Introduction
Breast cancer is the most common type of cancer in
women and is the second leading cause of cancer death following
lung cancer, resulting from coordinated actions between estrogen
receptor (ER) and growth factor receptor especially for HER2 and
various survival signaling pathways (1). Increasing efforts are conducted on
identifying not only agents that selectively target cancer cells
but also signaling pathways that promote or inhibit cancer
progression. Targeting a specific pathway is critical to successful
treatment of breast cancer, as cancer cells reflect the balance
between cell death and survival (2). ERs are a group of nuclear receptor
(NR) transcription factors, which could be activated by estrogen
hormone. There are two classical nuclear ER subtypes, ERα and ERβ.
Approximately 70% of breast cancer cases are ER-positive (3). Signaling of ERα is involved in normal
breast development, as well as in growth and progression of breast
cancer. HER2, an orphan receptor with intrinsic tyrosine kinase
activity, regulates cell growth, differentiation and survival
(4,5). Excess HER2 signaling leads to
numerous oncogenic processes including cell proliferation, survival
and carcinogenesis (6) via the
RAS/Raf1/MEK/ERK and the PI3K/Akt pathways. Moreover, crosstalk
between ERα and HER2 signaling pathways fundamentally contribute to
the development and aggressiveness of cancer (7).
Apoptosis is a form of programmed cell death which
is precisely regulated and plays important roles during
embryogenesis and immunology (8–11).
Apoptosis is characterized by a number of well-defined features
including cellular morphological changes, chromatin condensation,
DNA fragmentation and caspase activation. Apoptosis (program cell
death) proceeds through two main pathways, the extrinsic and
intrinsic pathways. The extrinsic pathway is triggered through the
binding of death ligands such as tumor necrosis factor-α (TNF-α),
cluster of differentiation 95 ligand (CD95L)/Fas ligand (FasL) or
TNF-related apoptosis-inducing ligand (TRAIL) to death receptors on
the cell surface resulting in formation of death-inducing signaling
complexes (DISC) and activation of initiator caspases-8 (12). On the contrary, the intrinsic or
mitochondrial pathway is initiated by cellular or DNA damage
characterized by depolarization of mitochondrial membrane e
controlled by the Bcl-2 family proteins leading to cytochrome
c release into cytoplasm and caspase-9 activation (13). Both pathways promote cleaved
caspase-3 expression then PARP degradation, DNA fragmentation and
apoptotic cell death. Apoptosis is regulated by a variety of
signaling pathways including the PI3K/Akt pathway. Protein kinase B
(Akt) has been shown to regulate apoptosis related proteins such as
B cell lymphoma-2 (Bcl-2), Bcl-2 associated X protein (Bax) and
cysteine aspartic acid specific protease (caspase-3) and is
crucially involved in anticancer drug induced apoptosis of cancer
cells (14–17).
MAPK signaling, targeted for cancer prevention and
treatment, includes a three-tiered kinase core, where MAP3K
activates MAP2K that activates MAPKs including ERK1/2 which
promotes cell growth, cell survival and differentiation whereas
JNK1/2 and p38 promote cell apoptosis (18,19).
Moreover, α-mangostin also showed decreased inactive-MAPK(s) by
dephosphorylation of c-Raf at Ser259. Therefore, downregulation of
phosphorylated-ERK1/2 may be an appropriate alternative therapy for
breast cancer patients (20).
Various signaling pathways were activated upon the
binding of estrogen to ERα including Ras-Raf-MAP kinase and
PI3K/Akt signaling pathways (21).
ERα phosphorylation is involved in tamoxifen resistance including
Ser104/106 and Ser118 in breast cancer patients (22). These regions of ERα were
phosphorylated by ERK1/2 and MEK1/2 (23,24).
ERK/MAPK and PI3K/Akt pathways rapidly activated by the
ERα-estrogen complex also have a critical role in estrogen action
as a survival agent. Furthermore, p38 phosphorylated-ERα (at
Ser294) in coordination with S-phase kinase-associated protein 2
(Skp2) which stimulated phosphorylation of ERα at Ser64 led to
proteosomal degradation of ERα. In addition, inhibition of p38 and
Skp2 affected the ERα function (25). Indeed, these pathways enhanced
Bcl-2 expression, blocked p38 activation and reduced caspase-3
activation (26,27).
Mangosteen (Garcinia mangostana Linn.) family
Guttiferae has been used for hundreds of years around the
world, mainly in Southeast Asia. Mangosteen is round, dark purple
or reddish fruit with white juicy pulp possessing slightly acidic
and sweet flavor, known as the ‘queen of fruits’ (28). The juicy pulp contains essential
nutrients such as carbohydrates, fat, protein, vitamins and trace
metals. The fruit hulls were used as a traditional herbal medicine
for the treatment of abdominal pain, dysentery, wound infections,
eczema, suppuration and chronic ulcer (29–31).
Xanthones, the most abundant polyphenolic compounds present in
mangosteen pericarp, have long been reported to possess multiple
health-promoting properties (32).
The dominant xanthone extracted from the fruit hulls of G.
mangostana, α-mangostin, has been demonstrated to possess
antioxidant (33–35), antibacterial (36,37),
antifungal (38),
anti-inflammatory (39,40), renoprotective activities (41). Effects of α-mangostin have been
reported in numerous preclinical tumor models such as colon cancer
(42), colorectal (43), leukemia (44), chondrosarcoma (45).
In the present study, we investigated the hypothesis
that α-mangostin could inhibit cell proliferation and induce
apoptosis associated with HER2/PI3K/Akt and MAPK signaling pathway
in human breast carcinoma T47D cells.
Materials and methods
Chemical
Hoechst 33342, fetal bovine serum (FBS), crystal
violet, 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium
bromide (MTT), phenylmethylsulphonylfluoride (PMSF),
2-(4-(2-hydroxyethyl)-1-piperazinyl)-ethanesulphonic acid (HEPES),
ethylenediaminetetraacetic acid (EDTA), Ethylene glycol-bis
(2-aminoethylether)-N,N,N′,N′-tetraacetic acid (EGTA) and
phenol:chloroform:isoamyl alcohol (25:24:1) were purchased from
Sigma-Aldrich (St. Louis, MO, USA). Dulbecco's modified Eagle's
medium (DMEM) and Roswell Park Memorial Institute (RPMI)-1640
medium were purchased from Gibco (Grand Island, NY, USA). Propidium
iodide (PI), Hoechst 33342 (2,5′-Bi-1H-benzimidazole,
2′-(4-ethoxyphenyl)-5-(4-methyl-1-piperazinyl)-23491-52-3), JC-1
(5,5′,6,6′-tetrachloro-1,1′,3,3′-tetraethyl-imidacarbocyanine
iodide) and DNase free RNase A were purchased from Thermo Fisher
Scientific Inc. (Pittsburgh, PA, USA). Dimethylsulfoxide (DMSO) was
purchased from Merck Calbiochem (San Diego, CA, USA). α-Mangostin
was isolated and purified by A. Krajarng according to our previous
report (45). The structure of
G. mangostana is shown in Fig.
1A (46).
Cell culture
Breast cancer cell lines MDA-MB-468, AU565, SKBR3
and T47D were obtained from the American Type Culture Collection
(ATCC; Manassas, VA, USA). MDA-MB-468 was maintained as a monolayer
in DMEM. AU565, SKBR3 and T47D were maintained as a monolayer in
RPMI-1640. The media were supplemented with 10% fetal bovine serum
(FBS) 100 U/ml penicillin, 100 μg/ml streptomycin (PAA
Laboratories, Pasching, Austria). Cells were cultured in 5%
CO2 at 37°C. The medium was refreshed every 2–3
days.
Evaluation of proliferation inhibitory
potential (MTT assay)
The cytotoxicity of α-mangostin was determined by
MTT assay. MDA-MB-468, AU565, SKBR3 and T47D cells were seeded at a
density of 1×104 cells/well in a 96-well plate and
allowed to grow for 24 h. Cells were then treated with α-mangostin
at 7.5, 15 and 30 μM, whereas the control group was treated with
0.03% DMSO. After incubation for 4, 8, 12, 16, 20 and 24 h, MTT
solution (0.5 mg/ml) was added to each well and was further
incubated for 3 h at 37°C. DMSO was added to each well to
solubilize water insoluble purple formazan crystals. The absorbance
at 570 nm was measured using a microplate reader (Multiskan EX;
Thermo Electron Corp., Vantaa, Finland). Survival percentage (%)
was calculated relative to the control.
The IC50 of T47D cells was 7.5 μM, which
was less than Vero cells (14.26 μM), whereas those of SKBR3,
MDA-MB-468 and AU565 were 23.88, 22.23 and 43.63 μM, respectively.
Thus, T47D cells were chosen for further study. The reason for
choosing different times depended on which step of apoptosis occurs
at each period, as each protein will be expressed at different
times. The survival proteins were reduced with increasing of
time.
Colony formation assay
T47D cell suspensions were seeded at a density of
4×104 cells/well in a 35-mm tissue culture dish and
allowed to grow for 24 h at 37°C. Cells were treated with 15 and 30
μM α-mangostin for 3 and 6 h. After 14 days colonies were fixed
with 100% methanol and stained with 2% (w/v) crystal violet
(Suvchem Laboratory Chemicals, Mumbai, India). The number of
colonies ≥50 cells was counted and colony formation efficiency was
calculated.
Nuclear morphology staining with Hoechst
33342
T47D cells were seeded at a density of
5×104 cells/dish for 24 h then treated with 30 μM
α-mangostin for 3, 6, 9 and 12 h at 37°C. The cells were then
stained with 10 μM Hoechst 33342 and examined under fluorescence
microscope (IX73; Olympus, Tokyo, Japan).
DNA fragmentation analysis
T47D cells at 2×106 cells/well were
seeded in 60-mm tissue culture dish and treated with 0, 15 and 30
μM α-mangostin for 24 h. For DNA fragmentation assay, low molecular
weight DNA was isolated (47).
Briefly, T47D cells treated with α-mangostin were collected, washed
and resuspended in TE (10 mM Tris-HCl, pH 8.0 and 1 mM EDTA)
buffer. Then lysis buffer (5 mM Tris-HCl, pH 8.0 and 20 mM EDTA and
0.5% (v/v) Triton X-100) was added, resuspended and kept at −20°C
overnight. The mixture was centrifuged at 14,000 × g for 15 min.
The supernatant was then collected and extracted with
phenol:chloroform:isoamyl alcohol (25:24:1) followed by butanol and
precipitated with absolute ethanol. The pellet was dried and
dissolved in TE buffer, incubated with 100 μg/ml DNase free RNase A
for 2 h at 37°C and incubated with 200 μg/ml proteinase K (Vivantis
Inc., Oceanside, CA, USA) at 60°C. The sample DNA was analyzed
using 1.8% agarose gel (Vivantis) and visualized by ethidium
bromide staining.
Cell cycle analysis
T47D cells were seeded at 6×105
cells/well in a 6-well plate then treated with 0, 15 and 30 μM
α-mangostin for 12 h, washed with ice cold PBS, fixed with 70%
ethanol and stored at −20°C then resuspended in DNA staining
solution (50 μg/ml propidium iodide, 200 μg/ml DNase free RNase A,
0.1% Triton X-100, 0.1% sodium citrate). Cells were washed and
incubated at 4°C overnight in the dark and analyzed for DNA content
on a BD Accuri C6 flow cytometer (Accuri Cytometers, Inc., Ann
Arbor, MI, USA) at 488 nm excitation. The forward scatter and red
fluorescence above 600 nm were measured.
Measurement of mitochondrial membrane
potential (ΔΨm)
The changes in mitochondrial membrane potential
(ΔΨm) were determined using JC-1, a lipophilic fluorescent cation
that incorporates into the mitochondrial membrane. Intact living
cells stained with JC-1 exhibits pronounced red fluorescence,
whereas break-down of the mitochondrial membrane potential in
apoptotic cells shows decreased red fluorescence and reveals green
fluorescence. Cells were treated with 30 μM α-mangostin and the
control cells were treated with 0.03% DMSO, and then were stained
with 10 μg/ml JC-1.
Detection of cytochrome c release
Upon treatment of T47D cells in the absence or
presence of 30 μM α-mangostin, cells were resuspended in S-100
lysis buffer (20 mM HEPES, pH 7.5, 10 mM KCl, 1.5 mM
MgCl2, 1 mM EGTA, 1 mM EDTA and 250 mM sucrose) and
homogenized for 30–40 strokes. Cell were centrifuged at 500 × g for
5 min at 4°C (to eliminate nuclei and unbroken cells). The
supernatant was taken to a new tube, then centrifuged at 10,000 × g
for 30 min at 4°C. Pellet contained mitochondria and supernatant
contained cytosolic fraction. The protein was separated by 12%
sodium dodecyl sulphate polyacrylamide gel electrophoresis
(SDS-PAGE), transferred onto polyvinylidene fluoride (PVDF)
membranes (GE Healthcare, Buckinghamshire, UK) and subjected to
immunodetection of cytochrome c using a rabbit polyclonal
antibody against human cytochrome c (Cell Signaling
Technology, Beverly, MA, USA). The peroxidase activity of bound
secondary antibodies on the blots was detected by enhanced
chemiluminescence reagent (ECL; GE Healthcare) and detected under
chemiluminescent imaging system (GeneGnome gel documentation;
Synoptics Ltd., Cambridge, UK).
Western blot analysis
T47D cells were seeded at 6×105
cells/well in a 35-mm culture dish, exposed with 30 μM α-mangostin
and harvested at designated time-points. Cells were lysed with RIPA
(radioimmunoprecipitation assay) buffer (50 mM Tris-HCl, pH 7.5, 5
mM EDTA, 250 mM NaCl, 0.5% Triton X-100) supplemented with complete
mini-protease inhibitor cocktail (Roche Diagnostics GmbH, Mannheim,
Germany). The protein content was determined using Bio-Rad protein
assay kit (Bio-Rad Laboratories, Hercules, CA, USA), separated by
10–12% SDS-PAGE and transferred onto PVDF membrane using a Mini
Trans-Blot Cell® (Bio-Rad Laboratories). The blots were
incubated overnight at 4°C with appropriate primary antibodies
(Cell Signaling Technology). The membranes were washed and
incubated with appropriate secondary antibodies conjugated with
horseradish peroxidase (Cell Signaling Technology) for 1 h at room
temperature. Immunoreactive protein bands were detected by
chemiluminescence using ECL.
Statistical analysis
All data presented were obtained from at least three
independent experiments and are presented as mean ± standard
deviation (SD). Statistical significance was assessed by one-way
analysis of variance (ANOVA). Statistical analysis was performed
using the SPSS statistical software package (version 11.5) as well
as GraphPad Prism 3.03 (GraphPad Software, Inc., San Diego, CA,
USA). The protein band intensity was quantified by ImageJ
densitometer. Asterisks in the figures indicate that the
experimental values are significantly different from those of the
control (*P<0.05, **P<0.01).
Results
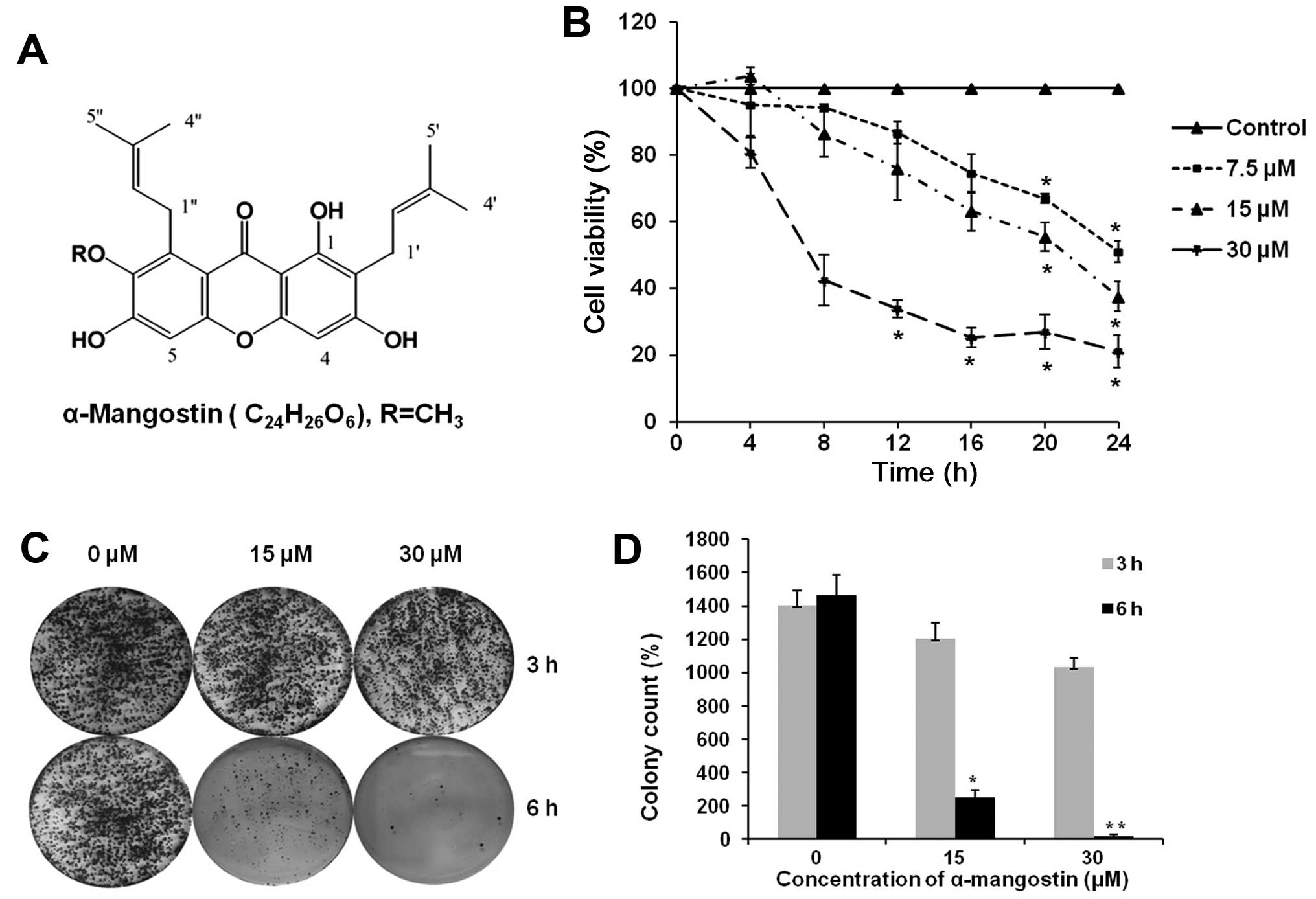
α-Mangostin decreases cell viability in
T47D human breast cancer cells
The effect of α-mangostin on cell viability in T47D
cells treated with α-mangostin at various concentrations and times
was determined and the results showed that the IC50
value was 7.5±0.5 μM at 24 h (Fig.
1A), and cell viability was dose- and time-dependent comparing
to the control cells.
α-Mangostin inhibits colony formation and
proliferation of breast cancer cells
The results indicated that α-mangostin inhibited
colony formation in T47D cells in a time- and dose-dependent manner
(Fig. 1B). T47D cells treated with
30 μM α-mangostin for 6 h grew slightly, but less than at 15 μM
α-mangostin. The quantitative data of colony formation assay were
calculated and compared with untreated control cells in three
independent experiments (Fig. 1C and
D).
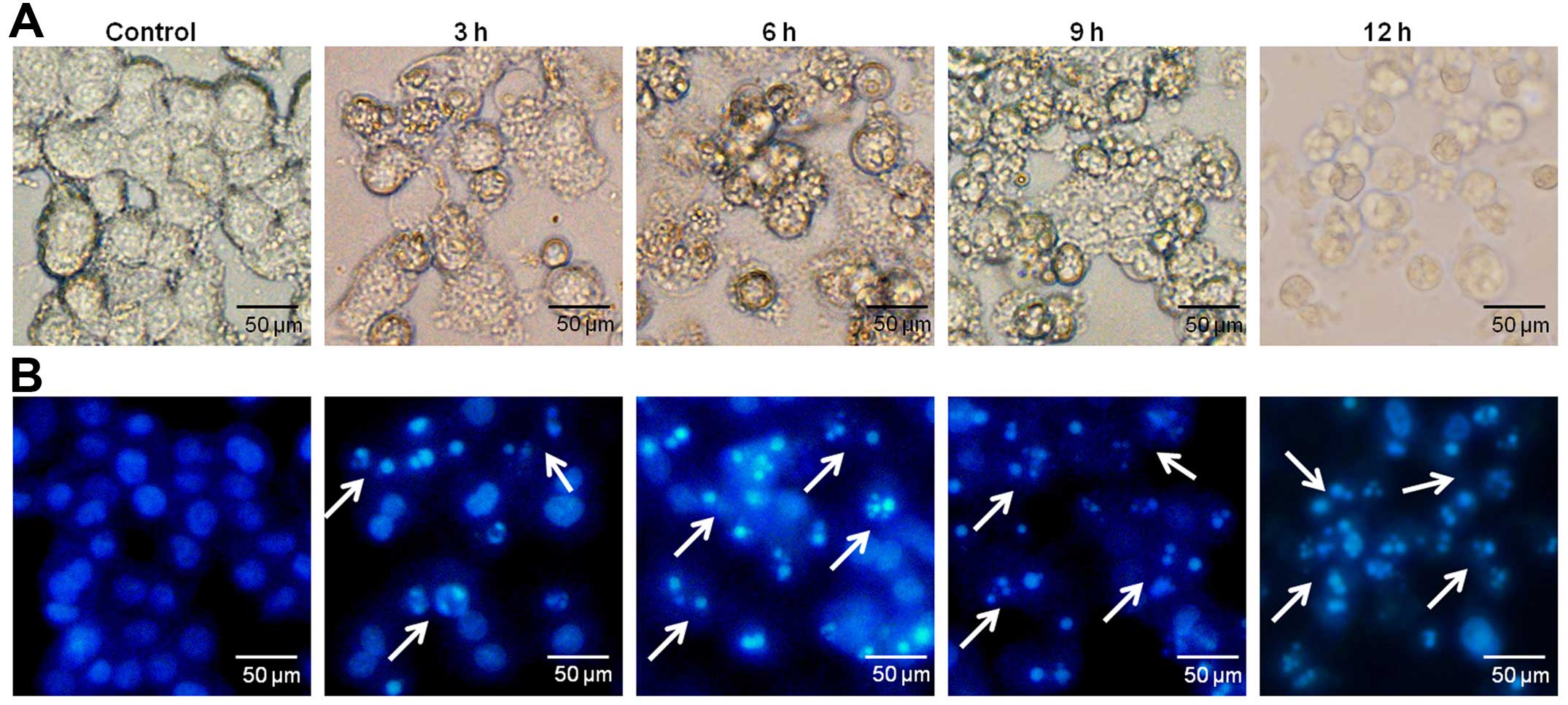
Morphological and nuclei changes
Treatment of T47D cells with 30 μM α-mangostin for
3, 6, 9 and 12 h showed evident morphological changes including
vacuolization, cell shrinkage and rounding (Fig. 2A). The result also showed chromatin
condensation, a characteristic of apoptotic cells (Fig. 2B).
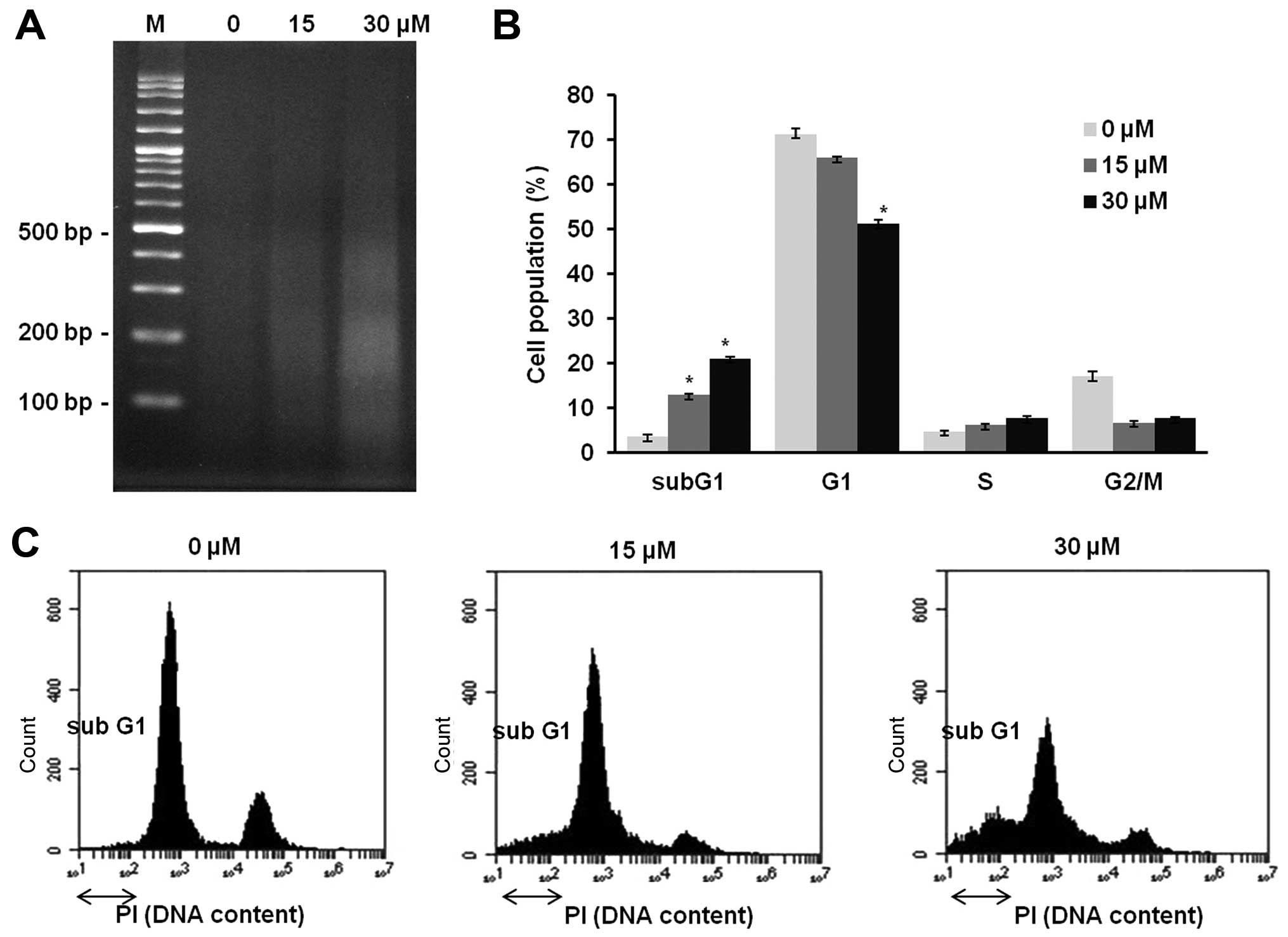
Cell cycle distribution and chromosomal
DNA fragmentation
DNA fragmentation, a ladder-like pattern, typical
DNA cleavage between nucleosome was visible dose-dependently after
incubation with α-mangostin, whereas, not in the control (Fig. 3A). The effect of α-mangostin on
cell cycle distribution showing two major peaks represented the
G0/G1 and G2/M phases, the sub G1 peak with reduced DNA content
represented apoptotic cells. The mean apoptotic population of T47D
cells was 3.3% under the control condition as compared to 12.6 and
21.0% upon treatment with 15 and 30 μM α-mangostin for 12 h,
respectively (Fig. 3B and C).
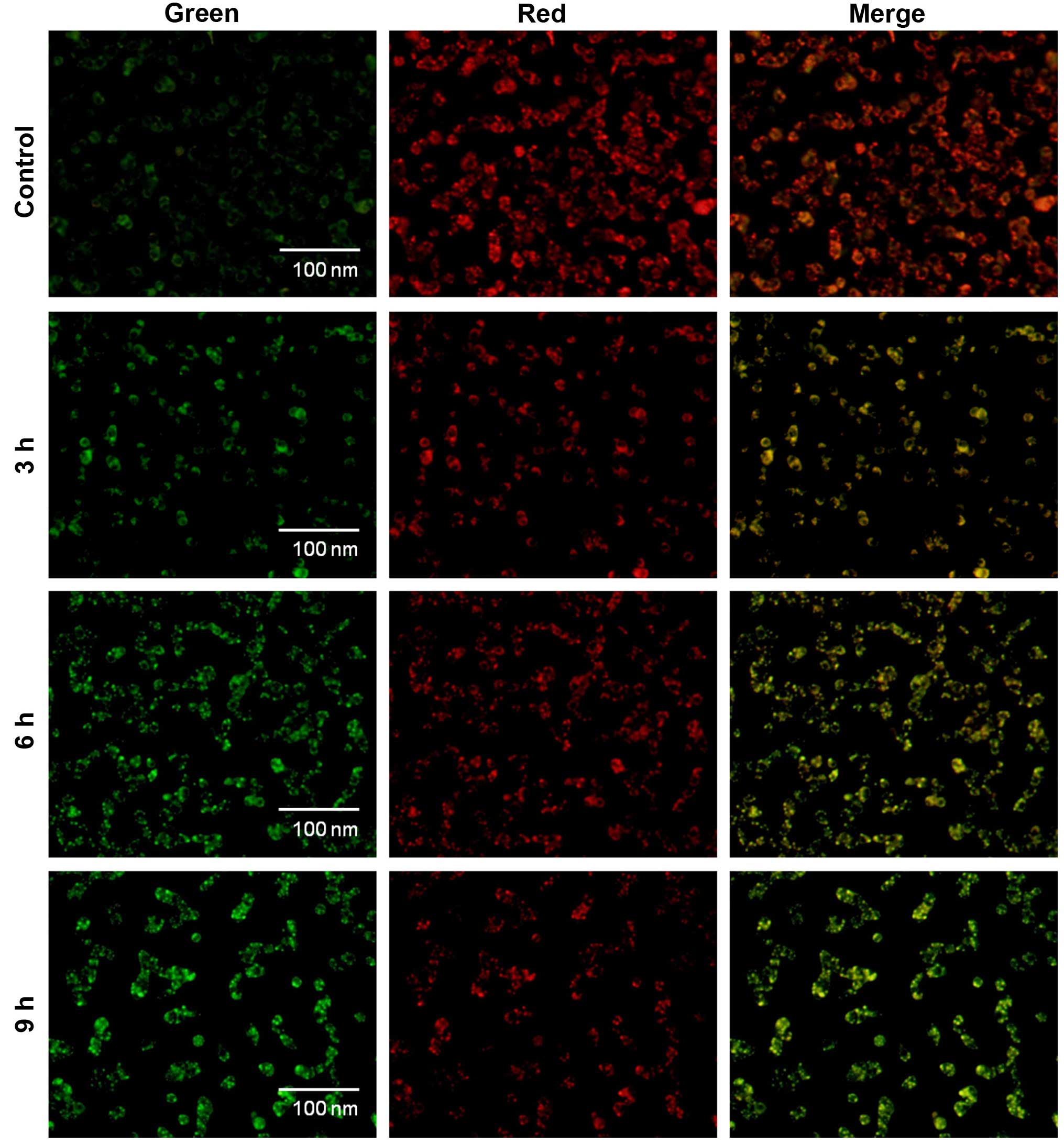
Effects of α-mangostin on the
mitochondrial membrane potential (ΔΨm)
Bax oligomers increased mitochondrial membrane
permeability resulting in cytochrome c release to the
cytosol. JC-1 staining was used to determine ΔΨm of T47D cells. At
higher membrane potentials or healthy cells, JC-1 forms a complex
known as J-aggregates with intense red fluorescence. In contrast,
in apoptotic or unhealthy cells with low ΔΨm, JC-1 remains in the
monomeric form with intense green fluorescence. The increased
intensity ratio of green to red fluorescence indicated
mitochondrial depolarization. The increased green/red ratio was
detected in T47D cells treated with 30 μM α-mangostin for 3, 6 and
9 h, while the control cells showed red fluorescence (Fig. 4) indicating that α-mangostin
induced loss of ΔΨm in T47D cells.
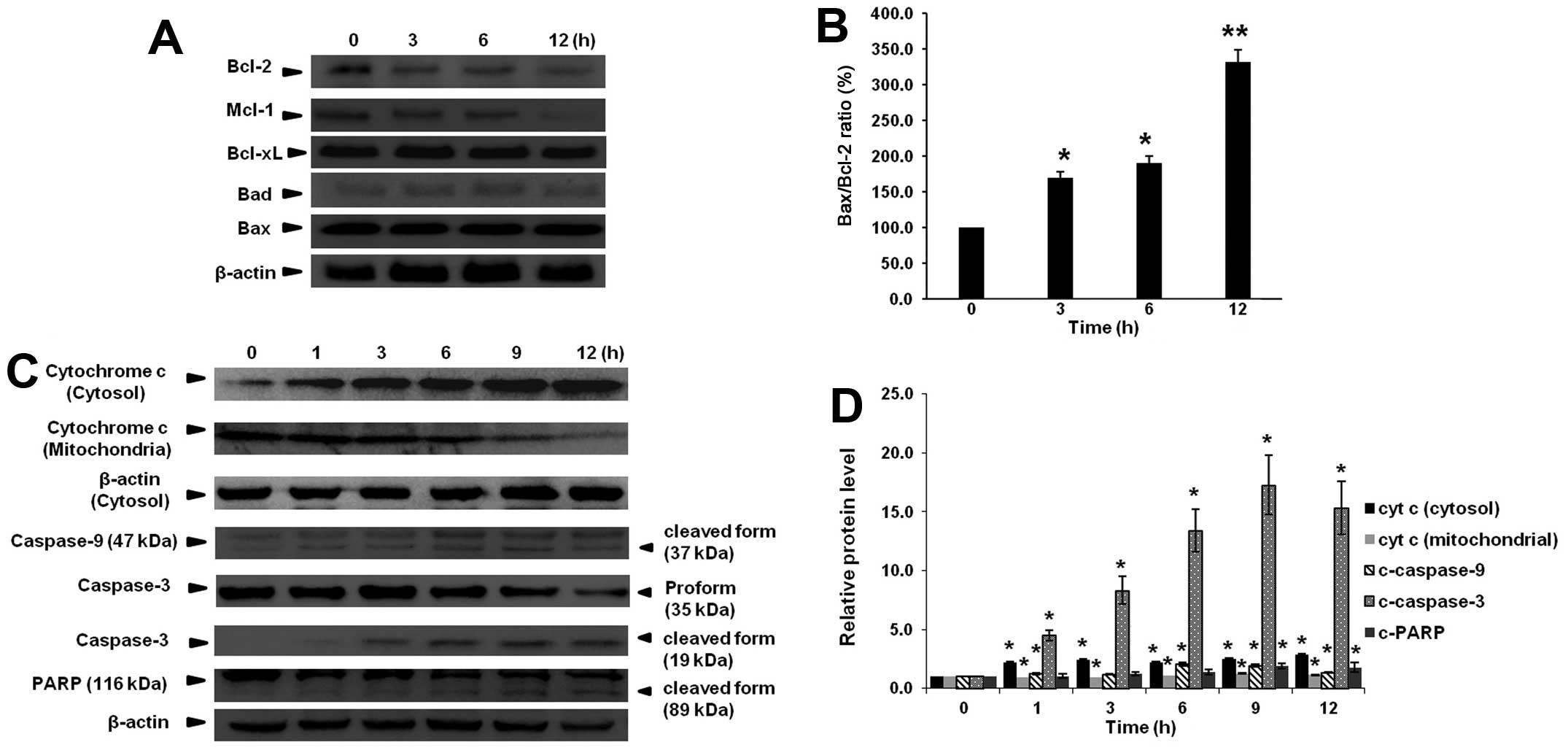
Effects of α-mangostin on the expression
of Bcl-2 family proteins and induction of cytochrome c release
The Bcl-2 family is a group of proteins regulating
cell survival and cell death. The insertion of Bax into
mitochondrial membrane induces the opening of the mitochondrial
voltage-dependent anion channel (VDAC). The results showed that
α-mangostin decreased myeloid cell leukemia (Mcl-1) expression but
not B-cell lymphoma-extra large (Bcl-xL) and Bcl-2-associated death
promoter (Bad) expression (Fig.
5A). The Bax/Bcl-2 ratio was increased significantly in each
α-mangostin treated group comparing to the control (Fig. 5B). Bax oligomers increased
mitochondrial membrane permeability and cytochrome c release
from mitochondria to cytosol in a time-dependent pattern (Fig. 5C).
Effects of α-mangostin on caspase-3,
caspase-9 and cleaved PARP activation
The expression of active forms of caspases was
determined by western blot analysis. As shown in Fig. 5C, cleaved caspase-3 and cleaved
caspase-9 were induced upon exposure to α-mangostin. Thus,
α-mangostin-induced apoptosis was mediated by caspase-3 and may be
associated with the activation of intrinsic pathway (via
caspase-9). In addition, apoptosis induction by α-mangostin was
accompanied by expression of the apoptosis marker, cleaved PARP
(Fig. 5C).
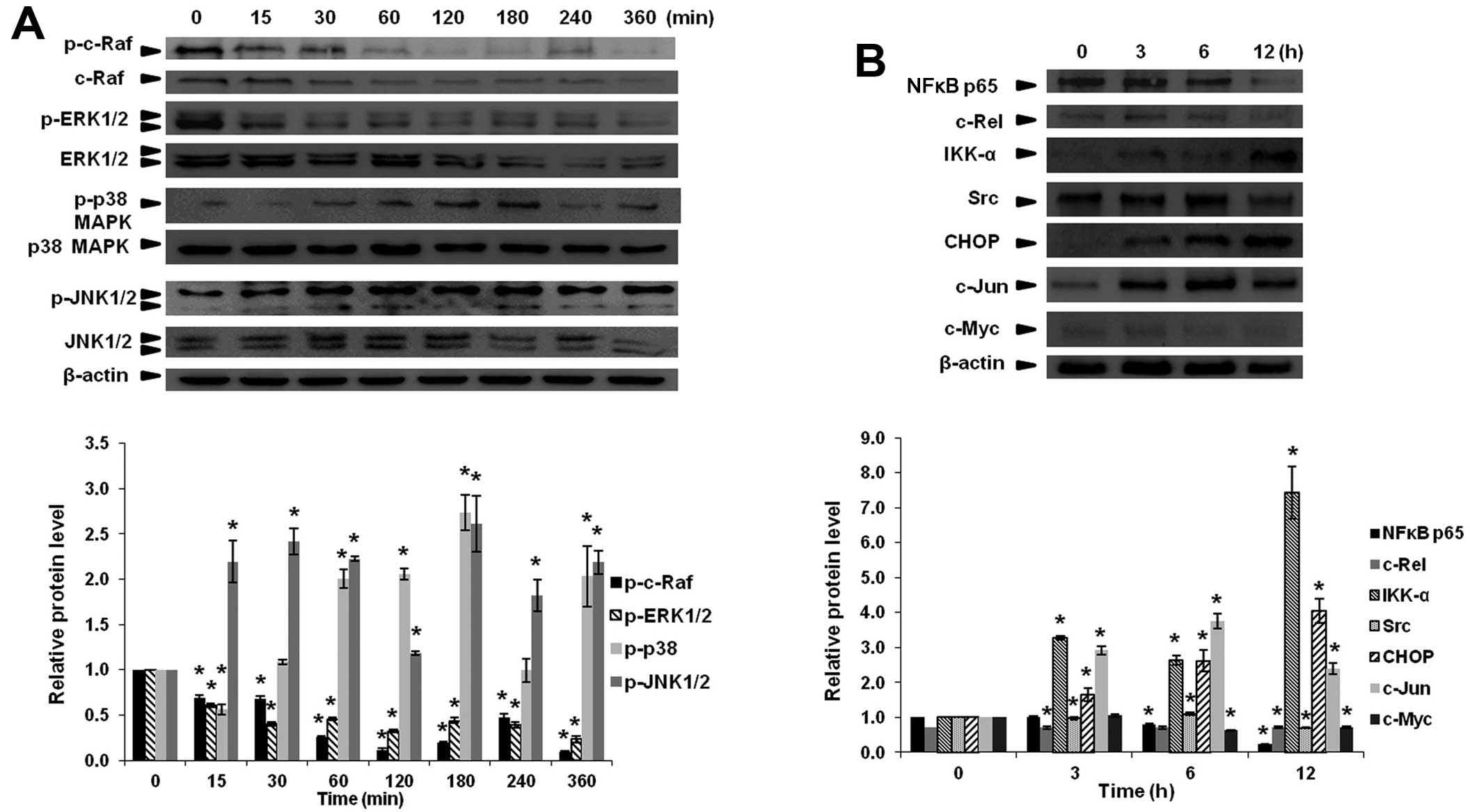
Modulation of MAPK signaling during
apoptosis induction by α-mangostin
MAPK are important signaling components that control
cellular proliferation, differentiation, motility, survival and
apoptosis. The results showed that α-mangostin induced
phosphorylated-p38 expression at 30 min and the highest level was
detected at 2 h. In addition, phosphorylated-JNK1/2 was induced at
15 min and the highest level was at 6 h. Furthermore, decreased
phosphorylated-ERK1/2 at 15 min was detected. Phosphorylated-c-Raf
was significantly decreased at 15 and 30 min (Fig. 6A). Once activated,
phosphorylated-c-Raf activated the dual specific protein kinases
mitogen-activated protein kinase kinase 1 (MEK1) and MEK2, which in
turn activated the serine/threonine-specific protein kinase ERK1/2.
In summary α-mangostin modulated the MAPK(s) pathway resulting in
apoptosis induction.
Evaluation of transcription factor and
downstream oncogenic products of PI3K/Akt and MAPK
α-Mangostin enhanced IκB kinases α (IKKα) and Src
expression whereas decreased nuclear transcription factor kappa B
(NF-κB p65) and c-Rel expression (Fig.
6B). This could result in cell proliferation inhibition and
inflammation in cancer cells as well as increased sensitivity to
antitumor agent and ultimately apoptotic cell death (48,49).
Moreover, α-mangostin also activated C/EBP homologous protein
(CHOP) and c-Jun expression, while suppressed c-Myc expression
(Fig. 6B).
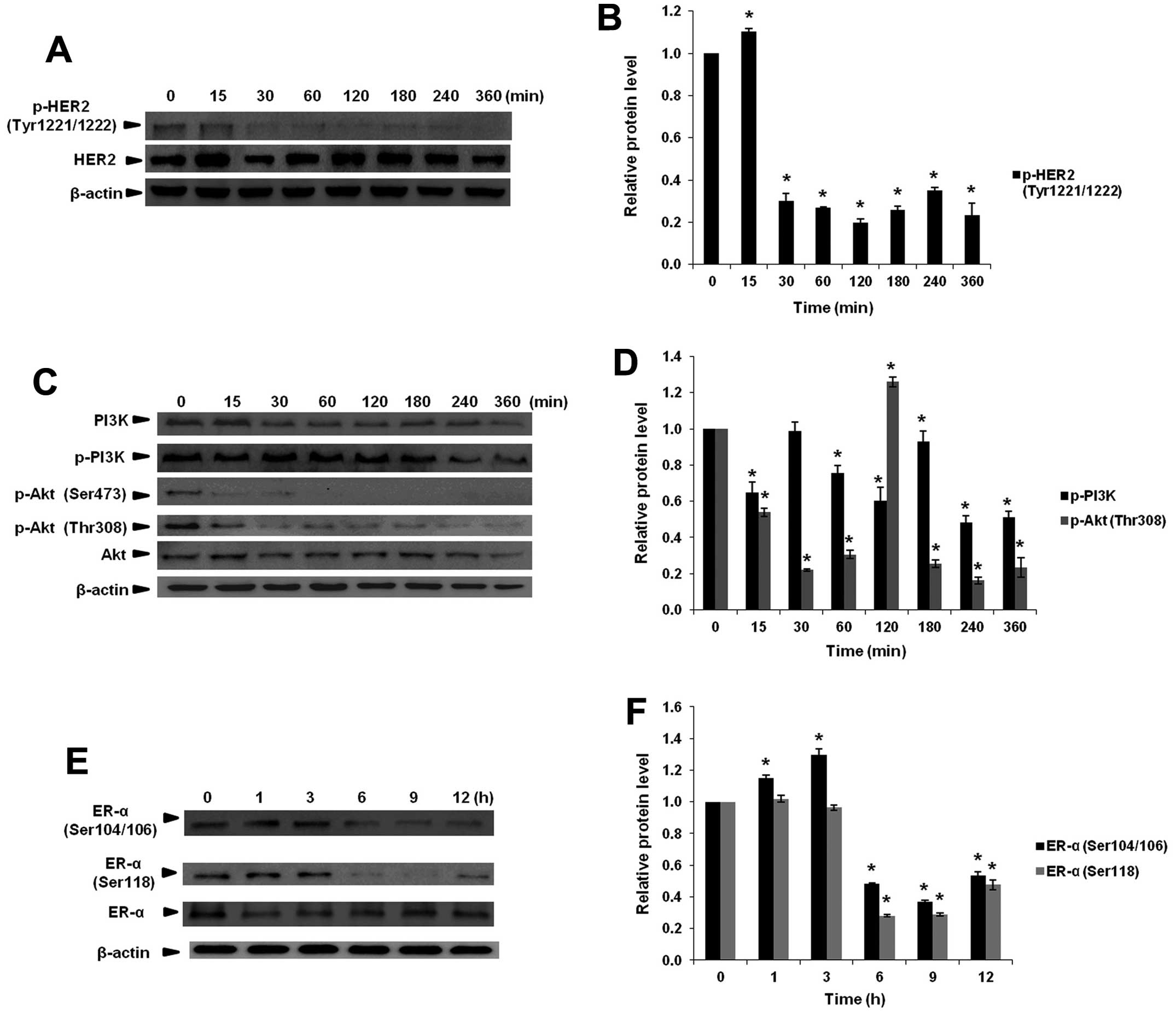
α-Mangostin inhibits HER2 activation and
downstream PI3K/Akt signaling pathway
Activation of the HER2 network results in
autophosphorylation of C-terminal tyrosine leading to the
recruitment of cytoplasmic signal transducer that regulates
cellular processes such as proliferation, apoptosis inhibition and
transformation. Therefore, we examined whether α-mangostin could
activate dephosphorylation of phosphorylated-HER2 (p-HER2).
Treatment of T47D cells with 30 μM α-mangostin resulted in
substantially decreased phosphorylated-HER2 Tyr1221/1222 at 30 min
(Fig. 7A), which led to
inactivation of RAS/Raf1/MEK/ERK and PI3K/Akt transduction cascade.
HER2 stimulates tumor cell growth and renders cellular
chemo-resistance involved with the HER2 tyrosine kinase domain.
PI3K/Akt signaling is the major
anti-apoptosis pathway that confers survival advantage and drug
resistance of cancer cells
Our data clearly indicated the inhibition of
PI3K/Akt signaling cascade by α-mangostin in T47D cells (Fig. 7B). In addition, Akt phosphorylation
at Ser473 and Thr308 was highly suppressed by α-mangostin resulting
in inhibition of cell proliferation and apoptosis induction.
Effect of α-mangostin on inhibition of
ERα activation
ERα is a member of NR family transcription factors
that is involved in cell proliferation and survival. Once activated
by estrogen, the ERα is able to translocate into the nucleus and
bind to specific DNA response elements, known as estrogen response
elements (EREs), to regulate different gene activities. The results
showed the decreased phosphorylation of Ser104/106 and Ser118 of
ERα upon treatment with 30 μM α-mangostin at 6 h (Fig. 7C) implying that α-mangostin could
inhibit cell proliferation and cell survival through the
suppression of ERα phosphorylation in T47D cells.
Discussion and Conclusion
The goal of the present study was to search for a
natural bioactive compound from a plant as anti-breast cancer agent
with high specificity to ER and progesterone receptor (PR) positive
T47D breast cancer cells. Approximately 75% of all breast cancers
are ER-positive and 65% are PR-positive cells. Mammary ductal
carcinoma is the most common type of breast cancer in women with
poor prognosis, resistant to chemotherapy and radiation treatment.
Apoptosis-inducing agents are being investigated as a tool for the
management of cancer treatment. α-Mangostin, one of the xanthones,
is the secondary metabolite from mangosteen pericarp possessing
various biological activities (35). For this reason, we investigated the
potential of α-mangostin for human breast carcinoma cell treatment.
The results showed that α-mangostin could inhibit cell growth in a
time- and dose-dependent manner with an IC50 value of
7.5±0.5 μM.
The present study showed that α-mangostin decreased
Mcl-1 expression (Fig. 5A) as well
as increased Bax/Bcl-2 ratio (Fig.
5B). The cytosolic cytochrome c level was increased
corresponding to the loss of ΔΨm (Fig.
5C). Cytosolic cytochrome c activated procaspase-9 by
binding to apoptotic protease activating factor 1 (Apaf1) and
activated downstream effector caspase (including caspase-3) then
triggered apoptosis induction through the intrinsic pathway.
Our results illustrated that α-mangostin modulated
MAPK(s) (Fig. 6A) and inactivated
ERK1/2 resulting in decreased NF-κBp 65, c-Rel and c-Myc but
increased IKKα expression, whereas, activation of p38 and JNK1/2
led to CHOP and c-Jun expression, respectively. A limitation of the
study at the present stage, is that further experiments with
inhibitors are required. There are many related studies including
phosphorylation of c-Jun leading to activator protein 1 (AP-1)
formation which involved pro-apoptotic protein transcription and
apoptosis induction (50). The
JNK1/2 played a key role in CHOP induction and cell death by ER
stress in rat hepatocytes (51).
Moreover, CHOP/death receptor 5 (DR5) upregulation was mediated via
JNK1/2 phosphorylation in α-TEA induced apoptosis in MDA-MB-231 and
MCF-7 cells (52). Our results
showed rapid cell growth inhibition and apoptosis induction in T47D
cells by α-mangostin at 15 min. α-Mangostin also decreased p-HER2
at Tyr1221/1222 as compared with proform-HER2 expression (Fig. 7A) leading to downregulation of
Ras-Raf-MAP kinase and PI3K/Akt transduction pathways (53). PI3K/Akt signaling pathway is
essential for cell survival and the expression of a constitutively
active PI3K/Akt pathway induces multidrug resistance and prevents
apoptosis in a variety of cell types. Our results showed that
α-mangostin could decrease the phosphorylation of PI3K and Akt at
Ser473 and Thr308 (Fig. 7B).
Interestingly, we found that p-Akt at both sites were decreased
significantly suggesting that Akt may be a target of α-mangostin in
breast cancer. Previous studies showed that inhibition of the
PI3K/Akt pathway with doxorubicin, trastuzumab, etoposide,
tamoxifen increased apoptosis induction implying that measuring Akt
activity would directly be beneficial to breast cancer patients
(54,55). Thus, our results implied that
inhibition of the PI3K/Akt pathway by α-mangostin may be of great
benefit when combining with traditional cytotoxic chemotherapy,
immune therapy and endocrine based therapy.
In addition, estrogen is also involved in normal
breast development as well as in breast cancer growth and
progression. The biological actions of estrogen are mediated by
binding to nuclear ERα and ERα. Treatment with ER antagonists is
the current hormone therapy of choice for ERα-positive breast
cancer treatment. In the present study, we found that α-mangostin
could inhibit ERα phosphorylation in T47D cells at 6 h.
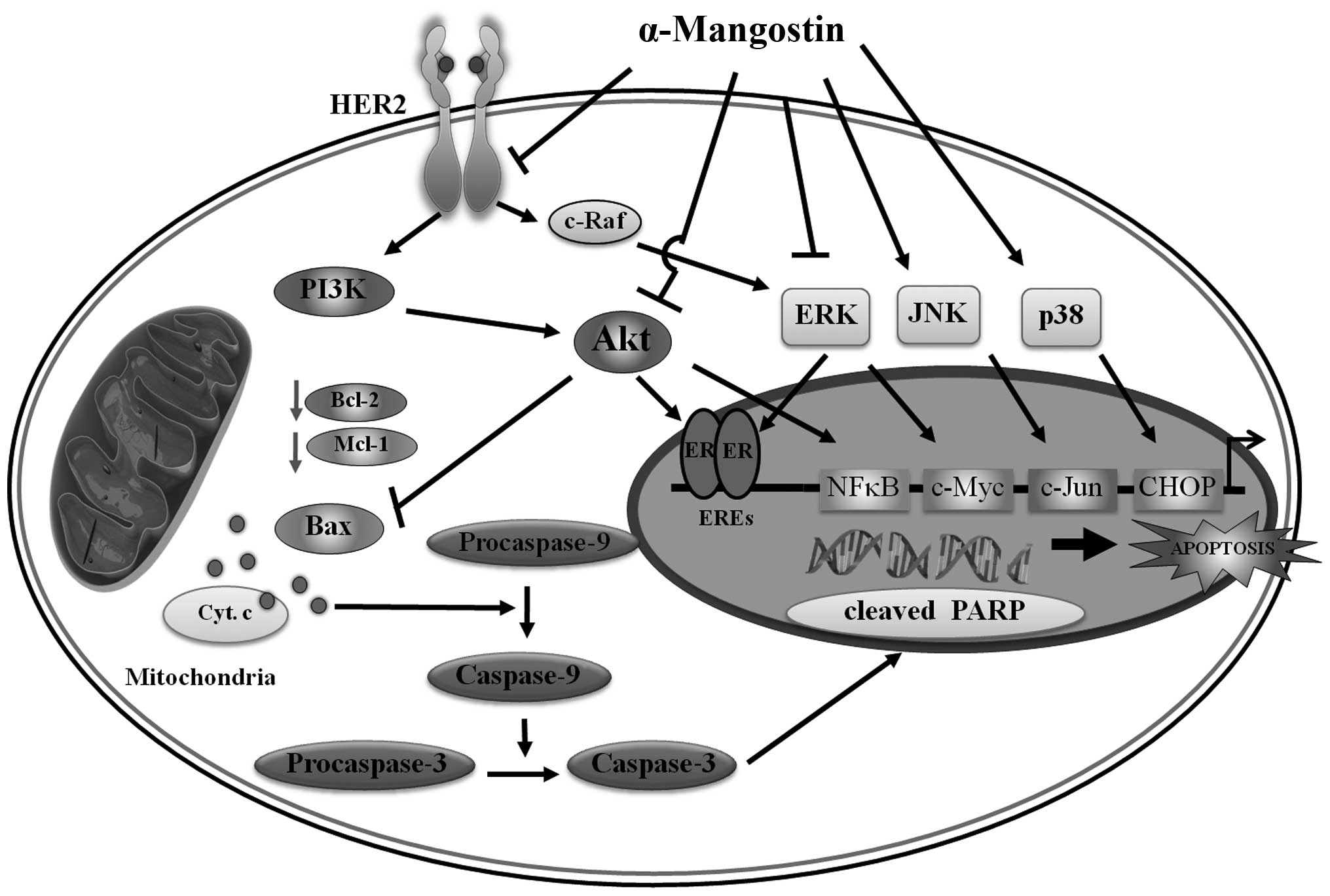
In conclusion, α-mangostin inhibited cell
proliferation and induced apoptosis associated with HER2/PI3K/Akt
and MAPK signaling pathway in human breast carcinoma T47D cells
(Fig. 8). Therefore, α-mangostin
may provide anticancer action in clinical application in human
breast cancer treatment.
Acknowledgements
We would like to thank the Royal Golden Jubilee
Ph.D. Program, Thailand Research Fund (grant no. PHD/0312/2550),
the Strategic Wisdom and Research Institute, Srinakharinwirot
University and the Research Division, Faculty of Medicine,
Srinakharinwirot University. We would also like thank Dr Aungkana
Krajarng for her help in α-mangostin preparation.
References
|
1
|
Osborne CK, Shou J, Massarweh S and Schiff
R: Crosstalk between estrogen receptor and growth factor receptor
pathways as a cause for endocrine therapy resistance in breast
cancer. Clin Cancer Res. 11:865s–870s. 2005.PubMed/NCBI
|
|
2
|
Parton M, Dowsett M and Smith I: Studies
of apoptosis in breast cancer. BMJ. 322:1528–1532. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Hanstein B, Djahansouzi S, Dall P,
Beckmann MW and Bender HG: Insights into the molecular biology of
the estrogen receptor define novel therapeutic targets for breast
cancer. Eur J Endocrinol. 150:243–255. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Yarden Y: Biology of HER2 and its
importance in breast cancer. Oncology. 61(Suppl 2): 1–13. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Yamamoto T, Ikawa S, Akiyama T, Semba K,
Nomura N, Miyajima N, Saito T and Toyoshima K: Similarity of
protein encoded by the human c-erb-B-2 gene to epidermal growth
factor receptor. Nature. 319:230–234. 1986. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Hynes NE and Stern DF: The biology of
erbB-2/neu/HER-2 and its role in cancer. Biochim Biophys Acta.
1198:165–184. 1994.PubMed/NCBI
|
|
7
|
Arpino G, Wiechmann L, Osborne CK and
Schiff R: Crosstalk between the estrogen receptor and the HER
tyrosine kinase receptor family: Molecular mechanism and clinical
implications for endocrine therapy resistance. Endocr Rev.
29:217–233. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Baselga J: Treatment of
HER2-overexpressing breast cancer. Ann Oncol. 21(Suppl 7):
vii36–vii40. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Kim KC, Kim JS, Son JK and Kim IG:
Enhanced induction of mitochondrial damage and apoptosis in human
leukemia HL-60 cells by the Ganoderma lucidum and Duchesnea
chrysantha extracts. Cancer Lett. 246:210–217. 2007. View Article : Google Scholar
|
|
10
|
Mow BM, Blajeski AL, Chandra J and
Kaufmann SH: Apoptosis and the response to anticancer therapy. Curr
Opin Oncol. 13:453–462. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Kim R: Recent advances in understanding
the cell death pathways activated by anticancer therapy. Cancer.
103:1551–1560. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Mellier G, Huang S, Shenoy K and Pervaiz
S: TRAILing death in cancer. Mol Aspects Med. 31:93–112. 2010.
View Article : Google Scholar
|
|
13
|
Chipuk JE, Moldoveanu T, Llambi F, Parsons
MJ and Green DR: The BCL-2 family reunion. Mol Cell. 37:299–310.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Li D, Qu X, Hou K, Zhang Y, Dong Q, Teng
Y, Zhang J and Liu Y: PI3K/Akt is involved in bufalin-induced
apoptosis in gastric cancer cells. Anticancer Drugs. 20:59–64.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Takeuchi H, Kim J, Fujimoto A, Umetani N,
Mori T, Bilchik A, Turner R, Tran A, Kuo C and Hoon DS: X-Linked
inhibitor of apoptosis protein expression level in colorectal
cancer is regulated by hepatocyte growth factor/C-met pathway via
Akt signaling. Clin Cancer Res. 11:7621–7628. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Lee SM, Lee CT, Kim YW, Han SK, Shim YS
and Yoo CG: Hypoxia confers protection against apoptosis via
PI3K/Akt and ERK pathways in lung cancer cells. Cancer Lett.
242:231–238. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Bak Y, Kim H, Kang JW, Lee DH, Kim MS,
Park YS, Kim JH, Jung KY, Lim Y, Hong J, et al: A synthetic
naringenin derivative, 5-hydroxy-7,4′-diacetyloxyflavanone-N-phenyl
hydrazone (N101-43), induces apoptosis through up-regulation of
Fas/ FasL expression and inhibition of PI3K/Akt signaling pathways
in non-small-cell lung cancer cells. J Agric Food Chem.
59:10286–10297. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Xia Z, Dickens M, Raingeaud J, Davis RJ
and Greenberg ME: Opposing effects of ERK and JNK-p38 MAP kinases
on apoptosis. Science. 270:1326–1331. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Sebolt-Leopold JS: Development of
anticancer drugs targeting the MAP kinase pathway. Oncogene.
19:6594–6599. 2000. View Article : Google Scholar
|
|
20
|
Hilger RA, Scheulen ME and Strumberg D:
The Ras-Raf-MEK-ERK pathway in the treatment of cancer. Onkologie.
25:511–518. 2002. View Article : Google Scholar
|
|
21
|
Yager JD and Davidson NE: Estrogen
carcinogenesis in breast cancer. N Engl J Med. 354:270–282. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
de Leeuw R, Neefjes J and Michalides R: A
role for estrogen receptor phosphorylation in the resistance to
tamoxifen. Int J Breast Cancer. 2011:2324352011. View Article : Google Scholar
|
|
23
|
Thomas RS, Sarwar N, Phoenix F, Coombes RC
and Ali S: Phosphorylation at serines 104 and 106 by Erk1/2 MAPK is
important for estrogen receptor-alpha activity. J Mol Endocrinol.
40:173–184. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Weitsman GE, Li L, Skliris GP, Davie JR,
Ung K, Niu Y, Curtis-Snell L, Tomes L, Watson PH and Murphy LC:
Estrogen receptor-alpha phosphorylated at Ser118 is present at the
promoters of estrogen-regulated genes and is not altered due to
HER-2 overexpression. Cancer Res. 66:10162–10170. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Bhatt S, Xiao Z, Meng Z and
Katzenellenbogen BS: Phosphorylation by p38 mitogen-activated
protein kinase promotes estrogen receptor α turnover and functional
activity via the SCF(Skp2) proteasomal complex. Mol Cell Biol.
32:1928–1943. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Marino M, Acconcia F, Bresciani F, Weisz A
and Trentalance A and Trentalance A: Distinct nongenomic signal
transduction pathways controlled by 17beta-estradiol regulate DNA
synthesis and cyclin D(1) gene transcription in HepG2 cells. Mol
Biol Cell. 13:3720–3729. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Marino M, Acconcia F and Trentalance A:
Biphasic estradiol-induced AKT phosphorylation is modulated by PTEN
via MAP kinase in HepG2 cells. Mol Biol Cell. 14:2583–2591. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Gutierrez-Orozco F and Failla ML:
Biological activities and bioavailability of mangosteen xanthones:
A critical review of the current evidence. Nutrients. 5:3163–3183.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Sakagami Y, Iinuma M, Piyasena KG and
Dharmaratne HR: Antibacterial activity of alpha-mangostin against
vancomycin resistant Enterococci (VRE) and synergism with
antibiotics. Phytomedicine. 12:203–208. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Gopalakrishnan G, Banumathi B and Suresh
G: Evaluation of the antifungal activity of natural xanthones from
Garcinia mangostana and their synthetic derivatives. J Nat Prod.
60:519–524. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Sidahmed HM, Abdelwahab SI, Mohan S,
Abdulla MA, Taha MME, Hashim NM, Hadi AHA, Vadivelu J, Fai ML,
Rahmani M, et al: alpha-Mangostin from Cratoxylum arborescens
(Vahl) blume demonstrates anti-ulcerogenic property: A mechanistic
study. Evid Based Complement Alternat Med. 2013:4508402013.
View Article : Google Scholar
|
|
32
|
Huang HJ, Chen WL, Hsieh RH and Hsieh-Li
HM: Multifunctional effects of mangosteen pericarp on cognition in
C57BL/6J and triple transgenic Alzheimer's mice. Evid Based
Complement Alternat Med. 2014:8136722014. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Pedraza-Chaverrí J, Reyes-Fermín LM,
Nolasco-Amaya EG, Orozco-Ibarra M, Medina-Campos ON,
González-Cuahutencos O, Rivero-Cruz I and Mata R: ROS scavenging
capacity and neuroprotective effect of alpha-mangostin against
3-nitropropionic acid in cerebellar granule neurons. Exp Toxicol
Pathol. 61:491–501. 2009. View Article : Google Scholar
|
|
34
|
Pedraza-Chaverri J, Cárdenas-Rodríguez N,
Orozco-Ibarra M and Pérez-Rojas JM: Medicinal properties of
mangosteen (Garcinia mangostana). Food Chem Toxicol. 46:3227–3239.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Devi Sampath P and Vijayaraghavan K:
Cardioprotective effect of alpha-mangostin, a xanthone derivative
from mangosteen on tissue defense system against
isoproterenol-induced myocardial infarction in rats. J Biochem Mol
Toxicol. 21:336–339. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Suksamrarn S, Suwannapoch N, Phakhodee W,
Thanuhiranlert J, Ratananukul P, Chimnoi N and Suksamrarn A:
Antimycobacterial activity of prenylated xanthones from the fruits
of Garcinia mangostana. Chem Pharm Bull (Tokyo). 51:857–859. 2003.
View Article : Google Scholar
|
|
37
|
Nguyen PT and Marquis RE: Antimicrobial
actions of α-mangostin against oral streptococci. Can J Microbiol.
57:217–225. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Kaomongkolgit R, Jamdee K and Chaisomboon
N: Antifungal activity of alpha-mangostin against Candida albicans.
J Oral Sci. 51:401–406. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Chen LG, Yang LL and Wang CC:
Anti-inflammatory activity of mangostins from Garcinia mangostana.
Food Chem Toxicol. 46:688–693. 2008. View Article : Google Scholar
|
|
40
|
Chairungsrilerd N, Furukawa K, Ohta T,
Nozoe S and Ohizumi Y: Pharmacological properties of
alpha-mangostin, a novel histamine H1 receptor antagonist. Eur J
Pharmacol. 314:351–356. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Sánchez-Pérez Y, Morales-Bárcenas R,
García-Cuellar CM, López-Marure R, Calderon-Oliver M,
Pedraza-Chaverri J and Chirino YI: The alpha-mangostin prevention
on cisplatin-induced apoptotic death in LLC-PK1 cells is associated
to an inhibition of ROS production and p53 induction. Chem Biol
Interact. 188:144–150. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Nabandith V, Suzui M, Morioka T, Kaneshiro
T, Kinjo T, Matsumoto K, Akao Y, Iinuma M and Yoshimi N: Inhibitory
effects of crude alpha-mangostin, a xanthone derivative, on two
different categories of colon preneoplastic lesions induced by 1,
2-dimethylhydrazine in the rat. Asian Pac J Cancer Prev. 5:433–438.
2004.PubMed/NCBI
|
|
43
|
Nakagawa Y, Iinuma M, Naoe T, Nozawa Y and
Akao Y: Characterized mechanism of alpha-mangostin-induced cell
death: Caspase-independent apoptosis with release of endonuclease-G
from mitochondria and increased miR-143 expression in human
colorectal cancer DLD-1 cells. Bioorg Med Chem. 15:5620–5628. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Matsumoto K, Akao Y, Yi H, Ohguchi K, Ito
T, Tanaka T, Kobayashi E, Iinuma M and Nozawa Y: Preferential
target is mitochondria in alpha-mangostin-induced apoptosis in
human leukemia HL60 cells. Bioorg Med Chem. 12:5799–5806. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Krajarng A, Nakamura Y, Suksamrarn S and
Watanapokasin R: α-Mangostin induces apoptosis in human
chondrosarcoma cells through downregulation of ERK/JNK and Akt
signaling pathway. J Agric Food Chem. 59:5746–5754. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Tewtrakul S, Wattanapiromsakul C and
Mahabusarakam W: Effects of compounds from Garcinia mangostana on
inflammatory mediators in RAW264.7 macrophage cells. J
Ethnopharmacol. 121:379–382. 2009. View Article : Google Scholar
|
|
47
|
Sebastian KS and Thampan RV: Differential
effects of soybean and fenugreek extracts on the growth of MCF-7
cells. Chem Biol Interact. 170:135–143. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Sethi G, Sung B and Aggarwal BB: Nuclear
factor-kappaB activation: From bench to bedside. Exp Biol Med
(Maywood). 233:21–31. 2008. View Article : Google Scholar
|
|
49
|
Tabas I and Ron D: Integrating the
mechanisms of apoptosis induced by endoplasmic reticulum stress.
Nat Cell Biol. 13:184–190. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Dhanasekaran DN and Reddy EP: JNK
signaling in apoptosis. Oncogene. 27:6245–6251. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Li J and Holbrook NJ: Elevated
gadd153/chop expression and enhanced c-Jun N-terminal protein
kinase activation sensitizes aged cells to ER stress. Exp Gerontol.
39:735–744. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Tiwary R, Yu W, Li J, Park SK, Sanders BG
and Kline K: Role of endoplasmic reticulum stress in alpha-TEA
mediated TRAIL/ DR5 death receptor dependent apoptosis. PLoS One.
5:e118652010. View Article : Google Scholar
|
|
53
|
Ménard S, Tagliabue E, Campiglio M and
Pupa SM: Role of HER2 gene overexpression in breast carcinoma. J
Cell Physiol. 182:150–162. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Clark AS, West K, Streicher S and Dennis
PA: Constitutive and inducible Akt activity promotes resistance to
chemotherapy, trastuzumab, or tamoxifen in breast cancer cells. Mol
Cancer Ther. 1:707–717. 2002.PubMed/NCBI
|
|
55
|
Brognard J, Clark AS, Ni Y and Dennis PA:
Akt/protein kinase B is constitutively active in non-small cell
lung cancer cells and promotes cellular survival and resistance to
chemotherapy and radiation. Cancer Res. 61:3986–3997.
2001.PubMed/NCBI
|