Introduction
Gastric carcinoma (GC) is one of the most common
malignant tumors. Currently, combined surgical operation and
chemotherapy play an important role in the comprehensive treatment
of GC (1,2). However, multidrug resistance (MDR)
and its adverse effectiveness on chemotherapy lead to a
considerable number of cases where the effects of chemotherapy are
not ideal (1,2). Therefore, studying the mechanism of
MDR in GC is important in order to improve the efficacy of
chemotherapy and prolong the survival time of patients.
It has been reported that there are several
mechanisms whereby GCs become resistant to chemotherapeutic agents.
One mechanism is that P-glycoprotein (P-gp, 170 kDa) and its coding
gene multidrug resistance protein 1 (MDR1) play a role in
drug-excreted pump function (3).
Many studies have shown that MDR1/P-gp is closely related to the
effectiveness of chemotherapy. For example, the survival rate of GC
patients is short, the remission rate is low and the recurrence
rate is high when MDR1 is positive (4). Cyclin-dependent kinase inhibitor 1B
(p27Kip1) is one of the anti-oncogenes that were first
discovered by Polyak et al (5) in a variety of cancer cell MDR.
p27Kip1 was significantly downregulated in paclitaxel
(TAX), cisplatin and carboplatin-induced drug-resistant SKOV3
ovarian cancer cells (6).
Overexpression of p27Kip1 caused cell cycle arrest at S
phase, and promoted the pro-apoptotic reaction of cisplatin in
human ovarian cancer SKOV3 cells (6).
A large number of studies have demonstrated that
p27Kip1 plays an important role in the modulation of MDR
through its degradation. In drug-resistant human myeloma cell
lines, it was found that facilitation of S-phase kinase-associated
protein 2 (Skp2) expression promoted cell cycle progression and
p27Kip1 degradation (7). Cell adhesion-mediated drug resistance
(CAM-DR) is a primary factor leading to relapse after chemotherapy
(8). It was demonstrated that
increasing the p27Kip1 level or disturbing
p27Kip1 phosphorylation at the Thr (187) site inhibited
CAM-DR in human myeloma cell lines (9). Skp2 is an F-box family member that is
the specific substrate recognition subunit of the skp1-cullin-F-box
(SCF) ubiquitin ligase complex. Skp2 also promotes the degradation
of p27Kip1 through the ubiquitin proteasome pathway
(10). The degradation of
p27Kip1 could also be increased by the Kip1
ubiquitination-promoting complex (KPC) (11). In a study of GC drug resistance,
stable downregulation of the transcription factor E2F1 increased
the sensitivity of cisplatin-resistant SGC7901/cisplatin cells, and
inhibited the expression of drug resistance associated proteins,
such as MDR1 and Skp2 (12). Other
studies have also shown that E3 ubiquitin ligase Skp2 (nuclear) and
KPC (cytoplasmic)-induced phosphorylation of p27Kip1 on
Thr (187) and Ser (10) sites are
prerequisites for p27Kip1 degradation (11,13).
GAS, an important gastrointestinal (GI) hormone,
stimulates parietal cells to secret gastric acid (HCl) and
participate in epithelial proliferation of the GI tract (14). There are five main types of GAS -
GAS-14, -17, -34, -52 and -71. GAS-17 (GAS) accounts for >90% of
total gastric secretion, deriving from gastroduodenal G-cells in
most mammals (15,16). The secretion of GAS is elicited by
GAS-releasing peptide (GRP), which regulates gastric acid secretion
and enteric motor function (17).
Importantly, it has been demonstrated that GAS plays a role in
promoting cancer cell proliferation and anti-apoptosis. It has been
suggested that GAS has the potential to modulate MDR in cancer
cells (18). In CCK2 receptor
(CCK2R) expressing human gastric adenocarcinoma cells, GAS reduced
the expression of S-phase kinase associated protein 2
(p27Kip1) through increasing the expression of miR-222
(19). Additionally, recent
research has shown that in GC cells, GAS can significantly decrease
the level of p27Kip1 and increase cyclin E protein
expression through inducing the nuclear translocation of CacyBP/SIP
(20).
Although GAS has been shown to regulate the growth
of GC cells, the latest studies of GAS in our laboratory have found
that GAS accelerated cell cycle progression and proliferation of GC
and is associated with promoting the migration and invasion through
the β-catenin/T-cell factor-4 (TCF-4) pathway (21). It is still necessary to investigate
whether GAS functionally regulates the MDR of GC cells. The purpose
of the present study was to evaluate the effects of GAS on MDR in
GC. Hence, the effects and mechanisms of GAS on drug resistance of
GC were investigated. The results revealed that the expression of
GAS in GC tissues of drug-resistant patients and GC cell lines was
significantly upregulated. Further studies indicated that
suppression of GAS inhibited MDR through the degradation of
p27Kip1 in multidrug-resistant variants of GC cell
lines.
Materials and methods
Tissues
Surgical specimens of cancer tissues were collected
from 17 patients with advanced GC. All patients (ten males and
seven females with an average age of 58.3 years, ranging from 48 to
71 years) received neoadjuvant chemotherapy based on docetaxel
(DTX) and cisplatin (CDDP) prior to surgical removal of the tumors
(Table I). The response rates to
chemotherapy for the patients were evaluated as follows: a complete
response was considered as a good response, a partial response was
considered as a moderate response, stable disease was considered as
a poor response, and progressive disease was considered as no
response. Twelve of the patients had no or poor response and the
other five had moderate response to chemotherapy. The study was
performed with the approval of the Ethics Committee of the Second
Affiliated Hospital of Xi'an Jiaotong University. Written informed
consent was obtained from each patient before initiation of the
study.
 | Table IClinicopathological variables and the
expression of GAS in GC patients. |
Table I
Clinicopathological variables and the
expression of GAS in GC patients.
| Clinicopathological
variable | Cases | GAS expression | t-value | P-value |
|---|
| Age (mean ± SD),
58.3±7.1 years (range) (48–71) | | | 0.828 | 0.421 |
| <59 | 9 | 1.488±0.495 | | |
| ≥59 | 8 | 1.281±0.536 | | |
| Sex | | | 1.050 | 0.310 |
| Female | 7 | 1.56±0.51 | | |
| Male | 10 | 1.69±0.26 | | |
| Chemotherapy
regimen | | | 1.428 | 0.174 |
|
5-Fluorouracil | 11 | 1.517±0.474 | | |
| Irinotecan | 6 | 1.159±0.531 | | |
| Docetaxel
(DTX) | 17 | | | |
| Cisplatin
(CDDP) | 17 | | | |
| Response to
chemotherapy | | | 3.933 | 0.001 |
| No or poor | 12 | 1.62±0.46 | | |
| Moderate | 5 | 0.68±0.72 | | |
| Metastasis | | | 0.475 | 0.642 |
| Yes | 14 | 1.58±0.23 | | |
| No | 3 | 1.20±0.70 | | |
Cell culture
The human gastric adenocarcinoma cell line SGC7901
(obtained from Xijing Hospital, the Fourth Military Medical
University, Xi'an, Shaanxi, China) and its multidrug-resistant
variants SGC7901/VCR and SGC7901/ADR (established and maintained in
our laboratory) were cultured in RPMI-1640 medium supplemented with
10% fetal calf serum (Gibco BRL, Grand Island, NY, USA) in a
humidified atmosphere containing 5% CO2 at 37°C. To
maintain the MDR phenotype, VCR (final concentration 1
µg/ml) and Adriamycin (ADR, final concentration 0.4
µg/ml) were added to the culture media of SGC7901/VCR and
SGC7901/ADR cells, respectively.
Quantitative real-time polymerase chain
reaction (qRT-PCR)
Total RNA from GC tissues or cells was extracted
using TRIzol (Invitrogen), and cDNA was synthesized using an M-MLV
Reverse Transcriptase kit (Invitrogen). Then, the detection of
specific products was performed using SYBR green (Applied
Biosystems, Foster City, CA, USA) technology. Amplification
involved a denaturation step (95°C for 5 min, 1 cycle), and
amplification and quantification were repeated for 40 cycles (95°C
for 5 sec and 60°C for 1 min, respectively). The data of the
relative mRNA expression levels were calculated using the
2−∆∆Ct method and are presented as the fold change of
transcripts for genes. The sample mean value was calculated and
expressed as the cycle threshold (Ct). mRNA expression was
calculated as the difference (∆Ct) between the Ct value of the
target gene and the Ct value of the inner control.
2−∆∆Ct means the fold change in the target mRNA
expression. The mRNA expression expressed as the mean ± SD. Primer
sequences were as follows: GAS, forward,
5′-GAGCTACCCTGGCTGGAGCAGCAG-3′; reverse,
5′-CTCATCCTCAGCACTGCGGCGGCC-3′. Pgp, forward,
5′-GCAAGAGGAGCAGCTTAT-GAAG-3′; reverse,
5′-ACTCCCTACCTTCAAGTTGAGG-3′. β-actin, forward,
5′-AGGTCATCACCATTGGCAAT-3′; reverse,
5′-ACTCGTCATACTCCTGCTTG-3′.
Protein extraction
Total proteins from tissue and cell samples were
extracted using RIPA buffer (50 mM Tris-HCl, 150 mM NaCl, 2 mM
EDTA, 1% NP-40 and 0.1% SDS). For nuclear protein and cytoplasmic
protein extraction, protein was isolated using a Nuclear and
Cytoplasmic Protein Extraction kit (Beyotime, Haimen, China)
according to the manufacturer's instructions. Briefly, cells were
harvested and washed with PBS, and then centrifuged at 8,000 g for
10 min. The cell sediments were collected and resuspended in 20 ml
of PBS and mixed with 200 ml of buffer A containing 1 mM
phenylmethylsulfonyl fluoride (PMSF). After vortexing, cells were
placed in an ice bath for 10 min. Thereafter, 10 ml of buffer B was
added and the cells were again vortexed and placed in an ice bath
for 1 min followed by centrifugation at 12,000 g for 5 min at 4°C.
The supernatants containing cytoplasmic protein were collected and
used for further study. The remaining sediments were collected and
resuspended in 50 ml of a nuclear protein extraction agent and
subjected to an ice bath for 30 min with vortexing at intervals of
2 min. After centrifugation (12,000 g for 10 min at 4°C), the
supernatants containing nuclear protein were collected and stored
at −70°C for further analysis.
Western blotting (WB)
The protein quantification was measured using a BCA
protein assay kit (Beyotime Institute of Biotechnology) according
to the manufacturer's instructions. Proteins (20 µg) were
separated in a 10% SDS-PAGE gel and then electroblotted onto a
nitrocellulose membrane (Bio-Rad). The membrane was blocked with 5%
non-fat milk and then incubated with rabbit anti-GAS (1:1,000,
ab14182, Abcam, Cambridge, UK), Bcl-2 (1:700, ab59348, Abcam), MRP1
(1:600, ab32574, Abcam), Pgp (1:800, ab170904, Abcam), Skp2 (1:700,
ab19877, Abcam), KPC (1:900, ab151317, Abcam), p27Kip1
(1:600, Ab-10, Abcam), Thr (187) (1:600, sc-16324, Santa Cruz
Biotechnology, Santa Cruz, CA, USA), Ser (10) (1:400, ab36727, Abcam) and β-actin
(1:1,000, ab8227, Abcam) at 4°C overnight. Then, the membrane was
incubated with HRP conjugated anti-rabbit IgG (1:10,000, ab97064,
Abcam) for 1 h at room temperature. The signals were visualized
using ECL reagents (Beyotime). The protein bands were visualized
using an enhanced chemiluminescence detection system. Densitometry
values were analyzed using ImageJ 1.43 software (National
Institutes of Health, NIH, MD, USA), which were then normalized to
β-actin.
Immunofluorescence (IF)
We transfected SGC7901 cells with pCDNA3.1(+)/GAS or
pCDNA3.1(+)/GAS -NC. At 48 h after the transfection, the cells were
fixed in 3% paraformaldehyde in PBS at room temperature for 8 min,
then permeabilized with 0.2% Triton X-100 for 15 min at room
temperature. After washing in PBS, the cells were incubated with
Skp2 (1:400, Abcam) or KPC (1:600, Abcam) primary antibody at 4°C
overnight. After washing, the cells were incubated with
FITC-labeled secondary antibody (Pierce, Rockford, IL, USA) at room
temperature for 2 h avoiding light. The nuclear was stained with
DAPI for 5 min and washed with PBS at three times. Adding
anti-quenching reagent, and mounting was performed avoiding light.
The cells were examined under a Nikon fluorescence microscope
(Image Systems, Columbia, MD, USA).
Immunohistochemistry (IHC)
GC tissues were fixed in neutral buffered formalin
for 2 h at room temperature and for 18 h at 4°C and then tissues
were embedded in paraffin and sliced. The thickness of the tissue
serial sections was 3.5 µm. The serial sections mounted on
Superfrost Plus glass slides, and subjected to deparaffinization
and rehydration. The endogenous peroxidase activity of was blocked
by incubation in 0.3% H2O2 in methanol for 30
min. The sections were incubated overnight at 4°C with rabbit
anti-GAS antibodies (1:500, Abcam). The sections were exposed to
biotin-labeled secondary antibodies for 1 h and then developed with
DAB-H2O2. Immunohistochemical staining was
performed as described previously (22,23).
Cell transfection assay
The construction of the GAS over-expression vector
[pCDNA3.1(+)/GAS], and recombinant lentiviral vectors for siRNA
specific for GAS (siGAS) and Skp2 (siSkp2) was performed by Sangon
Biotechnology Co. Ltd. (Shanghai, China). All vectors were
transfected into cells according to the manufacturer's instructions
and following previous studies (24–29).
Briefly, for the functional study of GAS, SGC7901/VCR and
SGC7901/ADR cells were transfected with pCDNA3.1(+)/GAS or siGAS
for 48 h; for mechanistic experiments, SGC7901/VCR and SGC7901/ADR
or paclitaxel (TAX) and vincristine (VCR) combination (TAX-VCR)
treated SGC7901 which were pre-transfected with pCDNA3.1(+)/GAS
were further transfected with the siRNA of Skp2 (siSkp2) for 24 h.
Cells transfected with null vectors served as negative controls
(-NC). Following transfection, the medium was replaced with
complete RPMI-1640 medium supplemented with serum and
antibiotics.
MTT assay
To determine the drug sensitivity of the cells,
SGC7901/VCR, SGC7901/ADR and SGC7901 cells were plated in a 96-well
plate at a density of 5×103 cells/well in RPMI-1640
containing 10% FBS and were transfected respectively with the
above-mentioned vectors for 48 h. For the SGC7901/VCR and
SGC7901/ADR groups, 20 µl of 5 mg/ml MTT was then added per
well, and the culture was incubated for an additional 4 h. The
supernatant was then discarded and 150 µl/well of dimethyl
sulfoxide (DMSO) was added. The absorbance was measured at a
wavelength of 490 nm (A490). For the SGC7901 group, TAX (1
µM, Sigma, St. Louis, MO, USA) (30) and VCR (10 µM, Sigma)
(31) were added together in
SGC7901 cells, but not SGC7901/VCR or SGC7901/ADR. The culture
medium and the drug were replaced at 24-h intervals to maintain the
drug concentration. The following MTT assay steps are consistent
with the former. Each assay was performed in triplicate.
Apoptosis assay
Each group of cells was collected and detected using
an Annexin V Fluorescein Isothiocyanate kit (AV-FITC, BD
Pharmingen, San Diego, CA, USA) on a BD FACSCalibur™ system (BD,
Franklin Lakes, NJ, USA) according to the manufacturer's
instructions. In brief, the groups of cells were consistent with
the MTT assays and cells from the different conditions were
suspended in 100 ml of binding buffer, at a density of
1×106 cells per ml. Then, cells were incubated with
AV-FITC and propidium iodide (PI) for 15 min, and analyzed using
Beckman CXP software (Beckman Coulter, Brea, CA, USA) on a FC-500
flow cytometer (Beckman Coulter).
Statistical analysis
Statistical analysis was performed using GraphPad
Prism 5 software (GraphPad Software, Inc., La Jolla, CA, USA). Data
were analyzed using Student's t-test between two groups, and
categorical data were examined using the χ2 test. Data
are presented as the mean ± SD. P<0.05 was considered
statistically significant.
Results
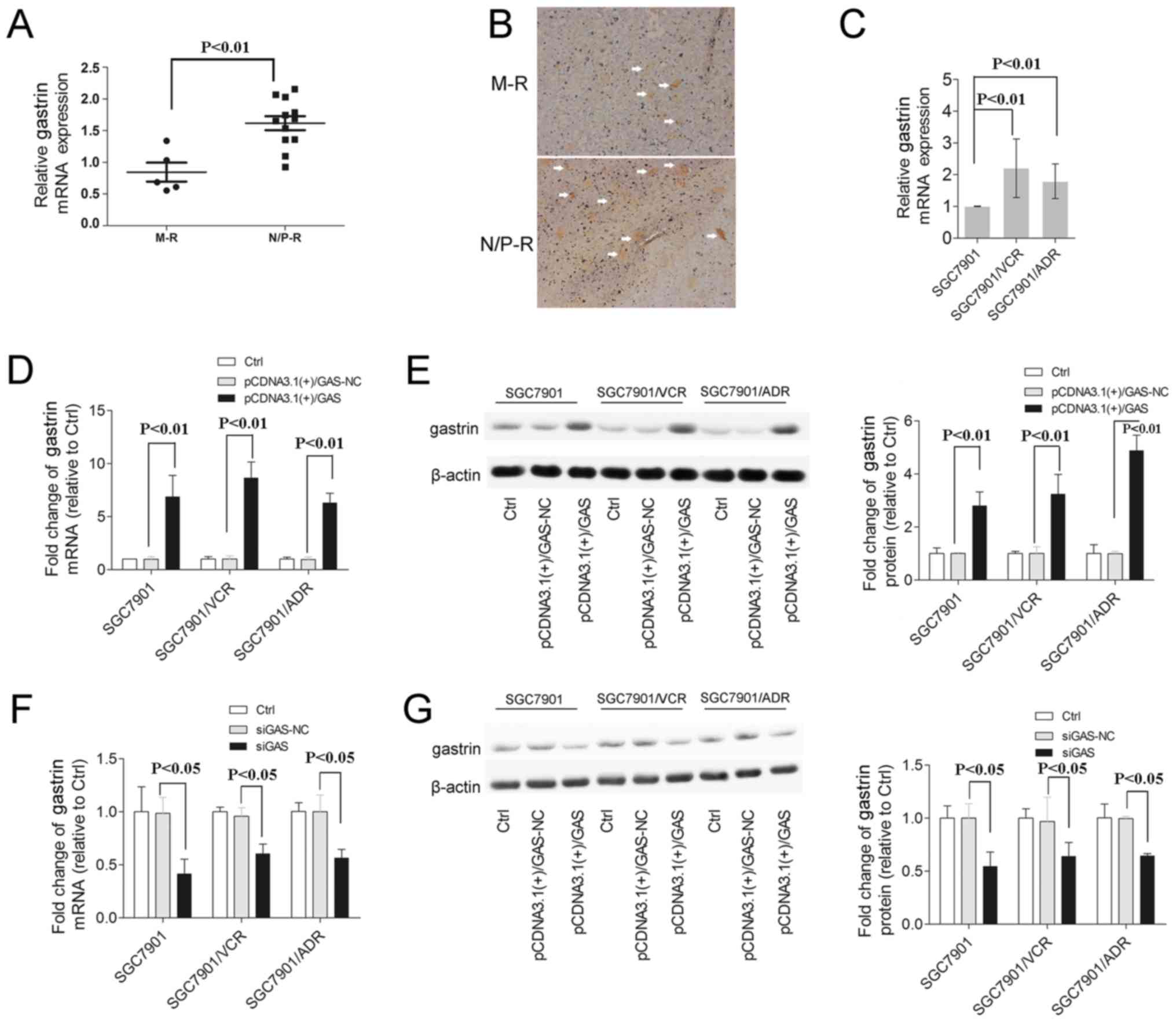
GAS is overexpressed in drug-resistant GC
cells
Recently, GAS was demonstrated to participate in the
growth and metastasis of GC cells (21,32).
Firstly, in order to determine the clinical relevance of GAS
expression in chemoresistance of GC patients, we used qRT-PCR to
evaluate the expression of GAS in human GC specimens.
Interestingly, GAS was highly expressed in non-responders or poor
responders (N/P-R). As shown in Fig.
1A, the mRNA levels of GAS was ~2.0-fold higher in N/P-R than
in moderate responders (M-R) (Fig.
1A, P<0.01).
Additionally, GAS-positive cells were stained brown.
Obviously, GAS-positive cells in the N/P-R group are much more
numerous than in the M-R group (Fig.
1B). Secondly, the mRNA level of GAS in drug-resistant cell
lines was evaluated. Our results revealed a high expression of GAS
in SGC7901/VCR and SGC7901/ADR cells compared with the SGC7901
group (Fig. 1C, P<0.01).
GAS inhibits drug sensitivity and
increases viability of human gastric cancer cells
We constructed the GAS overexpression vector
pCDNA3.1(+)/GAS and its silencing vector siGAS, and the effects of
transfection were tested. After 48 h of transfection, we found that
GAS was significantly upregulated in pCDNA3.1(+)/GAS
vector-transfected SGC7901/VCR, SGC7901/ADR and SGC7901 cells
(Fig. 1D and E) (P<0.01) and
downregulated in siGAS transfected SGC7901/VCR, SGC7901/ADR and
SGC7901 cells (P<0.05) compared with the NC groups (Fig. 1F and G).
To further investigate whether GAS contributes to
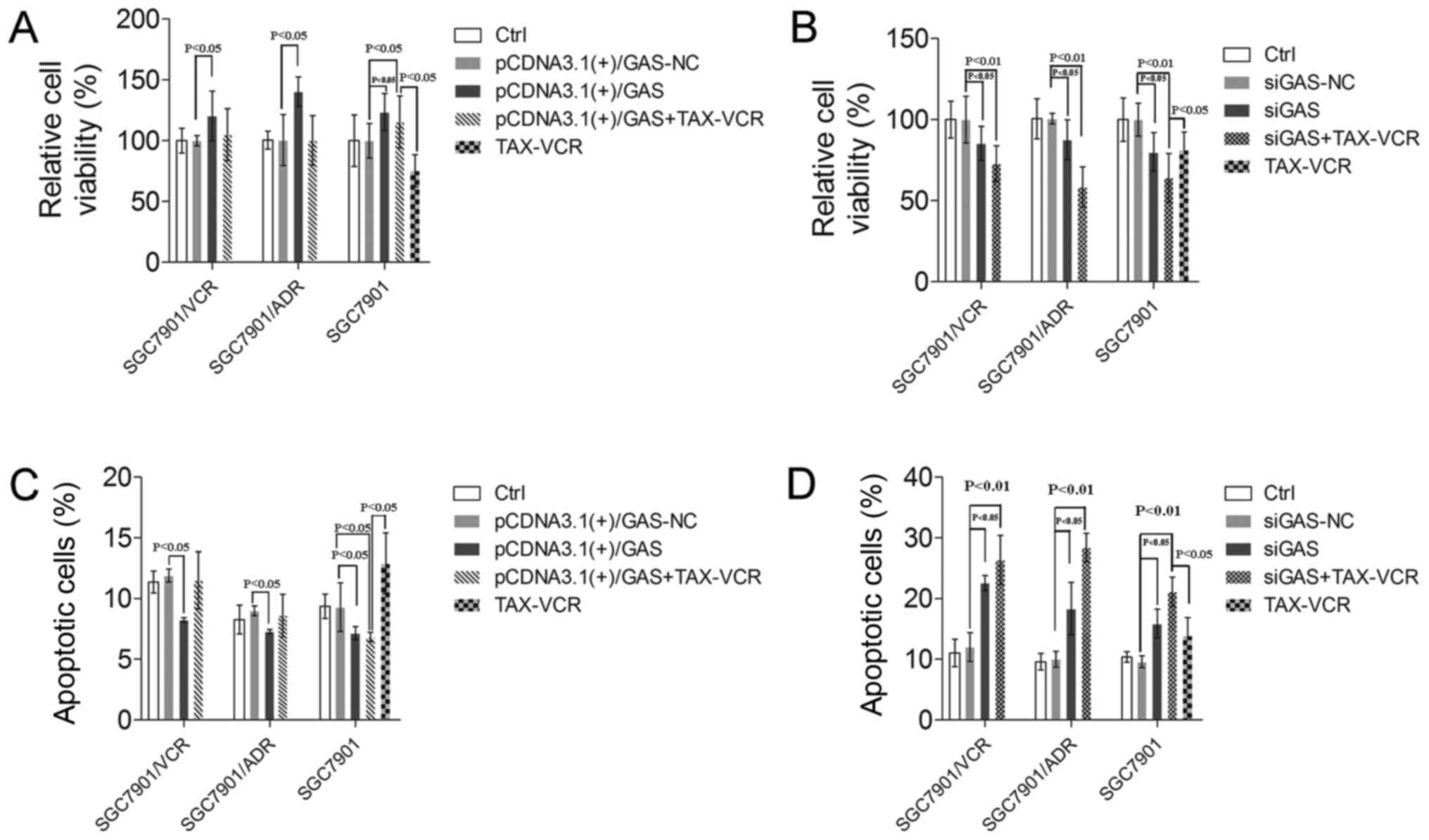
MDR in GC, it was force expressed or knocked down in GC cell lines.
MTT results showed that the transfection of pCDNA3.1(+)/GAS
markedly increased the cell viability of the drug-resistant GC cell
lines including SGC7901/VCR and SGC7901/ADR compared with
pCDNA3.1(+)/GAS-NC-transfected cells (P<0.05) (Fig. 2A). The viability of
pCDNA3.1(+)/GAS-transfected SGC7901 cell lines was still
upregulated on treatment with a TAX-VCR combination compared with
pCDNA3.1(+)/GAS-NC (Fig. 2A).
Additionally, SGC7901/VCR and SGC7901/ADR cells were transfected
with siGAS, revealing the opposite results. siGAS significantly
decreased the viability of SGC7901/VCR and SGC7901/ADR cells
(P<0.05) (Fig. 2B). We also
found that the viability of siGAS-transfected SGC7901 cell lines
was downregulated on treatment with the TAX-VCR combination
compared with siGAS-NC (Fig.
2B).
siGAS increases the drug sensitivity of
human gastric cancer cells through testing apoptosis
Previous studies have revealed that GAS play the
role of anti-apoptosis in a variety of cancer cell types (18). Therefore, we tested the effects of
GAS on the apoptosis of drug-resistant GC cell lines. The results
showed that transfection of the pCDNA3.1(+)/GAS vector decreased
the apoptosis rates in SGC7901/VCR, SGC7901/ADR and TAX-VCR
co-treated SGC7901 cells compared with the pCDNA3.1(+)/GAS-NC group
(Fig. 2C). However, siGAS groups
revealed opposite results and siGAS promoted the sensitivity of
SGC7901 to TAX-VCR (Fig. 2D).
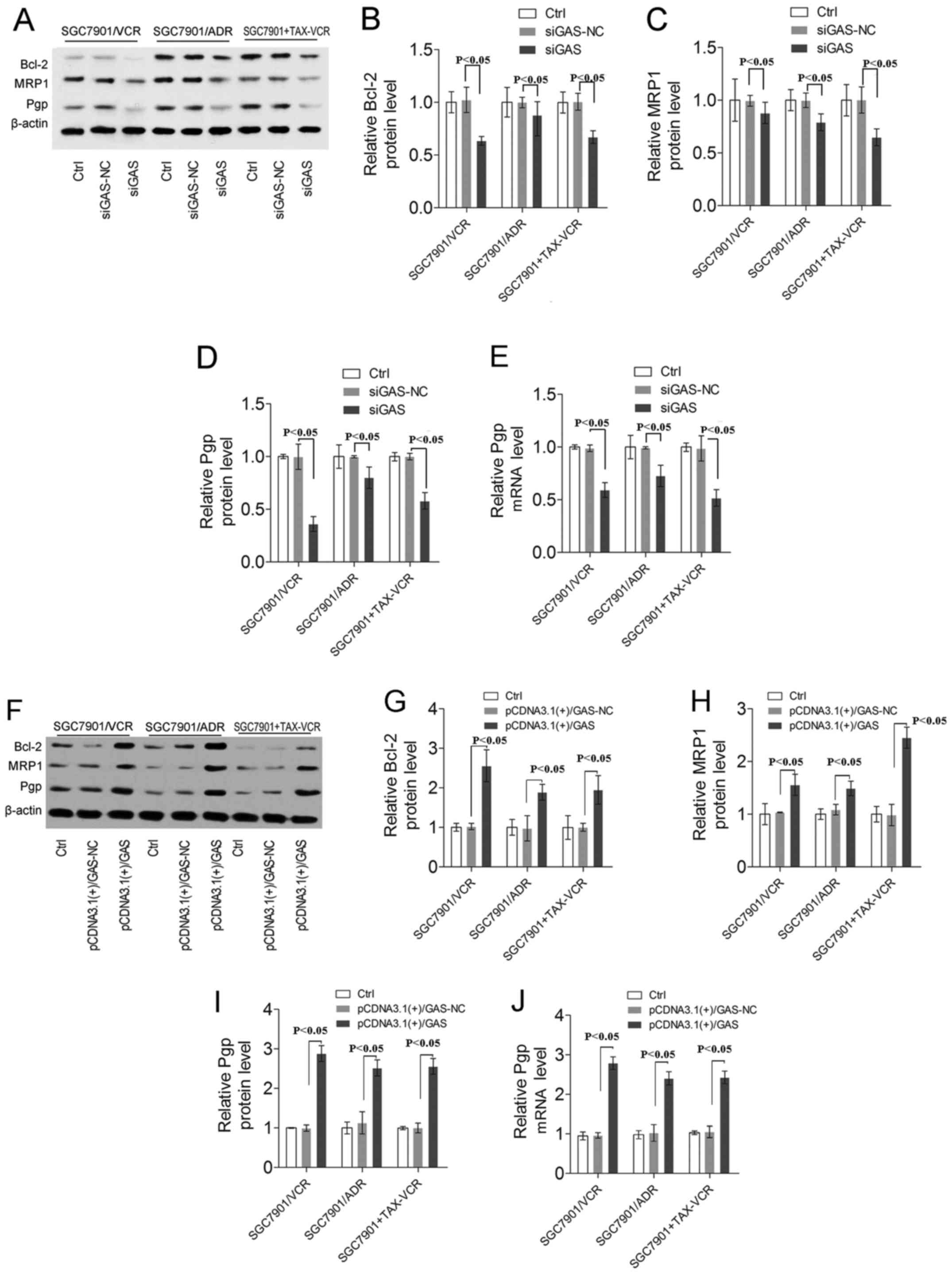
Further studies indicated that transfection of siGAS significantly
down-regulated Bcl-2 MRP1 and Pgp expression in SGC7901/VCR,
SGC7901/ADR and TAX-VCR co-treated SGC7901 cells (Fig. 3A–E, P<0.05). However, the
opposite results were found in the GAS overexpression group
(Fig. 3F–J, P<0.05). These
results indicate that GAS confers decreased sensitivity to TAX or
VCR and might be a key promoter of MDR in human GC cells.
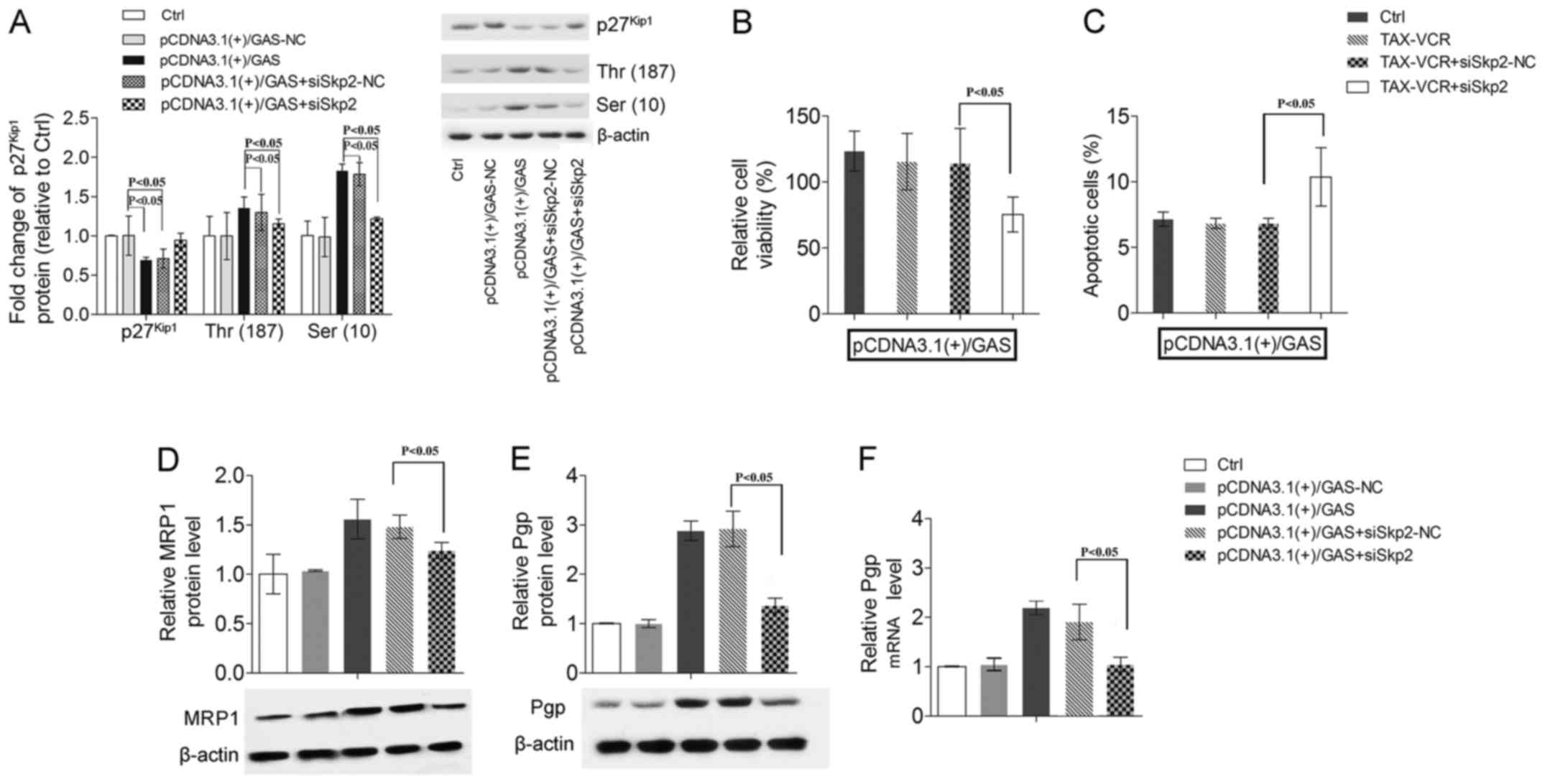
GAS inhibits the expression of
p27Kip1 in GC cell lines
Previous studies have shown that GAS can inhibit the
expression of p27Kip1 and p27Kip1 can
participate in drug resistance in a variety of cancer cells
(6,33). Overexpression of p27Kip1
could lead to cell cycle arrest in S phase, and promote the
pro-apoptosis response of cisplatin (6). In our study, we found that the
phosphorylation levels of p27Kip1 on Thr (187) and Ser
(10) sites were higher in
SGC7901/VCR and SGC7901/ADR cells than in normal SGC7901 cells, but
the levels of p27Kip1 were lower (Fig. 4A–C, P<0.05). In pCDNA3.1(+)/GAS
transfected SGC7901 cells, we found similar results:
pCDNA3.1(+)/GAS significantly inhibited p27Kip1
expression, but increased the phosphorylation levels of
p27Kip1 on Thr (187) and Ser (10) sites (Fig. 4D, P<0.05). Additionally, using
immunofluorescence localization methods and WB, we found that the
nuclear accumulation of Skp2 and the cytoplasmic accumulation of
KPC were increased by GAS overexpression (Fig. 4E–G, P<0.05). These results
indicate that GAS may regulate and induce the degradation of
p27Kip1 during the emergence of drug resistance in human
gastric cancer cells.
p27Kip1 induces drug
sensitivity of pCDNA3.1(+)/GAS-transfected SGC7901
Studies have shown that depletion of Skp2 could
inhibit the degradation of p27Kip1 (34). Therefore we constructed recombinant
lentiviral siRNA vectors specific for Skp2 (siSkp2). SGC7901 cells
that were transfected with pCDNA3.1(+)/GAS were further transfected
with siSkp2. We tested the phosphorylation levels of
p27Kip1 at Thr (187) and Ser (10) sites, and found that they were all
downregulated and p27Kip1 levels were upregulated
compared with the siSkp2-NC group (Fig. 5A, P<0.05). SGC7901 cells
transfected with pCDNA3.1(+)/GAS and treated with TAX-VCR were
further transfected with siSkp2, and we found the cell viability
was significantly inhibited and the cell apoptosis was markedly
promoted compared with the siSkp2-NC group (Fig. 5B and C, P<0.05). Additionally,
siSkp2 attenuated the high level of MRP1 and Pgp in SGC7901 cells
induced by GAS-overexpression (Fig.
5D–F, P<0.05).
These results indicated that inhibition of
p27Kip1 degradation could attenuate the effects of GAS
overexpression. All of these results suggested that GAS induces the
MDR of SGC7901 via the degradation of p27Kip1.
Discussion
In this study, we demonstrated a stimulatory role of
GAS in MDR in human GC cells. Firstly, we provided evidence that
GAS is upregulated in the SGC7901/VCR and SGC7901/ADR cell lines
and in non- or poorly-responding gastric tumor patients. Secondly,
further analysis demonstrated that the overexpression of GAS partly
abolished the drug sensitivity of SGC7901 cells while the silencing
of GAS increased the drug sensitivity of SGC7901, SGC7901/VCR and
SGC7901/ADR cells. A mechanistic assay confirmed that the
degradation of p27Kip1 was the downstream mechanism of
GAS in regulating MDR in GC cells. These findings suggest a
potentially important role for GAS in regulating MDR.
MDR is a great impediment to the success of
chemotherapy for GC. It is believed that GAS plays an important
role in the process of cell migration and might be involved in the
regulation of cancer development and drug resistance in digestive
tract and other cancers (18).
Firstly, it was reported that GAS mediates proliferative responses
in a variety of cancer cell model systems (21) and in particular has been implicated
in accelerating the development of gastrointestinal cancers
(35). Secondly, Bcl-2 is known as
a key regulator of the progression of apoptosis and MDR in multiple
types of cancer (36). In a study
of large intestine carcinoma, high expression of GAS in large
intestine cancer tissue samples was accompanied by high expression
of Bcl-2, which indicates that the regulation and control of GAS in
cell apoptosis of large intestine carcinoma may be directly related
to the abnormal expression of bcl-2 (37) Furthermore, in a clinical study of
resistance to chemotherapy in small cell lung cancer (SCLC),
pro-GAS releasing peptide (pro-GRP) was identified as a surrogate
marker of Bcl-2 amplification and changes correlated with changes
in tumor volume (38). Thirdly,
the secretion of GAS was controlled by proton pump inhibitors
(PPI), which could protect colorectal cancer (CRC) cells from
developing chemo- or radiotherapeutics resistance (39). Rintoul and Sethi (40) suggested that SCLC cells produce a
variety of growth factors, cytokines and inflammatory mediators,
including insulin-like growth factor-1, GRP and interleukin-8 via
autocrine and paracrine effects, and these proteins are able to
protect SCLC cells from chemotherapy-induced apoptosis. In the
present study, we found direct evidence of GAS downregulating the
drug sensitivity of GC cells. The MDR of SGC7901 was upregulated by
GAS overexpression and the MDR of SGC7901/VCR and SGC7901/VCR was
downregulated by GAS silencing. These results demonstrated that GAS
could play a positive role in the formation and development of
human GC.
SGC7901/VCR and SGC7901/VCR are the MDR variants of
SGC7901, and we found remarkable upregulation of GAS along with
MRP1, Pgp and Bcl-2 overexpression, while the downregulation of
p27Kip1 was also demonstrated in these two cell lines
compared with SGC7901. Interestingly, we overexpressed GAS using
pCDNA3.1(+)/GAS in SGC7901, and found the overexpression of GAS
upregulated MRP1, Pgp and Bcl-2, but accelerated the degradation of
p27Kip1 with a coincident upregulation of the
phosphorylation levels of p27Kip1 on Thr (187) and Ser
(10) sites. Recently,
p27Kip1 has been shown to be involved in MDR in a
variety of cancer cell lines. It was reported that
p27Kip1 was significantly downregulated in TAX,
cisplatin and carboplatin-induced drug resistance in ovarian cancer
cells, and when the expression of p27Kip1 was restored,
the sensitivity of cisplatin cells was increased (6). Overexpression of p27Kip1
causes cell cycle arrest at S phase, and promotes the pro-apoptotic
reaction of cisplatin (6). More
importantly, GAS could inhibit the expression of p27Kip1
by increasing the expression of miR-222 (19). In GC cells, GAS induced the nuclear
translocation of CacyBP/SIP. CacyBP-SIP nuclear translocation
significantly decreased the level of the cell cycle inhibitor
p27Kip1, increasing cyclin E protein expression
(20), so CacyBP/SIP nuclear
translocation might be one of the potential mechanisms of the loss
of p27Kip1. More importantly, we found that nuclear
accumulation of Skp2 was also upregulated by gastrin. However, in
the study of Niu et al, CacyBP/SIP nuclear translocation did
not affect the level of Skp2 (20). Thus we believe that there is an
intimate connection between p27Kip1 and GAS in
regulating drug resistance of GC, and nuclear accumulation of Skp2
might be the key mechanism that can correlate between increased
gastrin and degradation of p27Kip1.
A large number of studies have demonstrated that the
degradation of p27Kip1 plays an important role in the
modulation of MDR. First, in drug-resistant human myeloma cell
lines, it was found that facilitation of Skp2 expression promoted
cell cycle progression and suppressed p27Kip1 expression
(7). Second, cell
adhesion-mediated drug resistance (CAM-DR) in many types of cancer
is a primary factor leading to relapse after chemotherapy (8). It was demonstrated that increasing
p27Kip1 level or disturbing p27Kip1
phosphorylation at Thr (187) inhibited CAM-DR in human myeloma cell
lines (9) Third, Skp2 can promote
the degradation of p27Kip1 through the ubiquitin
proteasome pathway (10), while
the degradation of p27Kip1 can also be increased by KPC
(11). In a study of drug
resistance in GC, stable downregulation of the transcription factor
E2F1 increased the sensitivity of SGC7901/cisplatin (a
cisplatin-resistant GC cell line), and inhibited the expression of
drug resistance associated proteins such as MDR1 and Skp2 (12). Other studies also show that E3
ubiquitin ligase Skp2 (nuclear) and KPC (cytoplasmic)-induced
phosphorylation of p27Kip1 on Thr (187) and Ser
(10) sites is a prerequisite for
p27Kip1 degradation (11,13).
In the present study, we found a significant
accumulation of Skp2 in the nucleus and accumulation of KPC in the
cytoplasm of GC cells. Our results are consistent with previous
studies. It has been shown that the accumulation of
p27Kip1 was increased in Skp2 defect-mice. Skp2 can
interact with p27Kip1 ubiquitinating it (11,41–43).
E3 ubiquitin ligase Skp2 (in the nucleus) and KPC (in the
cytoplasm)-induced phosphorylation of p27Kip1 on Thr
(187) and Ser (10) sites is a
prerequisite for degradation (11,13).
In agreement with these studies, p27Kip1 restored the
MDR that was inhibited by silencing of GAS. The silencing of both
Skp2 and KPC showed similar results. All of these studies indicate
that the degradation of p27Kip1 is a key component of
the pathway downstream of GAS in modulating multidrug resistance of
GC cells.
In conclusion, our findings indicated that GAS
promotes the introduction of MDR in GC cells by p27Kip1
degradation through a mechanism promoting phosphorylation levels of
p27Kip1 on Thr (187) and Ser (10) sites and inducing the accumulation
of E3 ubiquitin ligase Skp2 (nuclear) and KPC (cytoplasmic). Given
its triggered effect on MDR, GAS holds promise to be a novel
therapeutic target for chemotherapy in GC patients.
Acknowledgments
The authors would like to thank the members of the
Second Affiliated Hospital of Xi'an Jiaotong University and Xi'an
Central Hospital for providing technical support and helpful
discussions concerning the present study.
References
|
1
|
Glavinas H, Krajcsi P, Cserepes J and
Sarkadi B: The role of ABC transporters in drug resistance,
metabolism and toxicity. Curr Drug Deliv. 1:27–42. 2004. View Article : Google Scholar
|
|
2
|
Varma MV, Ashokraj Y, Dey CS and
Panchagnula R: P-glycoprotein inhibitors and their screening: A
perspective from bioavailability enhancement. Pharmacol Res.
48:347–359. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Marie JP, Zittoun R and Sikic BI:
Multidrug resistance (mdr1) gene expression in adult acute
leukemias: Correlations with treatment outcome and in vitro drug
sensitivity. Blood. 78:586–592. 1991.PubMed/NCBI
|
|
4
|
Huang Z, Zhang S, Shen Y, Liu W, Long J
and Zhou S: Influence of MDR1 methylation on the curative effect of
interventional embolism chemotherapy for cervical cancer. Ther Clin
Risk Manag. 12:217–223. 2016.PubMed/NCBI
|
|
5
|
Polyak K, Lee MH, Erdjument-Bromage H,
Koff A, Roberts JM, Tempst P and Massagué J: Cloning of
p27Kip1, a cyclin-dependent kinase inhibitor and a
potential mediator of extracellular antimitogenic signals. Cell.
78:59–66. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Zhao Y, Li Q, Wu X and Chen P:
Upregulation of p27Kip1 by demethylation sensitizes
cisplatin-resistant human ovarian cancer SKOV3 cells. Mol Med Rep.
14:1659–1666. 2016.PubMed/NCBI
|
|
7
|
Yang Y, Chen Y, Saha MN, Chen J, Evans K,
Qiu L, Reece D, Chen GA and Chang H: Targeting phospho-MARCKS
overcomes drug-resistance and induces antitumor activity in
preclinical models of multiple myeloma. Leukemia. 29:715–726. 2015.
View Article : Google Scholar
|
|
8
|
Sun L, Liu L, Liu X, Wang Y, Li M, Yao L,
Yang J, Ji G, Guo C, Pan Y, et al: Gastric cancer cell adhesion to
laminin enhances acquired chemotherapeutic drug resistance mediated
by MGr1-Ag/37LRP. Oncol Rep. 32:105–114. 2014.PubMed/NCBI
|
|
9
|
Fei M, Hang Q, Hou S and Ruan C: Cell
adhesion to fibronectin down-regulates the expression of Spy1 and
contributes to drug resistance in multiple myeloma cells. Int J
Hematol. 98:446–455. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Carrano AC, Eytan E, Hershko A and Pagano
M: SKP2 is required for ubiquitin-mediated degradation of the CDK
inhibitor p27. Nat Cell Biol. 1:193–199. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Masumoto K and Kitagawa M: E3 ubiquitin
ligases as molecular targets in human oral cancers. Curr Cancer
Drug Targets. 16:130–135. 2016. View Article : Google Scholar
|
|
12
|
Yan LH, Wang XT, Yang J, Kong FB, Lian C,
Wei WY, Luo W, Xie YB and Xiao Q: Reversal of multidrug resistance
in gastric cancer cells by E2F-1 downregulation in vitro and in
vivo. J Cell Biochem. 115:34–41. 2014. View Article : Google Scholar
|
|
13
|
Hnit SS, Xie C, Yao M, Holst J, Bensoussan
A, De Souza P, Li Z and Dong Q: p27(Kip1) signaling:
Transcriptional and post-translational regulation. Int J Biochem
Cell Biol. 68:9–14. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Friis-Hansen L, Sundler F, Li Y, Gillespie
PJ, Saunders TL, Greenson JK, Owyang C, Rehfeld JF and Samuelson
LC: Impaired gastric acid secretion in gastrin-deficient mice. Am J
Physiol. 274:G561–G568. 1998.PubMed/NCBI
|
|
15
|
Rehfeld JF, Hansen CP and Johnsen AH:
Post-poly(Glu) cleavage and degradation modified by O-sulfated
tyrosine: A novel post-translational processing mechanism. EMBO J.
14:389–396. 1995.PubMed/NCBI
|
|
16
|
Goetze JP, Eiland S, Svendsen LB, Vainer
B, Hannibal J and Rehfeld JF: Characterization of gastrins and
their receptor in solid human gastric adenocarcinomas. Scand J
Gastroenterol. 48:688–695. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Merali Z, McIntosh J and Anisman H: Role
of bombesin-related peptides in the control of food intake.
Neuropeptides. 33:376–386. 1999. View Article : Google Scholar
|
|
18
|
Han YM, Park JM, Kangwan N, Jeong M, Lee
S, Cho JY, Ko WJ and Hahm KB: Role of proton pump inhibitors in
preventing hypergastrinemia-associated carcinogenesis and in
antagonizing the trophic effect of gastrin. J Physiol Pharmacol.
66:159–167. 2015.PubMed/NCBI
|
|
19
|
Lloyd KA, Moore AR, Parsons BN, O'Hara A,
Boyce M, Dockray GJ, Varro A and Pritchard DM: Gastrin-induced
miR-222 promotes gastric tumor development by suppressing p27kip1.
Oncotarget. 7:45462–45478. 2016.PubMed/NCBI
|
|
20
|
Niu YL, Li YJ, Wang JB, Lu YY, Liu ZX,
Feng SS, Hu JG and Zhai HH: CacyBP/SIP nuclear translocation
regulates p27Kip1 stability in gastric cancer cells. World J
Gastroenterol. 22:3992–4001. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Zhuang K, Yan Y, Zhang X, Zhang J, Zhang L
and Han K: Gastrin promotes the metastasis of gastric carcinoma
through the β-catenin/TCF-4 pathway. Oncol Rep. 36:1369–1376.
2016.PubMed/NCBI
|
|
22
|
Varro A, Kenny S, Hemers E, McCaig C,
Przemeck S, Wang TC, Bodger K and Pritchard DM: Increased gastric
expression of MMP-7 in hypergastrinemia and significance for
epithelial-mesenchymal signaling. Am J Physiol Gastrointest Liver
Physiol. 292:G1133–G1140. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Kumar JD, Steele I, Moore AR, Murugesan
SV, Rakonczay Z, Venglovecz V, Pritchard DM, Dimaline R, Tiszlavicz
L, Varro A, et al: Gastrin stimulates MMP-1 expression in gastric
epithelial cells: Putative role in gastric epithelial cell
migration. Am J Physiol Gastrointest Liver Physiol. 309:G78–G86.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Singh P, Owlia A, Varro A, Dai B,
Rajaraman S and Wood T: Gastrin gene expression is required for the
proliferation and tumorigenicity of human colon cancer cells.
Cancer Res. 56:4111–4115. 1996.PubMed/NCBI
|
|
25
|
Hayashi N, Kayo T, Sugano K and Takeuchi
T: Production of bioactive gastrin from the non-endocrine cell
lines CHO and COS-7. FEBS Lett. 337:27–32. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Supriatno HK, Harada K, Hoque MO, Bando T,
Yoshida H and Sato M: Overexpression of p27(Kip1) induces growth
arrest and apoptosis in an oral cancer cell line. Oral Oncol.
38:730–736. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Yin Y, Grabowska AM, Clarke PA, Whelband
E, Robinson K, Argent RH, Tobias A, Kumari R, Atherton JC and
Watson SA: Helicobacter pylori potentiates epithelial:mesenchymal
transition in gastric cancer: Links to soluble HB-EGF, gastrin and
matrix metalloproteinase-7. Gut. 59:1037–1045. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Sumimoto H, Yamagata S, Shimizu A, Miyoshi
H, Mizuguchi H, Hayakawa T, Miyagishi M, Taira K and Kawakami Y:
Gene therapy for human small-cell lung carcinoma by inactivation of
Skp-2 with virally mediated RNA interference. Gene Ther. 12:95–100.
2005. View Article : Google Scholar
|
|
29
|
Hristova NR, Tagscherer KE, Fassl A,
Kopitz J and Roth W: Notch1-dependent regulation of p27 determines
cell fate in colorectal cancer. Int J Oncol. 43:1967–1975.
2013.PubMed/NCBI
|
|
30
|
Del Vecchio CA, Feng Y, Sokol ES, Tillman
EJ, Sanduja S, Reinhardt F and Gupta PB: De-differentiation confers
multidrug resistance via noncanonical PERK-Nrf2 signaling. PLoS
Biol. 12:e10019452014. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Conway RM, Madigan MC, Billson FA and
Penfold PL: Vincristine- and cisplatin-induced apoptosis in human
retinoblastoma. Potentiation by sodium butyrate. Eur J Cancer.
34:1741–1748. 1998. View Article : Google Scholar
|
|
32
|
Gutkind JS: The pathways connecting G
protein-coupled receptors to the nucleus through divergent
mitogen-activated protein kinase cascades. J Biol Chem.
273:1839–1842. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Stendahl M, Nilsson S, Wigerup C, Jirström
K, Jönsson PE, Stål O and Landberg G: p27Kip1 is a
predictive factor for tamoxifen treatment response but not a
prognostic marker in premenopausal breast cancer patients. Int J
Cancer. 127:2851–2858. 2010. View Article : Google Scholar
|
|
34
|
Tsvetkov LM, Yeh KH, Lee SJ, Sun H and
Zhang H: p27(Kip1) ubiquitination and degradation is regulated by
the SCF(Skp2) complex through phosphorylated Thr187 in p27. Curr
Biol. 9:661–664. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Marshall KM, Laval M, Estacio O, Hudson
DF, Kalitsis P, Shulkes A, Baldwin GS and Patel O: Activation by
zinc of the human gastrin gene promoter in colon cancer cells in
vitro and in vivo. Metallomics. 7:1390–1398. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Reed JC, Miyashita T, Takayama S, Wang HG,
Sato T, Krajewski S, Aimé-Sempé C, Bodrug S, Kitada S and Hanada M:
BCL-2 family proteins: Regulators of cell death involved in the
pathogenesis of cancer and resistance to therapy. J Cell Biochem.
60:23–32. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Mao JD, Wu P, Xia XH, Hu JQ, Huang WB and
Xu GQ: Correlation between expression of gastrin, somatostatin and
cell apoptosis regulation gene bcl-2/bax in large intestine
carcinoma. World J Gastroenterol. 11:721–725. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Gandhi L, Camidge DR, Ribeiro de Oliveira
M, Bonomi P, Gandara D, Khaira D, Hann CL, McKeegan EM, Litvinovich
E, Hemken PM, et al: Phase I study of Navitoclax (ABT-263), a novel
Bcl-2 family inhibitor, in patients with small-cell lung cancer and
other solid tumors. J Clin Oncol. 29:909–916. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
De Milito A and Fais S: Proton pump
inhibitors may reduce tumour resistance. Expert Opin Pharmacother.
6:1049–1054. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Rintoul RC and Sethi T: Extracellular
matrix regulation of drug resistance in small-cell lung cancer.
Clin Sci (Lond). 102:417–424. 2002. View Article : Google Scholar
|
|
41
|
Hara T, Kamura T, Kotoshiba S, Takahashi
H, Fujiwara K, Onoyama I, Shirakawa M, Mizushima N and Nakayama KI:
Role of the UBL-UBA protein KPC2 in degradation of p27 at G1 phase
of the cell cycle. Mol Cell Biol. 25:9292–9303. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Nakayama K, Nagahama H, Minamishima YA,
Matsumoto M, Nakamichi I, Kitagawa K, Shirane M, Tsunematsu R,
Tsukiyama T, Ishida N, et al: Targeted disruption of Skp2 results
in accumulation of cyclin E and p27(Kip1), polyploidy and
centrosome overduplication. EMBO J. 19:2069–2081. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Nakayama K, Nagahama H, Minamishima YA,
Miyake S, Ishida N, Hatakeyama S, Kitagawa M, Iemura S, Natsume T
and Nakayama KI: Skp2-mediated degradation of p27 regulates
progression into mitosis. Dev Cell. 6:661–672. 2004. View Article : Google Scholar : PubMed/NCBI
|