Introduction
Prostate cancer is the most commonly diagnosed
cancer and second leading cause of cancer-related death in men in
Western countries (1). The most
common treatment is hormone deprivation; however, during disease
progression, the malignant cells lose their androgen receptors and
thereby their hormone dependence and therapeutic responsiveness.
The resulting androgen-independent prostate cancer (AIPC)
progresses and metastasizes, with no effective therapeutic options
at present (2). Although the
cellular composition of prostate cancer is heterogeneous, AIPC
cells are suggested to possess cancer stem cell (CSC)
characteristics (3). According to
the hypothesis, CSCs mediate tumor formation, progression, and
metastasis and do not respond to chemoor radiotherapy (4); therefore, CSCs may be enriched by
treatment, increasing resistance in late-stage cancer. Due to their
ability for self-renewal and differentiation, CSCs can generate all
the cellular subtypes of the original tumor. In addition, these
cells overexpress CSC markers, including ALDH1, CD44, CD133 and
c-Met (3).
Human tumor necrosis factor (TNF)-related apoptosis
ligand (TRAIL) has achieved promising therapeutic results by
targeting only malignant cells and leaving normal cells undamaged.
Although phase 1/2 clinical trials have demonstrated a favorable
toxicity profile for recombinant soluble TRAIL, there is limited
evidence of antitumor activity (5); this may be due to the short exposure
of tumor cells to low concentrations of TRAIL, which has a short
half-life. Moreover, TRAIL signaling does not always cause
apoptosis in cancer cells. Indeed, some studies have shown that
TRAIL may induce a prosurvival response via signaling factors that
include nuclear factor (NF)-κB, mitogen-activated protein kinase
(MAPK), and Akt (5). This finding
is corroborated by data obtained in prostate cancer and
glioblastoma that have indicated that some cancer cells are
resistant to TRAIL; it has been demonstrated that these cells are
CSCs (6). A recent study proposes
that the dietary agent sulforaphane, a mustard oil and
isothiocyanate present in high concentrations in broccoli and
cauliflower (7), sensitizes
prostate cancer cells to TRAIL-induced apoptosis (8), though the authors did not focus on
CSCs. Our recent results demonstrate that sulforaphane sensitizes
resistant pancreatic CSCs to TRAIL-induced apoptosis through the
inhibition of basal and TRAIL-induced NF-κB activity (9). Based on several promising animal and
epidemiological studies, prospective clinical trials with
sulforaphane-enriched broccoli sprout extracts are ongoing in the
US to examine the effects on atypical nevi and bladder and prostate
cancer (10). A pilot study at our
clinic has been initiated to evaluate the effects of
sulforaphane-enriched broccoli sprouts on patients with advanced
pancreatic cancer.
The present study utilized the DU145 and PC3 cell
lines, with highly enriched CSC features, and primary prostate CSCs
to demonstrate that colony- and spheroid formation is strongly
affected by sulforaphane, whereas TRAIL had only minor activity in
targeting CSC features. However, the combination of both agents
acted synergistically, indicating that sulforaphane shifted the
balance of TRAIL-induced survival and suicide signaling to the cell
death pathway. The results may be due to the sulforaphane-mediated
inhibition of NF-κB activity and stem cell signaling, including
tumor engraftment and growth in vivo. Our data provide new
mechanistic insight into the synergistic effects of TRAIL and
sulforaphane on prostate CSCs.
Materials and methods
Tumor cell lines
PC3 and DU145 prostate cancer cell lines were
obtained from American Type Culture Collection (Manassas, VA, USA)
and authenticated throughout culture by the typical morphology. The
cells were cultured in RPMI-1640 (PAA, Pasching, Austria)
supplemented with 10% heat-inactivated FCS (Sigma, Deisenhoffen,
Germany) and 25 mmol/l HEPES (PAA). Human prostate CSCs were
obtained from CELPROGEN (San Pedro, CA, USA) and cultivated in
‘Human Prostate Cancer Complete Growth Medium’ (CELPROGEN) on
‘Human Prostate Cancer Stem Cell Matrix’-coated tissue containers
(CELPROGEN). To maintain the authenticity of the cell lines, frozen
stocks were prepared from the initial stocks, and a new frozen
stock was thawed every three months for the experiments. The
established cell lines were recently authenticated by a commercial
service (Multiplexion, Heidelberg, Germany). Mycoplasma-negative
cultures were ensured by monthly mycoplasma tests.
Cytotoxic agents
D, L-Sulforaphane (Sigma-Aldrich, St. Louis, MO,
USA) was dissolved in ethanol to generate a 100 mM stock.
Recombinant Super Killer TRAIL was obtained from AXXORA (Lörrach,
Germany) and was dissolved in TRAIL buffer AXXORA (Lörrach) to
generate a 100 μg/ml stock solution. The final
concentrations of solvents in the media were ≤0.1%.
Gel retardation analysis of NF-κB
binding
The preparation of nuclear protein extracts and the
bandshift reaction using the Light Shift®
Chemiluminescent EMSA kit were performed as we recently described
(11).
Colony-forming assay
Treated cells were seeded at a cell density of 500
cells/well in complete medium in 6-well tissue culture plates
(TPP), and the colony-forming assay was performed as we recently
described (12).
Spheroid assay
For spheroid formation, the cells were cultured in
NeuroCult NS-A basal serum-free medium (human) (StemCell
Technologies, Vancouver, Canada) supplemented with 2 μg/ml
Heparin (StemCell Technologies), 20 ng/ml hEGF (R&D Systems,
Wiesbaden-Nordenstadt, Germany), 10 ng/ml hFGF-b (PeproTech,
Hamburg, Germany), and NeuroCult NS-A Proliferation Supplements
(StemCell Technologies). For the evaluation of the first generation
of sphere formation, the cells were seeded at clonal density
(5×102 cells/ml) in 12-well low-adhesion plates in 1 ml
medium per well. Upon sphere formation, the spheres were
dissociated, and the cell number was evaluated. The viable cells
were reseeded under the same conditions as mentioned above to
evaluate the potential of secondary sphere formation.
ALDH1 activity
ALDEFLUOR substrate (5 μl; Aldagen, Inc.,
Durham, NC, USA) was added to 1×106 treated PC3 cells in
500 μl assay buffer and incubated for 60 min at 37°C.
Pre-treatment with the ALDH1 inhibitor diethylaminobenzaldehyde was
used as a negative control.
Western blot analysis
Following treatments, the proteins were isolated,
and a western blot analysis was performed as described (9). The following antibodies were used:
mouse mAbs against human Notch 1, CXCR4 (Abcam, Cambridge, UK), and
β-actin (Sigma-Aldrich) and rabbit polyclonal Abs against human
Jagged1 (Abcam), SOX2 and Nanog (Cell Signaling, Danvers, MA,
USA).
Human pluripotent stem cell antibody
array
Nitrocellulose membranes on which capture antibodies
had been spotted and the detection reagents were obtained as a kit
from R&D Systems® (R&D Systems). According to
the instructions of the manufacturer, the protein extracts were
harvested after cell lysis and incubated overnight with the
nitrocellulose membranes. After washing, the membranes were
incubated with biotinylated secondary antibodies and
streptavidin-HRP and chemiluminescent detection reagents were used
to detect binding.
Adipogenic differentiation assay
Tumor cells (1×105) were seeded and
treated in 6-well plates. To induce adipogenic differentiation, the
medium was changed to NH AdipoDiff Medium, 2 ml per well. The
medium was refreshed every third day; after 14 days, the fat
droplets of adipogenic cells were stained with Oil Red O
(Sigma-Aldrich).
Transplantation of tumor cells on
fertilized chicken eggs
This assay was performed as described recently
(13), but with modifications.
Fertilized white leghorn chicken eggs (Geflügelzucht Hockenberger,
Eppingen, Germany) were incubated at a humidity of 45–55% and
37.8°C in digital motor breeders Type 168/D incubator (Siepmann
GmbH, Herdecke, Germany). At day 4 of embryonic development, 2–3 ml
of albumen was removed with a syringe, allowing detachment of the
embryo. A small window was cut into the eggshell and then sealed
with tape. At day 8 of embryonic development, small handmade rings
from Thermanox™ cover discs (Thermo Scientific, Schwerte, Germany)
were placed on the CAM, and 1×106 pre-treated or
untreated tumor cells mixed with Matrigel at a ratio of 1:1 were
deposited into the rings of the viable embryos. For the in
ovo treatment of xenografts, a Whatman paper saturated with
sulforaphane solution (10 μM) was deposited next to
untreated tumors at day 11. At day 12, a TRAIL solution (5 ng/ml)
was dropped onto the Whatman paper until it was saturated. At day
18, the xenografts were resected to determine the tumor engraftment
rate and the tumor volume. All the embryos that died before day 18
were excluded from further analyses. The tumor volumes were
estimated by the following formula: Volume = 4/3 × ∏ ×
r3 (r = 1/2 × square root of diameter 1 × diameter 2)
(13).
Immunofluorescence staining of tumor
xenograft tissue
For immunofluorescence staining, frozen xenograft
tissue was sectioned, and the staining was performed according to a
standard protocol. In short, the tissue was fixed in 4% PFA for 10
min; rabbit polyclonal Abs against human CD44 (Gene Tex, Irvine,
CA, USA), CXCR4, and c-Met (Abcam) and mouse polyclonal Abs against
RelA (Rockland, Gilbertsville, USA), Nanog (Cell Signaling), EpCam
(kindly provided by Dr G. Moldenhauer), ALDH (Becton-Dickinson,
Heidelberg, Germany), and CD133 (Millipore, Bergisch Gladbach,
Germany) were used as the primary Abs. The nuclei were stained with
DAPI (4,6-diamidino-2′-phenylindol, 1 μg/ml). Goat
anti-rabbit Alexa Fluor 488 IgG, goat anti-rabbit Alexa Fluor 594
IgG, goat anti-mouse Alexa Fluor 594 IgG, and goat anti-mouse Alexa
Fluor 488 IgG (Invitrogen, Camarillo, CA, USA) were used as the
secondary Abs.
Immunohistochemical staining of tumor
xenograft tissue
For the immunohistochemical staining of primary
spheroidal cultures and xenograft tissue, the Avidin/Biotin
blocking kit (Vector, Burlingame, CA, USA) was used according to
the instructions of the manufacturer. Endogenous peroxidase was
quenched by 0.3% H2O2 in methanol. The
primary Abs used are described above in the immunofluorescence
section. Biotinylated goat anti-rabbit or anti-mouse IgG (Vector)
served as the secondary Abs. The signal was enhanced using the ABC
Elite kit (Vector). The samples were counterstained with
hematoxylin (Dako, Glostrup, Denmark) and mounted in Pro Tags Mount
Aqua (Quartett, Berlin, Germany). Omission of the primary Abs
served as a negative control. The signals for immunohistochemistry
and immunofluorescence staining were detected at ×400 magnification
using a Leica DMRB fluorescence microscope (Leica, Wetzlar,
Germany). The images of representative fields were obtained using a
SPOT™ FLEX 15.2 64-Mp pixel-shifting digital color camera
(Diagnostic, Instruments, Inc. USA) and analyzed with SPOT
Basic/Advanced 4.6 software.
H&E histochemical staining of liver
sections
Liver tissue obtained from the chicken embryos was
sectioned and stained with eosin (Sigma-Aldrich) and hematoxylin
(Dako) following a standard protocol. The signal was detected at
×400 magnification using a Leica DMRB microscope (Leica). The
images of representative fields were obtained using a SPOT FLEX
15.2 64-Mp pixel-shifting digital color camera (Diagnostic,
Instruments, Inc.) and analyzed with SPOT Basic/Advanced 4.6
software.
Statistical analysis
The quantitative data are presented as the mean ±
SD. The data were analyzed using Student’s t-test for statistical
significance. Variances in the tumor volumes were evaluated with
the Kruskal-Wallis test and Mann-Whitney test with the Bonferroni
correction. *p<0.05 was considered statistically
significant and **p<0.001 as statistically highly
significant.
Results
The combination of sulforaphane and TRAIL
is superior to single treatments in reducing self-renewal
potential
As model cell lines, we used the human prostate cell
lines DU145 and PC3, which are derived from advanced AIPC
metastases. These cells harbor highly enriched CSC characteristics
(9,14–25),
as summarized in Table I. We
performed an electrophoretic mobility shift assay (EMSA) to measure
the effect of TRAIL and sulforaphane on NF-κB activity. PC3 and
DU145 cells were pre-treated with sulforaphane for 24 h to mimic a
steady-state level of nutritional uptake, and the cells were then
treated with TRAIL for an additional 24 h - either alone or in
combination. This treatment schedule was used in all the following
assays. Both cell lines exhibited basal NF-κB activity, which was
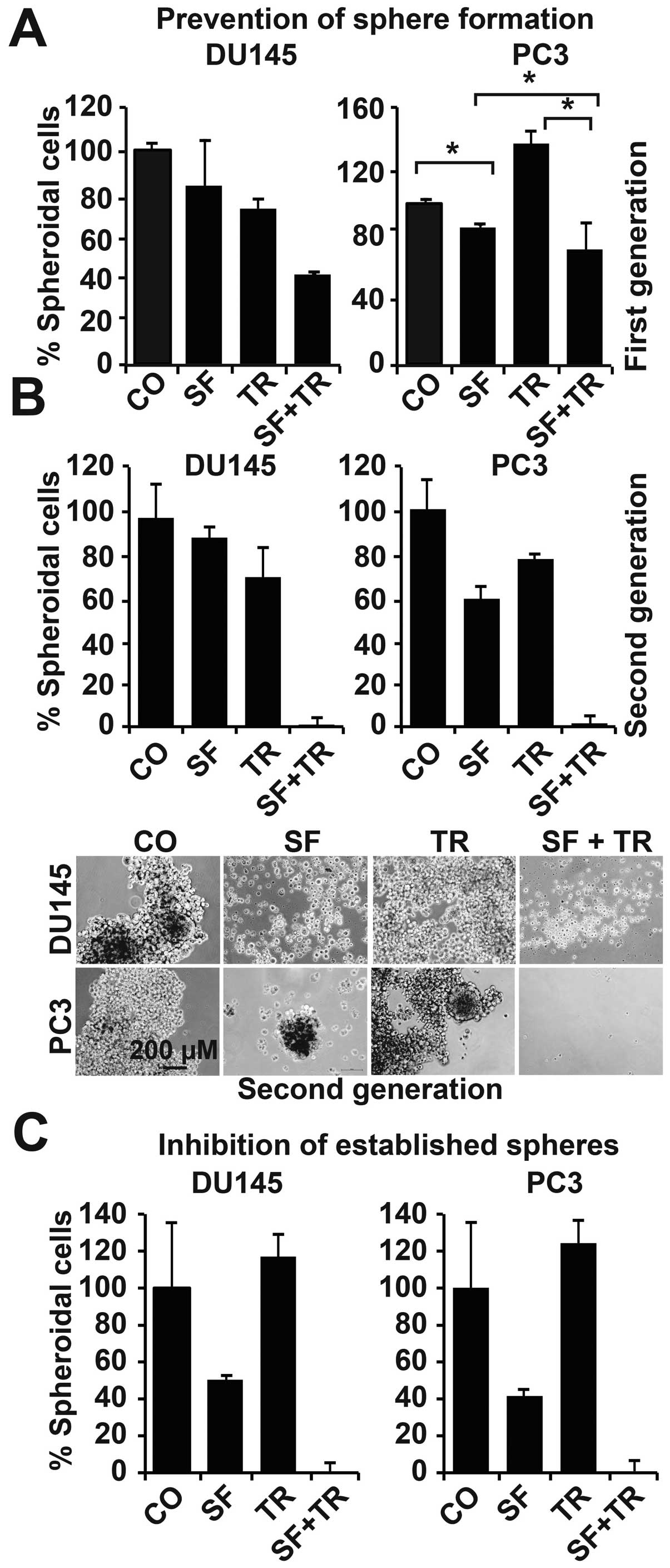
higher in the PC3 cells and further increased by TRAIL (Fig. 1A). Sulforaphane inhibited this
basal NF-κB activity and completely prevented TRAIL-induced NF-κB
activity in the combination treatment. To examine the effect on
self-renewal potential, the cells were seeded at a low density
after treatment, and the formation of colonies was evaluated after
10 days (Fig. 1B). Sulforaphane
significantly reduced clonogenic cell division to ∼50%, whereas
TRAIL had only minor effects; however, synergism occurred in the
combination treatment, as colony formation was almost completely
inhibited. To confirm these results in vivo, we
xenotransplanted untreated or in vitro-treated PC3 cells to
the chorioallantois membrane (CAM) of fertilized chicken eggs; nine
days later, the self-renewal capacity was evaluated by determining
of the ability to form tumors in vivo, a feature attributed
to CSCs (Fig. 1C). Sulforaphane
reduced the rate of tumor engraftment of the untreated control
cells from 78 to 43%, TRAIL to 38%, and the double treatment to
13%. The average volume of the tumors that developed in each group
was 40, 7, 15 and 2 mm3 for the control, sulforaphane,
TRAIL and combination groups, respectively. To further investigate
these results, the self-renewal potential of the cells was
evaluated by an analysis of the anchorage-independent spheroidal
growth, another typical feature of stem cells. Two different types
of sphere assays were performed to measure: i) the effect of
treatment on spheroid formation, and ii) the effect of treatment on
established spheroids. In the first assay, we seeded pretreated
cells at clonal density under conditions that favor the
proliferation of stem cells; seven days later, the number of
spheroidal-growing cells was determined by dissociating the
spheroids and counting the viable cells (Fig. 2A). The percentage of
spheroidal-growing cells was reduced by sulforaphane in both cell
lines to ∼80%; TRAIL also led to a reduction of 80% in the DU145
cells but to enhanced spheroid formation of 130% in the PC3 cells.
The combined treatment significantly further reduced the percentage
of spheroidal-growing cells to 40% in the DU145 cells, whereas the
reduction was not significant in the PC3 cells. Furthermore, the
cells were damaged by the combined treatment, which was
demonstrated by re-seeding equal numbers of the cells at clonal
density for spheroid formation without additional treatment and
determining the number of spheroidal-growing cells after seven days
(Fig. 2B). A substantial number of
cells derived from the sulforaphaneor TRAIL-treated groups were
still able to form spheroids, whereas none of the cells derived
from the combination group formed spheroids. In a third test, we
evaluated the efficacy of the treatment in reducing the percentage
of cells derived from established spheres (Fig. 2C). The cells were seeded at clonal
density for spheroid formation and treated three days later; the
efficacy was evaluated after an additional seven days. Sulforaphane
alone reduced the percentage of spheres to ∼40%, whereas TRAIL
slightly induced sphere formation in both cell lines. Moreover, the
co-treatment completely destroyed sphere formation. These results
demonstrate that the combination of sulforaphane and TRAIL, but not
the single agents, is sufficient to completely eliminate the
self-renewal potential of DU145 and PC3 cells.
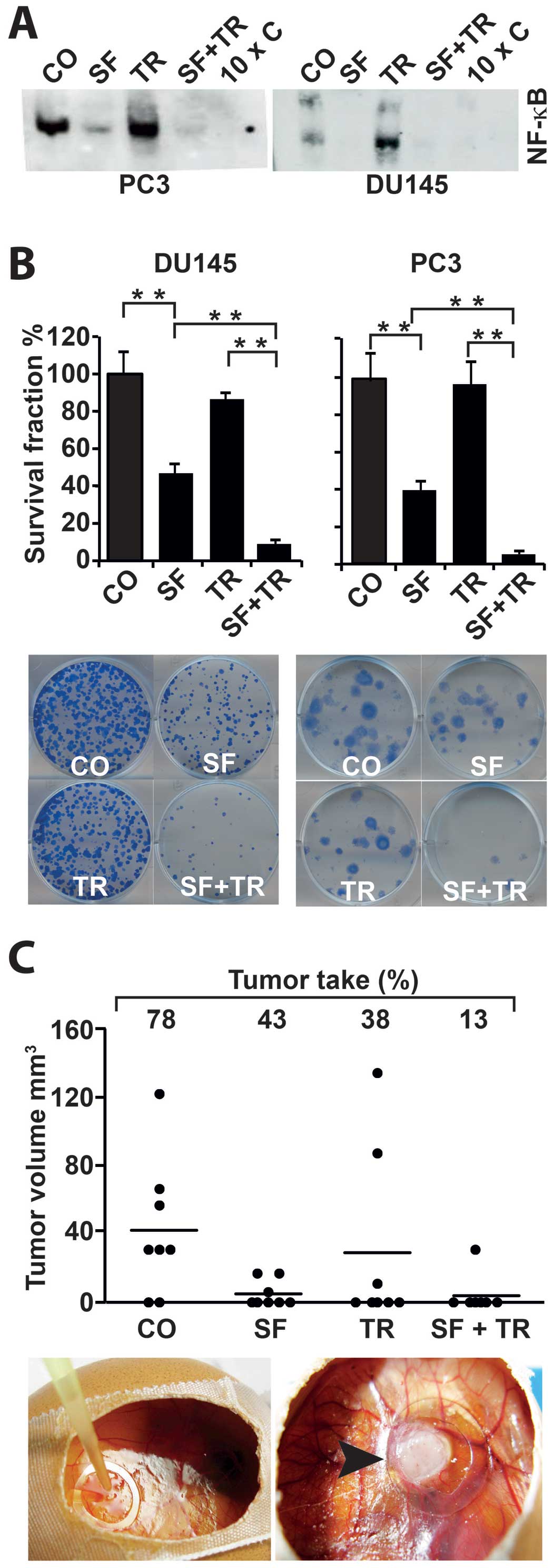 | Figure 1.The combination of sulforaphane and
TRAIL is superior to single treatments in reducing self-renewal
potential. (A) DU145 and PC3 cells were left untreated or were
treated with 10 μM sulforaphane; after 24 h, TRAIL was added
at a concentration of 5 ng/ml. After an additional 24 h, the
nuclear proteins were harvested and DNA binding was analyzed by
EMSA using a biotin-labeled oligonucleotide probe for the NF-κB
promoter consensus sequence. The specific NF-κB shifts are marked.
Competition with a 10-fold excess of unlabeled oligonucleotide (10
× C) served as a control for the binding specificity. (B)
Twenty-four hours after TRAIL treatment, the cells were trypsinized
and re-plated in a normal medium at a low density (500 cells/well)
in 6-well plates. Ten days later, the cells were stained, and
colonies containing >50 cells were counted under a dissecting
Zeiss Stemi DV4 microscope. Images of the fixed and stained
colonies are presented in the lower panel. The data are presented
as the mean of three independent experiments, and SD are shown
(*p<0.05, **p<0.01). (C) Following
in vitro treatment as described above, PC3 cells were
transplanted onto the chorioallantoic membrane of fertilized
chicken eggs at day 8 of embryonic development. Nine days later
(day 17), the developed xenograft tumors were resected, and the
tumor engraftment rates and tumor volumes were evaluated. The tumor
volumes are presented as black dots and tumor engraftment is
presented as the percentage of grown tumors relative to the number
of each treatment group. Below the diagram, representative images
show the transplantation of the tumor cells into a plastic ring on
the CAM (left images), and a developed PC3 xenograft is marked with
an arrow (right image). |
 | Table I.CSC characteristics of established
human AIPC cell lines. |
Table I.
CSC characteristics of established
human AIPC cell lines.
| DU145 | PC3 | Ref(s). |
|---|
| ATCC no. | HTB-81 | CRL-1435 | ATCC |
| Source | Brain
metastasis | Bone metastasis,
grade IV | ATCC |
| p53 status | Mutant | Absent | (14) |
| Growth
characteristics | Adherent | Adherent | ATCC |
| Colony-forming
capacity | High | High | (15) |
| Spheroid-formation
capacity | None | None | (9) |
| ALDH activity | 2.4±0.3 | 6.3±1.0 | (16) |
| Growth in nude
mice | Rapid, 100
CD44+/CD24− cells form a tumor in 5 of 5 mice
within 80 days | Rapid, 100 PC3
cells form a tumor in 1 of 8 mice within 90 days | (17,18) |
|
CD44+/CD24− | 7–10% | Detectable | (9,17,19,20) |
| CD133 | 0.01% | Setectable | (15,21) |
| α2β1 | 1–10% | 100% | (22) |
| Invasiveness in
Matrigel | > 20%, highly
invasive | Highly
invasive | (23,24) |
| E-cadherin
expression | Low | Low | (23,25) |
The combination of sulforaphane and TRAIL
is superior to single treatments in reducing CSC-related
signaling
To study the influence of sulforaphane and TRAIL on
stem cell signaling, we performed an antibody protein array to
detect the levels of human pluripotent stem cell markers. PC3 cells
were treated; the proteins were isolated after 24 h and incubated
with the array membranes (Fig.
3A). TRAIL showed no or minor effects, whereas sulforaphane
reduced the amount of Nanog, Sox2, E-cadherin, GATA-4, HNF-3β,
SOX17, Otx2, TP63, Snail, VEGF R2 and HCG. Moreover, the
combination treatment further reduced the levels of Oct-3/4,
HNF-3β, PDX-1, Otx2, TP63, GSC, Snail, VEGF R2 and HCG. These
results demonstrate that sulforaphane, particularly in combination
with TRAIL, reduces the levels of proteins required for
self-renewal, differentiation, cell migration, the
epithelialmesenchymal transition (EMT) and tumorigenesis (26–41)
(Table II). To further elucidate
the obtained array results, we examined CSC marker proteins in the
PC3 and DU145 cells by a western blot analysis (Fig. 3B). The expression of the CXCR4
receptor, which is involved in migration and metastasis (42), was inhibited following the
sulforaphane-only treatment and the combination with TRAIL further
reduced the expression. Similar results were found for the Notch 1
receptor and its ligand Jagged1, which are regulators of asymmetric
and symmetric division, progression, and metastasis in prostate
cancer (43). Correspondingly,
sulforaphane, but not TRAIL, inhibited the expression of SOX2 and
Nanog, which are important regulators of self-renewal potential
(28), and the effects were
stronger with the combination. Because ALDH1 activity has been
reported to be required for self-renewal potential (44), we examined the enzymatic activity
using a substrate assay and flow cytometry in PC3 cells (Fig. 3C). Sulforaphane significantly
reduced the ALDH1 activity from ∼30 to 12%; conversely, TRAIL
increased the activity to 45% and the combination treatment to 5%.
We next evaluated the influence of the treatments on the
differentiation potential of PC3 and DU145 cells; after treatment,
the cell culture medium was exchanged for an adipocytic
differentiation medium, and the formation of adipocytes was
evaluated by staining the cells with Oil Red O after fourteen days
(Fig. 3D). Although the untreated
cells exhibited a high percentage of red cells, reflecting fat
droplets, such fat droplets were completely absent in all the
treatment groups, indicating that sulforaphane and TRAIL inhibited
the cell differentiation potential. These data suggest that
sulforaphane strongly inhibits stem cell signaling, ALDH1 activity,
and differentiation potential, though TRAIL has only minor effects;
both agents in combination resulted in the strongest effect.
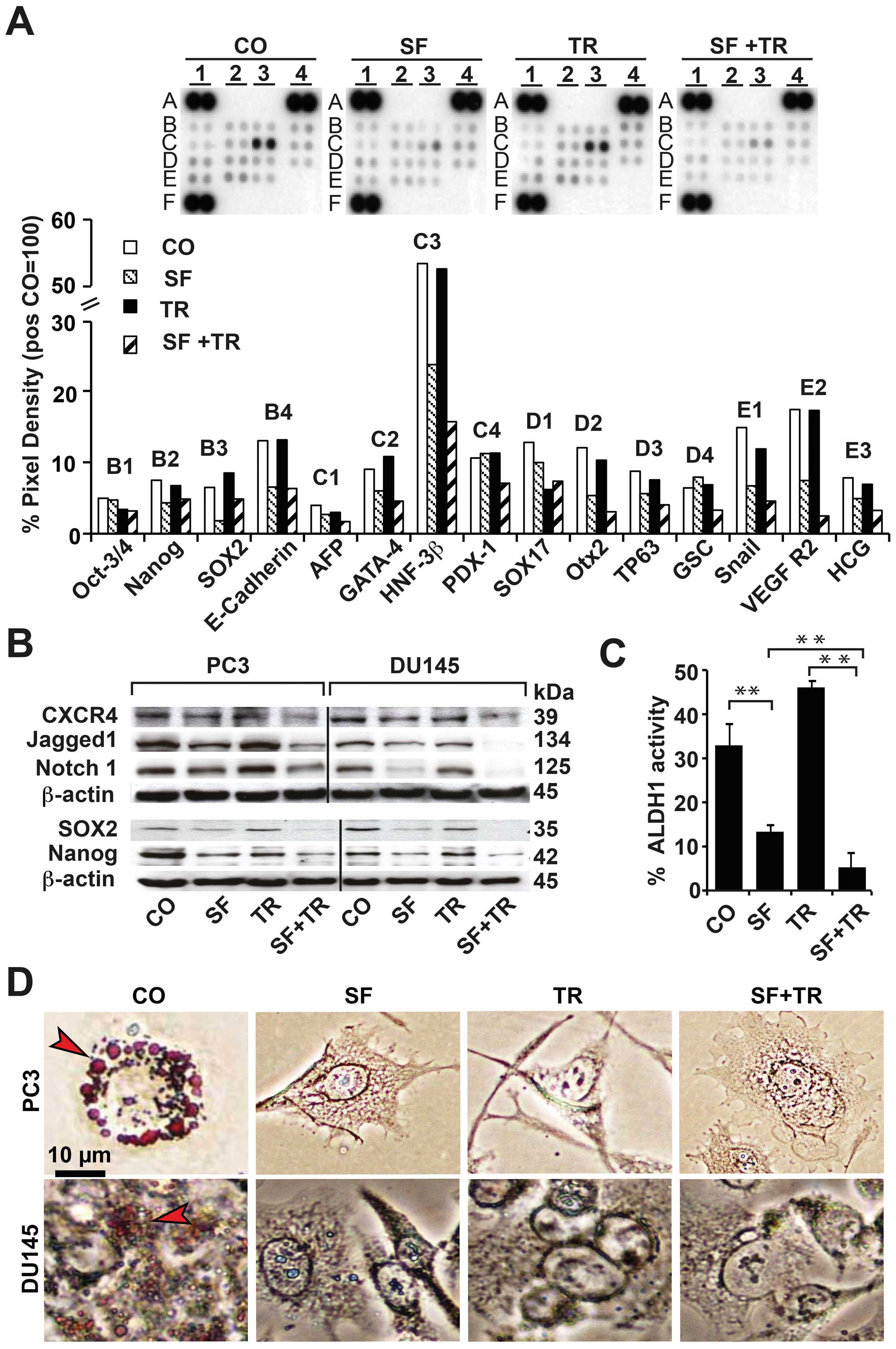 | Figure 3.Sulforaphane, but not TRAIL, strongly
inhibits stem cell-associated signaling and differentiation and
their combination enhances these effects. (A) PC3 cells were
treated as described in Fig. 1A.
Twenty-four hours later, the proteins were isolated and a human
pluripotent stem cell array was performed. The binding of proteins
to antibodies spotted in duplicate on the membranes was detected
using biotinylated secondary antibodies, streptavidin-HRP and
chemiluminescence. After normalization to reference spots (positive
control A1, A4 and F1; PBS-negative control E4), the pixel density
was quantified using ImageJ software. (B) Proteins were also
harvested, and a western blot analysis was performed to detect the
expression of CXCR4, Jagged1, Notch 1, SOX2 and Nanog. The
detection of β-actin served as a control for equal conditions. (C)
For ALDH1 evaluation, PC3 cells were treated as described in
Fig. 1A; the activity of ALDH1 was
determined by a substrate assay and the turnover was analyzed by
flow cytometry. The data are presented as the percentage of
ALDH1-positive cells ± SD (*p<0.05,
**p<0.01). (D) DU145 and PC3 cells were treated as
described in Fig. 1A. Twenty-four
hours later, the medium was exchanged for NH AdipoDiff medium to
induce adipocyte differentiation. After 14 days, the cells were
stained with Oil Red O to detect the fat droplets in adipocytes.
Representative images are shown, and the arrows indicate the red
fat droplets. The scale bar indicates 10 μm. |
| Table II.Human pluripotent stem cell antibody
array. |
Table II.
Human pluripotent stem cell antibody
array.
| Marker | Function | Ref. |
|---|
| AFP | α-fetoprotein, the
fetal form of serum albumin, a tumor marker for hepatocellular
carcinoma, germ cell tumors and metastatic liver cancer. | (26) |
| Oct-3/4 | Transcription
factor involved in self-renewal. Associated with an
undifferentiated phenotype and tumors. | (27) |
| NANOG | Transcription
factor involved in maintaining the pluripotency of stem cells. | (28) |
| SOX2 | Transcription
factor essential for the self-renewal and pluripotency of stem
cells. | (29) |
| E-cadherin | Cell-cell adhesion
glycoprotein. Loss of function contributes to the progression of
cancer by increasing proliferation and invasion. | (30) |
| GATA-4 | Zinc-finger
transcription factor important in differentiation and
embryogenesis. Decreased GATA-4 expression has been associated with
carcinogenesis. | (31) |
| FOXA2 | DNA-binding
protein, the dysregulation of which has been linked to
inflammation, tumorigenesis, and EMT. | (32, 33) |
| PDX-1 | Transcription
factor involved in the reprogramming of differentiated mouse
pancreatic exocrine cells into β-cells. | (34) |
| SOX17 | Transcription
factor necessary for differentiation and the antagonization of
self-renewal. | (35) |
| Otx2 | Orthodenticle
homeobox 2 is a homeobox transcription factor that controls brain
morphogenesis and brain development. | (36) |
| p63 | Member of the p53
family that is important for development and interactions between
mesenchyme and epithelium. Loss may be involved in
tumorigenesis. | (37) |
| GSC | The goosecoid
homeobox gene is repressed during stem cell differentiation. | (38) |
| Snail | Zinc-finger
transcription factor involved in inducing EMT during cancer
progression via the downregulation of E-cadherin. | (39) |
| VEGF R2 | Main receptor for
VEGF involved in vasculogenesis, angiogenesis, proliferation,
migration, survival and increased permeability. | (40) |
| hCG | Peptide hormone
that meditates immune tolerance in pregnancy and tumors. | (41) |
The combination of sulforaphane and TRAIL
is superior to single treatments in reducing tumor growth and CSC
marker expression in a xenograft model
To evaluate whether sulforaphane, TRAIL, and both
agents together inhibit tumor growth and stem cell signaling in
in vivo-treated xenografts, we transplanted untreated PC3
cells at day 9 of embryonic development into fertilized chicken
eggs. The eggs were treated with sulforaphane at day 11, followed
by treatment with TRAIL at day 12, and the xenograft tumors were
resected at day 18. The average volume of the untreated xenograft
tumors was 20 mm3, and sulforaphane or TRAIL reduced
this to ∼15 mm3 (Fig.
4A). However, the combination of sulforaphane and TRAIL nearly
abolished tumor growth, with the average volume reduced to 4
mm3. The analysis of the body weight of the chicken
embryos revealed no significant difference between the treatment
groups and no developmental defects were detectable (Fig. 4B). Similarly, liver necrosis did
not occur, as evaluated by H&E staining of embryonal liver
sections and microscopy (Fig. 4C).
These results indicate that the treatment was well tolerated in
vivo, without the induction of obvious side effects in the
chicken embryos. Next, we analyzed apoptosis induction by staining
the tumor sections with an antibody specific for the cleaved
fragment of activated caspase-3 (Fig.
4D) and found that caspase-3 activity was increased in all the
treatment groups but was strongest following the combined
treatment. The tumor tissue was further analyzed by double
immunofluorescence staining (Fig.
4E), which demonstrated that sulforaphane and TRAIL reduced the
expression of the CSC markers CD133, CXCR4, Nanog, c-Met, EpCAM,
CD44, and ALDH1 and the proliferation marker Ki67; the inhibition
was much more pronounced with the combination of sulforaphane and
TRAIL. These results indicate that sulforaphane and TRAIL reduce
tumor growth in vivo, that their combination shows the
strongest effects, without obvious side effects, and that the
observed results are associated with the inhibition of CSC
markers.
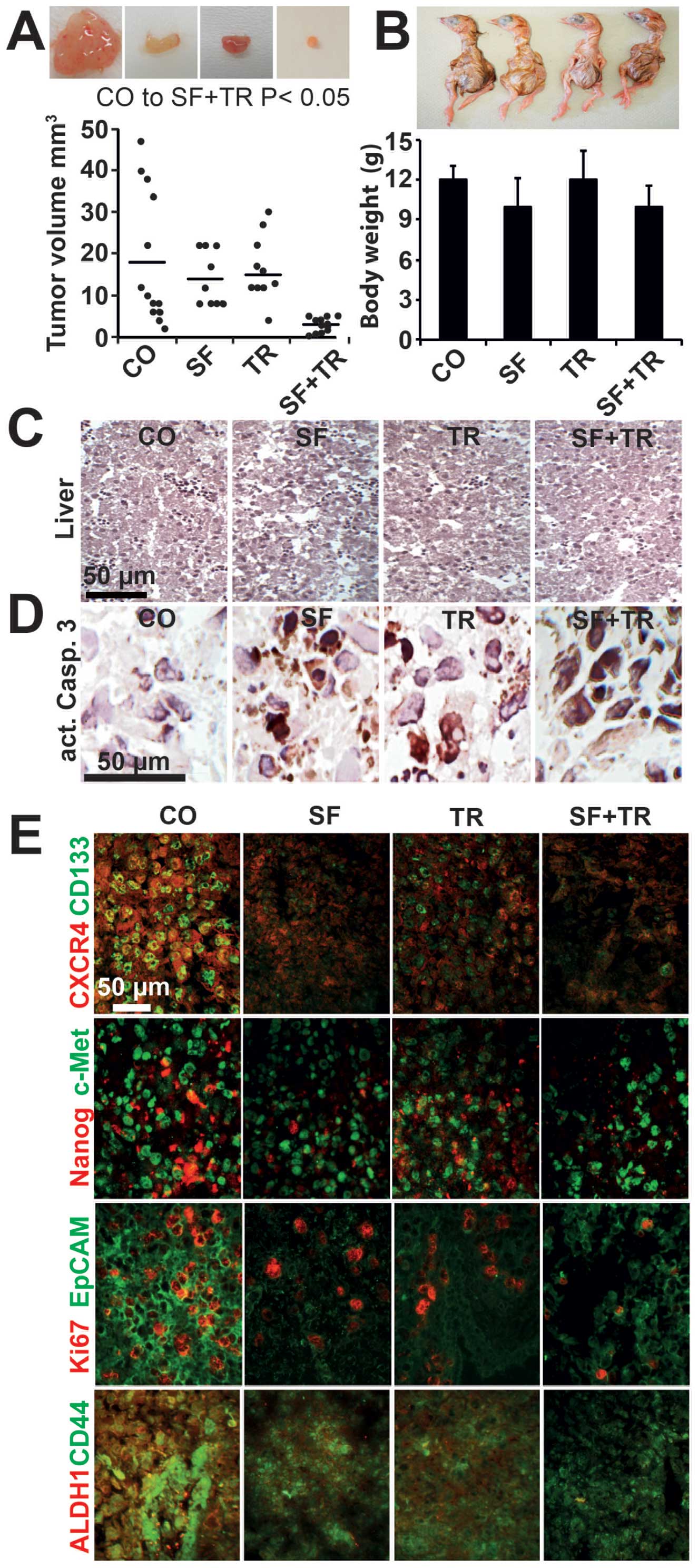 | Figure 4.TRAIL and sulforaphane inhibit tumor
growth in vivo and reduce CSC marker expression, with the
strongest effects after their combination. (A) Untreated PC3 cells
in Matrigel were transplanted into a plastic ring on the
chorioallantoic membrane of fertilized chicken eggs at day 9 of
embryonic development. At day 11, a 1-cm2 Whatman paper
saturated with 10 μM sulforaphane was placed directly
adjacent to the tumor plastic ring. At day 12, a 5 μg/ml
TRAIL solution was dropped onto the Whatman paper until saturation.
The tumor xenografts were resected at day 18 of embryonic
development, and the volumes were determined as described in
Materials and methods. The volumes of the individual tumors per
group are presented as black dots and the bars indicate the average
tumor size of each group. Representative images of the resected
tumors are shown in the upper panel. (B) After tumor resection, the
morphology and the body weights of the chicken embryos were
evaluated. The average body weights of the embryos per group are
presented in the diagram, and images of representative chicken
embryos are shown. (C) The livers of the embryos were sectioned,
and H&E staining was performed and visualized using microscopy.
(D) Slices of the tumor tissue were immunohistochemically stained
with an antibody for the detection of the cleaved fragment of
active caspase-3 and the signal was detected using microscopy. (E)
Slices of tumor tissue were double-immunofluorescence stained with
antibodies for the detection of CD133/CXCR4, Nanog/c-Met,
Ki67/EpCAM, and ALDH1/CD44 and positive signals were detected by
fluorescence microscopy. The bar indicates 50 μm. |
The combination of sulforaphane and TRAIL
is superior to single treatments in reducing the growth and stem
cell marker expression of primary prostate CSCs
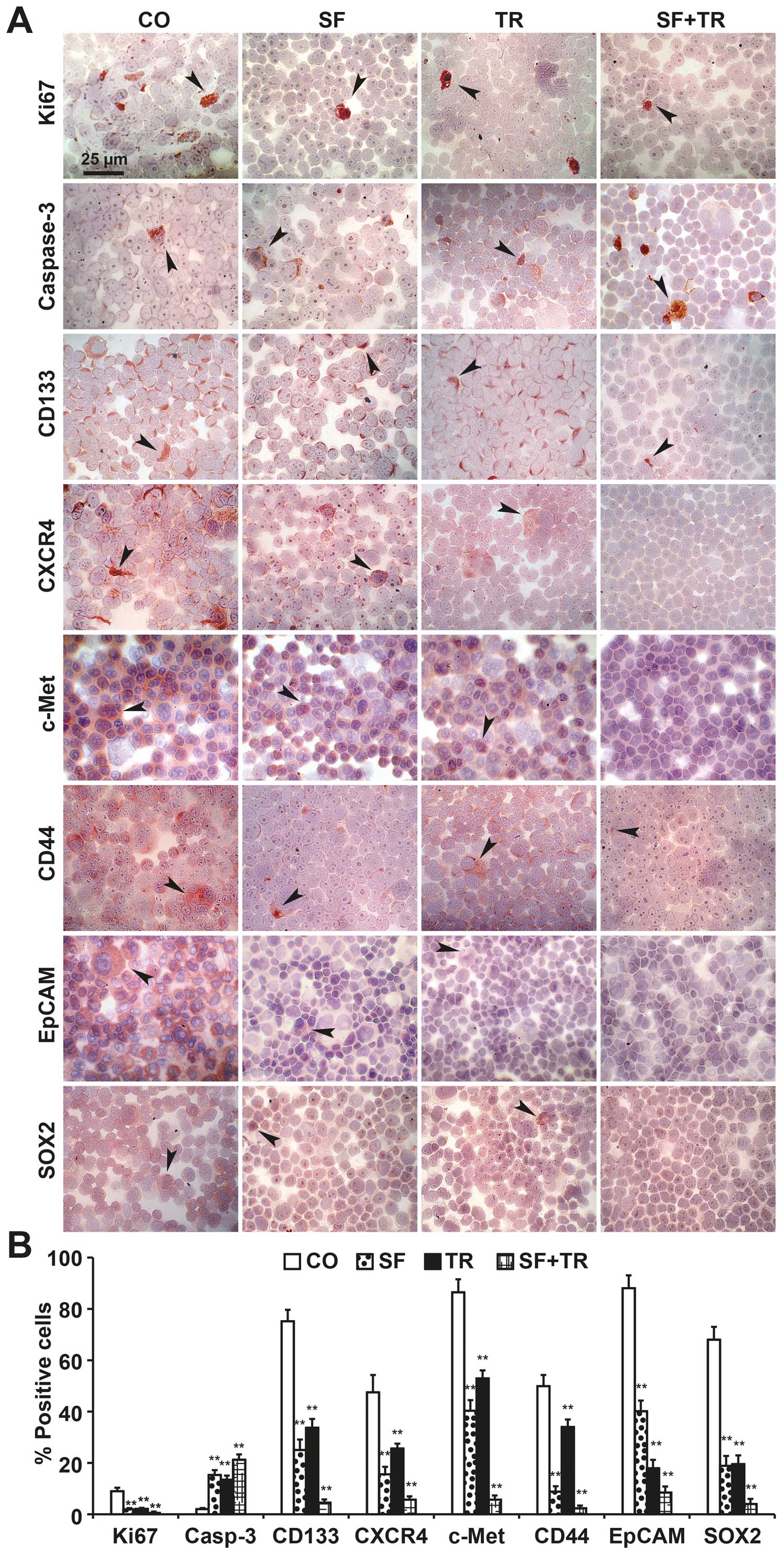
To evaluate the effect of sulforaphane and TRAIL in
a more patient-related model, we treated primary prostate CSCs that
were directly derived from patient tumors. Following the treatment,
the cells were cytospinned onto microscope slides, and positive
cells with enhanced apoptosis or inhibited proliferation and CSC
marker expression were detected by immunohistochemistry (Fig. 5A). The amount of positive cells was
quantified by the evaluation of the number of positive-stained
cells in 10 vision fields. Fig. 5B
shows representative images and a diagram of the enhanced levels of
the apoptosis marker ‘cleaved fragment of active caspase-3’ and
reduced levels of the proliferation marker Ki67 and the CSC markers
CD133, CXCR4, c-Met, CD44, EpCAM and SOX2. Each single treatment
was effective, though the combination of sulforaphane and TRAIL
exhibited the strongest effects. These data strengthen our concept
that a nutritional strategy for enhanced sulforaphane intake could
enhance the efficacy of TRAIL in attacking prostate CSCs.
Discussion
TRAIL is selectively toxic to malignant, but not to
non-malignant, cells and therefore is a promising anticancer agent
(5). We examined whether TRAIL is
able to eliminate AIPC cell lines enriched in CSC features and
primary prostate CSCs. We found an induction of NF-κB activity by
TRAIL that was associated with a minor efficacy of TRAIL in
inhibiting clonogenicity, tumor engraftment and growth, spheroid
formation and CSC signaling. In contrast, the treatment of cells
with sulforaphane was largely more effective than TRAIL in
eliminating CSC features. Furthermore, the combination of TRAIL
with sulforaphane completely prevented basal TRAIL-induced NF-κB
activity and synergistically led to a nearly complete elimination
of CSCs. This result is in accordance with our recent data obtained
for pancreatic CSCs, which exhibited enhanced basal NF-κB activity
due to increased binding of the c-Rel and RelA subunits, which was
further induced by TRAIL (9).
Because the downregulation of c-Rel by siRNA restores sensitivity
to TRAIL-induced apoptosis, the NF-κB prosurvival response induced
by TRAIL may compete with its proapoptotic signals, and NF-κB
activity may be a crucial regulator of the cellular outcome of
survival or suicide.
Our present study supports the recent idea that
prostate CSCs are resistant to TRAIL-induced apoptosis (6). This finding is in agreement with a
prior study that reported that the TRAIL resistance of glioblastoma
CSCs was due to the insufficient expression of the death receptors
DR4 and DR5 and the inhibition of the CD95/Fas domain (45). In the same study, TRAIL resistance
was circumvented by additional cisplatin treatment, which restored
the expression of death receptors and Fas domain activity (45). Although our report does not address
the expression of TRAIL receptors, we cite a paper that
demonstrated the upregulation of the death receptors DR4 and DR5 in
TRAIL-resistant PC3 and LNCap prostate cancer cells by
sulforaphane, which enhanced TRAIL-induced apoptosis in
vitro and in orthotopically growing PC3 xenografts transplanted
into the prostate gland of immunodeficient mice (8).
For our in vivo studies, we used the
xenotransplantation of PC3 cells into the CAM of fertilized chicken
eggs. TRAIL was capable of reducing tumor engraftment and tumor
growth to ∼50%, demonstrating the reduction of tumorigenic cells.
Sulforaphane had similar effects in vivo, and tumor growth
was completely inhibited by the combination treatment. These
results suggest that the inhibition of TRAIL-induced survival
signaling by sulforaphane might have switched the TRAIL response to
apoptosis induction and that this, together with the cytotoxicity
of sulforaphane, resulted in synergism and the complete elimination
of the tumorigenic cells. Most importantly, neither the single
treatments nor the combined agents had any obvious side effects on
the chicken embryos, as developmental defects, weight loss or liver
necrosis did not occur. In contrast, caspase-3 activity was
induced, and proliferation and the expression of CSC markers CD133,
CXCR4, c-Met, Nanog, EpCAM, CD44 and ALDH1 were almost completely
abolished after the in ovo treatment of the PC3 xenograft
tumors. These effects may be due to the observed interference of
sulforaphane with NF-κB activity, an assumption that is underscored
by the recent finding that tumor-initiating stem-like cells in
human prostate cancer exhibit increased NF-κB signaling (46).
In our in vitro studies, we employed an
antibody protein array and western blot analysis to confirmed the
above described in vivo data. Our results showed that
sulforaphane strongly inhibits the expression of the CSC proteins
Nanog, SOX2, CXCR4, Jagged1, and Notch 1 and that of Snail, a
mediator of the epithelial-mesenchymal transition (47). In addition, we found a
sulforaphane-mediated inhibition of ALDH1 activity, which is known
to be high in tumor-initiating and metastasis-initiating cells in
human prostate cancer (16).
Although TRAIL was not effective in reducing these progression
markers, it synergistically enhanced the effect of sulforaphane in
combination.
The targeting of prostate CSCs by sulforaphane may
be expected to occur in patients, as exemplified by two recent
epidemiological studies. Data from a prospective Canadian
epidemiological study suggest that a high consumption of
cruciferous vegetables, which contain high amounts of sulforaphane
and related bioactive agents, is associated with the inhibition of
metastasis (48). In the latter
study, the dietary patterns of 1138 men with prostate cancer were
evaluated, with a median follow-up of 4.2 years. Although the
consumption of vegetables in general significantly reduced the
relative risk (RR) of metastasis to 0.41, cruciferous vegetables
showed the highest effect among all vegetables (crucifers in
general RR=0.60; cabbage RR=0.64; cauliflower RR=0.48; broccoli RR=
0.55); of the crucifers, broccoli and cauliflower were most
effective, with a significant effect at almost 1 serving per week
and 3–5 servings being more effective. In a similar study, Richman
and colleagues (49) prospectively
examined the post-diagnostic intake of vegetables, particularly
cruciferous vegetables, and the relationship with prostate cancer
progression risk. These authors reported a 59% reduced risk of
prostate cancer progression following cruciferous vegetable intake
in men after diagnosis, with an inverse association for total
vegetables that was not significant.
For future therapeutic application of our
experimental strategy, we suggest a dietary intervention together
with the targeted delivery of a protected TRAIL protein to tumors
to avoid dilution in the body or inactivity due to the short
half-life. In this regard, we are currently evaluating the
suitability of the transduction of mesenchymal stem cells with
TRAIL-expressing oncolytic adenoviruses. The supporting
cytotoxicity of oncolytic adenoviruses may shift the balance of
TRAIL signaling from survival to tumor-specific apoptosis.
Abbreviations:
|
AIPC
|
androgen-independent prostate
cancer;
|
|
CSC
|
cancer stem cell
|
Acknowledgements
We thank Dr G. Moldenhauer for
providing the anti-EpCAM antibody, Dr K. Müller-Decker, Dr E.
Ryschich and Dr F. Fortunato for helpful discussion, and S.
Bregulla and S. Ogrodnik for excellent technical assistance. This
study was supported by grants from the German Cancer Aid (Deutsche
Krebshilfe 109362), German Research Community (DFG HE 3186/11-1),
Federal Ministry of Education and Research (BMBF 031A213),
German-Israeli Foundation for Scientific Research and Development
(GIF 1058-7.11/2008), Heidelberger Stiftung Chirurgie, Stiftung für
Krebs und Scharlachforschung, and Hanns A. Pielenz Stiftung.
References
|
1.
|
Siegel R, Naishadham D and Jemal A: Cancer
statistics, 2012. CA Cancer J Clin. 62:10–29. 2012. View Article : Google Scholar
|
|
2.
|
Feldman BJ and Feldman D: The development
of androgen-independent prostate cancer. Nat Rev Cancer. 1:34–45.
2001. View
Article : Google Scholar : PubMed/NCBI
|
|
3.
|
Sharpe B, Beresford M, Bowen R, Mitchard J
and Chalmers AD: Searching for prostate cancer stem cells: markers
and methods. Stem Cell Rev. 9:721–730. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
4.
|
Abbott A: Cancer: the root of the problem.
Nature. 442:742–743. 2006. View
Article : Google Scholar : PubMed/NCBI
|
|
5.
|
Stuckey DW and Shah K: TRAIL on trial:
preclinical advances in cancer therapy. Trends Mol Med. 19:685–694.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
6.
|
Szegezdi E, O’Reilly A, Davy Y, et al:
Stem cells are resistant to TRAIL receptor-mediated apoptosis. J
Cell Mol Med. 13:4409–4414. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
7.
|
Herr I and Büchler MW: Dietary
constituents of broccoli and other cruciferous vegetables:
implications for prevention and therapy of cancer. Cancer Treat
Rev. 36:377–383. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
8.
|
Shankar S, Ganapathy S and Srivastava RK:
Sulforaphane enhances the therapeutic potential of TRAIL in
prostate cancer orthotopic model through regulation of apoptosis,
metastasis, and angiogenesis. Clin Cancer Res. 14:6855–6866. 2008.
View Article : Google Scholar
|
|
9.
|
Kallifatidis G, Rausch V, Baumann B, et
al: Sulforaphane targets pancreatic tumour-initiating cells by
NF-kappaB-induced anti-apoptotic signalling. Gut. 58:949–963. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
10.
|
Herr I, Lozanovski V, Houben P, Schemmer P
and Buchler MW: Sulforaphane and related mustard oils in focus of
cancer prevention and therapy. Wien Med Wochenschr. 163:80–88.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
11.
|
Liu L, Salnikov AV, Bauer N, et al:
Triptolide reverses hypoxia-induced EMT and stem-like features in
pancreatic cancer by NF-kappa B downregulation. Int J Cancer. (In
press).
|
|
12.
|
Apel A, Herr I, Schwarz H, Rodemann HP and
Mayer A: Blocked autophagy sensitizes resistant carcinoma cells to
radiation therapy. Cancer Res. 68:1485–1494. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
13.
|
Balke M, Neumann A, Szuhai K, et al: A
short-term in vivo model for giant cell tumor of bone. BMC Cancer.
11:2412011. View Article : Google Scholar : PubMed/NCBI
|
|
14.
|
van Bokhoven A, Varella-Garcia M, Korch C,
et al: Molecular characterization of human prostate carcinoma cell
lines. Prostate. 57:205–225. 2003.PubMed/NCBI
|
|
15.
|
Pfeiffer MJ and Schalken JA: Stem cell
characteristics in prostate cancer cell lines. Eur Urol.
57:246–255. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
16.
|
van den Hoogen C, van der Horst G, Cheung
H, et al: High aldehyde dehydrogenase activity identifies
tumor-initiating and metastasis-initiating cells in human prostate
cancer. Cancer Res. 70:5163–5173. 2010.
|
|
17.
|
Hurt EM, Kawasaki BT, Klarmann GJ, Thomas
SB and Farrar WL: CD44+CD24(-) prostate cells are early
cancer progenitor/stem cells that provide a model for patients with
poor prognosis. Br J Cancer. 98:756–765. 2008.
|
|
18.
|
Li H, Chen X, Calhoun-Davis T, Claypool K
and Tang DG: PC3 human prostate carcinoma cell holoclones contain
self-renewing tumor-initiating cells. Cancer Res. 68:1820–1825.
2008. View Article : Google Scholar
|
|
19.
|
Patrawala L, Calhoun T,
Schneider-Broussard R, et al: Highly purified CD44+
prostate cancer cells from xenograft human tumors are enriched in
tumorigenic and metastatic progenitor cells. Oncogene.
25:1696–1708. 2006.
|
|
20.
|
Glinsky GV: ‘Stemness’ genomics law
governs clinical behavior of human cancer: implications for
decision making in disease management. J Clin Oncol. 26:2846–2853.
2008.
|
|
21.
|
Luk SU, Yap WN, Chiu YT, et al:
Gamma-tocotrienol as an effective agent in targeting prostate
cancer stem cell-like population. Int J Cancer. 128:2182–2191.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
22.
|
Patrawala L, Calhoun-Davis T,
Schneider-Broussard R and Tang DG: Hierarchical organization of
prostate cancer cells in xenograft tumors: the
CD44+alpha2beta1+cell population is enriched
in tumor-initiating cells. Cancer Res. 67:6796–6805. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
23.
|
Klarmann GJ, Hurt EM, Mathews LA, et al:
Invasive prostate cancer cells are tumor initiating cells that have
a stem cell-like genomic signature. Clin Exp Metastasis.
26:433–446. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
24.
|
Shukla S, Maclennan GT, Hartman DJ, Fu P,
Resnick MI and Gupta S: Activation of PI3K-Akt signaling pathway
promotes prostate cancer cell invasion. Int J Cancer.
121:1424–1432. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
25.
|
Zhao Y, Yan Q, Long X, Chen X and Wang Y:
Vimentin affects the mobility and invasiveness of prostate cancer
cells. Cell Biochem Funct. 26:571–577. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
26.
|
Tomasi TB Jr: Structure and function of
alpha-fetoprotein. Annu Rev Med. 28:453–465. 1977. View Article : Google Scholar : PubMed/NCBI
|
|
27.
|
Niwa H, Miyazaki J and Smith AG:
Quantitative expression of Oct-3/4 defines differentiation,
dedifferentiation or self-renewal of ES cells. Nat Genet.
24:372–376. 2000. View
Article : Google Scholar : PubMed/NCBI
|
|
28.
|
Mitsui K, Tokuzawa Y, Itoh H, et al: The
homeoprotein Nanog is required for maintenance of pluripotency in
mouse epiblast and ES cells. Cell. 113:631–642. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
29.
|
Niwa H, Ogawa K, Shimosato D and Adachi K:
A parallel circuit of LIF signalling pathways maintains
pluripotency of mouse ES cells. Nature. 460:118–122. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
30.
|
Hazan RB, Qiao R, Keren R, Badano I and
Suyama K: Cadherin switch in tumor progression. Ann NY Acad Sci.
1014:155–163. 2004. View Article : Google Scholar
|
|
31.
|
Akiyama Y, Watkins N, Suzuki H, et al:
GATA-4 and GATA-5 transcription factor genes and potential
downstream antitumor target genes are epigenetically silenced in
colorectal and gastric cancer. Mol Cell Biol. 23:8429–8439. 2003.
View Article : Google Scholar
|
|
32.
|
Liu M, Lee DF, Chen CT, et al: IKKalpha
activation of NOTCH links tumorigenesis via FOXA2 suppression. Mol
Cell. 45:171–184. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
33.
|
Song Y, Washington MK and Crawford HC:
Loss of FOXA1/2 is essential for the epithelial-to-mesenchymal
transition in pancreatic cancer. Cancer Res. 70:2115–2125. 2010.
View Article : Google Scholar
|
|
34.
|
Zhou Q, Brown J, Kanarek A, Rajagopal J
and Melton DA: In vivo reprogramming of adult pancreatic exocrine
cells to beta-cells. Nature. 455:627–632. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
35.
|
Niakan KK, Ji H, Maehr R, et al: Sox17
promotes differentiation in mouse embryonic stem cells by directly
regulating extraembryonic gene expression and indirectly
antagonizing self-renewal. Genes Dev. 24:312–326. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
36.
|
Bai RY, Staedtke V, Lidov HG, Eberhart CG
and Riggins GJ: OTX2 represses myogenic and neuronal
differentiation in medulloblastoma cells. Cancer Res. 72:5988–6001.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
37.
|
Flores ER and Lozano G: The p53 family
grows old. Genes Dev. 26:1997–2000. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
38.
|
Kalisz M, Winzi M, Bisgaard HC and Serup
P: EVEN-SKIPPED HOMEOBOX 1 controls human ES cell differentiation
by directly repressing GOOSECOID expression. Dev Biol. 362:94–103.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
39.
|
Davidson NE and Sukumar S: Of Snail, mice,
and women. Cancer Cell. 8:173–174. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
40.
|
Holmes DI and Zachary IC: Vascular
endothelial growth factor regulates stanniocalcin-1 expression via
neuropilin-1-dependent regulation of KDR and synergism with
fibroblast growth factor-2. Cell Signal. 20:569–579. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
41.
|
Triozzi PL and Stevens VC: Human chorionic
gonadotropin as a target for cancer vaccines. Oncol Rep. 6:7–17.
1999.
|
|
42.
|
Gassmann P, Haier J, Schluter K, et al:
CXCR4 regulates the early extravasation of metastatic tumor cells
in vivo. Neoplasia. 11:651–661. 2009.PubMed/NCBI
|
|
43.
|
Leong KG and Gao WQ: The Notch pathway in
prostate development and cancer. Differentiation. 76:699–716. 2008.
View Article : Google Scholar
|
|
44.
|
Douville J, Beaulieu R and Balicki D:
ALDH1 as a functional marker of cancer stem and progenitor cells.
Stem Cells Dev. 18:17–25. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
45.
|
Ding L, Yuan C, Wei F, et al: Cisplatin
restores TRAIL apoptotic pathway in glioblastoma-derived stem cells
through up-regulation of DR5 and down-regulation of c-FLIP. Cancer
Invest. 29:511–520. 2011. View Article : Google Scholar
|
|
46.
|
Rajasekhar VK, Studer L, Gerald W, Socci
ND and Scher HI: Tumour-initiating stem-like cells in human
prostate cancer exhibit increased NF-kappaB signalling. Nat Commun.
2:1622011. View Article : Google Scholar : PubMed/NCBI
|
|
47.
|
Peinado H, Olmeda D and Cano A: Snail, Zeb
and bHLH factors in tumour progression: an alliance against the
epithelial phenotype? Nat Rev Cancer. 7:415–428. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
48.
|
Kirsh VA, Peters U, Mayne ST, et al:
Prospective study of fruit and vegetable intake and risk of
prostate cancer. J Natl Cancer Inst. 99:1200–1209. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
49.
|
Richman EL, Carroll PR and Chan JM:
Vegetable and fruit intake after diagnosis and risk of prostate
cancer progression. Int J Cancer. 131:201–210. 2012. View Article : Google Scholar : PubMed/NCBI
|