Introduction
Cell polarity is defined as asymmetrical cell shapes
and/or asymmetrical protein distributions and cell functions and
contributes to normal tissue integrity and development (1). Polarity complexes, formed by polarity
proteins, are required for the maintenance of cell polarity. Three
apical-basal polarity complexes have been identified to date. The
Crumbs (CRB) complex is formed by CRB (CRB1-3 in mammals), protein
associated with Lin seven 1 (PALS1) and PALS1-associated tight
junction protein (PATJ) (2–4). The
partitioning defective (PAR) complex comprises PAR3 and 6, and
atypical protein kinase C (aPKC). The CRB and PAR complexes are
localized in the apical membrane domain and promote
apical-membrane-domain identity (5,6).
Their function is antagonized by the basolaterally localized
Scribble complex. The Scribble complex is composed of lethal giant
larvae (Lgl), scribble (Scrib) and discs large (Dlg), which work
together to promote basolateral membrane identity (7).
Most human cancers originate from epithelial cells,
a cell type in which polarity proteins play crucial roles in
maintaining epithelial structures (8). Disruption of cell polarity is a
hallmark of cancer. Recent evidence indicates that the expression
of a number of polarity proteins is altered in human cancers. A
study has shown that polarity protein PAR6, a component of the PAR
complex, is overexpressed in breast cancer-derived cell lines and
advanced primary human breast cancers (9). Another component of the PAR complex,
aPKC protein, is also found to be increased in ovarian cancers and
non-small cell lung cancers (10,11).
However, loss of PAR3 can promote breast cancer metastasis
(12,13). PAR6 and aPKC may act as tumor
promoters while PAR3 may serve as a tumor suppressor (14). Numerous studies have shown that
components of the Scribble complex were correlated with cancers.
Lgl2, a homolog of Lgl, is downregulated in colorectal cancer and
loss of Lgl2 may be involved in metastasis (15), which suggest that Lgl2 may be a
tumor suppressor. Dlg1, a homolog of Dlg, is also a tumor
suppressor and loss of Dlg1 promotes the acquisition of
invasiveness (16). Interestingly,
the roles of the polarity protein Scrib in tumorigenesis seem to be
complicated: in some cases it induces cell migration, whereas in
others it is associated with inhibition of invasion and metastasis
(17–20). Although evidence is increasing on
involvement of deregulated polarity proteins in cancers, little is
currently known about the roles of the CRB (Drosophila),
especially the roles of CRB3, a homolog of the CRB.
CRB3 is widely expressed in epithelial tissues.
Previous experiments have suggested that CRB may act as a tumor
suppressor in Drosophila imaginal disc epithelium (21). Loss of CRB3 expression promotes
mammalian epithelial tumor progression, while restoration of CRB3
expression restrains cell migration and metastasis (22). These data suggest that CRB may
possess tumor suppressive potential not only in Drosophila
but also in mammalian cells. However, no study has been performed
on the role of deregulated CRB3 in progression toward malignancy in
human tumors. Renal cell carcinoma (RCC) accounts for 3% of all
malignant tumors. Patients with RCC have a high risk of relapse and
metastasis (23). Clear cell renal
cell carcinoma (ccRCC), a main histologic type of RCC, accounts for
a majority of kidney cancer-associated deaths (24,25).
The prognosis of the disease is difficult to predict due to lack of
effective clinical prognostic factors. In this study, we aimed to
investigate the role of CRB3 in human ccRCC. We found that CRB3
protein was weakly expressed in human ccRCC but strongly expressed
in the adjacent normal kidney tissues. This is also the first study
to investigate the relationships between the expression of CRB3 and
the clinicopathological features of ccRCC patients. We found that
negative CRB3 expression was correlated with shorter overall
survival (OS) in ccRCC patients. Altogether, our results showed
that CRB3 may be an independent favorable prognostic factor for
patients with ccRCC.
Materials and methods
Cell culture
All cell lines were obtained from Shanghai Institute
of Biochemistry and Cell Biology, Chinese Academy of Sciences
(Shanghai, China). According to the American Type Culture
Collection (ATCC) (Manassas, VA, USA), 786-O and 769-P were
classified as ccRCC cell lines and ACHN was classified as the RCC
cell line. OS-RC-2 was the RCC cell line and was maintained as
previously described (26). The
human embryonic kidney 293T (HEK 293T) cell line and Madin-Darby
canine kidney (MDCK) cell line were maintained in Dulbecco’s
modified Eagle’s medium (DMEM) supplemented with 10% fetal bovine
serum (FBS) and 1% penicillin-streptomycin (all from HyClone,
Logan, UT, USA). The ccRCC cell lines 786-O and 769-P and RCC cell
line ACHN were cultured in RPMI-1640 (HyClone) supplemented with
10% FBS and 1% penicillin-streptomycin. The cells were incubated in
5% CO2 at 37°C.
Antibodies
The rabbit polyclonal anti-CRB3 antibody was
purchased from Sigma-Aldrich which was used for
immunohistochemistry (IHC) (1:200 dilution, HPA013835; St. Louis,
MO, USA). The following antibodies were used for western blot
analysis: rabbit polyclonal anti-CRB3 antibody (1:200 dilution,
SC-292449; Santa Cruz Biotechnology, Inc., Santa Cruz, CA, USA),
mouse monoclonal anti-E-cadherin antibody (1:500 dilution, ab1416;
Abcam, Cambridge, MA, USA), mouse monoclonal anti-N-cadherin
antibody (1:1,000 dilution, 610920; BD Biosciences, Bedford, MA,
USA), rabbit polyclonal anti-vimentin antibody (1:2,000 dilution,
10366-1-AP) and mouse monoclonal anti-GAPDH antibody (1:5,000
dilution, HRP-60004) (both from Wuhan Sanying Biotechnology, Hubei,
China). Anti-rabbit and anti-mouse HRP-conjugated secondary
antibodies were purchased from Santa Cruz Biotechnology, Inc.
(1:3,000 dilution, SC-2004 and SC-2005).
Transfection
The 786-O and HEK 293T cells were seeded in a 6-cm
dish one day before transfection. Then the 786-O cells were
transfected with GV168-CRB3 plasmid using TurboFect Transfection
Reagent (Thermo Scientific, Rockford, IL, USA) to overexpress CRB3
according to the manufacturer’s instructions. GV168-CRB3 plasmid
was purchased from Genechem Co., Ltd. (Shanghai, China). The HEK
293T cells were transfected with siRNA using Lipofectamine 2000
(Life Technologies, Carlsbad, CA, USA) to knock down CRB3
expression according to the manufacturer’s instructions. The siRNA
was purchased from Shanghai GenePharma Co., Ltd. (Shanghai, China).
Target sequences of oligonucleotides used were as follows: CRB3,
5′-UGG CAC UGU UGG UGC GGA ATT-3′; control (non-targeting), 5′-UUC
UCC GAA CGU GUC ACG UTT-3′. The cells were harvested 48 h after
transfection.
Tissue microarrays, patients and
followup
The human study was approved by the Ethics Committee
on Human Research of the First Affiliated Hospital of Xi’an
Jiaotong University (Shaanxi, China). Written informed consent was
obtained from all patients. Tissue microarrays (TMAs) were obtained
from Shanghai Biochip Co., Ltd. (Shanghai, China). The TMAs
contained 142 pairs of ccRCC and adjacent normal kidney tissues.
Six pairs of tissue samples were excluded from the study due to
lack of sufficient tumor tissue in the samples or fixation
artifacts. The remaining 136 pairs of tissue samples consisted of
tissues from 85 male and 51 female patients with a mean age of 59
years (range, 29–82 years). One-hundred and five patients were
followed up for 2–74 months, with a median follow-up of 63 months.
The follow-up data were summarized by the end of September, 2012.
OS was defined as the period from the surgery to death or to the
last known follow-up. Seventy-seven patients were still alive by
the end of the follow-up.
Immunohistochemistry and scoring
The paraffin-embedded TMA slides were deparaffinized
in xylene and rehydrated in ethanol with decreasing concentrations
(100, 95, 90 and 85%, 5 min each). After the slides were washed
five times (3 min each time) with phosphate-buffered saline (PBS),
3% H2O2 was used to block the endogenous
peroxidase activity for 20 min. After being rinsed, the slides were
placed in 0.01 M sodium citrate in a microwave oven (pH 6.0, 100 W,
6 min; 50 W, 13 min) for antigen retrieval. Then slides were cooled
at room temperature and added with goat serum to block non-specific
binding sites for 15 min. Without being washed, the slides were
incubated with the primary antibody in a moist box at 4°C
overnight. After being rinsed, the slides were incubated with the
streptavidin-peroxidase staining system kit according to the
manufacturer’s instructions (SP-9001; Beijing Zhongshan Golden
Bridge Biotechnology Co., Ltd., Beijing, China). Finally, the
slides were stained by using the 3,3′-diaminobenzidine (DAB) liquid
chromogen substrate kit (ZLI-9017; Beijing Zhongshan Golden Bridge
Biotechnology Co., Ltd.) and counterstained with hematoxylin,
followed by observation with Leica Microsystems GmbH (SCN 400;
Mannheim, Germany). The results of IHC were evaluated individually
in a blinded manner by three investigators who had no access to the
clinical or clinicopathological status of the specimens. The
intensity of immunohistochemical staining was scored as 0
(negative), 1 (weakly positive), 2 (moderately positive) or 3
(strongly positive). The extent of immunohistochemical staining was
assessed according to the percentage of positive cells: 0
(negative), 1 (1–25%), 2 (26–50%), 3 (51–75%) and 4 (76–100%). The
staining score for each field was calculated as the product of the
intensity and extent of the staining. The final staining score for
each sample was the mean of the staining scores of three randomly
selected fields. The expression level was considered negative if
the final score was 0–2 and positive if the final score was 3–12.
The expression level was considered low if the final score was 3–5
and high if the final score was 6–12.
CRB3 is not expressed in the adipose tissue
according to the data provided in EBI Expression Atlas database
(27). We performed IHC of the
adipose tissue using the primary antibody rabbit polyclonal
anti-CRB3 antibody and found that CRB3 protein was not expressed in
the adipose tissue. CRB3 was expressed in the ccRCC tissues
incubated with rabbit polyclonal anti-CRB3 antibody, and not
expressed in the ccRCC tissues incubated with CRB3 recombinant
protein and rabbit polyclonal anti-CRB3 antibody. Therefore, the
ccRCC tissue incubated with CRB3 recombinant protein and primary
antibody, and the adipose tissue were used as negative controls in
the study. Tissues incubated with PBS in place of the primary
antibody were also used as negative control to verify the
specificity of the IHC results.
Real-time polymerase chain reaction
analysis
Total RNA was extracted from cells by using the
E.Z.N.A.® Total RNA kit I (Omega Bio-Tek, Inc.,
Norcross, GA, USA) according to the manufacturer’s instructions.
Reverse transcription was performed by using the PrimeScript™ RT
Master Mix [Takara Biotechnology (Dalian) Co., Ltd., Liaoning,
China] according to the manufacturer’s instructions. Real-time
polymerase chain reaction (real-time PCR) was performed at an
annealing temperature of 60°C using the SYBR-Green I kit [Takara
Biotechnology (Dalian) Co., Ltd.]. The following primers were used
for real-time PCR reaction: CRB3 forward, 5′-CTT CTG CAA ATG AGA
ATA GCA CTG T-3′ and reverse, 5′-GAA GAC CAC GAT GAT AGC AGT GA-3′;
GAPDH forward, 5′-CTC CTC CAC CTT TGA CGC TG-3′ and reverse, 5′-TCC
TCT TGT GCT CTT GCT GG-3′. Results were analyzed using the
comparative Ct method with the housekeeping gene
glyceraldehyde-3-phosphate dehydrogenase (GAPDH) as an internal
control.
Western blot analysis
Proteins were extracted from cells using RIPA lysis
buffer (50 mM Tris-HCl pH 7.4, 150 mM NaCl, 1% Triton X-100, 0.1%
SDS, 1 mM EDTA, and 1% sodium deoxycholate) supplemented with a
protease inhibitor cocktail (Roche Diagnostics, Ltd., Basel,
Switzerland). Cell lysates were centrifuged at 12,000 rpm for 20
min at 4°C. Proteins were separated by 12% SDS-polyacrylamide gel
electrophoresis (SDS-PAGE) and transferred onto polyvinyl
difluoride (PVDF) membranes (0.22 μm; Millipore, Billerica, MA,
USA) by electroblotting. The membranes were blocked with 5% skimmed
milk containing Tris-buffered saline and 0.1% Tween-20 (TBST) for 2
h, followed by incubation with the primary antibody at 4°C
overnight. After the membranes were washed with TBST, they were
incubated with the secondary antibody. Immunoreactions were
visualized using the chemiluminescence reagent (Millipore)
according to the manufacturer’s instructions. GAPDH was used as a
protein loading control.
Wound-healing assay
786-O and HEK 293T cells were transfected with
GV168-CRB3 plasmid and siRNA, respectively. Cells were harvested 24
h after transfection and plated on 24-well cell culture plates.
After 12 h, scratch wounds were created manually in the monolayer
of cells with a P200 pipette tip, washed twice with PBS, and
incubated with fresh medium. The plates were photographed at the
identical locations 0 and 24 h after incubation by Phase Contrast
Microscope (20x; Nikon, Tokyo, Japan). Migration was determined by
comparing the two sets of images.
Cell migration and invasion assays
Migration and invasion assays were performed in
24-well Transwells with polycarbonate filters (8 μm pore size;
Corning, Inc., Corning, NY, USA). The Transwell for the invasion
assay was coated with Matrigel (1:5 dilution; BD Biosciences),
while the Transwell for the migration assay was not coated. 786-O
and HEK 293T cells were transfected with GV168-CRB3 plasmid and
siRNA, respectively. The cells were harvested 24 h after
transfection. 786-O cells were trypsinized and suspended with
RPMI-1640 without FBS. 786-O cells, 2×104 (for migration
assay) or 2×105 (for invasion assay), were added to the
upper wells and 600 μl RPMI-1640 medium containing 10% FBS was
added to the lower wells. Cells in the wells were incubated in 5%
CO2 at 37°C for 24 h for migration assay and 48 h for
invasion assay. After incubation, cells in the upper wells were
gently removed by scrubbing, fixed in 95% ethanol for 15 min and
stained with 0.4% crystal violet for 30 min. The number of
transmembrane cells was calculated as the mean of the cell numbers
in three randomly selected microscopic fields (20x; Nikon).
Migration and invasion assays of HEK 293T cells were performed
following the same procedure except that HEK 293T cells were
maintained with the DMEM medium and 5×104 cells were
added for migration assay and 5×105 cells for invasion
assay.
Statistical analysis
Statistical analyses were performed using the
Statistical Package for the Social Sciences (SPSS) software
(version 13.0; SPSS Inc., Chicago, IL, USA). The χ2 test
was used to investigate the possible relationships between CRB3
expression and clinicopathological characteristics. The
Mann-Whitney U test was used to compare differences between CRB3
protein expression in paired ccRCC and adjacent normal kidney
tissues. Survival analysis was done by using the Kaplan-Meier curve
and the log-rank test. Multivariate analysis was performed using
the Cox proportional hazard regression model to explore the effects
of CRB3 expression and ccRCC clinicopathological variables on
survival. In cellular studies, values are expressed as mean ± SEM.
Comparisons between two groups were conducted using the Student’s
t-test. All experiments were done in triplicate. All statistical
tests were two-sided. P<0.05 was considered statistically
significant.
Results
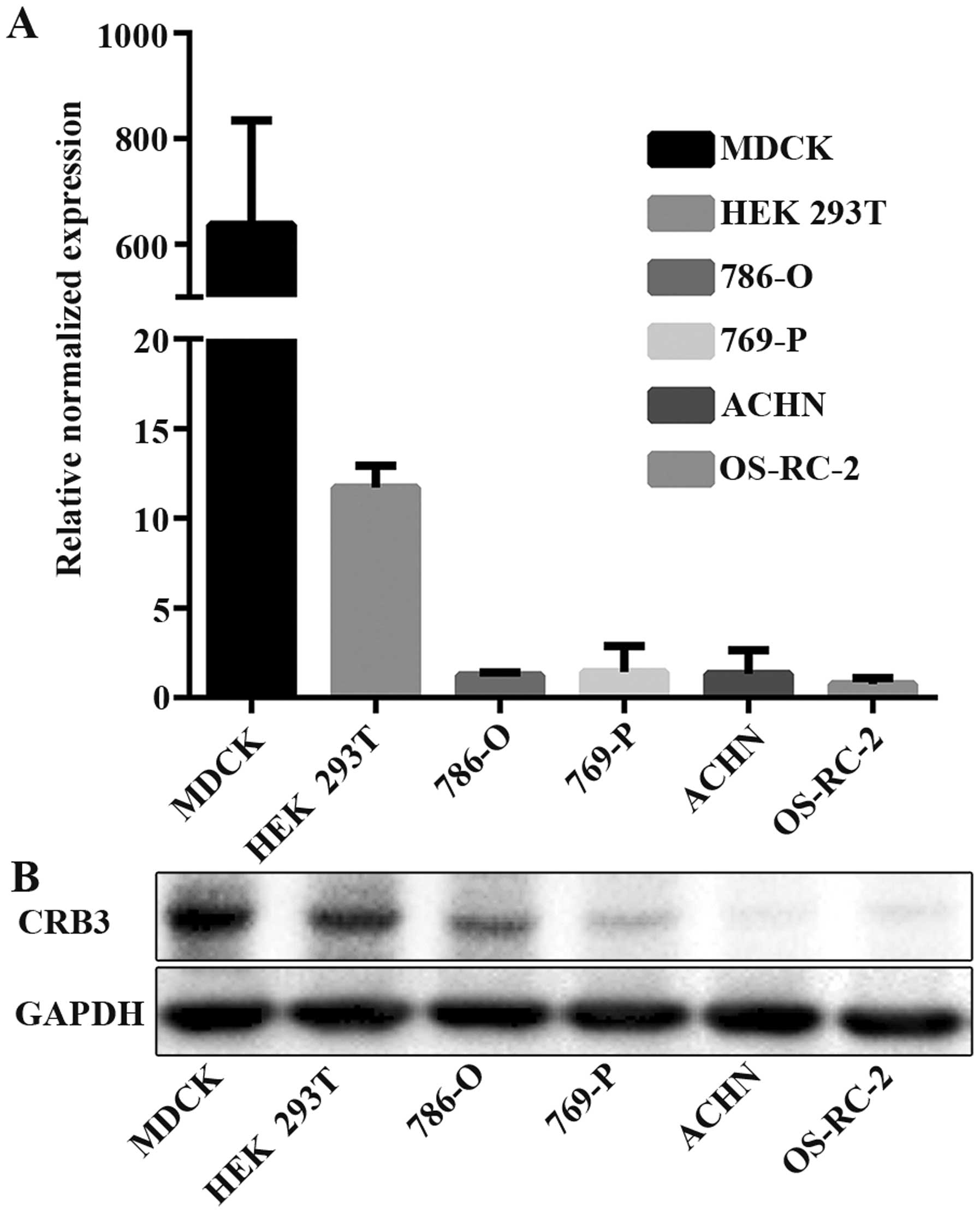
Low CRB3 expressions in ccRCC and RCC
cell lines
We first examined CRB3 mRNA and protein expressions
in the normal canine kidney epithelial cell line MDCK, HEK 293T
cell line, ccRCC cell lines 786-O and 769-P, and the RCC cell lines
ACHN and OS-RC-2. Real-time PCR and western blot analyses showed
that MDCK had the highest mRNA and protein expression levels of
CRB3 among the cell lines (Fig. 1A and
B). The mRNA and protein expressions of CRB3 in ccRCC and RCC
cell lines were relatively lower than those in MDCK.
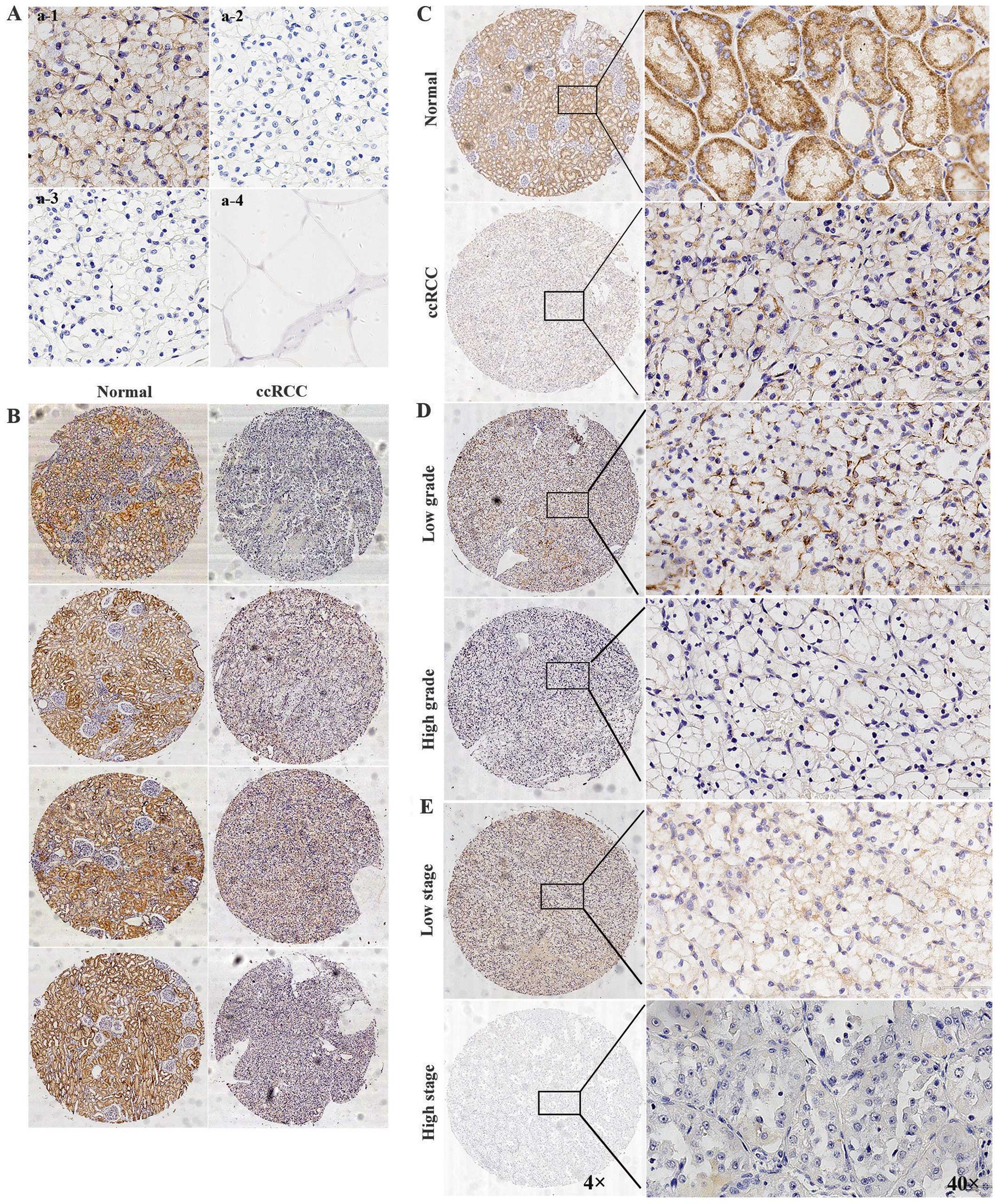
Distributions of CRB3 protein in paired
ccRCC and adjacent normal kidney tissues
Immunohistochemical analysis of TMAs was performed
to investigate whether CRB3 was differently expressed in paired
ccRCC and adjacent normal kidney tissues. The TMAs contained 136
pairs of ccRCC and adjacent normal kidney tissue samples. Due to
the small sample size, low and high CRB3 expressions were
classified as positive expressions. The clinicopathological
features of the ccRCC patients and their CRB3 expression levels are
listed in Table I. The specificity
of the primary antibody rabbit polyclonal anti-CRB3 antibody was
validated by using three negative controls (Fig. 2A). The results showed that CRB3 was
not expressed in the adipose tissue and the ccRCC tissues incubated
with CRB3 recombinant protein or PBS, which indicated that the
primary antibody used for IHC was specific and the IHC results are
sound. We found that CRB3 protein was predominantly expressed in
the membrane and cytoplasm of renal tubular epithelial cells
(Fig. 2B and C). CRB3 protein was
strongly expressed in adjacent normal kidney tissues and weakly
expressed in ccRCC tissues (Fig. 2B
and C). Sixty-nine percent of ccRCC tissues showed complete
loss of CRB3 expressions (n=136, 69%, P<0.001, Table II). The expression level of CRB3
was significantly correlated with histologic grade and TNM stage
(P<0.05, Fig. 2D and E,
Table I), but not with age,
gender, tumor size and site, and capsule invasion (Table I).
 | Table IRelationships between CRB3 expression
and clinicopathologic parameters of ccRCC patients. |
Table I
Relationships between CRB3 expression
and clinicopathologic parameters of ccRCC patients.
| | Expression
levels | | |
|---|
| |
| | |
|---|
| Variables | No. | Neg. | Pos. | χ2 | P |
|---|
| Age (years) | | | | 1.741 | 0.187 |
| <60 | 76 | 49 | 27 | | |
| ≥60 | 60 | 45 | 15 | | |
| Gender | | | | 0.230 | 0.632 |
| Male | 85 | 60 | 25 | | |
| Female | 51 | 34 | 17 | | |
| Histologic
grade | | | | 7.761 | 0.005b |
| I–II | 98 | 61 | 37 | | |
| III–IV | 38 | 33 | 5 | | |
| TNM stagec | | | | 5.378 | 0.020a |
| 1–2 | 110 | 70 | 40 | | |
| 3–4 | 20 | 18 | 2 | | |
| Tumor size
(cm)d | | | | 0.897 | 0.344 |
| <6.0 | 73 | 53 | 20 | | |
| ≥6.0 | 63 | 41 | 22 | | |
| Tumor site | | | | 0.431 | 0.512 |
| Left | 72 | 48 | 24 | | |
| Right | 64 | 46 | 18 | | |
| Capsule
invasion | | | | 0.197 | 0.657 |
| Neg. | 113 | 79 | 34 | | |
| Pos. | 23 | 15 | 8 | | |
| Table IICRB3 protein expressions in ccRCC and
adjacent normal kidney tissues. |
Table II
CRB3 protein expressions in ccRCC and
adjacent normal kidney tissues.
| | Expression
levels | | |
|---|
| |
| | |
|---|
| Variables | No. | Neg. | Low | High | Z | P |
|---|
| Normala | 136 | 11 | 48 | 77 | −10.175 | <0.001 |
| ccRCC | 136 | 94 | 25 | 17 | | |
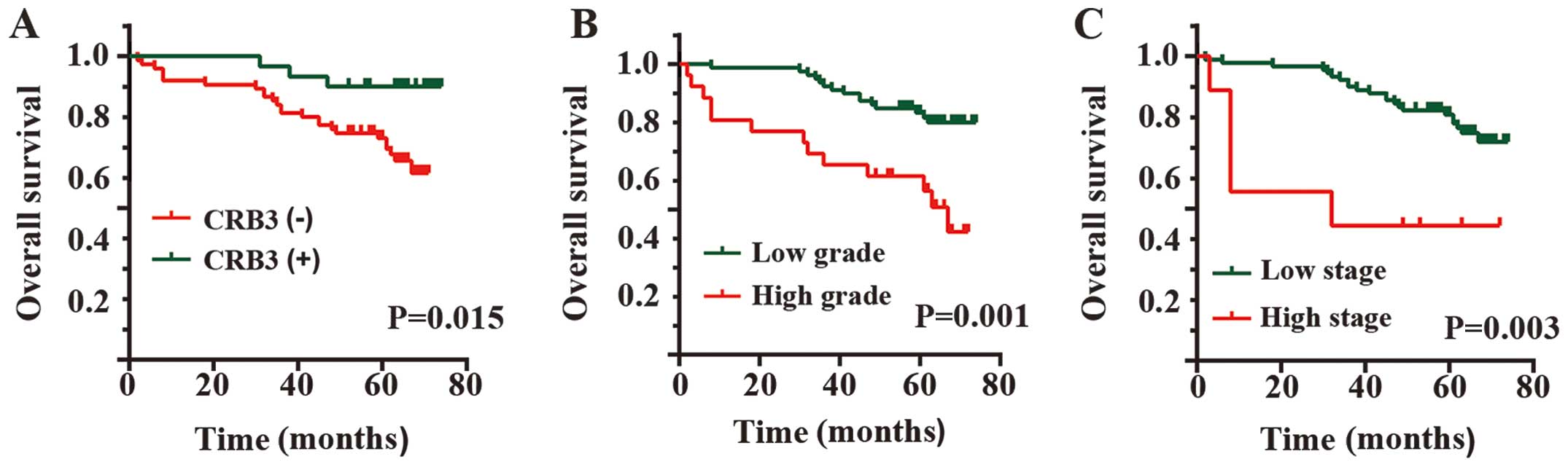
Prognostic significance of CRB3
The significant differences in the CRB3 expression
between the pairs of ccRCC and adjacent normal kidney tissues
inspired us to consider whether CRB3 could be used as a prognostic
predictor for ccRCC patients. The association of CRB3 expression
with OS was assessed by univariate analysis using the Kaplan-Meier
curve and the log-rank test. No significant difference was detected
in OS among patients with negative, low and high CRB3 expressions
(data not shown). Due to the small sample size, low and high CRB3
expressions were classified as positive expressions. Of the 105
patients followed up for survival, 30 patients showed positive
expression of CRB3 while the other 75 patients showed negative
expression. Patients with negative CRB3 expressions had a
significantly shorter OS than patients with positive expression
(P=0.015, log-rank test, Fig. 3A).
The result indicated that CRB3 may be a prognostic indicator for
patients with ccRCC. Univariate analysis also showed that patients
with different histologic grades or TNM stages showed significantly
different OS (data not shown). Here, due to the insufficient sample
size, we divided the histologic grades or TNM stages into low
histologic grade or TNM stage (histologic grade I-II or TNM stage
1–2) and high histologic grade or TNM stage (histologic grade
III-IV or TNM stage 3–4). Log-rank test showed that high histologic
grade or TNM stage was significantly associated with shorter OS
(histologic grade I-II vs. III-IV, P=0.001, Fig. 3B; TNM stage 1–2 vs. 3–4, P=0.003,
Fig. 3C).
Multivariate analysis was performed using the Cox
proportional hazard regression model to further determine the
effects of CRB3 expression and clinicopathological parameters on
the prognosis of ccRCC patients. The multivariate Cox proportional
hazard regression model mainly considered CRB3 expression and
clinicopathological parameters, including age, gender, histologic
grade, TNM stage, tumor size and site, and capsule invasion.
Results showed that positive CRB3 expression was linked to longer
OS [B=−1.288, Exp(B)=0.276, P=0.037, Table III], which indicated that
positive CRB3 expression may be an independent favorable prognostic
factor for OS and may be used to predict OS of ccRCC patients.
Table III also shows that
histologic grade may be a hazardous factor [B=1.250, Exp(B)=3.490,
P=0.001, Table III]. Age,
gender, TNM stage, tumor size and site, and capsule invasion were
not independent indicators for clinical outcome (Table III).
| Table IIIMultivariate Cox proportional hazard
regression analysis of overall survival in ccRCC patients.a |
Table III
Multivariate Cox proportional hazard
regression analysis of overall survival in ccRCC patients.a
| Variables | B | Exp(B) | 95% CI | P |
|---|
| Age | | | | |
| <60 | 0.305 | 1.356 | 0.590–3.117 | 0.473 |
| ≥60 | | | | |
| Gender | | | | |
| Female | 0.115 | 1.122 | 0.503–2.502 | 0.779 |
| Male | | | | |
| Histologic
grade | | | | |
| I-II | 1.250 | 3.490 | 1.632–7.464 | 0.001 |
| III-III | | | | |
| TNM stage | | | | |
| 1–2 | 0.577 | 1.780 | 0.420–7.539 | 0.434 |
| 3–4 | | | | |
| Tumor size | | | | |
| <6.0 | 0.398 | 1.489 | 0.606–3.661 | 0.386 |
| ≥6.0 | | | | |
| Tumor site | | | | |
| Left | 0.161 | 1.175 | 0.495–2.787 | 0.714 |
| Right | | | | |
| Capsule
invasion | | | | |
| No | 0.306 | 1.358 | 0.237–7.788 | 0.731 |
| Yes | | | | |
| CRB3 expression
level | | | | |
| Negative | −1.288 | 0.276 | 0.082–0.923 | 0.037 |
| Positive | | | | |
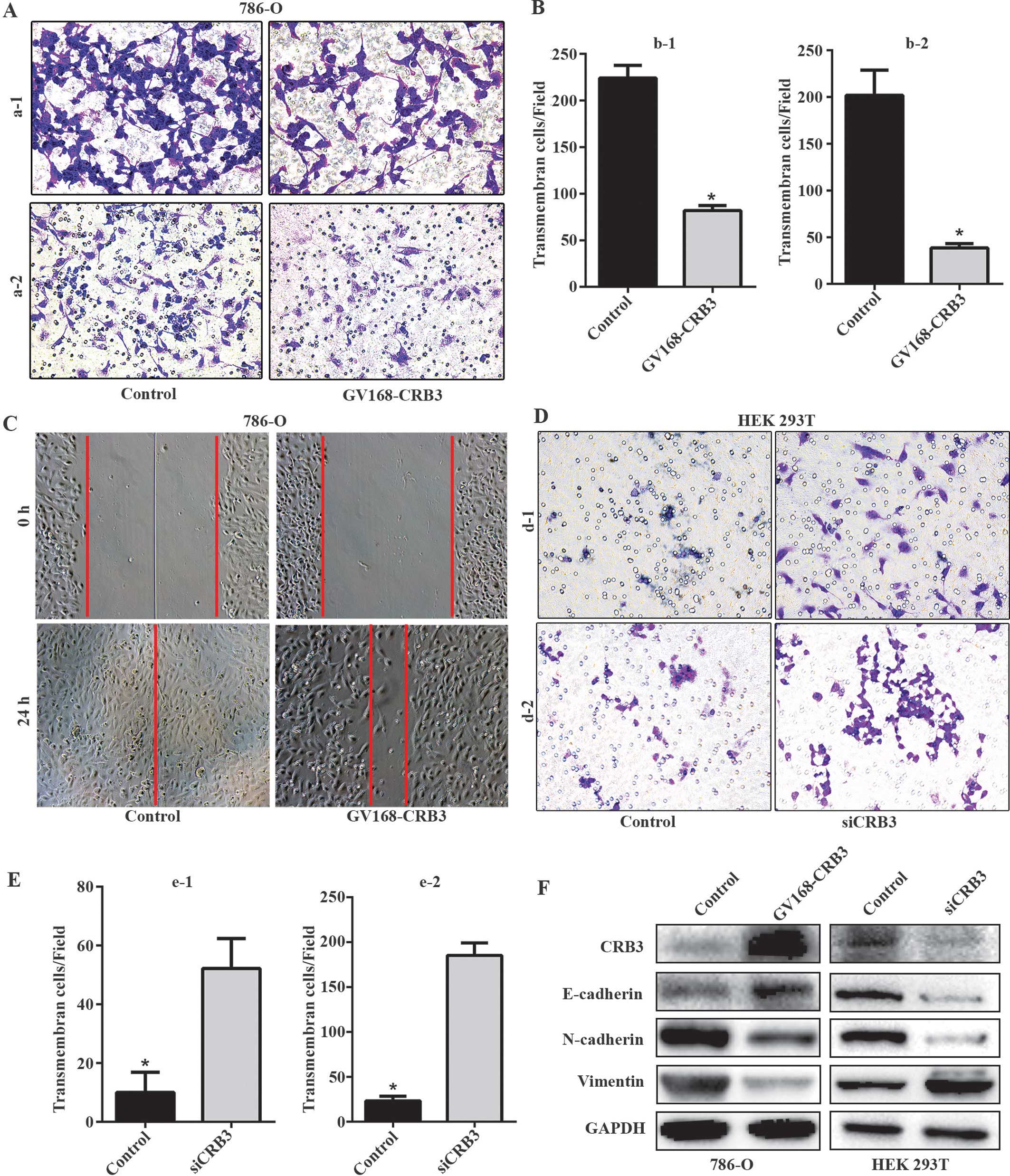
Roles of CRB3 in regulation of invasion
and migration
Invasion and metastasis are the hallmarks of
malignant tumor progression and the main causes of deaths in
cancer. Here, cell migration, invasion and wound-healing assays
were conducted to analyze the invasive and metastatic behavior
in vitro. Since the 786-O cell line had a low protein
expression of CRB3 and is a suitable host for transfection, 786-O
cells were transfected with GV168-CRB3 plasmid to overexpress CRB3.
Cell migration and invasion assays revealed that transfected 786-O
cells showed reduced migration and invasion, when compared with
control cells (Fig. 4A and B).
Wound-healing assay showed that the wound of control cells
disappeared, but the transfected cells failed to efficiently
migrate to fill the void, indicating that the migration rate of
786-O cells was reduced after transfection with GV168-CRB3 plasmid
(Fig. 4C). The results indicated
that CRB3 overexpression significantly inhibited the migration and
invasion of 786-O cells.
To investigate the effect of the loss of CRB3 on
cell invasion and migration, HEK 293T cells were transfected with
siRNA to knock down the CRB3 expression. The transfected HEK 293T
cells showed increased migration and invasion, when compared with
control cells, indicating that loss of CRB3 expression may promote
the migration and invasion of HEK 293T cells (Fig. 4D and E). Unfortunately,
wound-healing assay was not successful for HEK 293T cells, probably
due to the poor anchorage-dependent growth of the cells.
To investigate the roles of CRB3 in
epithelial-mesenchymal transition (EMT), we examined the expression
levels of the mesenchymal markers vimentin and N-cadherin and the
epithelial marker E-cadherin in 786-O cells transfected with
GV168-CRB3 plasmid and HEK 293T cells transfected with siRNA.
Western blot analysis revealed an increased E-cadherin expression
and concomitant decreased vimentin and N-cadherin expressions in
transfected 786-O cells (Fig. 4F).
However, transfection with siRNA induced decreased E- and
N-cadherin expressions and an increased vimentin expression in HEK
293T cells (Fig. 4F). The
expression changes of the mesenchymal and epithelial markers
indicated that CRB3 may be associated with EMT.
Discussion
Loss of cell polarity has been widely assumed to be
a key step in cancer progression. Most human cancers, including
ccRCC, originate from epithelial cells, a cell type in which
polarity proteins play crucial roles in maintaining epithelial
structures (8). The CRB polarity
complex, formed by CRB, PATJ and PALS1, participates in epithelial
polarity maintenance, apical membrane formation, and tissue
morphogenesis (14,28). Previous studies have reported the
association between disordered expressions of CRB and tumor
progression in Drosophila and non-human mammalian cells.
These reports inspired us to consider whether the deregulated CRB
expression is associated with progression toward malignancy in
human tumors, especially human ccRCC.
CRB3, one of the human CRB, is a small transmembrane
protein with a conserved cytoplasmic domain (29,30).
We found that CRB3 was strongly expressed in normal kidney
epithelial cells but weakly expressed in ccRCC and RCC cells. This
led us to investigate the expression pattern of CRB3 in human tumor
tissues. It has been found that CRB acts as a tumor suppressor in
Drosophila imaginal disc epithelium (21). Loss of CRB3 expression promotes
mammalian epithelial tumor progression while restoration of CRB3
expression restrains cell migration and metastasis (22). We found that ccRCC tissues had an
apparently lower protein expression of CRB3 than adjacent normal
kidney tissues, which is consistent with the above findings. CRB3
was predominantly expressed in the membrane and cytoplasm of renal
tubular epithelial cells. CRB3 was detected in the cytoplasm
probably because it is under dynamic intracellular trafficking
control (31,32) and in dynamic cycling and
degradation processes. We also investigated the link between the
expression of CRB3 and the prognosis of ccRCC patients, aiming to
find an effective clinical prognostic marker for human ccRCC.
Patients with negative CRB3 expression showed a significantly
shorter OS than patients with positive expression. This result
indicated that the positive CRB3 expression may be an independent
favorable factor for OS. Due to the limited numbers of the tissue
samples and histologic types, a study using a larger sample size is
needed to verify these findings. 786-O cells transfected with
GV168-CRB3 plasmid overexpressed CRB3 and showed remarkably lower
migration and invasion, when compared with the control cells. HEK
293T cells transfected with siRNA had a decreased CRB3 expression
and showed increased migration and invasion, when compared with
control cells. The results indicated that loss of CRB3 expression
may promote the cell migration and invasion. Since invasion and
metastasis can lead to deaths of cancer patients, these findings
may explain why ccRCC patients with negative CRB3 expressions had
poor prognosis.
EMT is characterized by loss of cell polarity,
enhanced cell motility and increased cell invasion (33). CRB3 expression is mediated by two
EMT inducers, namely, ZEB1 and Snail, which suggests that CRB3
expression is associated with EMT (15,34,35).
We also found that CRB3 may be associated with EMT. Loss of CRB3 in
HEK 293T cells decreased the E-cadherin expression and increased
the vimentin expression. On the other hand, overexpression of CRB3
in 786-O cells increased the E-cadherin expression and decreased
the vimentin expression. Dysfunctions of vimentin and E-cadherin
have been considered hallmarks of EMT. Vimentin is involved in
epithelial cell migration and invasion (36). Reduction in E-cadherin increases
cell mobility and promotes tumor cell invasion; increase in
E-cadherin has the reverse role (37–39).
The changes in the expression of EMT markers may be the causes of
altered cell migration and invasion. Thus, loss of CRB3 induced
E-cadherin reduction and vimentin increase, which may lead to
increase cell migration and invasion. Advanced cancers often
undergo a partial or full EMT, which disrupts epithelial junction
formation and may contribute to invasion and metastasis, the
hallmarks of malignant tumor progression and the main cause of
deaths in cancer (33). Patients
with negative CRB3 expression had a significantly shorter OS than
patients with positive expressions. Therefore, loss of CRB3 and EMT
may contribute to invasion and metastasis and cause the poor
prognosis of the ccRCC patients. Interestingly, loss of CRB3
induced inconsistent changes of N-cadherin expression in 786-O and
HEK 293T cells. The cause of this disparity was not clear. Further
studies are needed to solve this interesting problem.
Recent studies have also found that CRB3 is involved
in tight junction formation in mammalian epithelial cells and that
loss of CRB3 expression leads to tight junction defects (30,40,41).
Acquisition of tumorigenesis due to loss of CRB3 is associated with
deficient tight junction formation and restoration of CRB3
expression restores junctions and polarity while suppressing
migration and metastasis (22).
Our results showed that overexpression of CRB3 restrained invasion
and migration of 786-O cells. E-cadherin, essential for appropriate
tight junction formation (42),
was also increased in 786-O cells with CRB3 overexpression. These
observations and data may suggest that CRB3 overexpression
inhibited migration and invasion probably due to the proper tight
junction formation. Further studies are needed to reveal the
precise mechanism underlying the role of the increased CRB3
expression in inhibition of tumor migration and invasion.
Taken together, this study suggested that expression
of CRB3 may be used as an independent favorable prognostic factor
for patients with ccRCC. The identification of CRB3 as a prognostic
factor may contribute to selection of high-risk patients for
therapy, making it possible to improve the clinical outcome of the
patients. It will be of great interest to investigate the roles of
CRB3 expression in other human tumors and the precise molecular
mechanisms underlying the functions of CRB3.
Acknowledgements
This study was financially supported by Grants from
the National Natural Science Foundation of China (no. 81272342;
Beijing, China) and Program for New Century Excellent Talents in
University of the Ministry of Education of China (no. NCET-10-0677;
Beijing, China) to Dr Peijun Liu.
References
|
1
|
Nelson WJ: Adaptation of core mechanisms
to generate cell polarity. Nature. 422:766–774. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Tepass U, Theres C and Knust E: Crumbs
encodes an EGF-like protein expressed on apical membranes of
Drosophila epithelial cells and required for organization of
epithelia. Cell. 61:787–799. 1990. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Tepass U and Knust E: Crumbs and stardust
act in a genetic pathway that controls the organization of
epithelia in Drosophila melanogaster. Dev Biol. 159:311–326. 1993.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Bhat MA, Izaddoost S, Lu Y, Cho KO, Choi
KW and Bellen HJ: Discs Lost, a novel multi-PDZ domain protein,
establishes and maintains epithelial polarity. Cell. 96:833–845.
1999. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Margolis B and Borg JP: Apicobasal
polarity complexes. J Cell Sci. 118:5157–5159. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Suzuki A and Ohno S: The PAR-aPKC system:
lessons in polarity. J Cell Sci. 119:979–987. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Bilder D: Epithelial polarity and
proliferation control: links from the Drosophila neoplastic tumor
suppressors. Genes Dev. 18:1909–1925. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Assémat E, Bazellières E,
Pallesi-Pocachard E, Le Bivic A and Massey-Harroche D: Polarity
complex proteins. Biochim Biophys Acta. 1778:614–630. 2008.
View Article : Google Scholar
|
|
9
|
Nolan ME, Aranda V, Lee S, et al: The
polarity protein Par6 induces cell proliferation and is
overexpressed in breast cancer. Cancer Res. 68:8201–8209. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Eder AM, Sui X, Rosen DG, et al: Atypical
PKCiota contributes to poor prognosis through loss of apical-basal
polarity and cyclin E overexpression in ovarian cancer. Proc Natl
Acad Sci USA. 102:12519–12524. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Regala RP, Weems C, Jamieson L, et al:
Atypical protein kinase C iota is an oncogene in human non-small
cell lung cancer. Cancer Res. 65:8905–8911. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Xue B, Krishnamurthy K, Allred DC and
Muthuswamy SK: Loss of Par3 promotes breast cancer metastasis by
compromising cell-cell cohesion. Nat Cell Biol. 15:189–200. 2013.
View Article : Google Scholar
|
|
13
|
McCaffrey LM, Montalbano J, Mihai C and
Macara IG: Loss of the Par3 polarity protein promotes breast
tumorigenesis and metastasis. Cancer cell. 22:601–614. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Ellenbroek SI, Iden S and Collard JG: Cell
polarity proteins and cancer. Semin Cancer Biol. 22:208–215. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Spaderna S, Schmalhofer O, Wahlbuhl M, et
al: The transcriptional repressor ZEB1 promotes metastasis and loss
of cell polarity in cancer. Cancer Res. 68:537–544. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Sandoval GJ, Graham DB, Gmyrek GB, et al:
Novel mechanism of tumor suppression by polarity gene discs large 1
(DLG1) revealed in a murine model of pediatric B-ALL. Cancer
Immunol Res. 1:426–437. 2013. View Article : Google Scholar
|
|
17
|
Etienne-Manneville S: Polarity proteins in
migration and invasion. Oncogene. 27:6970–6980. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Dow LE and Humbert PO: Polarity regulators
and the control of epithelial architecture, cell migration, and
tumorigenesis. Int Rev Cytol. 262:253–302. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Martin-Belmonte F and Perez-Moreno M:
Epithelial cell polarity, stem cells and cancer. Nat Rev Cancer.
12:23–38. 2011.PubMed/NCBI
|
|
20
|
Dow LE, Elsum IA, King CL, Kinross KM,
Richardson HE and Humbert PO: Loss of human Scribble cooperates
with H-Ras to promote cell invasion through deregulation of MAPK
signalling. Oncogene. 27:5988–6001. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Ling C, Zheng Y, Yin F, et al: The apical
transmembrane protein Crumbs functions as a tumor suppressor that
regulates Hippo signaling by binding to Expanded. Proc Natl Acad
Sci USA. 107:10532–10537. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Karp CM, Tan TT, Mathew R, et al: Role of
the polarity determinant crumbs in suppressing mammalian epithelial
tumor progression. Cancer Res. 68:4105–4115. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Athar U and Gentile TC: Treatment options
for metastatic renal cell carcinoma: a review. Can J Urol.
15:3954–3966. 2008.PubMed/NCBI
|
|
24
|
Cohen HT and McGovern FJ: Renal-cell
carcinoma. N Engl J Med. 353:2477–2490. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Kaelin WG Jr: Treatment of kidney cancer:
insights provided by the VHL tumor-suppressor protein. Cancer.
115(Suppl 10): S2262–S2272. 2009. View Article : Google Scholar
|
|
26
|
Kinouchi T, Kotake T, Mori Y and Abe T:
Human renal cell carcinoma: establishment and characterization of a
new cell line (OS-RC-2). In Vitro Cell Dev Biol. 21:195–199. 1985.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Fagerberg L, Hallström BM, Oksvold P, et
al: Analysis of the human tissue-specific expression by genome-wide
integration of transcriptomics and antibody-based proteomics. Mol
Cell Proteomics. 13:397–406. 2014. View Article : Google Scholar
|
|
28
|
Bulgakova NA and Knust E: The Crumbs
complex: from epithelial-cell polarity to retinal degeneration. J
Cell Sci. 122:2587–2596. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Makarova O, Roh MH, Liu CJ, Laurinec S and
Margolis B: Mammalian Crumbs3 is a small transmembrane protein
linked to protein associated with Lin-7 (Pals1). Gene. 302:21–29.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Lemmers C, Michel D, Lane-Guermonprez L,
et al: CRB3 binds directly to Par6 and regulates the morphogenesis
of the tight junctions in mammalian epithelial cells. Mol Biol
Cell. 15:1324–1333. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Lu H and Bilder D: Endocytic control of
epithelial polarity and proliferation in Drosophila. Nat Cell Biol.
7:1232–1239. 2005. View
Article : Google Scholar : PubMed/NCBI
|
|
32
|
Roeth JF, Sawyer JK, Wilner DA and Peifer
M: Rab11 helps maintain apical crumbs and adherens junctions in the
Drosophila embryonic ectoderm. PLoS One. 4:e76342009. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Thiery JP and Sleeman JP: Complex networks
orchestrate epithelial-mesenchymal transitions. Nat Rev Mol Cell
Biol. 7:131–142. 2006. View
Article : Google Scholar : PubMed/NCBI
|
|
34
|
Aigner K, Dampier B, Descovich L, et al:
The transcription factor ZEB1 (deltaEF1) promotes tumour cell
dedifferentiation by repressing master regulators of epithelial
polarity. Oncogene. 26:6979–6988. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Whiteman EL, Liu CJ, Fearon ER and
Margolis B: The transcription factor snail represses Crumbs3
expression and disrupts apico-basal polarity complexes. Oncogene.
27:3875–3879. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Gilles C, Polette M, Mestdagt M, et al:
Transactivation of vimentin by beta-catenin in human breast cancer
cells. Cancer Res. 63:2658–2664. 2003.PubMed/NCBI
|
|
37
|
Onder TT, Gupta PB, Mani SA, Yang J,
Lander ES and Weinberg RA: Loss of E-cadherin promotes metastasis
via multiple downstream transcriptional pathways. Cancer Res.
68:3645–3654. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Stoops SL, Pearson AS, Weaver C, et al:
Identification and optimization of small molecules that restore
E-cadherin expression and reduce invasion in colorectal carcinoma
cells. ACS Chem Biol. 6:452–465. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Weng W, Yin J, Zhang Y, Qiu J and Wang X:
Metastasis-associated protein 1 promotes tumor invasion by
downregulation of E-cadherin. Int J Oncol. 44:812–818.
2014.PubMed/NCBI
|
|
40
|
Fogg VC, Liu CJ and Margolis B: Multiple
regions of Crumbs3 are required for tight junction formation in
MCF10A cells. J Cell Sci. 118:2859–2869. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Roh MH, Fan S, Liu CJ and Margolis B: The
Crumbs3-Pals1 complex participates in the establishment of polarity
in mammalian epithelial cells. J Cell Sci. 116:2895–2906. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Tunggal JA, Helfrich I, Schmitz A, et al:
E-cadherin is essential for in vivo epidermal barrier function by
regulating tight junctions. EMBO J. 24:1146–1156. 2005. View Article : Google Scholar : PubMed/NCBI
|