Introduction
Oral squamous cell carcinoma (OSCC) represents
>95% of cancers of the oral cavity (1). In developing countries, OSCC is the
6th most common type of cancer. OSCC has an overall 5-year survival
rate of <50% (1), and this poor
prognosis is due mainly to a high likelihood of metastasis to lymph
nodes in the neck (2–5). Despite recent advances in treatment
modalities, such as surgery, radiotherapy and chemotherapy, the
survival rates for patients with OSCC have not markedly improved
(6). Thus, metastasis to the lymph
nodes in the neck is a major prognostic factor in OSCC.
Tumor invasion and metastasis require tumor cells to
resolve and move through the extracellular matrix (ECM). Matrix
metalloproteinases (MMPs) and tissue inhibitors of
metalloproteinases (TIMPs) are important for the degradation of the
ECM during these processes (7).
MMP-2 is of particular importance in tumor-cell invasion as it can
deconstruct collagen type IV, the major constituent of basement
membranes (8). The proteolytic
activation of MMP-2 is controlled by TIMP-2, which is necessary for
the activation of pro-MMP-2 (9,10).
TIMP-2 forms a complex with membrane type-I metalloproteinase 1
(MT1-MMP) on cell membranes, and the TIMP-2/MT1-MMP complex
activates pro-MMP-2. Thus, an imbalance in MMP-2 and TIMP-2 may
disrupt the balance of ECM turnover and can thus lead to the
uncontrolled degradation of the ECM, including basement membranes,
and is the most likely cause of tumor invasion and metastasis
(7). Thus, it may be possible to
control pro-MMP-2 activation, and therefore tumor invasion and
metastasis, by regulating TIMP-2.
MicroRNAs (miRNAs or miRs), which are short,
non-coding RNAs of 20–22 nucleotides in length, that regulate gene
expression at the post-transcriptional level by interacting with
the 3′-untranslated regions (3′-UTRs) of a target gene (11). miRNAs are involved in a wide range
of biological functions, and can function as oncogenes or tumor
suppressors, according to the functions of their target genes. To
date, several miRNAs have been implicated in the progression of
OSCC, including miR-26a/b, miR-23b/27b, miR-155, miR-221 and
miR-205 (12–16). Recent studies have shown that
miR-205-5p is downregulated in various cancer cells, including
breast cancer, prostate cancer and oral cancer cells (17–19).
In addition, the transient overexpression of miR-205-5p in cancer
cells has been reported to suppress tumor progression by inhibiting
tumor-associated genes and upregulating tumor suppressor genes.
However, the signaling pathway and anti-tumorigenic functions of
miR-205-5p in various cancer cells are unclear.
Therefore, the present study was conducted to
determine the function of miR-205-5p in OSCC and the molecular
mechanisms through which miR-205-5p suppresses OSCC tumor
progression.
Materials and methods
OSCC clinical specimens and cell
lines
OSCC cell lines (HSC3 and SAS) obtained from the
Japanese Collection of Research Bioresources (Osaka, Japan) were
cultured as previously described (14). We authenticated HSC3 and SAS
through short tandem repeat (STR) profiling (PowerPlex 16 STR
system) in 2017 and ensured that no culture contamination had
occurred. We obtained 71 primary OSCC tissue, and 28 metastatic
lymph node tissue samples from 71 previously untreated subjects
with OSCC who visited the University of Tsukuba Hospital (Tsukuba,
Japan) between February 2008 and November 2010, as well as tissues
samples from 6 subjects who did not have cancer. Primary and
metastatic specimens were collected at the time of biopsy and neck
dissection, respectively. The samples were prepared for
formalin-fixed paraffin-embedded (FFPE) histology using standard
procedures. OSCC was diagnosed and classified based on the
tumor-node metastasis (TNM) system of the Union for International
Cancer Control (UICC). All OSCC cases were diagnosed clinically and
confirmed histologically by pathologists. The clinical
characteristics of the patients with OSCC are shown in Table I.
 | Table IClinical characteristics of patients
with OSCC patients. |
Table I
Clinical characteristics of patients
with OSCC patients.
| Primary OSCC
| Metastatic OSCC
|
|---|
| Total | Lymph
node-negative
(n=41) | Lymph
node-positive
(n=30) | Metastatic lymph
node
(n=28) |
|---|
| Age | | | | |
| <60 | 18 | 7 | 11 | 9 |
| ≥60 | 53 | 34 | 19 | 19 |
| Sex | | | | |
| Male | 47 | 24 | 23 | 21 |
| Female | 24 | 17 | 7 | 7 |
| Primary lesion | | | | |
| Tongue | 33 | 20 | 13 | 12 |
| Gum | 23 | 15 | 8 | 7 |
| Buccal mucosa | 7 | 4 | 3 | 3 |
| Other | 8 | 2 | 6 | 6 |
| pTNM stage | | | | |
| I–II | 28 | 28 | 0 | 0 |
| III–IV | 43 | 13 | 30 | 28 |
This study was reviewed and approved by the Ethics
Committee University of Tsukuba Hospital (approval no. 215). All
patients provided informed written consent prior to enrollment.
TaqMan-based reverse
transcription-quantitative PCR (RT-qPCR) assays of miRNA
expression
Total RNA was extracted using the miRNeasy Mini kit
(Qiagen, Venlo, The Netherlands) for cell lines and the miRNeasy
FFPE kit (Qiagen) for FFPE tissues. Reverse transcription was
performed using the TaqMan MicroRNA Reverse Transcription kit
(Applied Biosystems, Foster City, CA, USA). miR-205 expression was
measured using the TaqMan MicroRNA assay system (Applied
Biosystems) according to the manufacturer's instructions. PCR
reactions were performed using the CFX384 Real-Time system (Bio-Rad
Laboratories, Pleasanton, CA, USA). The small RNA, RNU6B, was used
as an internal control. Relative expression was calculated using
the comparative threshold (Ct) method (20).
TaqMan-based RT-qPCR assay of mRNA
expression
Total RNA was extracted using the RNeasy Mini kit
(Qiagen) and reverse-transcribed using the PrimeScript RT Reagent
kit (Takara, Shiga, Japan). PCR reactions were conducted using the
CFX384 Real-Time system (Bio-Rad Laboratories). Relative mRNA
expression was normalized against GAPDH. Relative expression was
calculated by the comparative threshold (Ct) method (20,21).
The following primers were used for RT-qPCR: TIMP-2 forward,
5′-GCCCCCGCCCGCCCAGCCCCCC-3′ and reverse,
5′-GCAACAATATCCACTTTACCAGAGTTAA-3′; and GAPDH forward,
5′-CAACGGATTTGGTCGTATTGG-3′ and reverse,
5′-GCAACAATATCCACTTTACCAGAGTTAA-3′.
Transfection with miR-205-5p mimic or
inhibitor or with siRNA
The cells were cultured to 70–80% confluence in
12-well plates and transfected with 50 nM miR-205-5p mimic or
inhibitor (hsa-miR-205 mirVana™ miRNA Mimic, #4464066;
mirVana™ miRNA Inhibitor, #4464084), or with 50 nM scramble
negative control (mirVana™ miRNA Mimic Negative Control,
#4464058; mirVana™ miRNA Inhibitor Negative Control,
#4464076) from Ambion (Austin, TX, USA). To knock down TIMP-2, the
cells were transfected with 25 nM TIMP-2 siRNA
(Silencer®Select siRNA, #4390824) or control siRNA
(Silencer®Negative control siRNA, #AM4611).
Lipofectamine RNAiMAX transfection reagent (Invitrogen, Carlsbad,
CA, USA) was used according to the manufacturer's instructions. The
cells were incubated at 37°C for 24 h after transfection, the
medium was exchanged for FBS-free medium, and the cells were
incubated for a further 48 h. Both the transfected cells and the
culture media were used for assays and analyses, as described
below.
Cell proliferation, migration and
invasion assays
The cells were cultured as previously described
(14). Cell proliferation assay
was performed as described in a previous study (22). The cells were transfected with 50
nM miR-205-5p mimic or inhibitor by reverse transfection and plated
in 96-well plates at 3×103 cells/well. After 72 h, cell
proliferation was determined by MTT assay using the MTT Cell Count
kit (Nakalai Tesque, Kyoto, Japan). Cell migration was measured
with the 96-well BME Cell-Invasion assay according to the
manufacturer's instructions (Trevigen, Inc., MD, USA). Invasiveness
was assayed by the three-dimensional Matrigel culture method
(23,24) as follows: a total of
8×105 transfected cells were suspended in 200 μl
Matrigel (Corning Inc., New York, NY, USA). A 150-μl drop of
Matrigel mixture containing transfected cells was polymerized on
the bottom of a 6-well microplate and incubated in 2 ml medium for
48 h. Photographs were acquired by fixed-point observation at ×40
magnification under a BZ-X700 microscope (Keyence, Osaka, Japan).
Invasiveness was assessed by the distance of cell migration. The
distance of cell migration represented the length from the gel
surface to the migratory front of cells that had invaded out of the
gel.
Identification of candidate genes
regulated by miR-205-5p
Possible miR-205-5p targets were identified by
bioinformatics analysis with the TargetScan algorithm and
integrated analysis across data for human cancer-cell lines and
mRNA microarray data to identify miRNAs whose expression correlated
with the inverse expression of the mRNA targets predicted in
silico.
Plasmid construction and dual-luciferase
reporter assay
Partial wild-type sequences of the TIMP-2 3′-UTR or
those with a deleted miR-205-5p target sites (positions 590–596 of
the TIMP-2 3′-UTR) were inserted between the
XhoI-PmeI restriction sites in the 3′-UTR of hRluc
gene in the psiCHECK-2 vector (Promega, Madison, WI, USA). The
protocol for vector construction was as previously described
(22,25). The synthesized DNA was cloned into
the psiCHECK-2 vector. The HSC3 cells were transfected with 50 ng
of the vector and 50 nM miR-205-5p mimic using Lipofectamine 3000
(Invitrogen). The activities of firefly and Renilla
luciferases in cell lysates were determined using a Dual-luciferase
Reporter assay system (Promega). Normalized data were calculated as
the ratio Renilla/firefly luciferase activities.
Western blot analysis
The cells were incubated at 37°C for 72 h after
transfection, and then cell lysates were prepared. In addition, the
culture media were centrifuged at 1,000 rpm and the supernatants
were collected. The supernatants were inspissated with a
Vivaspin2-10K (GE Healthcare UK Ltd., Little Chalfont, UK)
according to the manufacturer's instructions. Subsequently, 15
μg of protein lysates were separated on Mini-PROTEIN TGX
Gels (Bio-Rad, Hercules, CA, USA) and then transferred onto
Trans-Blot Turbo Mini PVDF membranes (Bio-Rad). The membranes were
probed with rabbit anti-TIMP-2 antibodies (1:1,000; cat. no. 5738;
Cell Signaling Technology, Danvers, MA, USA), rabbit MT1-MMP
antibodies (1:1,000; cat. no. ab51074; Abcam, Cambridge, UK), and
goat anti-actin antibodies (1:1000; cat. no. sc1615; Santa Cruz
Biotechnology, Inc., Dallas, TX, USA) overnight at 4°C. Proteins
were detected with Chemi-Lumi One Super (Nakalai Tesque), and
images of the blots were obtained with a ChemiDoc XRS Plus system
(Bio-Rad).
Gelatin zymography
MMP-2 enzyme activity was analyzed in the cell
supernatants by gelatin zymography using the Gelatin Zymography kit
(Cosmo Bio Co., Ltd., Tokyo, Japan) according to the manufacturer's
instructions. Images were obtained using the ChemiDoc XRS Plus
system (Bio-Rad).
MMP-2 activity assay
The enzymatic activated MMP-2 content and total
MMP-2 in the cell supernatants were determined using the QuickZyme
Human MMP-2 Activity assay (QuickZyme BioSciences, Leiden, The
Netherlands) according to the manufacturer's instructions.
Statistical analysis
Data for in vitro experiments were evaluated
using the Student's t-test. Associations among more than 3
variables and numerical values were analyzed using the
Kruskal-Wallis test with the Steel-Dwass test. The cut-off value of
miR-205-5p expression in primary OSCC tissues were evaluated by the
receiver operating characteristic (ROC) curve analysis. The
analysis of disease-specific survival was performed by the
Kaplan-Meier method and compared using the log-rank test.
Correlations between miR-205-5p and TIMP-2 expression were
evaluated by Spearman's rank test. All statistical analyses were
performed using JMP version 11 software. A P-value <0.05 was
considered to indicate a statistically significant difference.
Results
miR-205-5p expression in OSCC cell lines
and clinical specimens
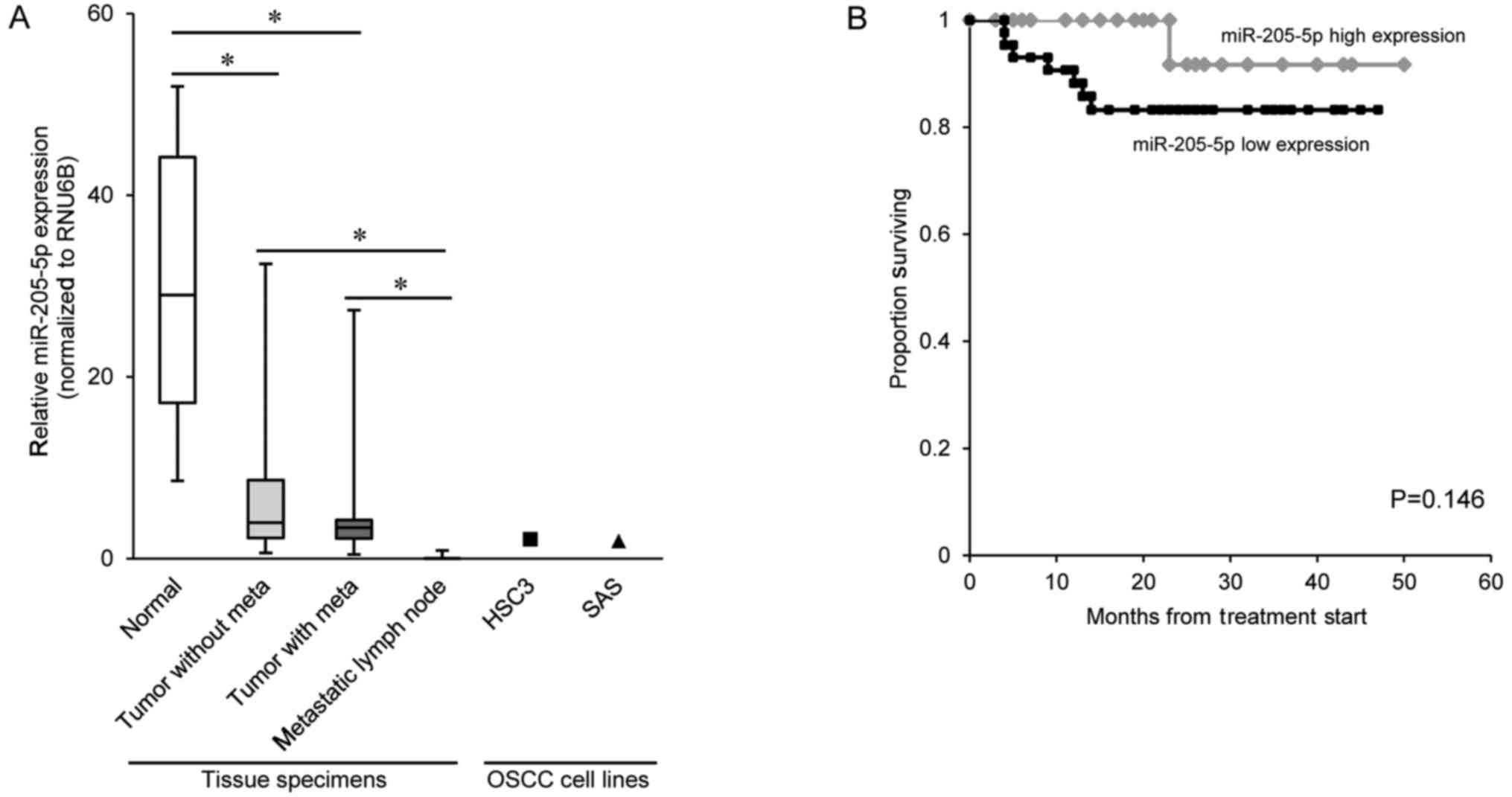
We evaluated miR-205-5p expression in 71 primary
OSCC specimens, 28 metastatic lymph node specimens (patient
characteristic shown in Table I),
and in the HSC3 and SAS cell lines. We found that miR-205-5p
expression was significantly lower in the primary OSCC tissue
samples than in normal oral mucosa tissue samples, and that the
expression was even lower in the metastatic lymph node tissue
samples than in the primary OSCC tissue samples (Fig. 1A). On the other hand, the
miR-205-5p expression was relatively low in primary OSCC tissue
with metastasis compared to that without metastasis (P=0.103).
Furthermore, we evaluated the power of miR-205-5p as the biomarker
of prognosis prediction. We determined the cut-off value (=3.74) of
miR-205-5p in 71 primary OSCC tissue by receiver operating
characteristic (ROC) curve analysis and analyzed disease-specific
survival by the Kaplan-Meier method and compared using the log-rank
test. The disease survival time was calculated from the date of the
patient's first visit. None of the 71 OSCC patients received
pre-operative treatment. Therefore, 'the time of first visit'
roughly equaled 'the time of treatment start'. There was no
difference significant between the miR-205-5p high expression group
and the miR-205-5p low expression group (Fig. 1B).
Effect of miR-205-5p mimic or inhibitor
on cell proliferation, migration and invasion
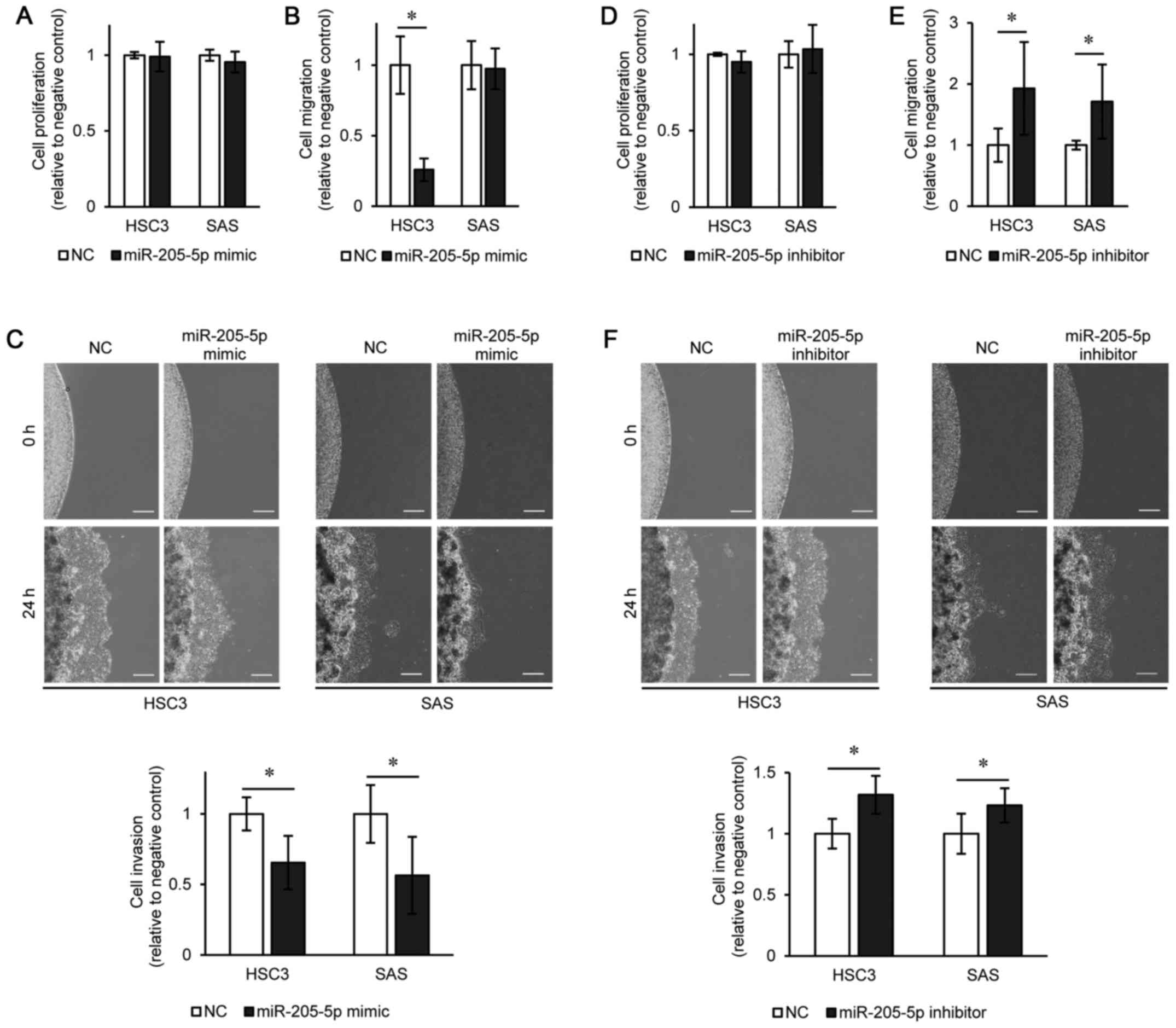
Proliferation was not suppressed in the HSC3 or SAS
cells transfected with miR-205-5p mimic compared to the negative
control (Fig. 2A). HSC3 cell
migration was significantly suppressed by transfection with
miR-205-5p mimic compared to the negative control; however, SAS
cell migration was not affected (Fig.
2B). The invasiveness of both the HSC3 and SAS cells was
significantly suppressed following transfection with miR-205-5p
mimic compared to the negative control (Fig. 2C). The number of cells that had
invaded out of the gel was almost proportional to the migration
distance. On the other hand, no significant differences in the
proliferation of the HSC3 and SAS cell transfected with miR-205-5p
inhibitor were observed compared to the negative control (Fig. 2D). Both migration and invasiveness
were significantly enhanced in the HSC3 and SAS cells transfected
with miR-205-5p inhibitor as compared to the negative control
(Fig. 2E and F).
Identification of miR-205-5p target genes
in OSCC
We screened the TargetScan database for possible
target genes containing a putative miR-205-5p-binding site in their
3′-UTR, and identified 5,974 candidate genes that may be regulated
by miR-205-5p. From these, we selected genes related to TIMPs and
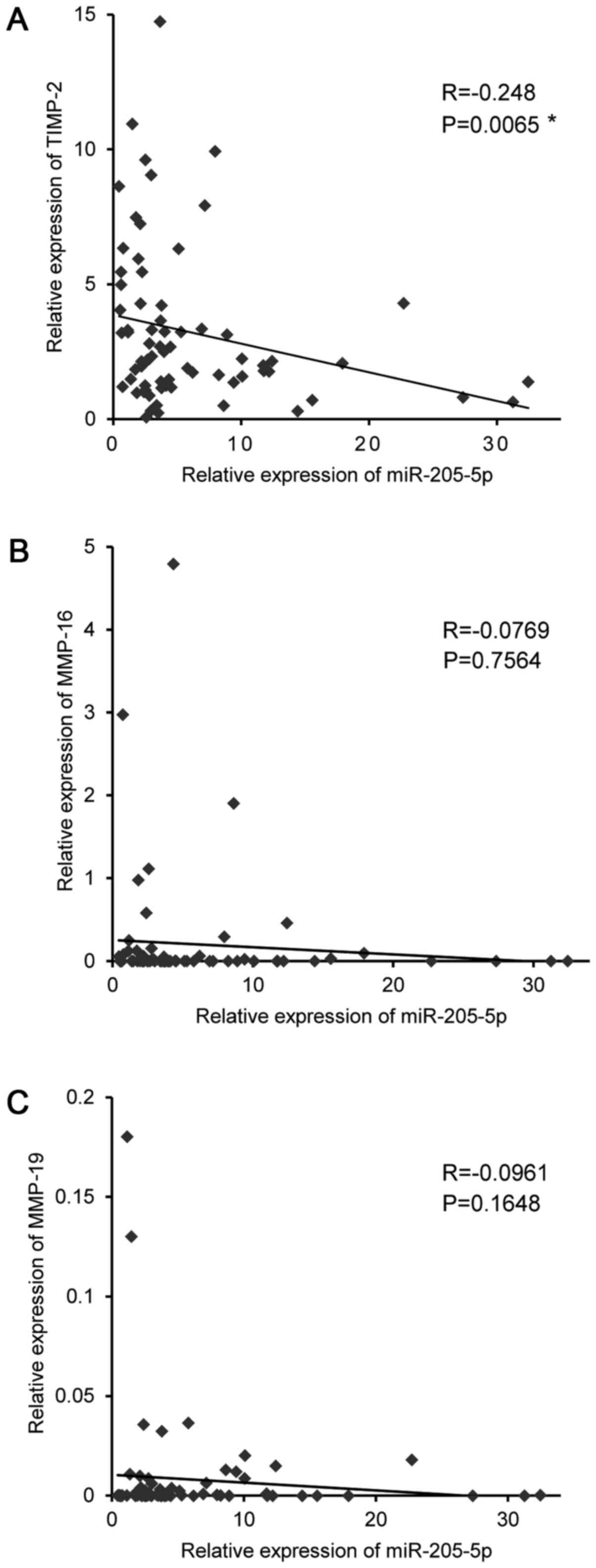
MMPs, and identified 3 candidate miR-205-5p targets (Table II). Among these candidates, we
focused on the TIMP-2 gene, as Spearman's rank test revealed a
significant negative correlation between the mRNA expression of
TIMP-2 and that of miR-205-5p in OSCC specimens (Fig. 3).
| Table IImiR-205-5p candidate target
genes. |
Table II
miR-205-5p candidate target
genes.
| Gene symbol | Representative
transcript | Gene name | Conserved | Poorly
conserved |
|---|
| TIMP-2 | NM_003255 | Metallopeptidase
inhibitor 2 | 0 | 1 |
| MMP-16 | NM_005941 | Matrix
metallopeptidase 16 | 0 | 1 |
| MMP-19 | NM_002429 | Matrix
metallopeptidase 19 | 0 | 1 |
TIMP-2 is a direct target of miR-205-5p
in OSCC cells
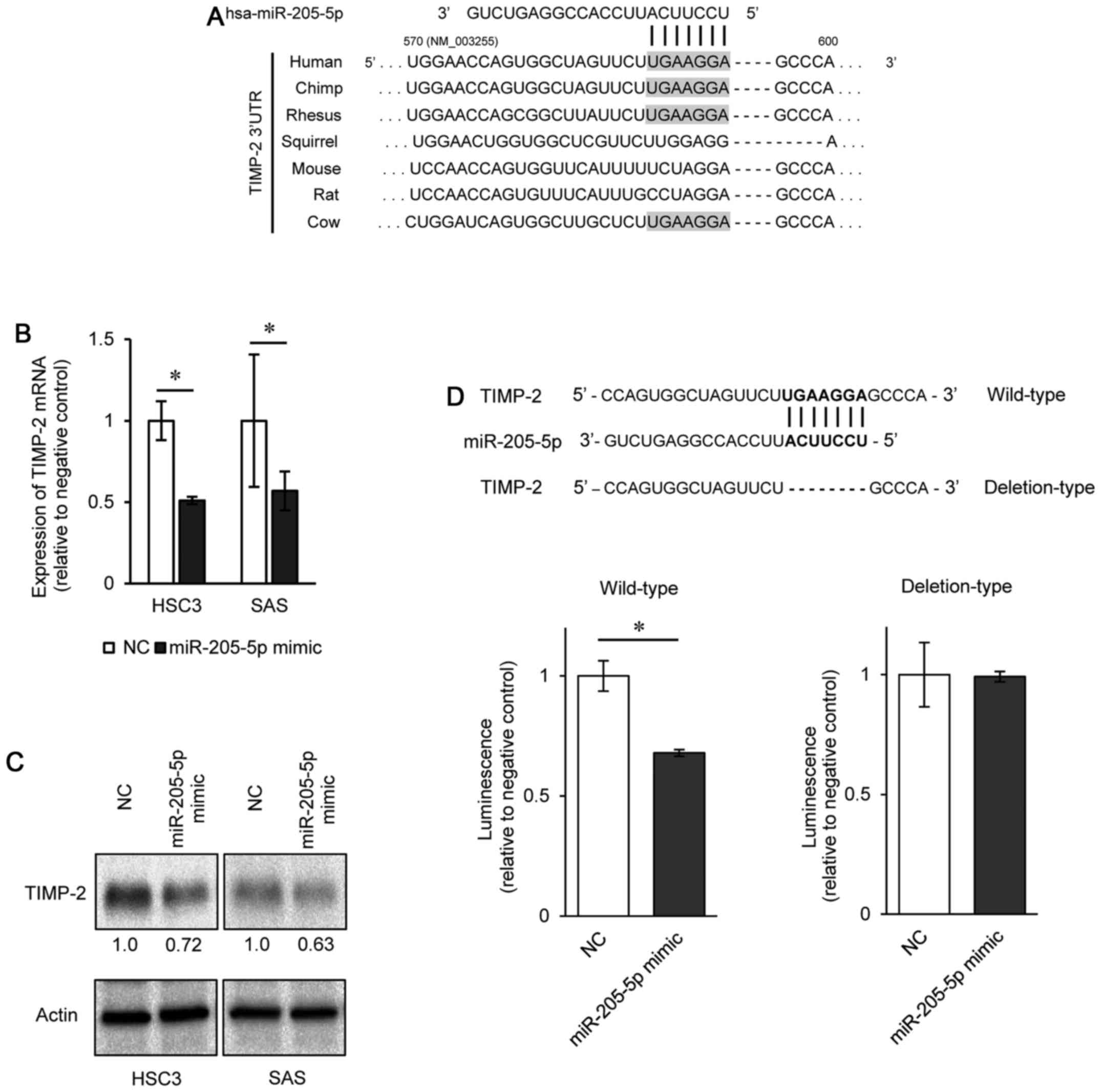
The TargetScan database revealed one putative,
poorly conserved miR-205-5p-binding site in the 3′-UTR of TIMP-2
(position 590–596) (Fig. 4A). We
thus examined TIMP-2 expression in miR-205-5p mimic-transfected
OSCC cells, and found that the TIMP-2 mRNA and protein expression
levels were suppressed in the cells transfected with miR-205-5p
mimic compared to the negative control (Fig. 4B and C). To determine whether
TIMP-2 mRNA contains a target site for miR-205-5p, we conducted
luciferase reporter assays using the HSC3 and SAS cells. The
TargetScan database revealed that there was one putative miR-205-5p
binding site in the TIMP-2 3′-UTR (position 590–596). We used
vectors encoding either a partial wild-type sequence (including the
predicted miR-205-5p target site) or deletion of the seed sequence
of the 3′-UTR of TIMP-2 mRNA. We found that the luciferase activity
was significantly suppressed by co-transfection with miR-205-5p
mimic and the vector carrying the wild-type 3′-UTR of TIMP-2
(Fig. 4D).
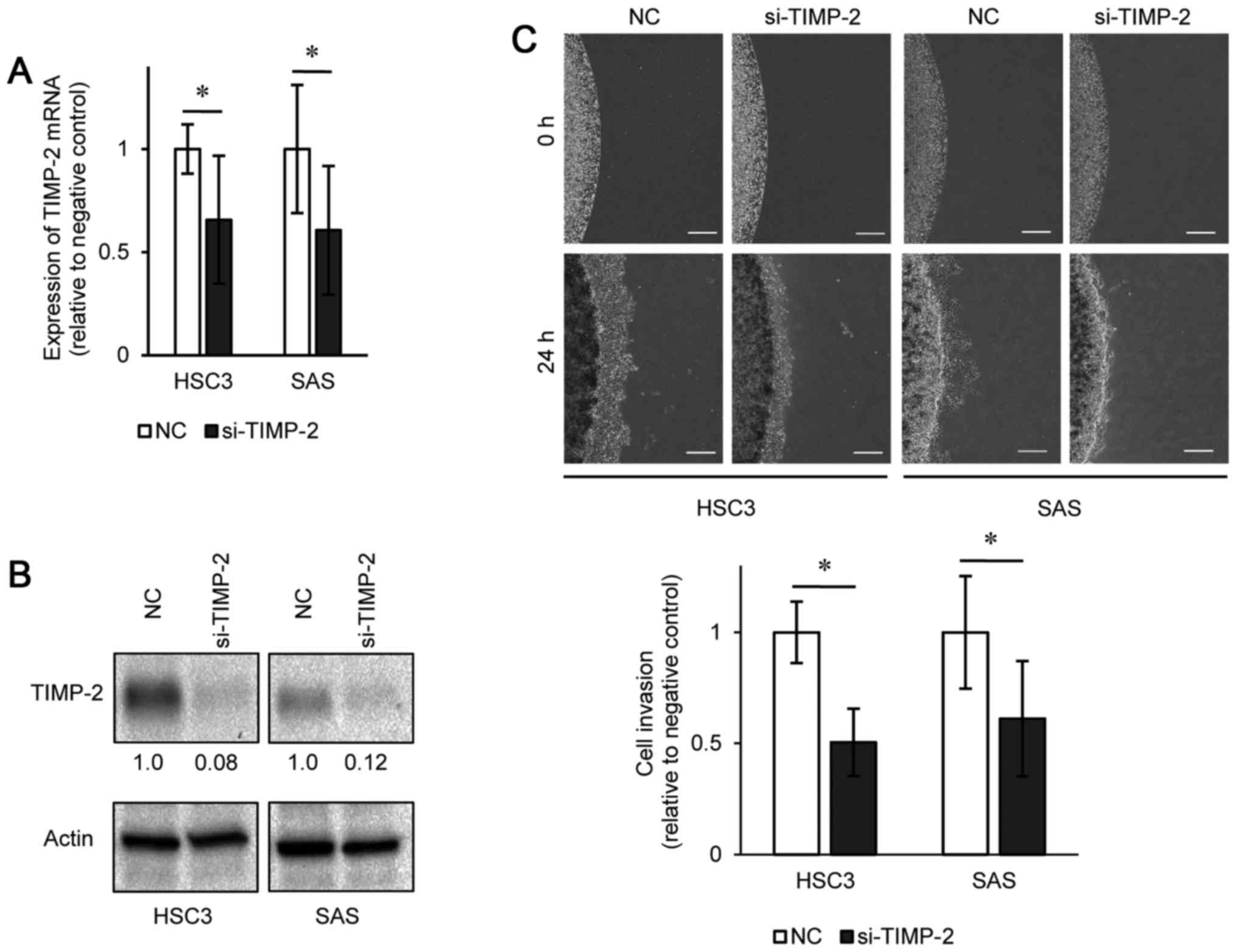
Effect of TIMP-2 silencing on OSCC cell
lines
We then investigated the functions of TIMP-2 in HSC3
and SAS cells by loss-of-function experiments using si-TIMP-2
transfection. The results of western blot analysis and RT-qPCR
indicated that si-TIMP-2 effectively downregulated TIMP-2
expression in both cell lines (Fig. 5A
and B). The invasiveness of both the HSC3 and SAS cells was
suppressed following transfection with si-TIMP-2 compared to the
negative control (Fig. 5C).
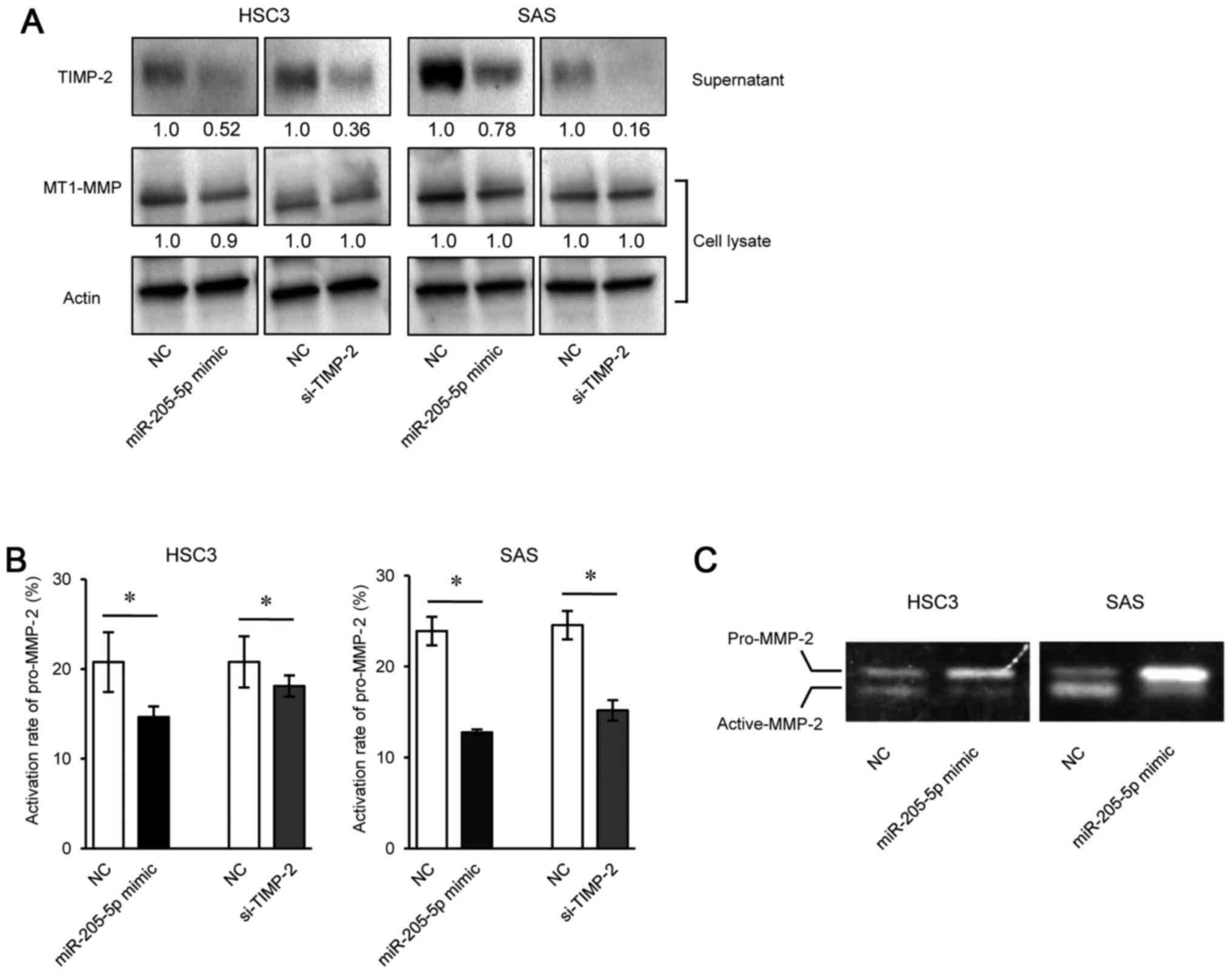
Suppression of TlMP-2 reduces the
activation of pro-MMP-2 via the T1MP-2/MT1-MMP complex
To investigate whether reducing TIMP-2 expression
affects the activation of pro-MMP-2 by MT1-MMP, we used western
blot analysis or gelatin zymography to measure the expression of
TIMP-2, MT1-MMP, pro-MMP-2 and active-MMP-2 in HSC3 and SAS cells
transfected with miR-205-5p mimic or si-TIMP-2. The results of
western blot analysis revealed that TIMP-2 expression was decreased
in the supernatant of both cells transfected with miR-205-5p mimic
or si-TIMP-2 compared to the negative control, and that MT1-MMP
expression was comparable in both cells transfected with miR-205-5p
mimic, si-TIMP-2, or negative control (Fig. 6A). We then measured pro-MMP-2
activation in the cell supernatants (see Materials and methods).
MMP-2 activity assay revealed that the rate of pro-MMP-2 activation
was significantly lower in both cells transfected with miR-205
mimic or si-TIMP-2 than in those transfected with the negative
control (Fig. 6B). The rate of
pro-MMP-2 activation represented the percentage of active-MMP-2 to
total MMP-2 (pro-MMP-2 + active-MMP-2). The results of gelatin
zymography revealed that activated pro-MMP-2 (active-MMP-2) was
decreased in both cells transfected with miR-205-5p mimic compard
with those transfected with the negative control (Fig. 6C).
Discussion
There is growing evidence to indicate the
involvement of miRNAs in several biological processes, including
human oncogenesis and metastasis (26); these include miR-29c, miR-335 and
miR-375 (27,28). Studies have shown that miR-155-5p
is downregulated in OSCC tissue, and that a low miR-155-5p level
correlates with a poor prognosis (14). Therefore, the investigation of
miRNA expression signatures in cancer specimens is an important
research avenue. In the present study, we focused on miR-205-5p,
and yielded 3 major findings: first, that miR-205-5p acted as a
tumor suppressor in OSCC cells (Figs.
1 and 2); second, that
miR-205-5p directly suppressed TIMP-2 expression in OSCC cells
(Fig. 4); and third, that by
suppressing TIMP-2 expression, miR-205-5p inhibited pro-MMP-2
activation (Fig. 6).
In this study, we observed that miR-205-5p
expression was significantly downregulated in primary OSCC tissues
compared to normal oral mucosa tissue, and that the expression was
even lower in metastatic lymph node tissue than in primary OSCC
tissue (Fig. 1A). We also
demonstrated that transfection with miR-205-5p suppressed the
invasiveness of both the HSC3 and SAS cells (Fig. 2C). These results suggest that
miR-205-5p acts as a tumor suppressor miRNA in OSCC, and that a
reduced miR-205-5p expression may significantly contribute to OSCC
invasion and metastasis. Recent studies have indicated that
miR-205-5p is downregulated in several types of cancer, including
breast, prostate and bladder cancer (29-31),
and that miR-205-5p suppresses several tumor types by targeting
oncogenes (16,19,32–34).
Other studies have shown that miR-205-5p suppresses cell
invasiveness in several types of cancer (18,35).
The role of miR-205-5p in suppressing cell invasion has also been
confirmed in in vivo models of breast cancer (30). These findings are consistent with
those of the present study. By contrast, Kaplan-Meier analysis
revealed that there was no difference between the miR-205-5p high
expression group and the miR-205-5p low expression group (Fig. 1B). A recent study also demonstrated
that a low expression of miR-205 was associated with a poor
prognosis in cervical cancer (36). Another previous study demonstrated
that a low expression of both miR-205 and let-7d was associated
with a poor prognosis in head and neck squamous-cell carcinoma
(HNSCC); however, this association was not observed with a low
expression of miR-205 only (37).
To reveal the correlation between the expression of miR-205-5p and
disease-specific survival in OSCC, further research is required,
examining more cases of OSCC.
A single miRNA can regulate several protein-coding
genes. Indeed, bioinformatics analyses have shown that at least
30-60% of the protein-coding genes in the human genome are
regulated by miRNAs (38).
Therefore, a reduced expression of tumor suppressor miRNAs may
cause an overexpression of oncogenic genes in cancer cells.
In this study, we identified TIMP-2 as a target of
miR-205-5p. Other recent studies have shown that TIMP-2 is an
oncogenic factor in several types of cancer (39,40).
Our data demonstrated a significant negative correlation between
the mRNA expression of TIMP-2 and that of miR-205-5p in OSCC
tissues (R=−0.248, P=0.0065) (Fig.
3). Furthermore, our luciferase reporter assays confirmed that
miR-205-5p miRNA directly binds the 3′-UTR of TIMP-2 (Fig. 4D). To the best of our knowledge,
this is the first study to show that TIMP-2 is directly regulated
by miR-205-5p in OSCC cells.
TIMP-2 is known to regulate MMPs through its
enzymatic activity. The data from the present study demonstrated
that silencing TIMP-2 suppressed the invasiveness of OSCC cells
(Fig. 5), suggesting that TIMP-2
promotes OSCC tumor progression. Recent immunohistochemical
analyses have identified an elevated TIMP-2 expression as an
indicator of aggressive behavior and a poor prognosis in patients
with HNSCC (41), and demonstrated
a strong correlation between a marked TIMP-2 expression and
lymph-node metastasis, as well as a poor prognosis, in the early
stages of OSCC (41–43). These data are consistent with the
findings of the present study, indicating that TIMP-2 may promote
OSCC tumor progression. Recent studies have shown that TIMP-2
promotes tumor progression via the activation of MMP-2, which is
mediated by MT1-MMP (9,10). TIMP-2 was long thought to suppress
tumor invasion by inhibiting MMP-2 in general, and when
overexpressed, TIMP-2 inhibited MMP-2 in vitro. However,
according to a model of cell-mediated MMP-2 activation, pro-MMP-2
binds TIMP-2 in a complex with MT1-MMP on the cell surface, forming
a pro-MMP-2/TIMP-2/MT1-MMP ternary complex. In this case, pro-MMP-2
is activated by the adjacent MT1-MMP that is not bound to TIMP-2
(9,10,44).
In the present study, although we observed that TIMP-2 expression
was decreased in the supernatant of miR-205-5p mimic-transfected
HSC3 and SAS cells, MT1-MMP expression was not affected (Fig. 6A). In addition, pro-MMP-2 was
activated at a lower rate in miR-205-5p mimic-transfected cells
(Fig. 6B and C). These results
suggest that miR-205-5p suppresses pro-MMP-2 activation by
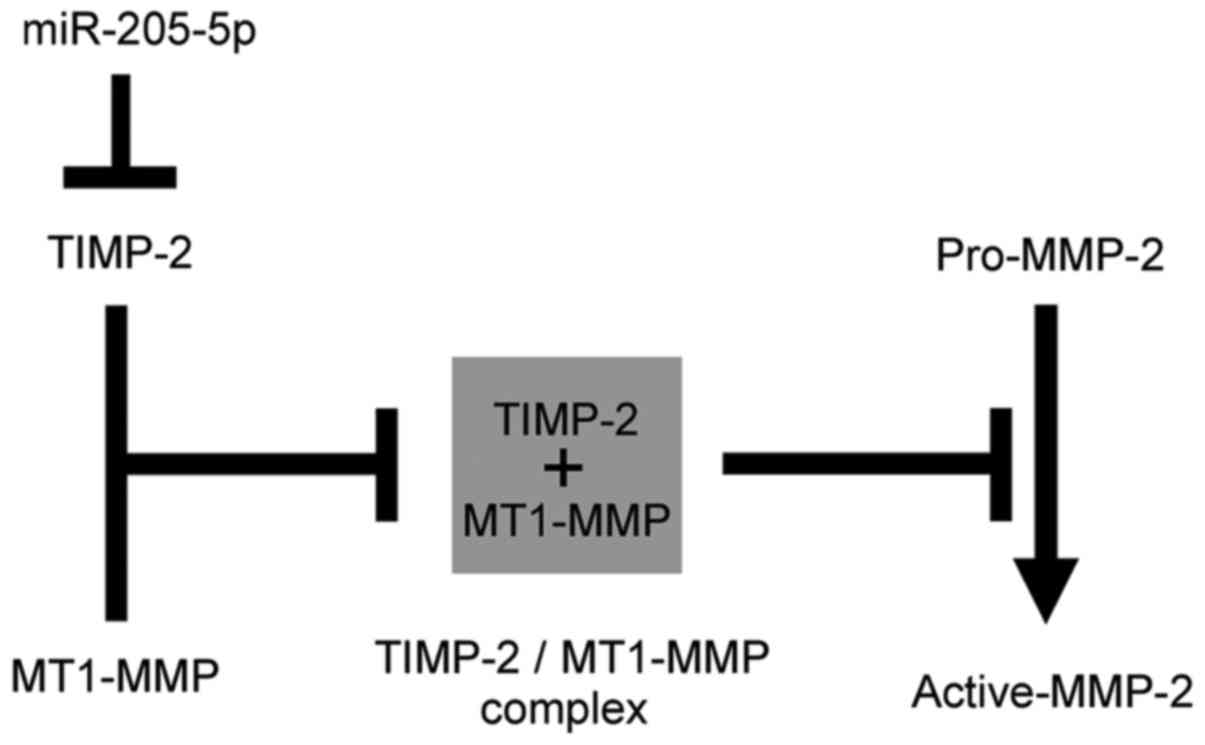
regulating TIMP-2, as illustrated in Fig. 7. Recent studies have shown that
TIMP-2 enhances pro-MMP-2 activation via MT1-MMP in vivo
(45–49), and that upregulating TIMP-2
promotes MMP-2 activation and the invasiveness of glioma cells
(47). Another study demonstrated
that the ratio of MMP-2 activation strongly correlated with TIMP-2
expression in SCC of the tongue (42). These findings are consistent with
those of the present study. Pro-MMP-2 activation depends on the
local TIMP-2 concentration; this activation occurs at a low
concentration of TIMP-2 relative to MT1-MMP, leaving sufficient
inhibitor-free MT1-MMP to initiate the activation of pro-MMP-2
(50,51). On the other hand, high TIMP-2
levels inhibit MMP-2 activation by blocking all of the available
MT1-MMP molecules (52). In the
present study, the amount of TIMP-2 secreted into the culture
medium appeared to be appropriate for regulating MMP-2 activation
in HSC3 and SAS cells. To further determine the role of miR-205-5p
in the regulation of TIMP-2 expression in OSCC, further studies
using animal models are warranted.
In conclusoin, the present study demonstrates that
miR-205-5p functions as a tumor suppressor in OSCC, that miR-205-5p
directly regulates TIMP-2 expression, and that miR-205-5p
suppresses pro-MMP-2 activation by regulating TIMP-2 expression in
OSCC cells (Fig. 7).
This identification of the tumor suppressor role of
miR-205-5p in OSCC may have significant therapeutic potential.
Currently, RNA interference is being implemented as a gene-specific
approach in molecular medicine. Thus, it may be possible to use
miR-205-5p to regulate specific genes related to tumor progression
as a novel therapeutic approach to the treatment of OSCC.
Acknowledgments
This study was supported by JSPS KAKENHI (grant nos.
25893024 and 15H05038), and was partly supported by the Extramural
Collaborative Research Grant of the Cancer Research Institute,
Kanazawa University.
Notes
[1] Competing
interests
The authors declare that they have no competing
interests.
References
|
1
|
Bhattacharya A, Roy R, Snijders AM,
Hamilton G, Paquette J, Tokuyasu T, Bengtsson H, Jordan RCK, Olshen
AB, Pinkel D, et al: Two distinct routes to oral cancer differing
in genome instability and risk for cervical node metastasis. Clin
Cancer Res. 17:7024–7034. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Ferlito A, Rinaldo A, Robbins KT, Leemans
CR, Shah JP, Shaha AR, Andersen PE, Kowalski LP, Pellitteri PK,
Clayman GL, et al: Changing concepts in the surgical management of
the cervical node metastasis. Oral Oncol. 39:429–435. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Greenberg JS, El Naggar AK, Mo V, Roberts
D and Myers JN: Disparity in pathologic and clinical lymph node
staging in oral tongue carcinoma. Implication for therapeutic
decision making. Cancer. 98:508–515. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Shingaki S, Takada M, Sasai K, Bibi R,
Kobayashi T, Nomura T and Saito C: Impact of lymph node metastasis
on the pattern of failure and survival in oral carcinomas. Am J
Surg. 185:278–284. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Woolgar JA, Rogers SN, Lowe D, Brown JS
and Vaughan ED: Cervical lymph node metastasis in oral cancer: The
importance of even microscopic extracapsular spread. Oral Oncol.
39:130–137. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Wikner J, Grobe A, Pantel K and Riethdorf
S: Squamous cell carcinoma of the oral cavity and circulating
tumour cells. World J Clin Oncol. 5:114–124. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Bhuvarahamurthy V, Kristiansen GO,
Johannsen M, Loening SA, Schnorr D, Jung K and Staack A: In situ
gene expression and localization of metalloproteinases MMP1, MMP2,
MMP3, MMP9, and their inhibitors TIMP1 and TIMP2 in human renal
cell carcinoma. ; Oncol Rep. 15:1379–1384. 2006.PubMed/NCBI
|
|
8
|
Kusukawa J, Sasaguri Y, Shima I, Kameyama
T and Morimatsu M: Expression of matrix metalloproteinase-2 related
to lymph node metastasis of oral squamous cell carcinoma. A
clinicopathologic study. Am J Clin Pathol. 99:18–23. 1993.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Imai K, Ohuchi E, Aoki T, Nomura H, Fujii
Y, Sato H, Seiki M and Okada Y: Membrane-type matrix
metalloproteinase 1 is a gelatinolytic enzyme and is secreted in a
complex with tissue inhibitor of metalloproteinases 2. Cancer Res.
56:2707–2710. 1996.PubMed/NCBI
|
|
10
|
Strongin AY, Collier I, Bannikov G, Marmer
BL, Grant GA and Goldberg GI: Mechanism of cell surface activation
of 72-kDa type IV collagenase Isolation of the activated form of
the membrane metalloprotease. J Biol Chem. 270:5331–5338. 1995.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Brodersen P and Voinnet O: Revisiting the
principles of microRNA target recognition and mode of action. Nat
Rev Mol Cell Biol. 10:141–148. 2009. View
Article : Google Scholar : PubMed/NCBI
|
|
12
|
Fukumoto I, Koshizuka K, Hanazawa T,
Kikkawa N, Matsushita R, Kurozumi A, Kato M, Okato A, Okamoto Y and
Seki N: The tumor-suppressive microRNA-23b/27b cluster regulates
the MET oncogene in oral squamous cell carcinoma. Int J Oncol.
49:1119–1129. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Fukumoto I, Hanazawa T, Kinoshita T,
Kikkawa N, Koshizuka K, Goto Y, Nishikawa R, Chiyomaru T, Enokida
H, Nakagawa M, et al: MicroRNA expression signature of oral
squamous cell carcinoma: Functional role of microRNA-26a/b in the
modulation of novel cancer pathways. Br J Cancer. 112:891–900.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Baba O, Hasegawa S, Nagai H, Uchida F,
Yamatoji M, Kanno NI, Yamagata K, Sakai S, Yanagawa T and Bukawa H:
MicroRNA-155-5p is associated with oral squamous cell carcinoma
metastasis and poor prognosis. J Oral Pathol. 45:248–255. 2016.
View Article : Google Scholar
|
|
15
|
He S, Lai R, Chen D, Yan W, Zhang Z, Liu
Z, Ding X and Chen Y: Downregulation of miR-221 inhibits cell
migration and invasion through targeting methyl-CpG binding domain
protein 2 in human oral squamous cell carcinoma cells. BioMed Res
Int. 2015:7516722015. View Article : Google Scholar
|
|
16
|
Elgamal OA, Park JK, Gusev Y,
Azevedo-Pouly AC, Jiang J, Roopra A and Schmittgen TD: Tumor
suppressive function of mir-205 in breast cancer is linked to HMGB3
regulation. PLoS One. 8:e764022013. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Iorio MV, Casalini P, Piovan C, Di Leva G,
Merlo A, Triulzi T, Ménard S, Croce CM and Tagliabue E:
microRNA-205 regulates HER3 in human breast cancer. Cancer Res.
69:2195–2200. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Majid S, Dar AA, Saini S, Yamamura S,
Hirata H, Tanaka Y, Deng G and Dahiya R: MicroRNA-205-directed
transcriptional activation of tumor suppressor genes in prostate
cancer. Cancer. 116:5637–5649. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Matsushima K, Isomoto H, Yamaguchi N,
Inoue N, Machida H, Nakayama T, Hayashi T, Kunizaki M, Hidaka S,
Nagayasu T, et al: miRNA-205 modulates cellular invasion and
migration via regulating zinc finger E-box binding homeobox 2
expression in esophageal squamous cell carcinoma cells. J Transl
Med. 9:302011. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Wang G, Wang L, Sun S, Wu J and Wang Q:
Quantitative measurement of serum microRNA-21 expression in
relation to breast cancer metastasis in Chinese females. Ann Lab
Med. 35:226–232. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Wei J, Zhang L, Li J, Zhu S, Tai M, Mason
CW, Chapman JA, Reynolds EA, Weiner CP and Zhou HH: MicroRNA-205
promotes cell invasion by repressing TCF21 in human ovarian cancer.
J Ovarian Res. 10:332017. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Kinoshita T, Nohata N, Hanazawa T, Kikkawa
N, Yamamoto N, Yoshino H, Itesako T, Enokida H, Nakagawa M, Okamoto
Y, et al: Tumour-suppressive microRNA-29s inhibit cancer cell
migration and invasion by targeting laminin-integrin signalling in
head and neck squamous cell carcinoma. Br J Cancer. 109:2636–2645.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Lee GY, Kenny PA, Lee EH and Bissell MJ:
Three-dimensional culture models of normal and malignant breast
epithelial cells. Nat Methods. 4:359–365. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Sakr M, Takino T, Sabit H, Nakada M, Li Z
and Sato H: miR-150 5p and miR-133a suppress glioma cell
proliferation and migration through targeting membrane-type-1
matrix metalloproteinase. Gene. 587:155–162. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Nishikawa R, Goto Y, Kojima S, Enokida H,
Chiyomaru T, Kinoshita T, Sakamoto S, Fuse M, Nakagawa M, Naya Y,
et al: Tumor-suppressive microRNA-29s inhibit cancer cell migration
and invasion via targeting LAMC1 in prostate cancer. Int J Oncol.
45:401–410. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Nelson KM and Weiss GJ: MicroRNAs and
cancer: Past, present, and potential future. Mol Cancer Ther.
7:3655–3660. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Liu L, Bi N, Wu L, Ding X, Men Y, Zhou W,
Li L, Zhang W, Shi S, Song Y, et al: MicroRNA-29c functions as a
tumor suppressor by targeting VEGFA in lung adenocarcinoma. Mol
Cancer. 16:502017. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Zhang JK, Li YS, Zhang CD and Dai DQ:
Up-regulation of CRKL by microRNA-335 méthylation is associated
with poor prognosis in gastric cancer. Cancer Cell Int. 17:282017.
View Article : Google Scholar
|
|
29
|
Chen Z, Tang ZY, He Y, Liu LF, Li DJ and
Chen X: miRNA-205 is a candidate tumor suppressor that targets ZEB2
in renal cell carcinoma. Oncol Res Treat. 37:658–664. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Wu H, Zhu S and Mo YY: Suppression of cell
growth and invasion by miR-205 in breast cancer. Cell Res.
19:439–448. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Wang N, Li Q, Feng NH, Cheng G, Guan ZL,
Wang Y, Qin C, Yin CJ and Hua LX: miR-205 is frequently
downregulated in prostate cancer and acts as a tumor suppressor by
inhibiting tumor growth. Asian J Androl. 15:735–741. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Gregory PA, Bert AG, Paterson EL, Barry
SC, Tsykin A, Farshid G, Vadas MA, Khew-Goodall Y and Goodall GJ:
The miR-200 family and miR-205 regulate epithelial to mesenchymal
transition by targeting ZEB1 and SIPl. Nat Cell Biol. 10:593–601.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Yue X, Wang P, Xu J, Zhu Y, Sun G, Pang Q
and Tao R: MicroRNA-205 functions as a tumor suppressor in human
glioblastoma cells by targeting VEGF-A. Oncol Rep. 27:1200–1206.
2012. View Article : Google Scholar
|
|
34
|
Adachi R, Horiuchi S, Sakurazawa Y,
Hasegawa T, Sato K and Sakamaki T: ErbB2 down-regulates
microRNA-205 in breast cancer. Biochem Biophys Res Commun.
411:804–808. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Zhang H and Fan Q: MicroRNA-205 inhibits
the proliferation and invasion of breast cancer by regulating AMOT
expression. Oncol Rep. 34:2163–2170. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Pang H and Yue X: miR-205 serves as a
prognostic factor and suppresses proliferation and invasion by
targeting insulin-like growth factor receptor 1 in human cervical
cancer. Tumour Biol. 39:10104283177013082017. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Childs G, Fazzari M, Kung G, Kawachi N,
Brandwein-Gensler M, McLemore M, Chen Q, Burk RD, Smith RV,
Prystowsky MB, et al: Low-level expression of microRNAs let-7d and
miR-205 are prognostic markers of head and neck squamous cell
carcinoma. Am J Pathol. 174:736–745. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Lewis BP, Burge CB and Bartel DP:
Conserved seed pairing, often flanked by adenosines, indicates that
thousands of human genes are microRNA targets. Cell. 120:15–20.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Grignon DJ, Sakr W, Toth M, Ravery V,
Angulo J, Shamsa F, Pontes JE, Crissman JC and Fridman R: High
levels of tissue inhibitor of metalloproteinase-2 (TIMP-2)
expression are associated with poor outcome in invasive bladder
cancer. Cancer Res. 56:1654–1659. 1996.PubMed/NCBI
|
|
40
|
Ree AH, Florenes VA, Berg JP, Maelandsmo
GM, Nesland JM and Fodstad O: High levels of messenger RNAs for
tissue inhibitors of metalloproteinases (TIMP-1 and TIMP-2) in
primary breast carcinomas are associated with development of
distant metastases. Clin Cancer Res. 3:1623–1628. 1997.
|
|
41
|
Ondruschka C, Buhtz P, Motsch C, Freigang
B, Schneider-Stock R, Roessner A and Boltze C: Prognostic value of
MMP-2, -9 and TIMP-1,-2 immunoreactive protein at the invasive
front in advanced head and neck squamous cell carcinomas. Pathol
Res Pract. 198:509–515. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Yoshizaki T, Maruyama Y, Sato H and
Furukawa M: Expression of tissue inhibitor of matrix
metalloproteinase-2 correlates with activation of matrix
metalloproteinase-2 and predicts poor prognosis in tongue squamous
cell carcinoma. Int J Cancer. 95:44–50. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Katayama A, Bandoh N, Kishibe K, Takahara
M, Ogino T, Nonaka S and Harabuchi Y: Expressions of matrix
metalloproteinases in early-stage oral squamous cell carcinoma as
predictive indicators for tumor metastases and prognosis. Clin
Cancer Res. 10:634–640. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Sato H, Takino T, Kinoshita T, Imai K,
Okada Y, Stetler Stevenson WG and Seiki M: Cell surface binding and
activation of gelatinase A induced by expression of
membrane-type-1-matrix metalloproteinase (MT1-MMP). FEBS Lett.
385:238–240. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Hernandez-Barrantes S, Toth M, Bernardo
MM, Yurkova M, Gervasi DC, Raz Y, Sang QA and Fridman R: Binding of
active (57 kDa) membrane type 1-matrix metalloproteinase (MT1-MMP)
to tissue inhibitor of metalloproteinase (TIMP)-2 regulates MT1-MMP
processing and pro-MMP-2 activation. J Biol Chem. 275:12080–12089.
2000. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Bernardo MM and Fridman R: TIMP-2 (tissue
inhibitor of metalloproteinase-2) regulates MMP-2 (matrix
metalloproteinase-2) activity in the extracellular environment
after pro-MMP-2 activation by MT1 (membrane type 1)-MMP. Biochem J.
374:739–745. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Lu KV, Jong KA, Rajasekaran AK, Cloughesy
TF and Mischel PS: Upregulation of tissue inhibitor of
metalloproteinases (TIMP)-2 promotes matrix metalloproteinase
(MMP)-2 activation and cell invasion in a human glioblastoma cell
line. Lab Invest. 84:8–20. 2004. View Article : Google Scholar
|
|
48
|
Kudo T, Takino T, Miyamori H, Thompson EW
and Sato H: Substrate choice of membrane-type 1 matrix
metalloproteinase is dictated by tissue inhibitor of
metalloproteinase-2 levels. Cancer Sci. 98:563–568. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Nishida Y, Miyamori H, Thompson EW, Takino
T, Endo Y and Sato H: Activation of matrix metalloproteinase-2
(MMP-2) by membrane type 1 matrix metalloproteinase through an
artificial receptor for proMMP-2 generates active MMP-2. Cancer
Res. 68:9096–9104. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Caterina JJ, Yamada S, Caterina NC,
Longenecker G, Holmbäck K, Shi J, Yermovsky AE, Engler JA and
Birkedal-Hansen H: Inactivating mutation of the mouse tissue
inhibitor of metalloproteinases-2 (Timp-2) gene alters proMMP-2
activation. J Biol Chem. 275:26416–26422. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Wang Z, Juttermann R and Soloway PD:
TIMP-2 is required for efficient activation of proMMP-2 in vivo. J
Biol Chem. 275:26411–26415. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Itoh Y, Ito A, Iwata K, Tanzawa K, Mori Y
and Nagase H: Plasma membrane-bound tissue inhibitor of
metalloproteinases (TIMP)-2 specifically inhibits matrix
metalloproteinase 2 (gelatinase A) activated on the cell surface. J
Biol Chem. 273:24360–24367. 1998. View Article : Google Scholar : PubMed/NCBI
|