Introduction
The malignant phenotype is characterized with
extensive complexity that present a daunting problem with respect
to treatment. Successful therapy depends on identification and
targeting of critical functional nodes that will terminate the
tumorigenic state by differentiation or death. This problem is
definitely characteristic of the incurable multiple myeloma (MM), a
malignancy of plasma cells that accumulate in the bone marrow (BM)
(1). It is accepted that effectual
MM treatment will need to address the cells in their supportive
microenvironment as well as target compound signaling cascades so
as to overcome cell heterogeneity and evolving resistance (1).
A unifying trait of MM cells is their extensive
protein synthesis (2–4) supported by expanded endoplasmic
reticulum (5) and activated
unfolded protein response (UPR) pathways (6–8).
Deregulation of protein synthesis has been linked to human cancers,
MM included, with elevated global translation as well as increased
synthesis of proteins integral to the malignant phenotype (4).
Protein synthesis is controlled by multiple stimuli
primarily at the initiation level as observed under
growth-promoting conditions and stress-induced synthesis shutdown
(2,9–11).
Translation initiation depends on recruitment of eukaryotic
initiation complex eIF4F, which is composed of three proteins: the
5′ cap binding eIF4E, scaffolding protein eIF4G, and RNA helicase
eIF4A. Many tumors, including subtypes of MM, display elevated
levels of translation initiation factors (3,4).
Moreover, eIF4E overexpression induces malignant transformation and
drug resistance (12), yet its
knockdown (KD) selectively decreases translation of specific
targets with no effect on global protein synthesis (11). Concordantly, the availability of
eIF4E for capped mRNA translation dictates protein repertoire as
well as synthesis rate (13).
Regulation of eIF4E integrates metabolic and mitogenic cues with
cell proliferation and growth (mTOR/4E-BP1 and PI3K/Ras/ERK/MNK).
In fact, constitutive mitogenic activation of PI3K/Akt/mTOR (an
established MM characteristic) in combination with activated Ras
(detected in ~50% of MM cases) (2,14)
may afford an explanation for elevated free eIF4E via mTOR/4E-BP1
and Ras/MNK/eIF4E phosphorylations in MM. Furthermore, several
eIF4E targets are elevated in MM and may well be attributed to its
enhanced activity: MMP9, Bcl2, BclXL, survivin, cyclin D1 (cycD1),
c-Myc, and VEGF (2,14). Interestingly, the frequent
deregulation of c-Myc in MM can rarely be explained by
translocation to an Ig locus; however elevated eIF4E translation
could explain elevated c-Myc synthesis (15). If so, this could also feedback on
eIF4E levels by increasing its transcription (15).
Previously, we demonstrated the downregulation of
specific tetraspanin family members (CD81N1, CD82N1) in MM
(16,17). Tetraspanins are transmembrane
proteins that orchestrate protein complexes thereby regulating
proteins segregation and proximity and influencing fundamental
biological processes, such as adhesion, migration, and
proliferation. The expression of certain family members is
correlated with malignant processes and prognosis (18,19).
We also reported that overexpression of tetraspanin
family members (CD81N1, CD82N1) normally silenced in myeloma,
caused activation of UPR and autophagic death in MM cell lines,
indicative of breached protein homeostasis (proteostasis) (6,16,17).
Recently, we showed that elevated protein synthesis induced by the
overexpressed tetraspanins was instrumental in the MM cell death
(20). We also observed an
attenuation of eIF4E regulators, mTOR and PI3K signaling cascades
yet elevated protein synthesis (20,21).
Hence, we speculated that protein synthesis regulated by these
cascades might have been perturbed by CD81N1/CD82N1 and involved in
determining the fate of MM cells. We investigated the tetraspanins’
role in protein synthesis, particularly their effect on eIF4E. We
proceeded to explore the efficacy of targeting eIF4E directly, to
MM therapy.
Materials and methods
Cell lines and reagents
MM cell lines RPMI-8226, CAG, U266, ARP-1 and ARK
were cultured in RPMI-1640 supplemented with 20% heat-inactivated
FBS and antibiotics (Biological Industries, Kibbutz Beit-Haemek,
Israel). RPMI-8226 constitutively expressing Akt was prepared by us
previously (21) by stably
transfecting Akt1/PKBα in pUSEamp or empty control vector
(Millipore, Darmstadt, Germany) and selecting single cell clones
with 1 mg/ml G418 (Clontech, Mountain View, CA, USA). Cultured
cells (20,000 cells/96-well culture plates) were treated with
ribavirin (RBV) (0–7 μM) (Sigma, St. Louis, MO, USA) diluted in
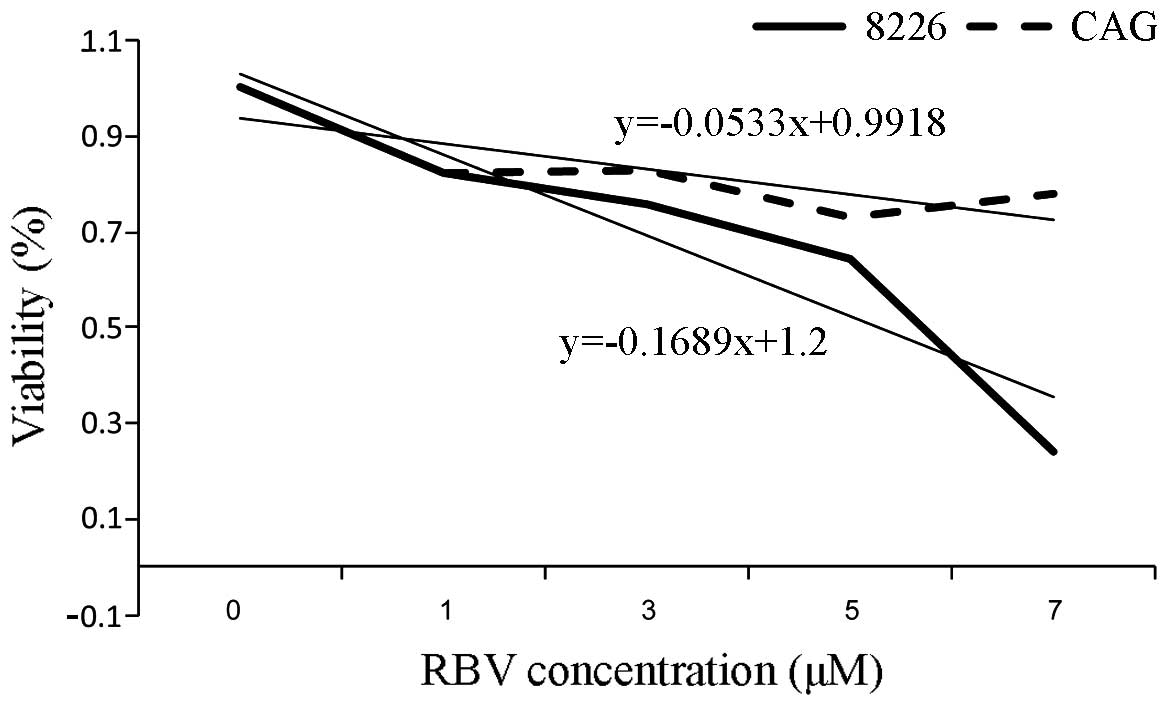
water. We determined 5 days RBV IC50 (4.1 μM for RPMI-8226, 9.2 μM
for CAG; Fig. 5). Velcade
(bortezomib) from the oncologic pharmacy at Meir Medical Center was
diluted in saline and used at 5 nM.
Study group
Eight peripheral blood samples obtained from healthy
volunteers aged 45–80 years and 18 BM samples obtained from
patients newly diagnosed (unless otherwise indicated) with MM were
studied. Details regarding myeloma isotype, burden (% monoclonal
CD138+ plasma cells/BM aspirate mononuclear cells),
light chain ratio (κ/λ), and β2 microglobulin levels are detailed
in Table I. Blood and BM
leucocytes were separated by Ficoll gradient (Sigma) according to
manufacturer’s instructions. The latter were hereafter regarded as
mononuclear cells enriched with MM cells. Drug-treated BMs were
cultured ≤5 days in RPMI-1640 supplemented with 20%
heat-inactivated FBS and antibiotics. Lack of significant death
under these conditions in control BM samples indicated that the
experimental model was appropriate. B cells tagged with anti-C
D19-Rd antibody (Dako, Glostrup, Denmark) were sorted from total
blood leukocytes (BD FAC SAria™ cell sorter; Tel Aviv University,
Tel Aviv, Israel). The study was approved by Meir Medical Center
Helsinki Committee, conducted according to Helsinki guidelines, and
all participants signed informed consent forms.
 | Table IClinical characteristics of the MM
patients. |
Table I
Clinical characteristics of the MM
patients.
| Patient no. | Gender | Age (years) | Diagnosis | Plasma cells in BM
aspirate mononuclear cells (%) | CD138+
of plasma cells (%) | Isotype (g/dl) | Light chain (κ/λ
ratio) | β2 microglobulin
(mg/l) |
|---|
| 1 | M | 59 | MM | 13 | 97 | Hypo | | κ 488 | 4.1 |
| 2 | M | 67 | MM | 20 | 90 | IgA | 3.1 | κ 3.2 | 2.1 |
| 3 | M | 60 | MM | 70 | 99 | Hypo | | κ 907 | 1.2 |
| 4 | M | 72 | MM | 40 | 89 | IgA | 4 | κ 166 | 4.7 |
| 5 | M | 53 | MM | 30 | 92 | Normo | | λ 0.02 | 3.4 |
| 6 | M | 68 | MM | 15 | 95 | IgG | 2.8 | κ 11.4 | 2.1 |
| 7 | M | 75 | MM | 15 | NT | IgG | 2.9 | κ 16.6 | 5.8 |
| 8 | M | 54 | MM | 80 | 95 | IgG | 3.6 | λ 0.00 | 5 |
| 9 | F | 50 | MM | 35 | 98 | Hypo | | λ 0.00 | NT |
| 10 | F | 55 | MM | 60 | 91 | IgA | 0.75 | λ 0.06 | 3.1 |
| 11 | F | 68 | MM | 30 | 55 | IgG | 3.5 | κ 133 | 2.2 |
| 12 | F | 62 | MM | 15 | 96 | IgG | 3 | λ 0.01 | 3.2 |
| 13 | F | 57 | SMM | 20 | NT | IgG | 3 | λ 0.03 | 2.7 |
| 14 | F | 54 | Relapse MM | 40 | 94 | Hypo | | λ 0.00 | 6.2 |
| 15 | F | 59 | SMM | 25 | 95 | IgG | 4.9 | κ 6127 | 2.5 |
| 16 | F | 52 | MM | 50 | 76 | Hypo | | λ 0.00 | 3.4 |
| 17 | F | 85 | MM | 35 | 94 | IgA | 0.85 | κ 1.73 | NT |
| 18 | F | 68 | MM | 50 | 93 | IgG | 2.6 | λ 0.1 | 3.5 |
Transient transfection of MM cell lines
and constitutive Akt/empty RPMI-8226
Purified plasmids pEGF P-N1 (N1/Mock), CD81N1N1-eGFP
(81N1) and CD82N1N1-eGFP (82N1) were separately introduced into
RPMI-8226, CAG, constitutive Akt-RPMI-8226, or empty RPMI-8226 as
described previously (17).
Fluorescence (≥10,000 events/analysis) was analyzed by flow
cytometer (FACS) (EPICS XL; Beckman Coulter, Nyon, Switzerland)
(17). Successful overexpression
of CD81N1, CD82N1, and Akt in our models was demonstrated by us
previously (17,21). Transfected and untransfected cells
were harvested 24 h post-transfection and stained with propidium
iodide (PI) (1 μg/ml) for 10 min. PI−/PI+
cells were enumerated by FAC S. PI− or
eGFP+/PI− were regarded as surviving cell
fractions, whereas PI+ and
eGFP+/PI+ were regarded as dead cells.
CD81N1N1/N1-transfected Akt/empty RPMI-8226 cells were harvested
19, 21 and 24 h post-transfection and assayed by immunoblotting for
protein levels.
siRNA transfection
Validated Alexa-labeled AllStars negative control
and eIF4E siRNAs (20 nM) (Qiagen, Valencia, CA, USA) were delivered
into MM cell lines using DMRIE-C.(Invitrogen Life Technologies,
Carlsbad, CA, USA). Fluorescence (≥10,000 events/analysis) was
analyzed by FACS (≥95%). Silencing of eIF4E was detected at the
transcript level by qPCR. Tested time points included 24, 48, 72,
96 and 120 h post-transfection. All analyses of siRNA-transfected
cells were done with unsorted cells.
Cell sorting
Transiently transfected cells (18/24 h
post-transfection) or CD19-R d-stained leucocytes were passed
through a syringe for clump dispersion. eGF P+ and
eGFP− cells or CD19+ and CD19−
cells (5×106 cells/ml 10% FBS PBS) were collected (BD
FAC SAria™ cell sorter; Tel Aviv University). eGFP−
cells treated with transfection reagent only were used to calibrate
the threshold of eGF P+ cells.
Quantitative reverse transcription
polymerase chain reaction (RT-qPCR)
Total RNA was extracted from 96 h siRNA-transfected
cells with PARIS kit (Applied Biosystems, Carlsbad, CA, USA). RNA
(1 μg) was reverse transcribed (GeneAmp RNA PCR; Applied
Biosystems) and amplified with QuantiFast SYBR-G reen RT-PCR kit
and eIF4E QuantiTect primers (cat. no. QT00046018; Qiagen)
according to manufacturer’s instructions. eIF4E expression was
normalized to β-actin. A standard sample dilution series was
executed with unknown samples for eIF4E and β-actin to verify the
linearity of the results.
Viability and proliferation (WST-1)
assay
Cell proliferation reagent WST-1 (Roche Diagnostics,
Basel, Switzerland) was used according to manufacturer’s
instructions. Absorbancies measured by a scanning multiwell
spectrophotometer (ELI SA reader, model Sunrise; Tecan, Salzburg,
Austria) were plotted on a linear scale. Cell survival was
estimated as percentage of solvent-treated control values.
Apoptosis/necrosis analysis
Cells (106) incubated with Annexin
V-PE.(250 μg/ml) (BioVision, Inc., Milpitas, CA, USA)/7AAD (0.05
μg/ml/15 min) (eBioscience, San Diego, CA, USA) were analyzed by
FAC S. Apoptotic Annexin+7AA Dand necrotic
Annexin+7AA D+ cells were enumerated. BM MM
samples were assayed for Annexin/7AAD in CD138+ cell
fraction thereby addressing MM cells only.
Western blotting
Cells were lysed 10 min on ice (6). Protein samples (300,000 cells) mixed
1:5 with loading buffer were separated by SDS-PAGE and transferred
to PVDF membrane. Membranes were blocked (5% milk powder) and
incubated with primary antibodies at 4°C overnight. Rabbit
anti-peIF4E (Ser209), total eIF4E, p4E-BP1 (Ser65), total 4E-BP1,
pMNK1 (Thr197/Thr202), total MNK1, cycD1, BiP/GR P78 and mouse
anti-c-Myc (1:1,000 dilution) were from Cell Signaling Technology,
Inc. (Danvers, MA, USA). Rabbit anti-GA DD153/CHOP (1:500) was from
Santa Cruz Biotechnology, Inc. (Santa Cruz, CA, USA) and rabbit
anti-ATF-6 (1:2,000) from Abcam (Cambridge, UK). The anti-pMNK1
antibody cross-reacts with pMNK2a and pMNK2b. Bound antibodies were
visualized using peroxidase-conjugated secondary antibody
(1:10,000, 75 min at 25°C; Jackson ImmunoResearch Laboratories,
West Grove, PA, USA) and ECL detection (Santa Cruz Biotechnology,
Inc.). Products were visualized with LA S3000 Imager (Fujifilm,
Greenwood, SC, USA). Integrated optical densities of immunoreactive
protein bands were measured as arbitrary units employing Multi
Gauge software (Fujifilm). All study results were normalized to
cell number and tubulin-loading control.
Statistical analysis
Student’s paired t-tests were used to analyze
differences between cohorts. Effects were considered significant
when p≤0.05. An antagonistic effect was verified by drugs’
interaction formula q = P(A + B)/P(A) + P(B) − P(A) × P(B)
(q<0.85, antagonist; q>1.15, synergist; 1.15>q>0.85,
additive) (22). All experiments
were conducted 3–7 separate times.
Results
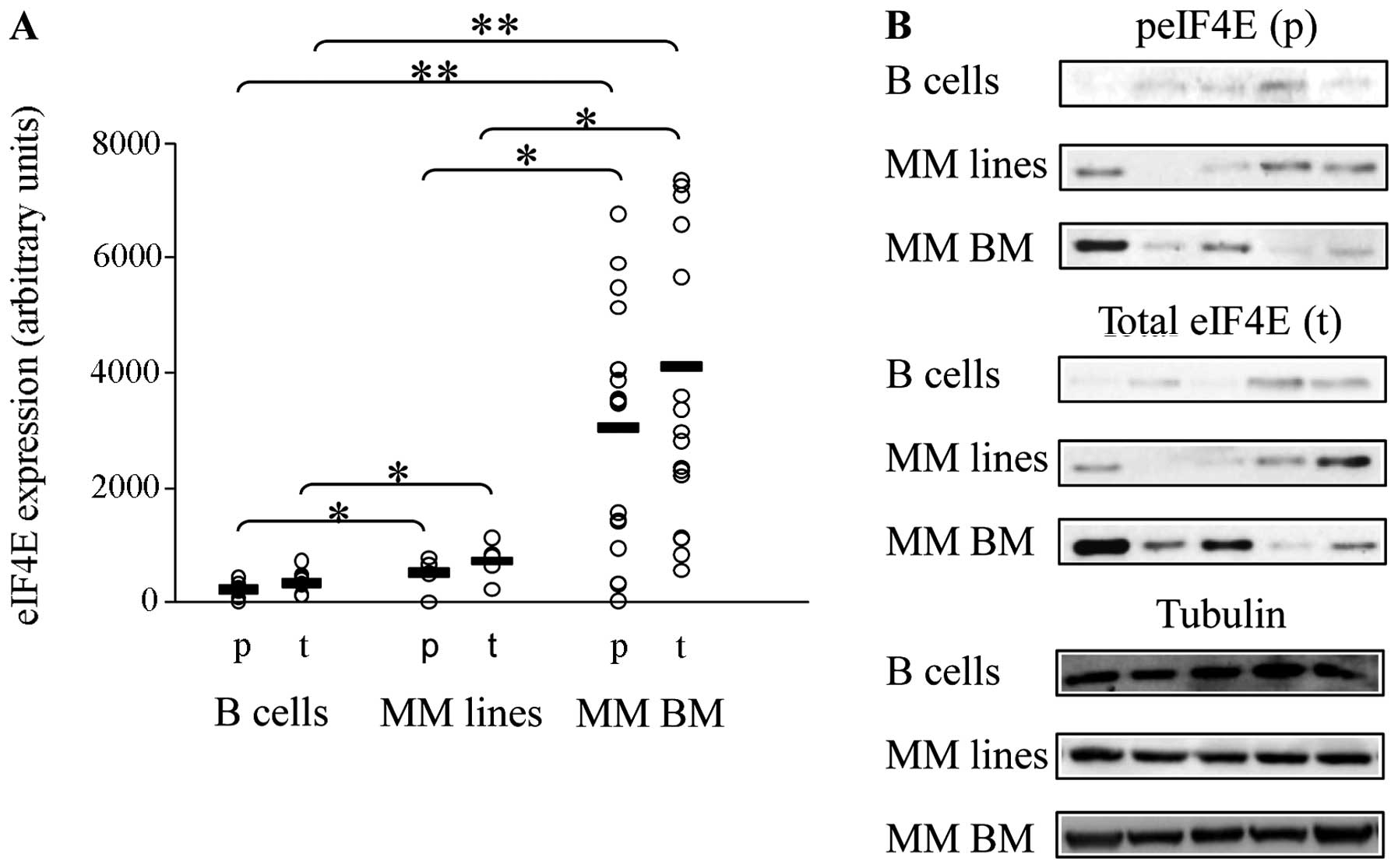
Baseline expression of eIF4E in MM
We determined the expression and baseline levels of
eIF4E protein in a panel of MM cell lines (n=5) and BM mononuclear
cells from MM patients (n=18) compared to peripheral B cells from
normal donors (n=8). The comparison to normal peripheral B cells
was implemented as done by others and us previously (16,23)
because normal plasma cells are not readily available and patients’
BM samples were limited in volume due to prior use for medical
purposes. This disallowed CD138+ cell sorting in
quantities necessary for immunoblotting so we assayed BM
mononuclear cells enriched with MM cells (Table I), as described in Materials and
methods. We determined eIF4E is expressed in all specimens
(immunoblot) (Fig. 1). Higher
levels of total and phosphorylated eIF4E were observed in MM cells
compared to B cells (cell lines: 2-fold change, BM samples:
~10-fold change; p<0.05) (Fig.
1). The differences in expression of phosphorylated and total
eIF4E between MM cell lines and BM samples were also significant
(p<0.05) and elaborated upon in the Discussion. These findings
suggest a potential role for eIF4E in MM.
Tetraspanins attenuated levels of
translation initiation factor eIF4E in MM cell lines
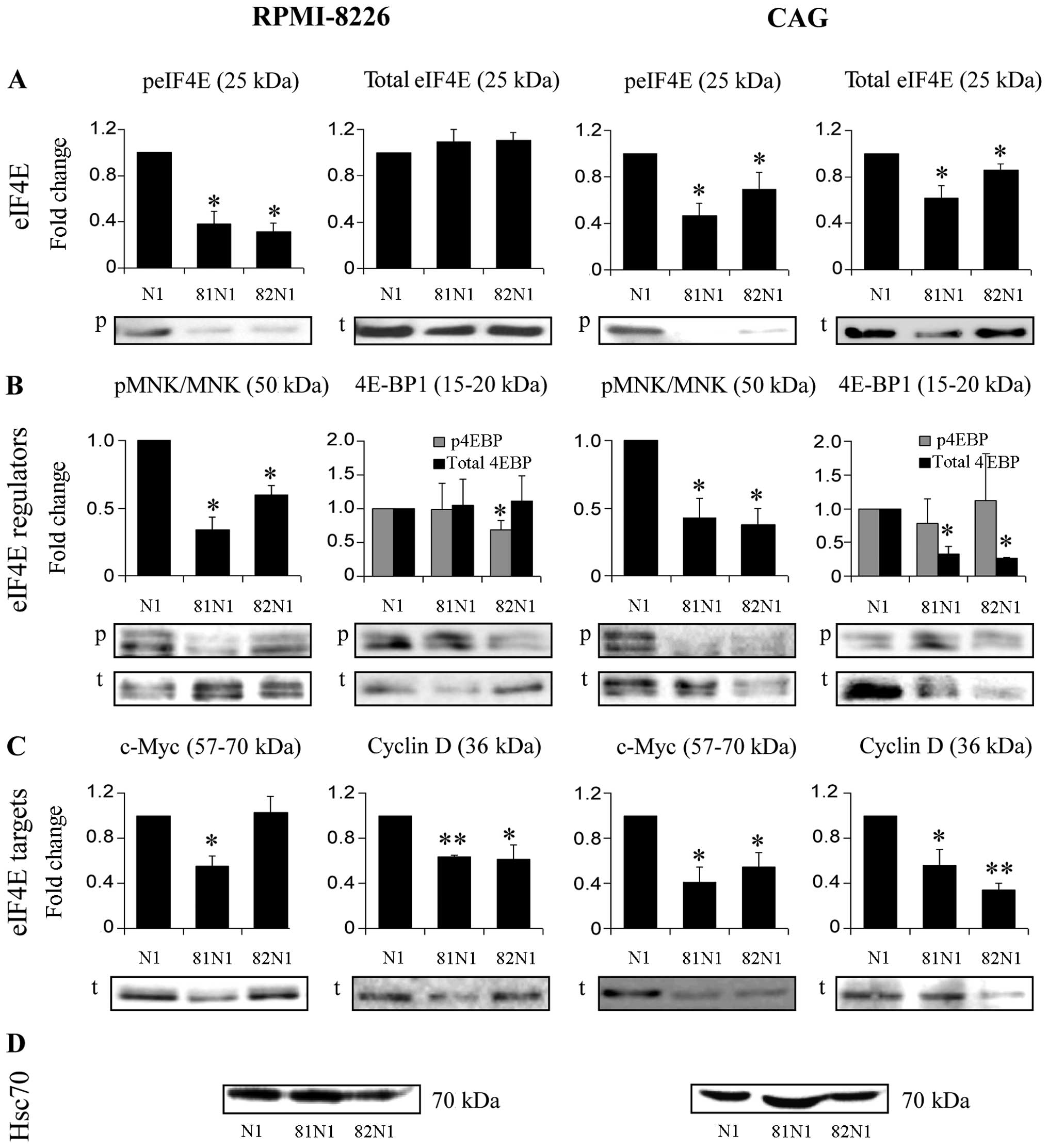
Next, we transiently transfected CD81N1 and CD82N1
into MM cell lines (RPMI-8226, CAG ). The actual overexpression of
the tetraspanins was corroborated by flow cytometry (using anti-C
D81N1 and anti-C D82N1 monoclonal antibodies) and microscopic
analyses (17). Subsequently we
assayed total and phosphorylated eIF4E protein levels in sorted
CD81N1/CD82N1/Mock (N1)-transfected MM cell lines (RPMI-8226, CAG
). A decrease in phosphorylated eIF4E protein levels was detected
in the MM cell lines with both tetraspanin vectors (~60%↓ in
RPMI-8226, ~40%↓ in CAG, p<0.05) (Fig. 2A). The cell lines differed in the
effect CD81N1 and CD82N1 had on total eIF4E levels. A decrease in
total eIF4E level was determined in CAG transfected with
CD81N1/CD82N1 (~40%↓ and ~15%↓, respectively, p<0.05) yet no
change in its levels were seen in RPMI-8226 (Fig. 2A).
Then we examined the expression levels of major
eIF4E regulators. Concordantly with the common tetraspanin-induced
decrease in phosphorylated eIF4E we also determined a decrease in
the phosphorylated active form of its kinase MNK1/2, in both cell
lines (40–65%↓, p<0.05) (Fig.
2B). Examination of the eIF4E-binding protein, 4E-BP1,
uncovered significant changes in its expression following
CD81N1/CD82N1 overexpression in both cell lines (Fig. 2B). RPMI-8226 was characterized with
decreased phosphorylated 4E-BP1 in CD82N1 overexpressing cells
(32%↓, p<0.05) but no change in CD81N1. CAG was typified by a
decrease in the total 4E-BP1 levels with both tetraspanin vectors
(55–65%↓, p<0.05) without changes in phosphorylated 4E-BP1.
We examined the expression of known eIF4E targets in
order to validate whether CD81N1/CD82N1 indeed caused a reduction
in eIF4E activity (Fig. 2C). A
common reduction in cycD1 was observed in both cell lines with
CD81N1 and CD82N1 (40–65%↓, p<0.05) (Fig. 2C). Reduced expression of c-Myc was
observed in CD81N1 transfected RPMI-8226 and CD81N1/CD82N1
transfected CAG (45–60%↓, p<0.05) (Fig. 2C).
Taken together, our findings so far indicate that
the tetraspanins reduced the activity of the translation initiation
factor eIF4E and its targets (Table
II).
| Table IISummary of the responses of MM cell
lines to eIF4E modulation. |
Table II
Summary of the responses of MM cell
lines to eIF4E modulation.
| Assayed
parameter | | Model |
|---|
| |
|
|---|
| | R PMI-8226 | CAG |
|---|
| |
|
|
|---|
| | C D81N1N1 | C D82N1N1 | CD81N1N1 | C D82N1N1 |
|---|
| eIF4E, regulators
and targets | Phospho-eIF4E | ↓ | ↓ | ↓ | ↓ |
| T otal eIF4E | = | = | ↓ | ↓ |
| 4E-BP1 | = | ↓ | ↓ | ↓ |
| MNK1/2 | ↓ | ↓ | ↓ | ↓ |
| c-Myc | ↓ | = | ↓ | ↓ |
| Cyclin D1 | ↓ | ↓ | ↓ | ↓ |
| Constitutive
Akt | peIF4E | ↑ | NT | NT | NT |
| pMKN1/2 | ↑ | NT | NT | NT |
| Rescue | ✓a | ✓a | ✓a | ✓a |
|
| Treatment | Assayed
parameter | Model |
|
| R PMI-8226 | CAG |
|
| siRNA eIF4E | C ell count | = | ↓ |
| Viability | ↓ | ↓ |
| Death | ↑ | ↑ |
| UPR | ↑ | ↑ |
| c-Myc | ↓ | ↓ |
| C yclin D1 | ↓ | ↓ |
| Ribavirin | C ell count | = | = |
| Viability | ↓ | ↓ |
| Death | = | = |
| UPR | ↑ | ↑ |
| c-Myc | ↓ | ↓ |
| C yclin D1 | ↓ | ↓ |
|
Ribavirin+Velcade | C ell count | ↓ | ↓ |
| Viability | ↓ | ↓ |
| Death | ↑ | ↑ |
| UPR | ↑ | ↑ |
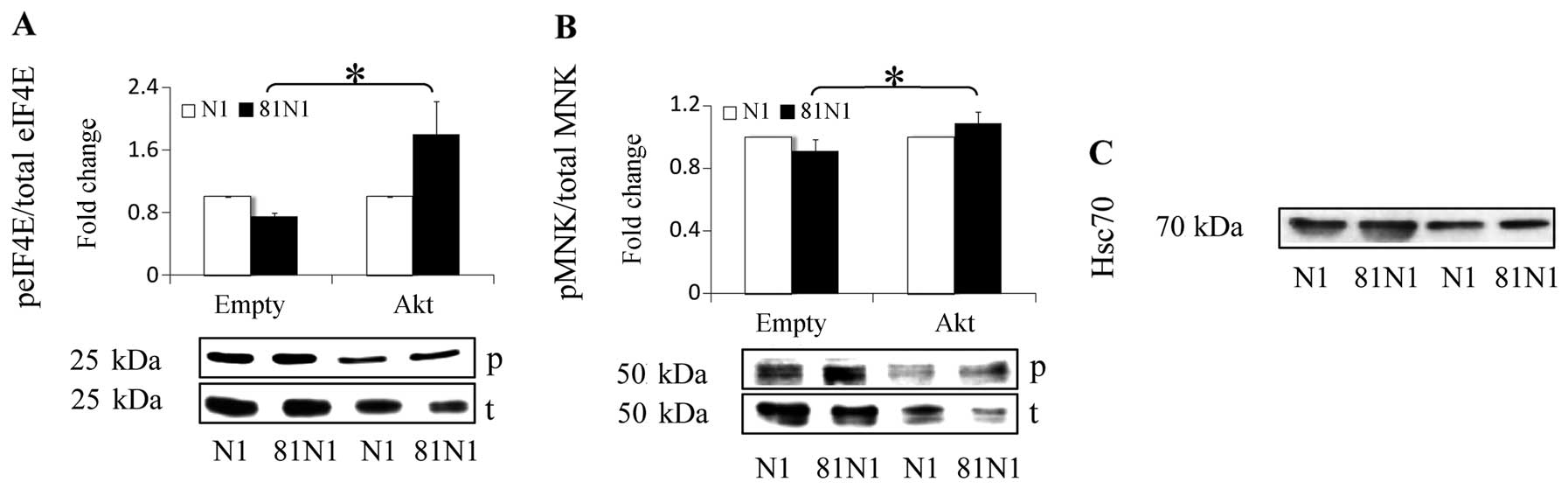
Constitutive Akt abrogates peIF4E
reduction in CD81N1-transfected RPMI-8226 and rescues cells from
consequent death
In a previous study we constructed and corroborated
a multiple myeloma model expressing constitutive Akt (21). Furthermore, we have demonstrated
the capability of constitutively active Akt to partially rescue the
MM cell lines (RPMI-8226, CAG ) from the tetraspanin
(CD81N1/CD82N1)-induced death (21). Here we wanted to explore the
possible involvement of eIF4E status in this phenomenon. Thus, we
used the same model of control (empty vector-RPMI-8226) or stably
transfected Akt-R PMI-8226 co-transfected with a Mock vector N1 or
CD81N1, and re-assessed cell death and levels of peIF4E and
pMNK1/2.
Increased peIF4E levels were observed in
CD81N1-transfected Akt-RPMI-8226 compared to the CD81N1-transfected
control empty vector-R PMI-8226 (105%↑, p<0.05) (Fig. 3A). This was coupled with a definite
rescue of the transfected cells from death (21) (28%, p<0.05). Analysis of the MM
cells 3 h earlier, as described in Materials and methods, displayed
elevated levels of pMNK1/2 in Akt-RPMI-8226 expressing CD81N1 cells
relative to the empty vector-RPMI-8226 co-transfected with CD81N1
(18%↑, p<0.05) (Fig. 3B). The
timing of elevated phosphorylated MNK1/2 is consistent with its
position in the signaling cascade, downstream of Akt and upstream
of eIF4E. These results demonstrate the co-regulation of survival
and peIF4E levels in tetraspanin-overexpressing MM cell lines
(Table II).
KD of eIF4E deleteriously affects MM cell
lines
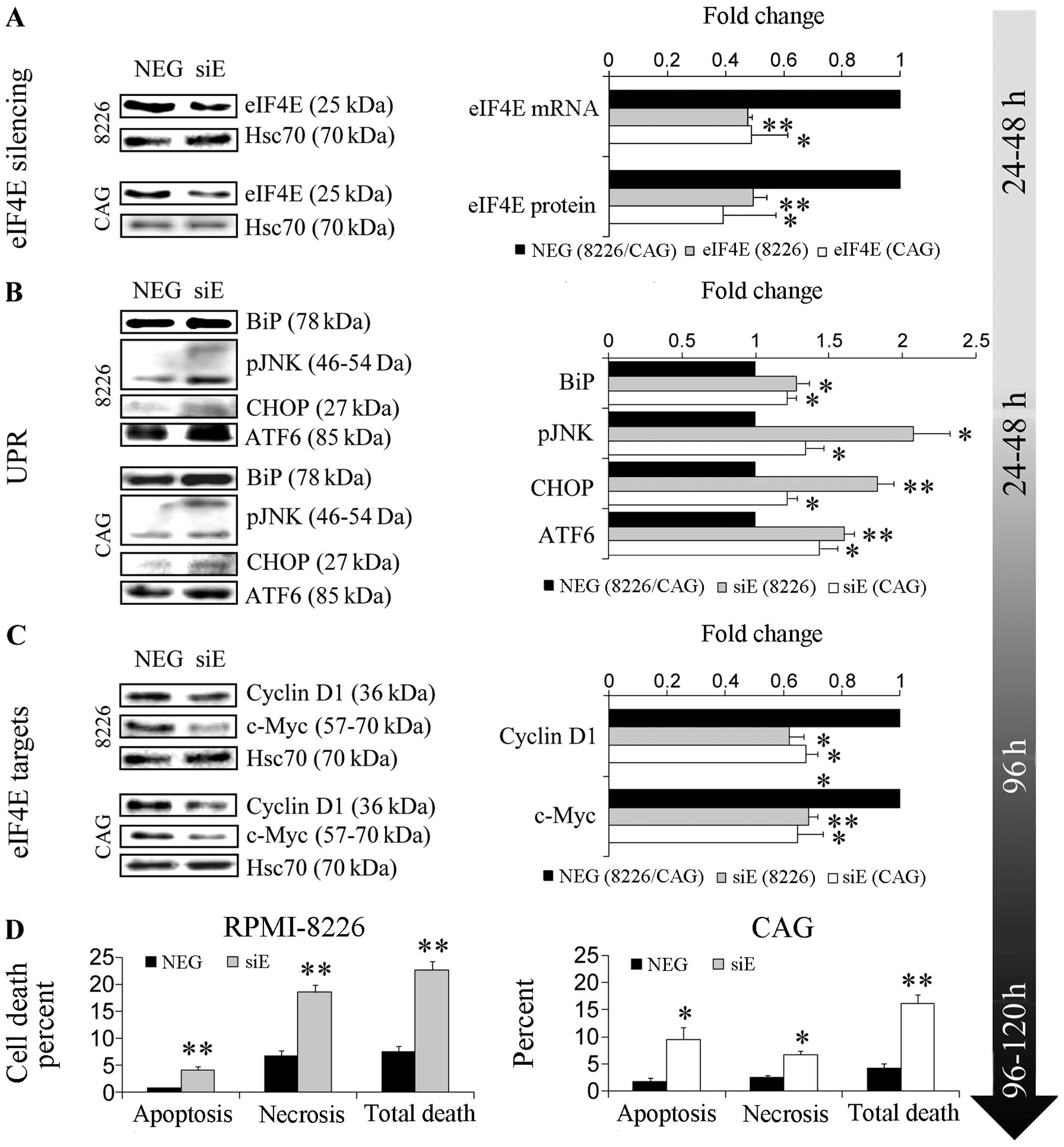
Next, we wanted to confirm directly that eIF4E
activity is indeed critical to MM cell lines’ growth and survival.
Thus, we introduced anti-eIF4E siRNA into the MM cell lines (~85%
transfection in RPMI-8226, ~95% in CAG ) and validated KD of eIF4E
at 24 and 48 h post-transfection (RNA and protein, respectively,
Fig. 4A). eIF4E KD was detected in
unsorted cells by qPCR (50%↓ in RPMI-8226 and CAG, p<0.01 and
p<0.05, respectively) and by immunoblotting (50%↓ in RPMI-8226,
p<0.01 and 60%↓ CAG, p<0.05) (Fig. 4A). We went on to assay the
consequences of eIF4E KD on the expression of its dependent
targets. Again, significant attenuation of cycD1 and c-Myc was
observed in both cell lines 96 h post-transfection demonstrating
that eIF4E activity was indeed compromised (30–40%↓ in RPMI-8226
and 30–35%↓ in CAG, p<0.01 and p<0.05, respectively)
(Fig. 4C). Subsequently, we
assessed the affect of eIF4E KD on the phenotype of MM cell lines
96–120 h post-transfection. Decreased viability was evidenced in
both cell lines (96 h post-transfection: 20%↓ in RPMI-8226; 10%↓ in
CAG; p<0.05). A decrease in cell count was also determined in
CAG only (96 h post-transfection: 33%↓; p<0.05). Analysis of
cell death demonstrated moderately increased total cell death in
both cell lines 96 h post-siRNA transfection in CAG and 120 h in
8226 (10–12%↑, p<0.01) attributed to apoptosis and necrosis
(2–8%↑ apoptosis, p<0.01 and 4–8%↑ necrosis, p<0.05)
(Fig. 4D, Table II). Higher death rates were
evident in both cell lines at 120 h post-transfection (15%↑ total
death, p<0.01). Next, we assayed eIF4E KD on MM cell
proteostasis by studying the expression of UPR proteins. A
significant elevation of all UPR signaling arms (PERK, IRE 1, and
ATF-6 represented by CHOP, pJNK and ATF-6, respectively) was
determined in both MM cell lines. RPMI-8226 displayed at 48 h
post-transfection elevated BiP (28%↑, p<0.05), CHOP (83%↑,
p<0.01), pJNK (107%↑, p<0.05), and ATF-6 (61%↑, p<0.01)
(Fig. 4B). CAG displayed at 24 h
post-transfection elevated BiP (22%↑, p<0.05), CHOP (22%↑,
p<0.05), pJNK (35%↑, p<0.05), and ATF-6 (44%↑, p<0.05)
(Fig. 4B). Taken together, these
findings suggest that inhibition of eIF4E perturbs proteostasis in
MM cell lines and enhances stress that leads to activation of the
UPR.
Competitive inhibition of eIF4E with RBV
elevates ER stress in MM cell lines
Our findings so far indicate that eIF4E diminution
may have anti-myeloma activity but the delivery of siRNA is
currently clinically difficult. Therefore, we examined the effect
of a clinically employed anti-viral drug, RBV suggested to act as a
mimic for the 5′ cap present on most human transcripts thereby
competing with eIF4E binding (24–27).
RBV doses were calibrated and IC50 was determined for each cell
line (Fig. 5), as described in
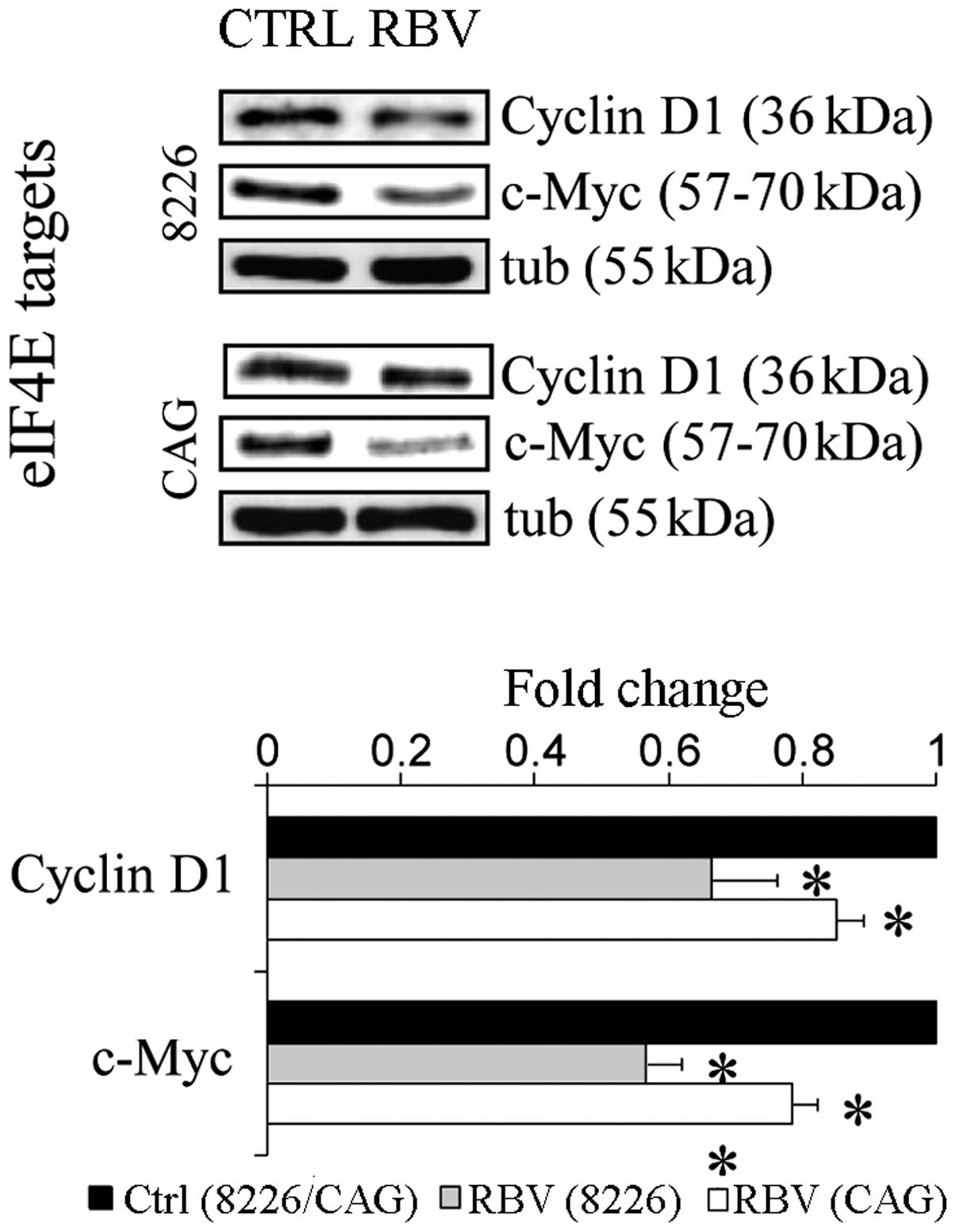
Materials and methods. As hypothesized, RBV (5 μM, 5 days)
(24) caused decreased expression
of eIF4E-dependent targets cycD1 and c-Myc in both cell lines
(15–35%↓ and 20–45%↓, respectively; p<0.05) (Fig. 6). Furthermore, RBV caused increased
UPR signals indicative of elevated ER stress in RPMI-8226 (35%↑
BiP, p<0.01; 48%↑ pJN K, p<0.05) and CAG (39%↑ BiP,
p<0.05; 35%↑ pJN K, p<0.05%, 44%↑ CHOP, p<0.01; 49%↑
ATF-6, p<0.05) (Fig. 7A,
Table II). RBV also caused mild
but significant reduction in the viability of MM cell lines (~10%↓,
p<0.05). No biologically significant differences in cell counts
or death were determined.
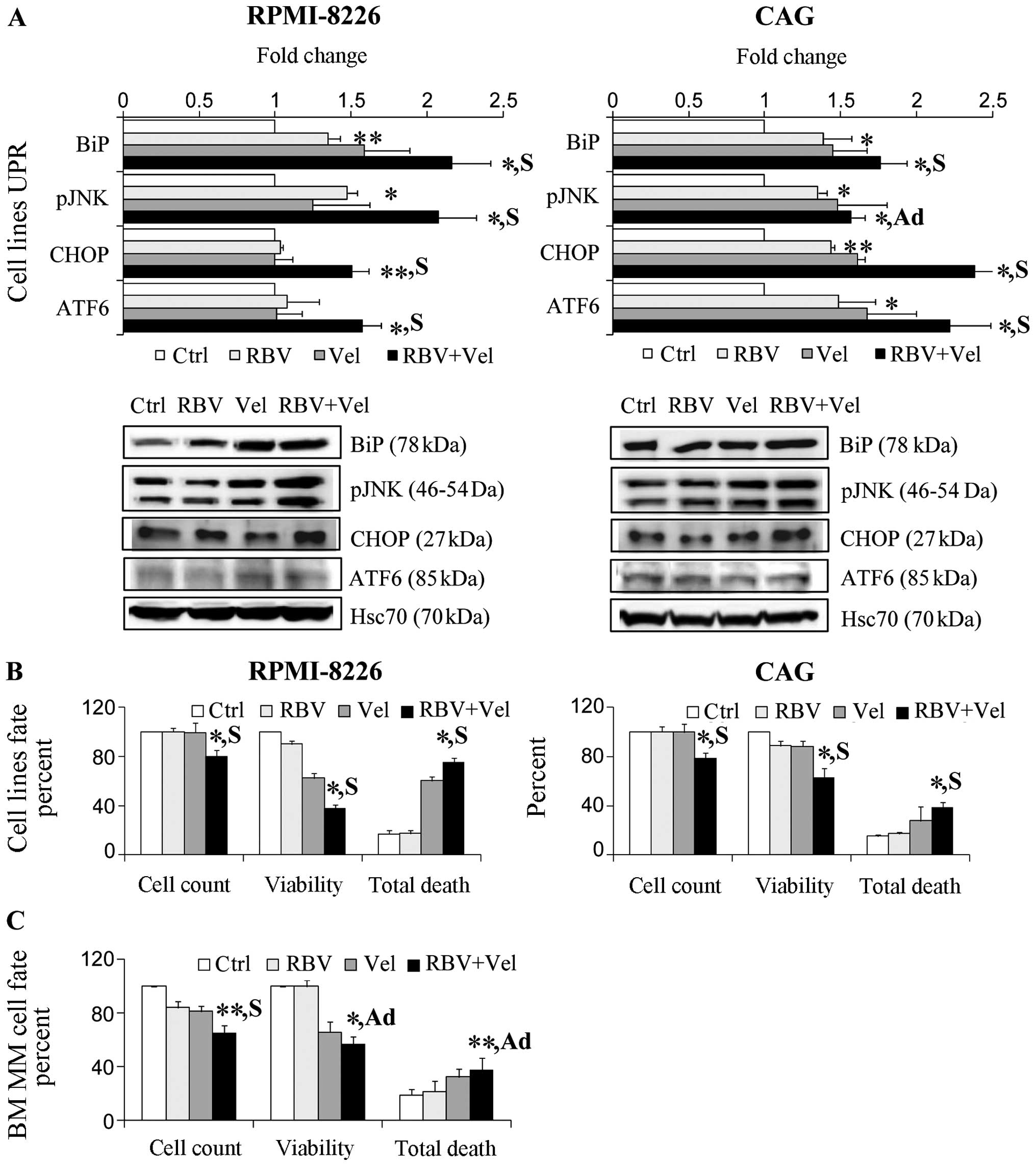 | Figure 7Combined ribavirin (RBV) and velcade
(Vel) is advantageous in anti-myeloma treatment. Multiple myeloma
(MM) cell lines and samples from bone marrow (BM) of MM patients
were treated with solvent, 5 μM RBV, 5 nM Vel or RBV and Vel
combination. (A) Combined RBV and Vel-affected unfolded protein
response (UPR) of MM cell lines. Representative immunoblots (bottom
panel) and graphic presentations (top panel) of UPR signaling arms
are presented (mean ± SE, n≥3). Hsc70 served as a loading control.
The effect of RBV and Vel treatment on (B) MM cell lines and (C) BM
CD138+ cell fraction count, viability, and death is
graphically presented. All assayed parameters measured in treated
cells were normalized to solvent treated control cells (mean ± SE,
n≥3). Statistically significant differences (*p<0.05,
**p<0.01), synergistic (S, q>1.15), and additive
(Ad, 0.85<q<1.15) effects of combined treatment are
indicated. |
Combined RBV and velcade has anti-myeloma
potential
Based on the observations we have made so far, we
hypothesized that RBV may sensitize MM cells to drugs that induce
ER-stress, such as velcade (bortezomib) (28,29).
Indeed, in combined treatment of MM cell lines with
5 μM RBV and 5 nM velcade for 5 days we witnessed significantly
increased UPR compared to the effect of each drug administered
alone (RPMI-8226: 116%↑ BiP, 107%↑ pJN K, 50%↑ CHOP, 57%↑ ATF-6,
p<0.05; CAG: 76%↑ BiP, 57%↑ pJN K, 138%↑ CHOP, 122%↑ ATF-6,
p<0.05). This combined effect is synergistic (q>1.15) or
additive (q=0.86) (Fig. 5A).
RPMI-8226 and CAG treated with combined RBV and velcade also
demonstrated reduced cell counts (20%↓, p<0.05), reduced
viability (40–72%↓, p<0.05), and increased cell death (63–75%↑,
p<0.05) (Fig. 6B, Table II) in comparison to each drug
administered alone (synergistic, q>1.15). Increased
proliferation coupled with elevated death would result in minimal
changes to steady state cell numbers yet higher extents of cell
death.
Next, we administered RBV alone and in combination
with velcade to primary mononuclear BM samples (enriched with MM
cells) (n=7). We ruled out the possibility that culture conditions
caused decreased viability by demonstrating no decrease in control
cells cultured up to 9 days (data not shown). Treatment of BM MM
cells with combined RBV and velcade showed a significantly
synergistic (q>1.15) and additive (q=1) deleterious effect
compared to the drugs administered individually (Fig. 7C). Particularly, this anti-myeloma
response was evidenced in diminished cell counts (35%↓, p<0.01),
viability (43%↓, p<0.01), and increased cell death in
CD138+ cell fraction (FAC S) (37%↑, p<0.05).
Discussion
The premise of our study was that CD81N1N1/CD82N1N1
overexpression breached the proteostasis of MM cell lines, based on
activation of UPR and induction of autophagic death (6). The increased fatal protein synthesis
caused by the re-expression of tetraspanins (20) suggested that their natural
downregulation in MM cells (16)
may sustain specific ‘proteostasis network’ characteristics suited
for the malignant cell survival and proliferation. Furthermore, the
reduced mTOR and Akt signals in our research model (21) were consistent with altered eIF4E
activity. Indeed, this study shows that CD81N1/CD82N1 transfection
decreases eIF4E expression and function and may reaffirm that
reduced CD81N1/CD82N1 in MM cells is instrumental to elevated eIF4E
expression and beneficial to MM cells survival.
eIF4E expression was described previously in MM
cells (3,30) but to the best of our knowledge its
importance was not addressed directly. We showed elevated
expression of eIF4E in myeloma cells (BM samples and cell lines).
Additionally, higher eIF4E levels were observed in BM myeloma cells
compared to MM cell lines. This phenomenon may be attributed to
signals originating from the BM microenvironment that affect eIF4E
and are absent in cell culture.
The significance of eIF4E to myeloma cell survival
and proliferation is evidenced in our study at multiple levels. We
demonstrated that the expression of c-Myc and cycD1 in RPMI-8226
and CAG is contingent on eIF4E activity. Both proteins are
oncogenes overexpressed in MM and important to cell growth and
survival. In many cases of MM there is no obvious mechanistic
explanation for elevated c-Myc expression and our results may
afford clarification of this issue (2,14).
The same is true for cases of elevated cycD1 that is not caused by
Ig translocation. Interestingly, in a previous study conducted on
the same model we showed that CD81N1N1/CD82N1N1 caused a decrease
in MMP9, which is also dependent on eIF4E (17,31).
Using constitutively expressing Akt-RPMI-8226 model we elaborated
upon previous findings (21) and
demonstrated that rescue from CD81N1-induced death is accompanied
by increased eIF4E phosphorylation. We also showed elevated
pMNK1/2. Taken together, these results suggest that the
tetraspanins critically attenuate Akt activity, causing decreased
pmTOR, pMNK1/2 and consequently diminished peIF4E levels.
siRNA and RBV mediated inhibition of eIF4E further
stratified its importance to MM cell line growth and survival. This
negative effect may be partially attributed to the downregulation
of c-Myc and cycD1. Of note, while anti-eIF4E siRNA caused MM cell
death RBV induced stress that did not result in death. Mechanistic
differences may underlie this disparity since anti-eIF4E siRNA
diminishes its expression, whereas RBV only abolishes its capped
mRNAs’ association thereby leaving eIF4E available for other
cellular functions such as mRNA transport (32). The use of siRNA and a specific
chemical inhibitor of eIF4E circumvent the limitations of the
artificial tetraspanin overexpression model and confirm the
accuracy of our findings. Moreover, our current observations and
previously published data on the tetraspanin-induced changes in the
MM cell line model all support the deduction that the regulation of
proteostasis is critically important to survival of MM cell lines
and that eIF4E expression level is instrumental to this
equilibrium. A reasonable assumption is that the
tetraspanin-induced death of the MM cell lines is a sum consequence
of the compiled effects as previously reported (6,20,21).
Our findings regarding eIF4E regulation comply with
published data and substantiate a role for both Ras/MNK and
mTOR/4E-BP1 (33). It is
established that eIF4E phosphorylation is carried out by MNK1/2,
and indeed we witnessed reduced phosphorylated MNK1/2 in
concordance with reduced phosphorylated eIF4E with both tetraspanin
vectors in both MM cell lines (33). The role of 4E-BP1 is more complex.
On the one hand, it binds eIF4E in the cytoplasm thereby preventing
its inclusion in the translation initiation factor eIF4F (10), on the other hand, it has been shown
to be instrumental to nuclear compartmentalization of eIF4E, where
the translation factor promotes export of specific mRNAs to the
cytoplasm for translation (34,35).
It was shown that fully phosphorylated 4E-BP1, manifested in
phosphorylated serine 65 residues, releases eIF4E (10). Free eIF4E is phosphorylated and
activated or ubiquitinated and degraded (10). We showed MM cell lines displayed
different regulatory mechanisms where 4E-BP1 is concerned.
RPMI-8226 displayed stable total eIF4E levels probably due to
increased protective binding to un-phosphorylated 4E-BP1. CAG
demonstrated decreased total eIF4E levels possibly explained by the
significantly depleted 4E-BP1 levels. Regulation of 4E-BP1 by
degradation was published in a model of sea urchins (36).
Recently, we reported that tetraspanins affect
global protein synthesis (20),
yet to the best of our knowledge this is the first account linking
tetraspanins to specific mechanisms regulating the process.
Previous studies described association of tetraspanin members with
cell proliferation and production of specific proteins failed make
a more general connection to cell proteostasis (37–39).
Most interesting are publications that described the tetraspanin
influence on expression of proteins that can now be categorized as
eIF4E-dependent targets (40–42).
Available therapeutic approaches unbalance proteostasis by limiting
protein synthesis and availability or affecting protein
degradation. Our results present a tetraspanin-dependent
attenuation of eIF4E as a means to critically unbalancing myeloma
cell proteostasis.
An unresolved conundrum is the conflicting
observations we have made that on the one hand tetraspanins elevate
global protein synthesis (20) yet
on the other hand cause decrease of eIF4E levels and activity.
While these findings are in concordance with eIF4E KD studies that
showed global protein synthesis is unaffected (43) we cannot currently explain by what
mechanism the tetraspanins promote protein production. Current
studies underway in our laboratory are exploring the possible
involvement of eIF4GI on the effect of tetraspanins.
Altogether, our results demonstrate that targeting
eIF4E in MM has therapeutic potential. This is especially promising
due to the various publications proclaiming that the initiation
factor is overexpressed in cancer cells, affording a certain
advantage in terms of selectivity. This approach is clinically more
feasible with the use of RBV, as we had shown since it is a widely
prescribed anti-viral drug and has limited and treatable
side-effects (44). The efficacy
of this strategy was previously demonstrated in acute myeloid
leukemia and in head and neck cancer (45,46).
Interestingly, a recently published case report described
regression of myeloma in a patient treated with RBV for HCV
infection (47). The advantageous
combination of RBV and velcade affords proof-of-concept to the
potential utility of simultaneously targeting various proteostasis
arms in MM treatment.
Acknowledgements
This study constitutes sections of the Ph.D. theses
of Victoria Zismanov and Oshrat Attar-Schneider, Sackler Faculty of
Medicine, Tel Aviv University, Israel. Victoria Zismanov and Oshrat
Attar-Schneider contributed equally to the research and preparation
of the manuscript. We are grateful to the staff of the
Hematological Laboratory at Meir Medical Center for their dedicated
technical support. This study was supported by the Tel Aviv
University Research Grant no. 0601242791.
References
|
1
|
Anderson KC: Targeted therapy of multiple
myeloma based upon tumor-microenvironmental interactions. Exp
Hematol. 35(Suppl 1): S155–S162. 2007. View Article : Google Scholar
|
|
2
|
Pratt G: Molecular aspects of multiple
myeloma. Mol Pathol. 55:273–283. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Agnelli L, Fabris S, Bicciato S, et al:
Upregulation of translational machinery and distinct genetic
subgroups characterise hyperdiploidy in multiple myeloma. Br J
Haematol. 136:565–573. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Barnhart BC and Simon MC: Taking aim at
translation for tumor therapy. J Clin Invest. 117:2385–2388. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Cenci S and Sitia R: Managing and
exploiting stress in the antibody factory. FEBS Lett.
581:3652–3657. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Zismanov V, Lishner M, Tartakover-Matalon
S, Radnay J, Shapiro H and Drucker L: Tetraspanin-induced death of
myeloma cell lines is autophagic and involves increased UPR
signalling. Br J Cancer. 101:1402–1409. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Carrasco DR, Sukhdeo K, Protopopova M, et
al: The differentiation and stress response factor XBP-1 drives
multiple myeloma pathogenesis. Cancer Cell. 11:349–360. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Patterson J, Palombella VJ, Fritz C and
Normant E: IPI-504, a novel and soluble HSP-90 inhibitor, blocks
the unfolded protein response in multiple myeloma cells. Cancer
Chemother Pharmacol. 61:923–932. 2008. View Article : Google Scholar
|
|
9
|
Larsson O, Li S, Issaenko OA, et al:
Eukaryotic translation initiation factor 4E induced progression of
primary human mammary epithelial cells along the cancer pathway is
associated with targeted translational deregulation of oncogenic
drivers and inhibitors. Cancer Res. 67:6814–6824. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Wang X and Proud CG: Methods for studying
signal-dependent regulation of translation factor activity. Methods
Enzymol. 431:113–142. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Montanaro L and Pandolfi PP: Initiation of
mRNA translation in oncogenesis: the role of eIF4E. Cell Cycle.
3:1387–1389. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Wu KD, Zhou L, Burtrum D, Ludwig DL and
Moore MA: Antibody targeting of the insulin-like growth factor I
receptor enhances the anti-tumor response of multiple myeloma to
chemotherapy through inhibition of tumor proliferation and
angiogenesis. Cancer Immunol Immunother. 56:343–357. 2007.
View Article : Google Scholar
|
|
13
|
Ramirez-Valle F, Braunstein S, Zavadil J,
Formenti SC and Schneider RJ: eIF4GI links nutrient sensing by mTOR
to cell proliferation and inhibition of autophagy. J Cell Biol.
181:293–307. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Kastrinakis NG, Gorgoulis VG, Foukas PG,
Dimopoulos MA and Kittas C: Molecular aspects of multiple myeloma.
Ann Oncol. 11:1217–1228. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Lin CJ, Cencic R, Mills JR, Robert F and
Pelletier J: c-Myc and eIF4F are components of a feedforward loop
that links transcription and translation. Cancer Res. 68:5326–5334.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Drucker L, Tohami T, Tartakover-Matalon S,
et al: Promoter hypermethylation of tetraspanin members contributes
to their silencing in myeloma cell lines. Carcinogenesis.
27:197–204. 2006. View Article : Google Scholar
|
|
17
|
Tohami T, Drucker L, Shapiro H, Radnay J
and Lishner M: Overexpression of tetraspanins affects multiple
myeloma cell survival and invasive potential. FASEB J. 21:691–699.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Lazo PA: Functional implications of
tetraspanin proteins in cancer biology. Cancer Sci. 98:1666–1677.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Charrin S, le Naour F, Silvie O, Milhiet
PE, Boucheix C and Rubinstein E: Lateral organization of membrane
proteins: tetraspanins spin their web. Biochem J. 420:133–154.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Zismanov V, Drucker L, Attar-Schneider O,
Matalon ST, Pasmanik-Chor M and Lishner M: Tetraspanins stimulate
protein synthesis in myeloma cell lines. J Cell Biochem.
113:2500–2510. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Lishner M, Zismanov V, Tohami T,
Tartakover-Matalon S, Elis A and Drucker L: Tetraspanins affect
myeloma cell fate via Akt signaling and FoxO activation. Cell
Signal. 20:2309–2316. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Su DF, Xu LP, Miao CY, Xie HH, Shen FM and
Jiang YY: Two useful methods for evaluating antihypertensive drugs
in conscious freely moving rats. Acta Pharmacol Sin. 25:148–151.
2004.PubMed/NCBI
|
|
23
|
Thiago LS, Perez-Andres M, Balanzategui A,
et al: Circulating clonotypic B cells in multiple myeloma and
monoclonal gammopathy of undetermined significance. Haematologica.
99:155–162. 2014. View Article : Google Scholar :
|
|
24
|
Assouline S, Culjkovic B, Cocolakis E, et
al: Molecular targeting of the oncogene eIF4E in acute myeloid
leukemia (AML): a proof-of-principle clinical trial with ribavirin.
Blood. 114:257–260. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Matassa DS, Amoroso MR, Agliarulo I, et
al: Translational control in the stress adaptive response of cancer
cells: a novel role for the heat shock protein TRAP1. Cell Death
Dis. 4:e8512013. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Issur M, Bougie I, Despins S and Bisaillon
M: Enzymatic synthesis of RNAs capped with nucleotide analogues
reveals the molecular basis for substrate selectivity of RNA
capping enzyme: impacts on RNA metabolism. PLoS One. 8:e753102013.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Tan K, Culjkovic B, Amri A and Borden KL:
Ribavirin targets eIF4E dependent Akt survival signaling. Biochem
Biophys Res Commun. 375:341–345. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Mujtaba T and Dou QP: Advances in the
understanding of mechanisms and therapeutic use of bortezomib.
Discov Med. 12:471–480. 2011.PubMed/NCBI
|
|
29
|
Moriya S, Che XF, Komatsu S, et al:
Macrolide antibiotics block autophagy flux and sensitize to
bortezomib via endoplasmic reticulum stress-mediated CHOP induction
in myeloma cells. Int J Oncol. 42:1541–1550. 2013.PubMed/NCBI
|
|
30
|
Attar-Schneider O, Drucker L, Zismanov V,
Tartakover-Matalon S, Rashid G and Lishner M: Bevacizumab
attenuates major signaling cascades and eIF4E translation
initiation factor in multiple myeloma cells. Lab Invest.
92:178–190. 2012. View Article : Google Scholar
|
|
31
|
Graff JR and Zimmer SG: Translational
control and metastatic progression: enhanced activity of the mRNA
cap-binding protein eIF-4E selectively enhances translation of
metastasis-related mRNAs. Clin Exp Metastasis. 20:265–273. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Culjkovic B, Topisirovic I, Skrabanek L,
Ruiz-Gutierrez M and Borden KL: eIF4E promotes nuclear export of
cyclin D1 mRNAs via an element in the 3′UTR. J Cell Biol.
169:245–256. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Wang X, Yue P, Chan CB, et al: Inhibition
of mammalian target of rapamycin induces phosphatidylinositol
3-kinase-dependent and Mnk-mediated eukaryotic translation
initiation factor 4E phosphorylation. Mol Cell Biol. 27:7405–7413.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Livingstone M, Larsson O, Sukarieh R,
Pelletier J and Sonenberg N: A chemical genetic screen for mTOR
pathway inhibitors based on 4E-BP-dependent nuclear accumulation of
eIF4E. Chem Biol. 16:1240–1249. 2009. View Article : Google Scholar
|
|
35
|
Rong L, Livingstone M, Sukarieh R, et al:
Control of eIF4E cellular localization by eIF4E-binding proteins,
4E-BPs. RNA. 14:1318–1327. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Saad H, Bellé R, Morales J, et al:
Initiation factors eIF4: from sea urchin embryonic development to
chronic lymphocytic leukemia. J Soc Biol. 201:307–315. 2007.(In
French). View Article : Google Scholar
|
|
37
|
Saito Y, Tachibana I, Takeda Y, et al:
Absence of CD9 enhances adhesion-dependent morphologic
differentiation, survival, and matrix metalloproteinase-2
production in small cell lung cancer cells. Cancer Res.
66:9557–9565. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Hasegawa M, Furuya M, Kasuya Y, et al:
CD151 dynamics in carcinoma-stroma interaction: integrin
expression, adhesion strength and proteolytic activity. Lab Invest.
87:882–892. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Maecker HT: Human CD81 directly enhances
Th1 and Th2 cell activation, but preferentially induces
proliferation of Th2 cells upon long-term stimulation. BMC Immunol.
4:12003. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Zöller M: Tetraspanins: push and pull in
suppressing and promoting metastasis. Nat Rev Cancer. 9:40–55.
2009. View Article : Google Scholar
|
|
41
|
Jee BK, Park KM, Surendran S, et al:
KAI1/CD82 suppresses tumor invasion by MMP9 inactivation via TIMP1
up-regulation in the H1299 human lung carcinoma cell line. Biochem
Biophys Res Commun. 342:655–661. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Suzuki M, Tachibana I, Takeda Y, et al:
Tetraspanin CD9 negatively regulates lipopolysaccharide-induced
macrophage activation and lung inflammation. J Immunol.
182:6485–6493. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Dua K, Williams TM and Beretta L:
Translational control of the proteome: relevance to cancer.
Proteomics. 1:1191–1199. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Jain MK and Zoellner C: Role of ribavirin
in HCV treatment response: now and in the future. Expert Opin
Pharmacother. 11:673–683. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Culjkovic B and Borden KL: Understanding
and targeting the eukaryotic translation initiation factor eIF4E in
head and neck cancer. J Oncol. 2009:9816792009. View Article : Google Scholar
|
|
46
|
Tamburini J, Green AS, Chapuis N, et al:
Targeting translation in acute myeloid leukemia: a new paradigm for
therapy? Cell Cycle. 8:3893–3899. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Panfilio S, D’Urso P, Annechini G, et al:
Regression of a case of Multiple Myeloma with antiviral treatment
in a patient with chronic HCV infection. Leuk Res Rep. 2:39–40.
2013.PubMed/NCBI
|