Applied Biosystems).
Statistical analysis
Statistical analysis of the data was done using the
Fisher’s exact test with p<0.05 as the criterion for
significance.
Results
LOI of the DMR at the DLK1-GTL2 locus in
ARMS and ERMS samples
Since the DLK1-GTL2 locus exhibits structural
and molecular similarity to the IGF2-H19 locus, we
investigated the imprinting status of the IG-DMR at the
DLK1-GTL2 locus, which is located between the two genes and
controls coordinated expression of both genes at this locus
(19). To address this question,
we performed the COBRA assay, in which PCR-amplified
bisulfite-treated DNA is cut with an appropriate restriction enzyme
(BstUI) to determine whether the fragment has retained or
lost the corresponding CpG-containing restriction enzyme
recognition site. The digested DNA was subsequently separated and
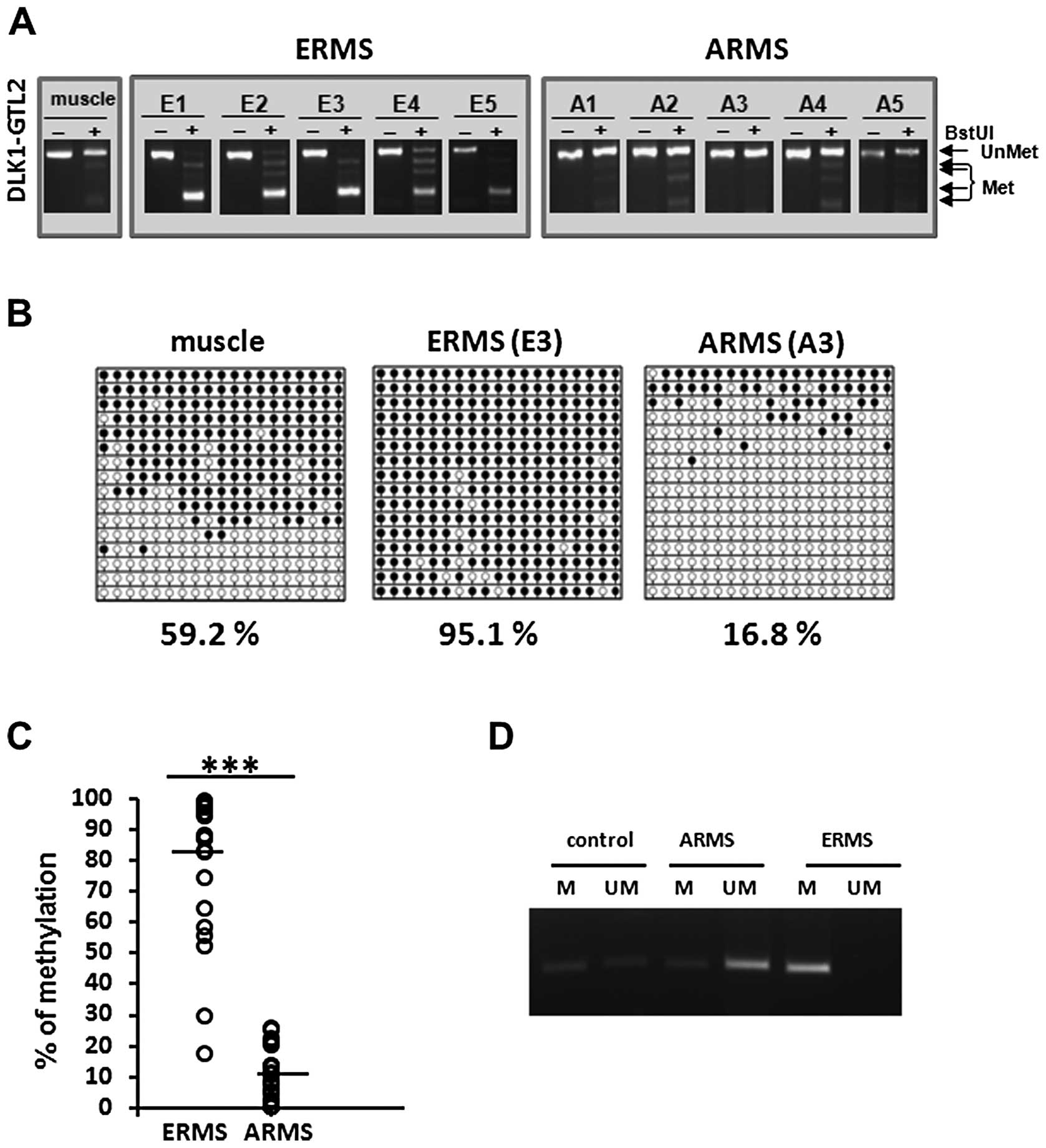
visualized by agarose gel electrophoresis. As shown in Fig. 1A, the BstU1 site was cut in
all five ERMS samples and thus this region was hypermethylated in
these cases. As there is often no evidence of a residual uncut of
hypomethylated allele, this finding is indicative of LOI in the
ERMS cases. By contrast, all five analyzed ARMS patient samples
have lost the BstU1 recognition site and thus exhibited
hypomethylation of the IG-DMR at the DLK1-GTL2 locus. On the
other hand since there is often no evidence of a residual cut of
hypermethylated allele, this finding is consistent with elimination
of imprinting (EOI) in these ARMS cases.
To further confirm these imprinting changes at the
DLK1-GTL2 locus in RMS, we employed bisulfite modification
of DNA followed by sequencing of selected ERMS (patient E3) and
ARMS (patient A3) samples. The PCR-amplified bisulfite-treated DNA
fragments were subcloned and multiple subclones were individually
sequenced. These sequencing results further supported our COBRA
results, as all the copies of the IG-DMR at the DLK1-GTL2
locus were hypermethylated in the ERMS case (consistent with LOI),
and were hypomethylated in the ARMS case (consistent with EOI)
(Fig. 1B). As expected, DNA
methylation of IG-DMRs in human skeletal muscle was ∼60% (19).
After these initial studies, to further confirm our
observation we analyzed an additional 17 ERMS and 18 ARMS patient
samples by the COBRA assay. Combined densitometric analysis of all
analyzed patient samples showed a highly relevant difference in
methylation pattern between ARMS and ERMS. The average methylation
of ARMS samples was ∼10%, whereas for ERMS samples, we observed 80%
methylation (Fig. 1C). Therefore,
these results demonstrate that the methylation pattern of the
IG-DMR significantly differs between the two subtypes of RMS.
Since distinguishing between ARMS and ERMS currently
requires histopathology analysis of patient samples, we developed
methylation-specific PCR primers that enable quick and specific
analysis of the methylation status of the DLK1-GTL2 locus
and thus can be potentially employed in RMS diagnostics (Fig. 1D).
The methylation pattern of the DMRs at
the IGF2-H19, PEG1, LIT1 and SNRPN loci do not show differences
between ARMS and ERMS samples
Next, by applying additional COBRA assays on the
same DNA samples derived from ERMS and ARMS tumors, we performed an
analysis of the methylation status of the DMRs at other imprinted
loci, such as IGF2-H19, PEG1, LIT1 and
SNRPN. As expected, we confirmed LOI of the DMR within
IGF2-H19 in all of the samples and at the same time some
degree of hypomethylation LOI of the LIT1 DMR in most of the
samples. At the same time, we did not observe any significant
changes in the methylation state of PEG1 and SNRPN
loci (Fig. 2A).
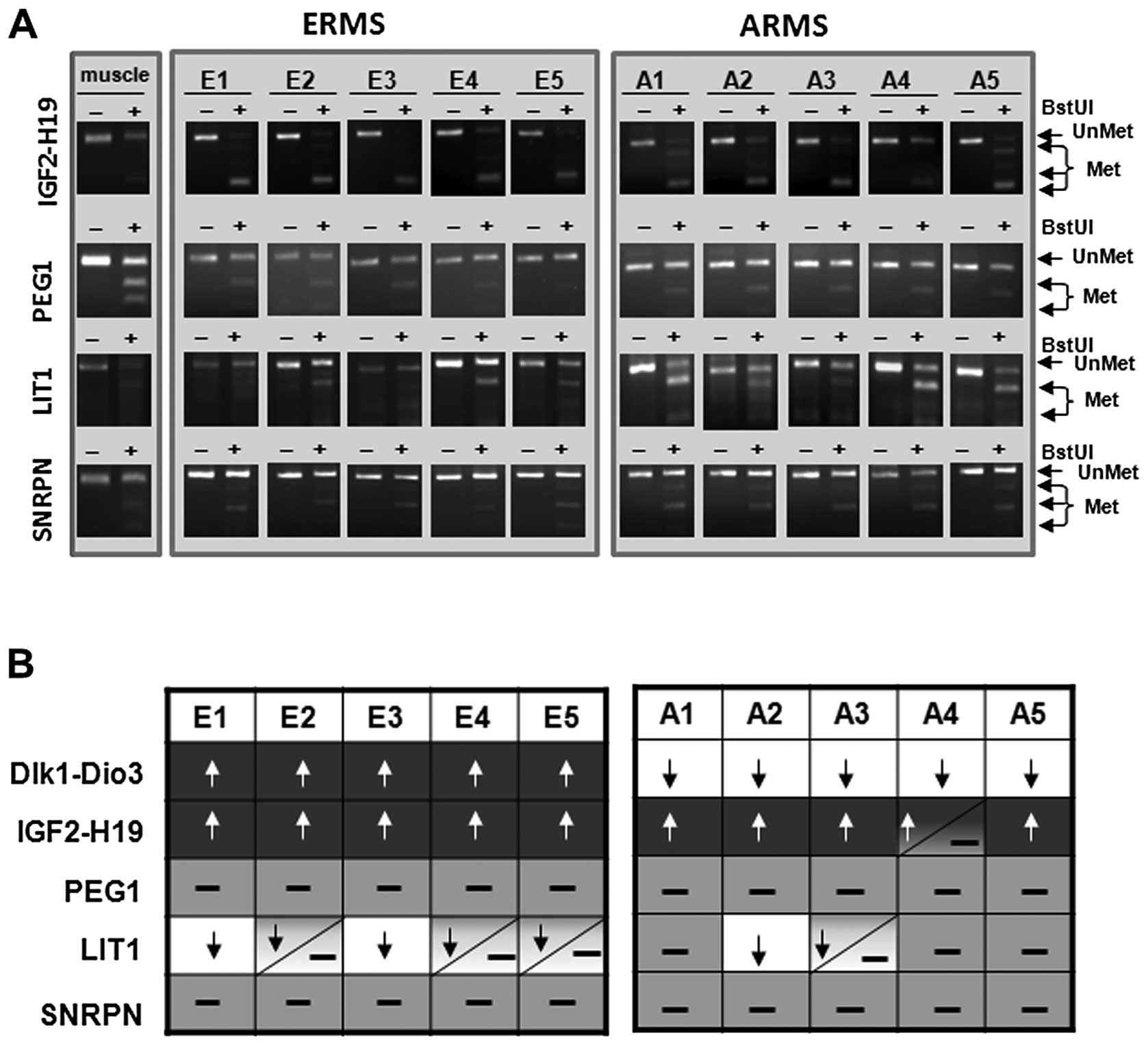 | Figure 2.Methylation pattern of the DMRs at
the IGF2-H19, PEG1, LIT1 and SNRPN loci
in ERMS and ARMS patient samples. (A) COBRA assay for DMRs at the
IGF2-H19, PEG1, LIT1 and SNRPN loci in
the presence of BstUI restriction enzyme. The unmethylated DNA
(UnMet) was not cleaved, in contrast to methylated DNA (Met),
because of a sequence change in the site recognized by the
restriction enzyme after the bisulfite reaction. (B) Summary of
methylation patterns of DMRs at the DLK1-GTL2,
IGF2-H19, PEG1, LIT1 and SNRPN loci in
DNA samples from normal skeletal muscles and patient DNA samples
(ERMS and ARMS). White color with black arrow indicates
hypomethylation (EOI), light grey indicates normal status (50%) and
dark color with white arrow indicates hypermethylation (LOI). |
Fig. 2B summarizes
the methylation pattern of DMRs at all loci tested by us in ERMS
and ARMS tumor samples and demonstrates that, for all of the DMRs
evaluated, imprinting at the DLK1-GTL2 locus differs between
ERMS and ARMS and may be of some diagnostic importance for example
as an addition to fusion assay.
LOI of the DMR at the DLK1-GTL2 locus
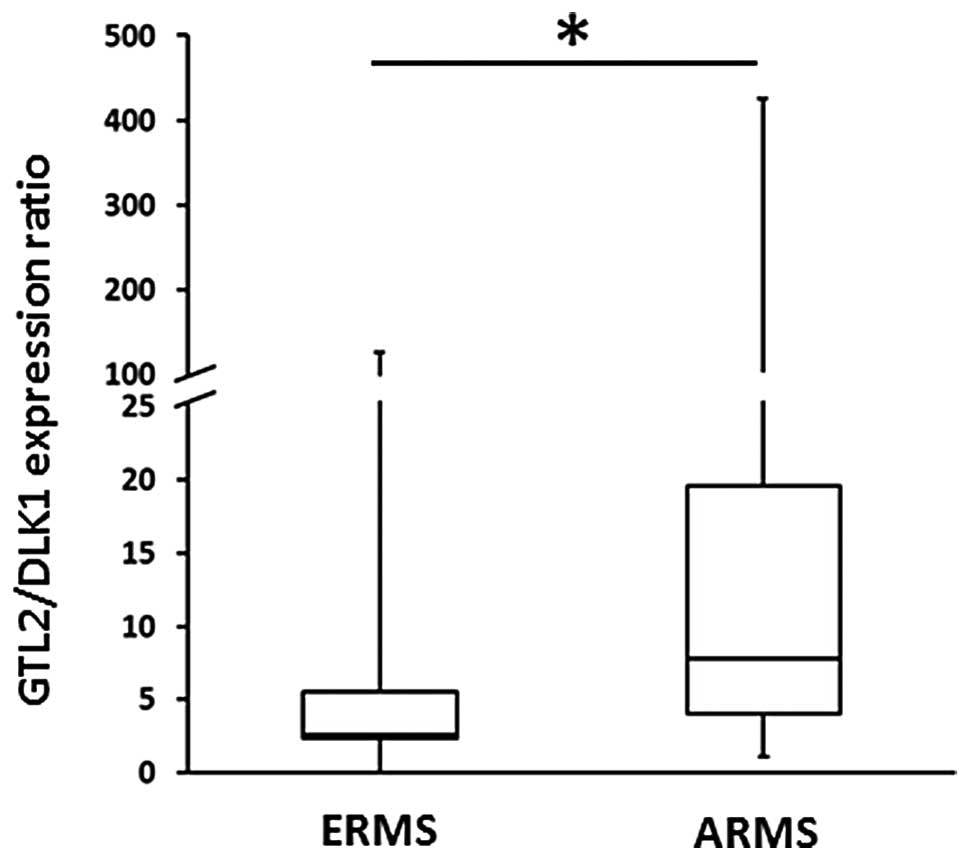
correlates with changes in GTL2/DLK1 mRNA ratio
Finally, to see whether changes in imprinting of
particular DMRs are translated into changes in RNA transcription,
we performed qRT-PCR analysis of the most important imprinted genes
in RNA extracts isolated from ERMS and ARMS samples (Fig. 3). To address this question we
compared the GTL2/DLK1 ratio in a large panel of ERMS and ARMS
tumors. The Fisher’s exact test analysis revealed a higher
GTL2/DLK1 ratio in ARMS patients as compared to ERMS; this finding
is consistent with LOI at the DLK1-GTL2 locus.
Discussion
The tandem DLK1-GTL2 gene locus is part of a
large genomic region located on chromosome 14 (14q32) known as
DLK1-DIO3 (20), which
contains three paternally imprinted and expressed genes,
DLK1, RTL1 and DIO3, as well as three
maternally expressed genes, GTL2 (MEG3), MEG8
(RIAN), and the antisense gene RTL1. Moreover, in
addition to two long non-coding intergenic RNAs (GTL2 and
MEG8), this region contains one of the largest miRNA
clusters in the human genome, encoding a total of 54 miRNAs, which
play an important role in tissue homeostasis and, when aberrantly
expressed, are involved in tumorigenesis (21). We focused on the DLK1-GTL2
locus in the 3’ end of this region, which is regulated by
imprinting of the IG-DMR. The proper expression of genes in this
locus from the paternally and maternally inherited genes
DLK1 and GTL2, respectively, is crucial for normal
development (19,22) and pluripotency of stem cells
(23).
Disordered imprinting has been implicated in the
pathogenesis of pediatric cancers. In particular, epigenetic
alterations in imprinting within the IGF2-H19 locus have
been observed in pediatric cancers, such as RMS and nephroblastoma
(Wilm’s tumor), which develop spontaneously or as a part of
Beckwith-Wiedemann syndrome (4).
Based on these observations, we became interested in potential
alterations to imprinting at the DLK1-GTL2 locus, which is
structurally similar to the IGF2-H19 locus. Such alterations
of imprinting are important because LOI of the IG-DMR at the
DLK1-GTL2 locus may result in overexpression of DLK1,
which stimulates development of skeletal muscle tissue (12), and downregulation of GTL2,
which, as recently postulated, may have an anti-oncogenic function
(21,24). In the case of EOI of the IG-DMR at
the DLK1-GTL2 locus, we would expect to see a reversed
pattern of expression for these genes, downregulation of
DLK1 and upregulation of GTL2.
In our studies, performed on DNA samples from 23
ERMS and 23 ARMS tumors, we observed that the IG-DMR at the
DLK1-GTL2 locus is hypermethylated (LOI) in ERMS and
hypomethylated (EOI) in ARMS cells. Therefore, in contrast to the
LOI that is consistently observed within the IGF2-H19 locus
in both ERMS and ARMS samples, methylation of the IG-DMR at the
DLK1-GTL2 locus is RMS-subtype specific. This different
pattern of methylation could have some diagnostic value. By
employing a specific pair of primers that recognize methylated DNA
sequences, we have demonstrated that it is possible to quickly
phenotype both subtypes of RMS by PCR.
To address if these changes in imprinting at
DLK1-GTL2 locus correspond to changes in mRNA expression we
analyzed by employing RQ-PCR the GTL2/DLK1 ratio in 10
fusion-negative and 16 PAX3-FOXO1 fusion-positive ARMS samples. Our
mRNA expression data correlated with imprinting status of the
DLK1-GTL2 locus and demonstrated a significantly higher
GTL2/DLK1 ratio in ARMS tumor samples compared to ERMS tumors.
In conclusion our data indicate that DLK1/GTL2 locus
is differently regulated by imprinting in ARMS vs. ERMS cells. We
developed an easy PCR-based assay to phenotype ERMS and ARMS, based
on the methylation status of the IG-DMR at the DLK1-GTL2
locus. However, we are aware that in addition to epigenetic
regulation some other mechanisms could be involved. In support of
this, an insulator protein located 18 kb upstream of the
DLK1-GTL2 locus, which plays an important role in regulating
imprinting of this locus, has been identified in human acute
leukemia cells (25). Thus,
regulation of expression of imprinted genes in RMS as well as
(probably) in other tumor cells requires further study.
Acknowledgements
This study was supported by NIH grants
2R01 DK074720 and R01HL112788 and the Stella and Henry Endowment to
M.Z.R.
References
|
1.
|
Davis E, Jensen CH, Schroder HD, et al:
Ectopic expression of DLK1 protein in skeletal muscle of padumnal
heterozygotes causes the callipyge phenotype. Curr Biol.
14:1858–1862. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
2.
|
Davicioni E, Anderson MJ, Finckenstein FG,
et al: Molecular classification of rhabdomyosarcoma - genotypic and
phenotypic determinants of diagnosis: a report from the Children’s
Oncology Group. Am J Pathol. 174:550–564. 2009.PubMed/NCBI
|
|
3.
|
Hettmer S and Wagers AJ: Muscling in:
Uncovering the origins of rhabdomyosarcoma. Nat Med. 16:171–173.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
4.
|
Chung WY, Yuan L, Feng L, Hensle T and
Tycko B: Chromosome 11p15.5 regional imprinting: comparative
analysis of KIP2 and H19 in human tissues and Wilms’ tumors. Hum
Mol Genet. 5:1101–1108. 1996.PubMed/NCBI
|
|
5.
|
Casola S, Pedone PV, Cavazzana AO, et al:
Expression and parental imprinting of the H19 gene in human
rhabdomyosarcoma. Oncogene. 14:1503–1510. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
6.
|
Ishida M and Moore GE: The role of
imprinted genes in humans. Mol Aspects Med. 34:826–840. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
7.
|
Reik W and Walter J: Genomic imprinting:
parental influence on the genome. Nat Rev Genet. 2:21–32. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
8.
|
Pannetier M and Feil R: Epigenetic
stability of embryonic stem cells and developmental potential.
Trends Biotechnol. 25:556–562. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
9.
|
Delaval K and Feil R: Epigenetic
regulation of mammalian genomic imprinting. Curr Opin Genet Dev.
14:188–195. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
10.
|
Geuns E, De Temmerman N, Hilven P, Van
Steirteghem A, Liebaers I and De Rycke M: Methylation analysis of
the intergenic differentially methylated region of DLK1-GTL2 in
human. Eur J Hum Genet. 15:352–361. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
11.
|
Sasaki H, Ishihara K and Kato R:
Mechanisms of Igf2/H19 imprinting: DNA methylation, chromatin and
long-distance gene regulation. J Biochem. 127:711–715. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
12.
|
Waddell JN, Zhang P, Wen Y, et al: Dlk1 is
necessary for proper skeletal muscle development and regeneration.
PLoS One. 5:e150552010. View Article : Google Scholar : PubMed/NCBI
|
|
13.
|
Markljung E, Jiang L, Jaffe JD, et al:
ZBED6, a novel transcription factor derived from a domesticated DNA
transposon regulates IGF2 expression and muscle growth. PLoS Biol.
7:e10002562009. View Article : Google Scholar : PubMed/NCBI
|
|
14.
|
Duan F, Smith LM, Gustafson DM, et al:
Genomic and clinical analysis of fusion gene amplification in
rhabdomyosarcoma: a report from the Children’s Oncology Group.
Genes Chromosomes Cancer. 51:662–674. 2012.
|
|
15.
|
Kerjean A, Dupont JM, Vasseur C, et al:
Establishment of the paternal methylation imprint of the human H19
and MEST/PEG1 genes during spermatogenesis. Hum Mol Genet.
9:2183–2187. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
16.
|
Higashimoto K, Urano T, Sugiura K, et al:
Loss of CpG methylation is strongly correlated with loss of histone
H3 lysine 9 methylation at DMR-LIT1 in patients with
Beckwith-Wiedemann syndrome. Am J Hum Genet. 73:948–956. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
17.
|
Kobayashi H, Sato A, Otsu E, et al:
Aberrant DNA methylation of imprinted loci in sperm from
oligospermic patients. Hum Mol Genet. 16:2542–2551. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
18.
|
Carr IM, Valleley EM, Cordery SF, Markham
AF and Bonthron DT: Sequence analysis and editing for bisulphite
genomic sequencing projects. Nucleic Acids Res. 35:e792007.
View Article : Google Scholar : PubMed/NCBI
|
|
19.
|
Kagami M, Sekita Y, Nishimura G, et al:
Deletions and epimutations affecting the human 14q32.2 imprinted
region in individuals with paternal and maternal upd(14)-like
phenotypes. Nat Genet. 40:237–242. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
20.
|
Ogata T, Kagami M and Ferguson-Smith AC:
Molecular mechanisms regulating phenotypic outcome in paternal and
maternal uniparental disomy for chromosome 14. Epigenetics.
3:181–187. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
21.
|
Benetatos L, Hatzimichael E, Londin E, et
al: The microRNAs within the DLK1-DIO3 genomic region: involvement
in disease pathogenesis. Cell Mol Life Sci. 70:795–814. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
22.
|
Li L, Forman SJ and Bhatia R: Expression
of DLK1 in hematopoietic cells results in inhibition of
differentiation and proliferation. Oncogene. 24:4472–4476. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
23.
|
Liu L, Luo GZ, Yang W, et al: Activation
of the imprinted Dlk1-Dio3 region correlates with pluripotency
levels of mouse stem cells. J Biol Chem. 285:19483–19490. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
24.
|
Zhou Y, Zhang X and Klibanski A: MEG3
noncoding RNA: a tumor suppressor. J Mol Endocrinol. 48:R45–R53.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
25.
|
Khoury H, Suarez-Saiz F, Wu S and Minden
MD: An upstream insulator regulates DLK1 imprinting in AML. Blood.
115:2260–2263. 2010. View Article : Google Scholar : PubMed/NCBI
|