Introduction
Ubiquitination is a post-translational modification
(PTM) in which ubiquitin binds to substrate proteins. Ubiquitin is
a 76 amino acid-long polypeptide protein that is covalently
attached to target proteins through an isopeptide bond between the
glycine at the carboxyl (C)-terminus of ubiquitin and the lysine at
the amino (N)-terminus of substrate proteins (1,2).
This reaction is carried out through a three-step process involving
ubiquitin-activating enzymes (E1s), ubiquitin-conjugating enzymes
(E2s) and ubiquitin ligases (E3s) (1–3).
There ubiquitin protein comprises seven lysine residues, K6, K11,
K27, K29, K33, K48 and K63, on which polyubiquitin chains may form.
Polyubiquitination may regulate a number of different cellular
functions, such as proteasomal degradation, mitophagy, translation,
receptor, endocytosis and sorting (4–6). The
K48-linked polyubiquitin chain has been previously reported to
induce proteasomal degradation of target substrate, whereas the
K63-linked polyubiquitin chain affects intracellular signaling, DNA
repair, endosomal-lysosomal pathway and degradation of proteins by
autophagy (7–9).
By contrast, deubiquitination is the process in
which ubiquitins are detached from target proteins.
Deubiquitinating enzymes (DUBs) remove ubiquitins from target
proteins by cleaving the isopeptide bond between the ubiquitin and
the protein. Approximately 100 DUB proteins are encoded in the
human genome, which are classified into 2 main categories: i)
Cysteine proteases, which include the ubiquitin-specific protease
(USP), the ubiquitin C-terminal hydrolase (UCH), the
Machado-Josephin disease protein (MJD), the ovarian tumor (OTU) and
the monocyte chemotactic protein-induced protease, and permutated
papain fold peptidases of dsRNA viruses and eukaryotes (PPPDE)
protein families; and ii) metalloproteases, including the
Jab1/Mov34/Mpr1-Pad1 N-terminal+ (JAMM) family (10). Of these, the USP protein family
contains the largest number of DUBs, which have three conserved
motifs, Cys-box, Asp/Asn-box and His-box, that are essential for
catalysis (11,12). Members of the UCH family share
close homology in their catalytic domains (13). OTU family members share homology
with the OTU gene and are known to regulate crucial
signaling pathways, including interferon, NF-κB p97-mediated
processes and DNA damage response (14,15).
Among the MJD family members, Ataxin 3 is known to regulate the
retrotranslocation of endoplasmic reticulum-associated degradation
substrates (16). Members of the
PPPDE family are reported to regulate the cell cycle in eukaryotes
(10). There are four different
JAMM domains in JAMM family; of these, three have been reported to
deubiquitinate their substrates, and one is related to
ubiquitin-like modification of Nedd8 (17,18).
Deubiquitination serves pivotal roles in cellular
homeostasis. For example, DUBs regulate DNA repair, protein
degradation, apoptosis, cell cycle and immune response (19,20).
The abnormal expression of DUBs may result in human diseases owing
to the misregulation of homeostasis and DUBs have been targeted for
treating diseases including cancer (21,22).
Therefore, DUB screening may be beneficial in analyzing biological
mechanisms and in establishing biomarkers for medical
diagnoses.
A biomarker is a biological molecule that may be
present as one of the components in the circulatory system,
including whole blood, serum, plasma and secretion. As it is also
detectable in specific tissues and body fluids, it may provide an
indication of the biological signs of abnormal processes and
diseases (23). In addition,
biomarkers may be valuable tracers that indicate the status of
human body, such that they may aid in determining prognosis,
progression and recurrence of the diseases (23). Clinical responses to treatments and
therapies may also be predicted by biomarkers (23). Thus, the discovery of new
biomarkers may be important in diagnosing and predicting various
human diseases, such as cancer. Multiplex polymerase chain reaction
(PCR) has been used to detect the level of gene expression in
biological samples (24).
Multiplex PCR is able to amplify several target genes by mixing
multiple primer pairs with different specificities to respective
genes (24). Therefore, it is a
useful tool to check the expression levels of various genes
properly and efficiently (24). By
using this method, gene expression levels in biological samples can
be examined, which may subsequently lead to the identification of
putative biomarkers. The aim of the present study was to develop a
screening tool for the identification of putative biomarkers using
multiplex PCR and primers for various DUB genes to detect
and quantify the mRNA expression levels of multiple DUBs
simultaneously. Therefore, the multiplex PCR platform for DUB
screening may be an important tool for biomarker
identification.
p53 is a tumor suppressor that serves an important
role in biological processes; for example, during cellular stress,
p53 is activated and may lead to cell cycle arrest or activate DNA
repair (24); if damaged DNA is
unable to be repaired, p53 induces apoptosis (25,26).
In the present study, differentially expressed DUBs were identified
that may be associated with the presence or absence of p53.
Multiplex PCR was performed to identify DUB genes that are
related to p53 signaling and the expression levels of these DUBs
were examined between HCT116 p53+/+ and
p53−/− cells.
USP5 was previously reported to decrease stability
of p53 (27). Ubiquitinated p53
competes with free polyubiquitin for recognition by proteasomal
degradation following suppression of USP5, and this competition
inhibits proteasomal degradation of p53 rather than decreases p53
ubiquitination. Moreover, suppression of USP5 increases p53 and FAS
levels in melanoma cells through the BRAF pathway (28). OTUD6A is a member of OTU family,
which is an important regulator for cell signaling cascade
(14). The OTU family categorizes
as the OTUB subfamily/Otubains, the OTUD subfamily, the A20-like
subfamily and the OTULIN subfamily (14). In contrast to the USP family, which
has an effect on most types of ubiquitin chains, the OTU family is
linkage-specific (14). However,
the function of OTUD6A is unknown. Results from the present study
DUB screening with multiplex PCR and subsequent protein expression
analysis revealed that the expression of USP5 and
OTUD6A were differentially expressed in HCT116
p53+/+ and p53−/− cells, which suggested that
USP5 and OTUD6A may be associated with p53.
Materials and methods
Cell culture, transfection and small
interfering (si)RNA treatment
Human colon cancer HCT116 p53+/+ and
HCT116 p53−/− cells were provide by Dr Albert J. Fornace
(Georgetown University, Washington, DC, USA) and were grown in
RPMI-1640 medium containing 10% FBS (both from Gibco; Thermo Fisher
Scientific, Inc., Waltham, MA, USA), and 1% penicillin/streptomycin
(Corning Life Sciences, Tewksbury, MA, USA) at 37°C in 5%
CO2 atmosphere.
Full-length p53 cDNA was subcloned into
pcDNA3-HA vector from a pcDNA3-Myc-p53 vector used in our
previous study (12). For
transfection of pcDNA3-HA-p53 into HCT116 p53+/+
and p53−/− cells, the cells were seeded
(8×105 cells/dish) in 60-mm culture dishes. Next day,
pcDNA3-HA-p53 was mixed with 10 mM polyethylenimine (PEI;
Polysciences, Inc., Warrington, PA, USA) and 150 mM NaCl was used,
and incubated for 15 min at room temperature. The construct was
transfected into both HCT116 p53+/+ and
p53−/− cells at various concentrations (0, 0.8, 1.5 and
3 µg) and an empty vector (pcDNA3-HA) was used for a control. The
amount of cDNA for transfection was optimized for subsequent
experiments based on the expression level of HA-p53 protein
following transfection with the differing amounts of cDNA
aforementioned. The cells were incubated at 37°C in 5%
CO2 atmosphere for 24 h and subsequently harvested for
further experimentation.
For p53 knockdown, HCT116 p53+/+ cells
were seeded (8×105 cells/dish) in 60-nm dishes 1 day
prior to transfection. p53-targeted siRNAs (si-p53) or
negative control siRNA (si-Ctrl; cat. no. SN-1001-CFG; Bioneer
Corporation, Daejeon, Korea) were transfected into HCT116
p53+/+ cells using Opti-MEM and
Lipofectamine® RNAiMAX (Invitrogen; Thermo Fisher
Scientific, Inc.) mixture according to the manufacturer's
instructions. The si-p53 sequences were: Forward 5′-CAC UAC AAC UAC
AUG UGU A-3′, reverse 5′-UAC ACA UGU AGU UGU AGU G-3′. siRNAs were
transfected at a concentration of 20 nM, as previously described
(12); cells were incubated at
37°C in 5% CO2 atmosphere for 48 h and subsequently
harvested for further experimentation.
Western blotting
Cells at 80–90% confluence in 60-mm or 100-mm
culture dishes were lysed in a lysis buffer (1 M Tris-HCl, pH 7.5;
1.5 M NaCl; 100 mM EDTA; 10% glycerol and 1% Triton X-100).
Following resuspension of cells with the lysis buffer, samples were
incubated for 20 min on ice and centrifuged at 16,200 × g at 4°C
for 20 min. The concentration of protein was determined with
Bio-Rad protein assay dye reagent concentrate (Bio-Rad, Inc.,
Hercules, CA, USA), according to the manufacturer's instructions. A
total of 30 µg of protein was loaded per lane and separated by 10%
SDS-PAGE (1.5 M Tris-HCl, pH 8.8; 1 M Tris-HCl, pH 6.8; 30%
acrylamide; 10% SDS; 10% ammonium persulfate and
tetramethylethylenediamine) and transferred onto microporus
polyvinylidene flouoride membranes (EMD Millipore, Billerica, MA,
USA). Membranes were blocked with 5% skim milk in Tris-buffered
saline + 0.05% Tween-20 for 30 min at room temperature and
incubated at 4°C overnight with the following primary antibodies:
Anti-p53 (1:1,000; cat. no. M7001; Dako; Agilent Technologies,
Inc., Santa Clara, CA, USA), anti-OTUD6A (1:1,000; cat. no.
24486-1-AP), anti-USP5 (1:1,000; cat. no. 10473-1-AP) (both from
ProteinTech Group, Inc., Chicago, IL, USA), anti-HA (1:1,000; cat.
no. 11 666 606 001; Roche, Basel, Switzerland) or anti-β-actin
(1:1,000; cat. no. sc-47778; Santa Cruz Biotechnology, Santa Cruz,
CA, USA). The membranes were subsequently incubated with a
horseradish peroxidase-conjugated goat anti-mouse immunoglobulin G
secondary antibody (1:10,000; cat. no. 074-1806; KPL, Inc.,
Gaithersburg, MD, USA) or goat anti-rabbit immunoglobulin G
secondary antibody (1:10,000; catalog no. GTX213110-01; GeneTex,
Inc., Irvine, CA, USA). Protein bands were visualized using the
Enhanced Chemiluminescence Reagent Solution (Young In Frontier,
Seoul, Korea). The densities of protein bands were normalized to
that of β-actin and analyzed by ImageJ (version 1.4.3.67; National
Institutes of Health, Bethesda, MD, USA).
Generation of multiplex PCR and reverse
transcription-quantitative PCR (RT-qPCR) primers
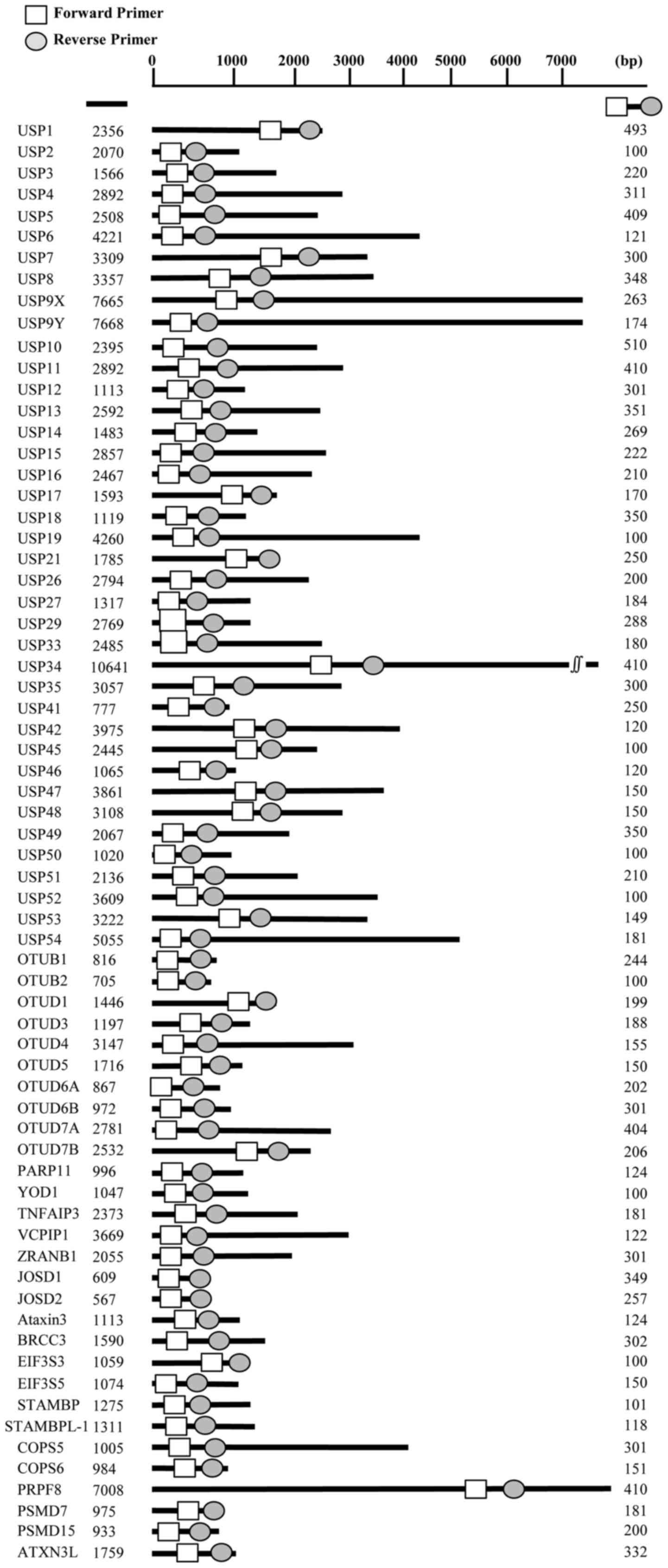
Primers for multiplex PCR were designed to specific
regions of each of 68 DUB genes to amplify sequences ranging
between 100 and 500 bp in length. A total of 10 groups of primer
sets were designed (Fig. 1 and
Table I). For RT-qPCR, the
following gene-specific primers were used: p53, forward,
5′-CTC CTG GCC CCT GTC ATC TTC-3′ and reverse, 5′-AGC GCC TCA CAA
CCT CCG TCA T-3′; USP5, forward, 5′-CGG GAC CAG GCC TTG
AA-3′ and reverse, 5′-TCG TCA ATG TGA CTG AAG ATC CA-3′;
OTUD6A, forward, 5′-TGG ATG ATC CGA AGA GTG AAC'-3′ and
reverse, 5′-TCT TGG AAC TTC TCC AGC TCC T-3′; and GAPDH,
forward, 5′-ATC CCA TCA CCA TCT TCC-3′ and reverse, 5′-CCA TCA CGC
CAC AGT TTC-3′.
 | Table IPrimers used for multiplex polymerase
chain reaction analysis. |
Table I
Primers used for multiplex polymerase
chain reaction analysis.
| Group | DUB gene | Primer
sequence
(5′→3′) | Size
(bp) |
|---|
| G1 | USP5 | F: GTC CAC AAA GAC
GAG TGC GCC T | 409 |
| R: AGG CTG AGT CGG
CCG ACA GTA | |
| USP8 | F: GAC GCC ACC TGC
ATC TAT AGA AG | 348 |
| R: GGA AAG TAA AAC
TGT CCT GCG CAA | |
| USP4 | F: GTA GAA GGC CAG
CAA CCC ATC G | 311 |
| R: ACT AGC ACC TGA
CCC TGG TAT AG | |
| USP9X | F: AGC TTC AAG GGT
TCC AGG ACA AG | 263 |
| R: GAA GAC TAT CTC
GCA ACA CTA TGG | |
| USP51 | F: GGA CCC CAG AGA
CTA GGA AAC G | 210 |
| R: CAT AAT CCT TAC
ACA TGA AGC A | |
| USP27 | F: CTC CAG CTT TAC
GAT CGG TTT AAG | 184 |
| R: CCG AAA CAG CGA
CGA CAT CTC AC | |
| USP47 | F: CAA TGA TCA ACA
TGT CAG CAG GA | 150 |
| R: TTT CTG GCT GGA
TCC TTC AGT CT | |
| USP42 | F: TTA CTC ATC CCA
CCC ATA GCC | 120 |
| R: TCA TGT GAG AGG
GAA GCT GTG GT | |
| USP45 | F: TGG GCT GTT CAG
ATC CAG TAG T | 100 |
| R: ACT GTC AGT CTC
CTT GGT GTA CAG | |
| G2 | USP1 | F: GAC CAA ATG TGT
GAA ATA GGT AAG C | 493 |
| R: GCA AGT AAG GAG
TAG AAG TAG GAG | |
| USP11 | F: TGG TGG AAG GCG
AGG ATT ATG TG | 410 |
| R: GCT GGG CCA AGT
GCC ATC TTT C | |
| USP13 | F: ACC CAG CTG GAC
AAT GGA GTC A | 351 |
| R: CAG CTT GAT GTC
ATT GTC CTG GA | |
| USP12 | F: GAA CTC TGA GTC
TGG TTA CAT CCT | 301 |
| R: GAG GAG CTG GTA
TCT CTG ATT TCA | |
| USP14 | F: TCA GTG TAT TCG
TTC TGT GCC TGA | 269 |
| R: CTC GCA TCA TTT
GTA TCC AAC ATT CA | |
| USP15 | F: AAA CCT CGC TCC
GGA AAG GGG A | 222 |
| R: CAG TTG GCA ACA
GTA TGT AAT CCA A | |
| USP54 | F: CGT GGT AGT GTA
CAA GGG ATG TTT | 181 |
| R: CTC CCA TGC ACT
TGT GAG TTG TAA | |
| USP48 | F: GCT GGT AGA TCG
GGA TAA TTC CA | 150 |
| R: AAC TCA TAG GGC
TCA GCT CCA G | |
| USP46 | F: CCA ATC CTG CTG
ATG TGG CAG TC | 120 |
| R: GCT GAT GGC TGG
AAA GAT GTA GTA | |
| USP52 | F: TCT GGC AAG GTT
TCC CTG AGA GA | 100 |
| R: GGT TGC CAT GCA
CAT CAA AGT CT | |
| G3 | USP10 | F: CCT CCA CAG CCC
GCA GTA TAT TT | 510 |
| R: GAG ATA GGA TCA
TCG CCA CCA TCT | |
| USP34 | F: CAG CCA TAG TGC
TGA AGT TCA AGT | 410 |
| R: GAC TGA CAT CAC
CAG ATT GTG CT | |
| USP18 | F: ATT GGA CAG ACC
TGC CTT A | 350 |
| R: AAG GAT TCC TTC
ACC CGG ATC G | |
| USP21 | F: TGA CAA AGC CGG
AAG TCC TGT A | 250 |
| R: AAA GGG CTT CAC
AGG TGC CAG A | |
| USP3 | F: CCT TGG GTC TGT
TTG ACT TGT TCA | 220 |
| R: CCA GTC CCA GCT
TGG TGT CAT TA | |
| USP16 | F: AAA CTT TAG AAC
CTG TGT GCA G | 210 |
| R: CCT GAG AAT TTC
TGC CAC AGC C | |
| USP33 | F: CCC TTG GTA CTT
GTCA GGA TTG TA | 180 |
| R: AAG CAT AAC ACC
ATA CTC GAA GAG | |
| USP53 | F: GAC ATT TCC AGA
GAA TGT GCT CTG | 149 |
| R: GAT CCA GAT TGG
AAA TGT GAA AGG | |
| USP19 | F: GTT CTT TCC TTC
ATC GTC AGG GTC | 100 |
| R: AGT GGG AGT AGC
CAA GAG ATC ATG | |
| G4 | JOSD1 | F: GTG AAT GTC ATT
ATG GCA GCA C | 349 |
| R: TCC TCC AAC TCT
GAT GAG CCT C | |
| BRCC3 | F: GAG TTC AGA GTA
TGA GAG AAT CG | 302 |
| R: CCT TTT CTT CTT
GTT GTA ATT CCT G | |
| JOSD2 | F: GTG TCT ACT ACA
ACC TGG ACT C | 257 |
| R: ATG AAG TGC TGG
CCT TTC CCA G | |
| EIF3S5 | F: TCT GCC TGG TCC
TGC TCT TCC A | 150 |
| R: TTG TCG ACA GTT
CCC AAC AGG G | |
| Ataxin3 | F: GTC CAA CAG ATG
CAT CGA CCA A | 124 |
| R: CGT CTA ACA TTC
CTG AGC CAT C | |
| STAMBP | F: GAA GCC CTC CTT
AGA TGT GTT | 101 |
| R: TGT CCA CCA CAG
GTG GCT TAG CT | |
| G5 | PRPF8 | F: TCT ATG ACG ACT
GGC TCA AGA C | 410 |
| R: ATC GCC ATG CTT
GTT GAC AGT G | |
| COPS5 | F: GCA GTG GTG ATT
GAT CCA ACA A | 301 |
| R: AGA CCT GAC CAG
TGG TAT AGT C | |
| PSMD14 | F: GGT TTG ACA CTT
CAG GAC TAC A | 200 |
| R: GAG GTC ATA AGT
ACA TCC ACAT G | |
| PSMD7 | F: ACG TCT TCA ACC
TGC TGC CAG A | 181 |
| R: TCC TGC CCT TCT
TTC TTC TCT G | |
| COPS6 | F: AGG TGT TCA AGG
AGC TGG AGT T | 151 |
| R: GGA AGA TCT GTG
TGC TTG GTC A | |
|
STAMBPL1 | F: TTC GAA GAT CAA
CTC AAG AAG CA | 118 |
| R: TCT GGT GTG TGG
AAA AGC AGG A | |
| EIF3S3 | F: GTC CAA ACT CTT
CAA ACC ACC A | 100 |
| R: AGT GAA CTC CTT
GAT GTT CTG G | |
| G6 | ZRANB1 | F: CTA GTG CAA GAC
CAA GGG TG | 301 |
| R: ACA CAT CTT TTA
GCC TTG GCC C | |
| OTUB1 | F: AGG AAC CTC AGC
AGC AGA AGC A | 244 |
| R: GTC TTG CGG ATG
TAC GAG TAC T | |
| OTUD1 | F: ATG GGG CAG ATG
CTG AAT GTG A | 199 |
| R: TGC ACC AGT TGT
CGT ACT CTG | |
| TNFAIP3 | F: CCG AGC TGT TCC
ACT TGT TAA CA | 181 |
| R: CAA CTT TGC GGC
ATT GAT GAG A | |
| OTUD5 | F: ATC GGA GGA GTC
ATG GAT TGA A | 150 |
| R: ACC TGG CGA GCC
TGT TTC TCC T | |
| VCPIP1 | F: GCT CGC TAT GGA
ATG GAC AAA C | 122 |
| R: ACA TGC TCT GGT
TCT ATG AGG | |
| OTUB2 | F: CAT TCT TCG GGA
CCA TCC TGA A | 100 |
| R: GTT CCC ATC CCC
TTT GGT CTT | |
| G7 | OTUD6B | F: AAG AAT GCT GTT
CCC AAG AAT G | 301 |
| R: CCA TAT GTC TGG
CTC CTG TTA A | |
| OTUD7B | F: ACT TCA CAG GGG
TGC CTT GTT | 206 |
| R: GTT CTT CCC TGT
AAC AAC AGG A | |
| OTUD3 | F: GAA GAC GAC CTG
AGA GAT GAA G | 188 |
| R: CTG GGC TCA AGA
TTC TCT TCT G | |
| OTUD4 | F: GCT CTG CTA TGT
GTC AGT CTC T | 155 |
| R: TTA CTT GCA ACT
GTC ATC CTC TG | |
| PARP11 | F: CAG CTA CAA GAT
AGA CTT TGC AG | 124 |
| R: GAT GGC CTC GTT
TTC ACA GAT G | |
| YOD1 | F: ACT TGC CCA TCC
AAT CTG GTG A | 100 |
| R: ACG TAA CTA GAA
GCA CCA CGT T | |
| G8 | USP35 | F: AAG TAC ATG CTC
CTG ACC TTC CA | 410 |
| R: CCC AGG TTG ATG
AGA CCA ATC TT | |
| OTUD7A | F: GCA GCA CTT CTA
CAT GAT CCT A | 405 |
| R: TGT GTA GAT TGG
CAT CTC CAG G | |
| USP26 | F: CAG CCA CCT GTG
AGA CCT GGT AA | 202 |
| R: CTG ATA ACT CTC
CGC AAG TAA G | |
| USP17 | F: GAG CAA CGC AAG
GAG AGC TCA AG | 172 |
| R: AGG GTA CCT TCG
ACT TTT CTG ACG | |
| USP50 | F: CTA TGA TAC CCT
TCC AGT TAA GG | 101 |
| R: TGG CAT TCA CGC
AGC ATG TGT TG | |
| G9 | USP49 | F: AGG ACT ACG TGC
TCA ATG ATA ACC | 402 |
| R: GCA GGA GCA GCC
GTG CAC TCT | |
| ATXN3L | F: TCA GAA GAA AGT
GAT GAG TCT GG | 332 |
| R: CTC TCA ATT GCT
CTC GAA CTT G | |
| USP7 | F: CTC TCA GAC CAT
GGG ATT TCC AC | 300 |
| R: ATT GGT GTG TAG
ATA TGC CCA CAG | |
| USP9Y | F: GAG GCT GTG AGT
GGC TGG AAG T | 174 |
| R: CGG ACG TGT ACC
ATT GTA AGA TAT G | |
| USP2 | F: TAT GGT GCC TAC
ACC CCG TCC T | 103 |
| R: TGA GGA AGC TGC
TGG TGG GGA C | |
| G10 | USP41 | F: GGT TCT GCT TCA
ATG ACT CCA ATA | 250 |
| R: AGC CAT CTC ACG
ATT GAC CGG CT | |
| OTUD6A | F: TGG ATG ATC CGA
AGA GTG AAC | 202 |
| R: TCT TGG AAC TTC
TCC AGC TCC T | |
| USP29 | F: GGG ATG ACT AAG
CTG AAA GAA GCT | 180 |
| R: TTT CAA AGT TAA
ACG CAG GTG ACT | |
| USP6 | F: CGT TGG AAT CAA
CAG CAG CAT TGA | 122 |
| R: CCA TCC ACT TGC
TCG TTC GTG TCA | |
RNA extraction, cDNA synthesis, multiplex
PCR and RT-qPCR
For RNA extraction, cells at 80–90% confluence in
100-mm dishes were lysed in a culture dish with 1 ml of TRIzol
reagent (Invitrogen; Thermo Fisher Scientific, Inc.). cDNA was
synthesized using ReverTra Ace qPCR Master Mix (Toyobo Life
Science, Osaka, Japan) according to the manufacturer's protocol.
GAPDH was used as an internal standard. For multiplex PCR,
2X Multiplex PCR Smart Mix (cat. no. SMP01-M25h; Solgent Co., Ltd.,
Daejeon, Korea) was used and cDNAs were amplified with the
following PCR thermocycling conditions: initial denaturation at
95°C for 15 min, followed by 40 cycles of denaturation at 95°C for
20 sec, annealing at 60°C for 40 sec, extension at 72°C for 1 min,
and final extension at 72°C for 3 min. GAPDH was used as a
control. All PCR products were separated by 3% agarose gel
electrophoresis and the gels were stained with RedSafe DNA Stain
(cat. no. 21141; Chembio, Medford, NY, USA) to visualize the
amplicons to confirm the amplification of specific cDNA bands of
the expected sizes. mRNA expression levels were normalized to
GAPDH and analyzed by ImageJ v1.4.3.6; the expression levels
of DUBs in HCT116 p53+/+ cells were considered as the
standard or baseline level of expression. RT-qPCR was performed
using a StepOne Real-Time PCR System (Thermo Fisher Scientific,
Inc.) according to the manufacture's protocol, and cDNA was
amplified using SYBR-Green PCR Master Mix (cat. no. 4309155;
Applied Biosystems; Thermo Fisher Scientific, Inc.). Relative
expression levels were normalized to GAPDH and compared
using the 2−ΔΔCq method (29).
Statistical analysis
Statistical significance was analyzed by ImageJ
(version 1.4.3.67) and GraphPad Prism version 5 (GraphPad Software,
Inc., La Jolla, CA, USA) from at least three independent
experiments using paired sample t-test. One-way analysis of
variance followed by Tukey's multiple comparisons post hoc test was
performed using GraphPad Prism version 5. P<0.05 was considered
to indicate a statistically significant difference.
Results
DUB screening through multiplex PCR to
identify p53-regulated DUBs
HCT116 p53+/+ and p53−/− cells
were used to examine differential expression levels of DUBs between
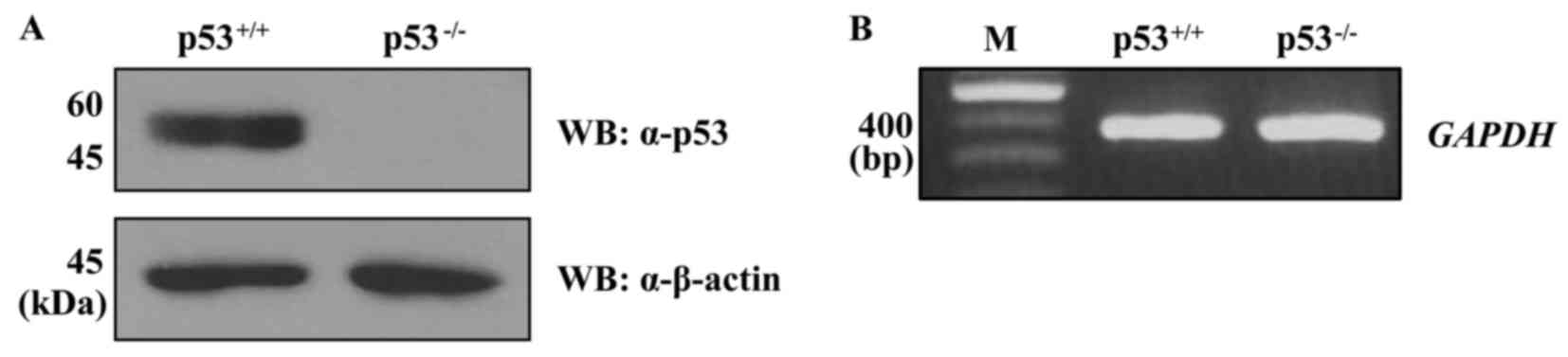
the two cell lines. First, the expression level of p53 protein in
the HCT116 p53+/+ and p53−/− cells was
confirmed (Fig. 2A). Subsequently,
RNA was extracted from each cell line and cDNA was synthesized to
use as templates for multiplex PCR using the DUB
gene-specific DNA primer groups G1-G10. For multiplex PCR, the
expression of GAPDH in the HCT116 p53+/+ and
p53−/− cells was determined at least three independent
times and used to normalize the gene expression data (Fig. 2B).
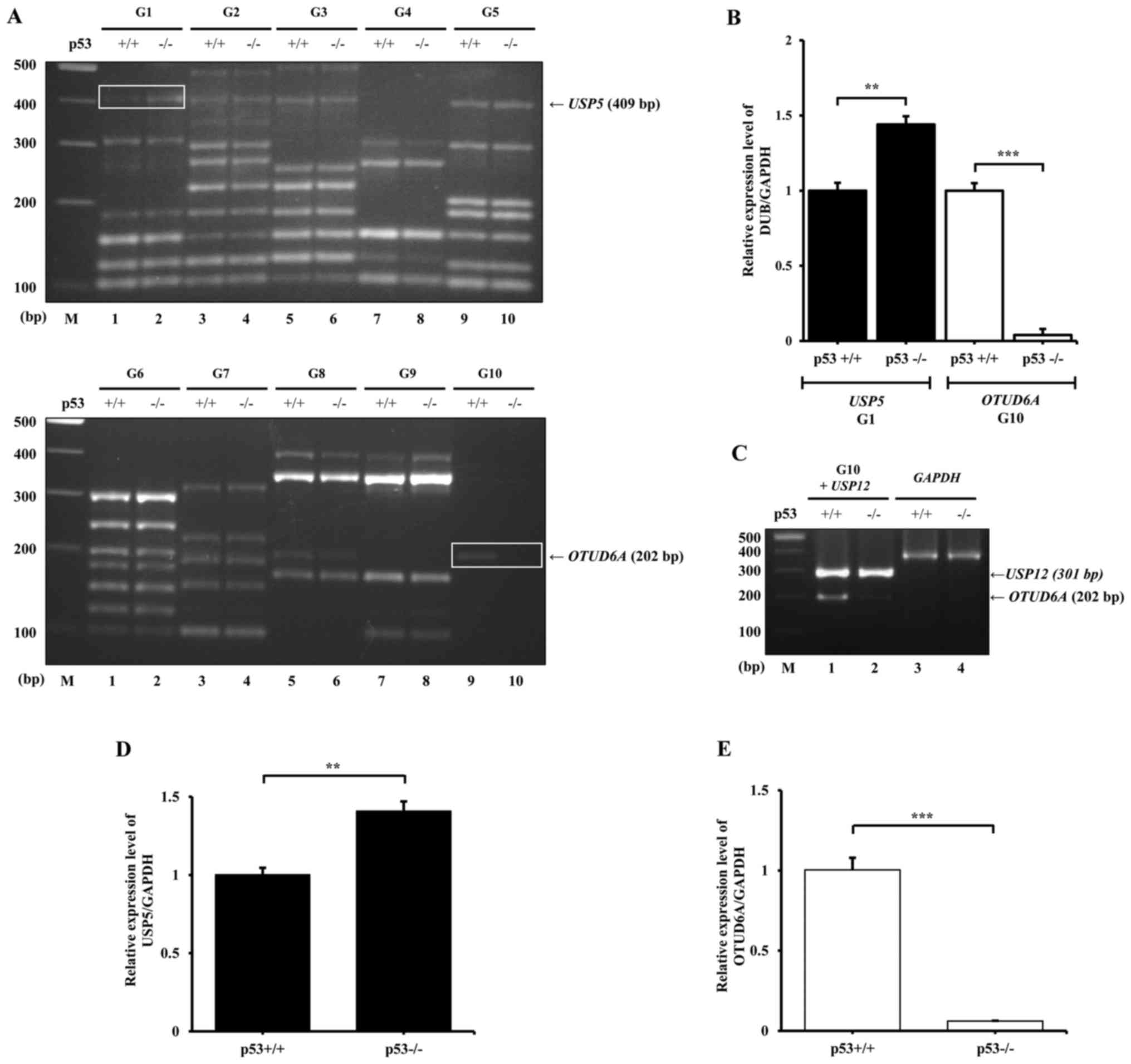
Following multiplex PCR, the PCR products were
analyzed by agarose gel electrophoresis and densitometric analysis
to compare the differential expression (Fig. 3A). The results demonstrated that
USP5 (G1) and OTUD6A (G10) exhibited the most notable
differential expression patterns between HCT116 p53+/+
and p53−/− cells. The results from densitometric
analysis indicated that the expression of USP5 in HCT116
p53−/− cells is a 1.47-fold higher compared with
USP5 expression in p53+/+cells, whereas the
expression of OTUD6A in HCT116 p53+/+ cells was
20-fold higher compared with expression levels in p53−/−
(Fig. 3B). These data were similar
to those reported in a previous study using RNA-sequencing, in
which USP5 was revealed to be highly expressed in HCT116
p53−/− (30). However,
the mRNA expression levels of USP6, USP29 and
USP41 in G10 were not detected. Therefore, multiplex PCR was
repeated using the USP12 primers from G2 as a positive
control spiked into the G10 primer set. Although the expression
level of USP12 was strong, the expression of USP6,
USP29 and USP41 remained undetectable (Fig. 3C). To verify the results from
multiplex PCR, the mRNA expression levels of USP5 and
OTUD6A were investigated by RT-qPCR. Similar to the
multiplex PCR results, the expression level of USP5 in
HCT116 p53−/− was 1.41-fold higher compared with
expression in HCT116 p53+/+ cells, and the expression of
OTUD6A in HCT p53−/− was 16.67-fold lower
compared with that in HCT116 p53+/+ cells (Fig. 3D and E, respectively).
p53 may influence USP5 and OTUD6A at the
protein expression level
Deubiquitination is a crucial PTM process for
regulating protein stability and function (31). Although the mRNA level may not
always correlate with the protein level, protein expression may be
partially predicted by the mRNA level (32). Therefore, the protein expression
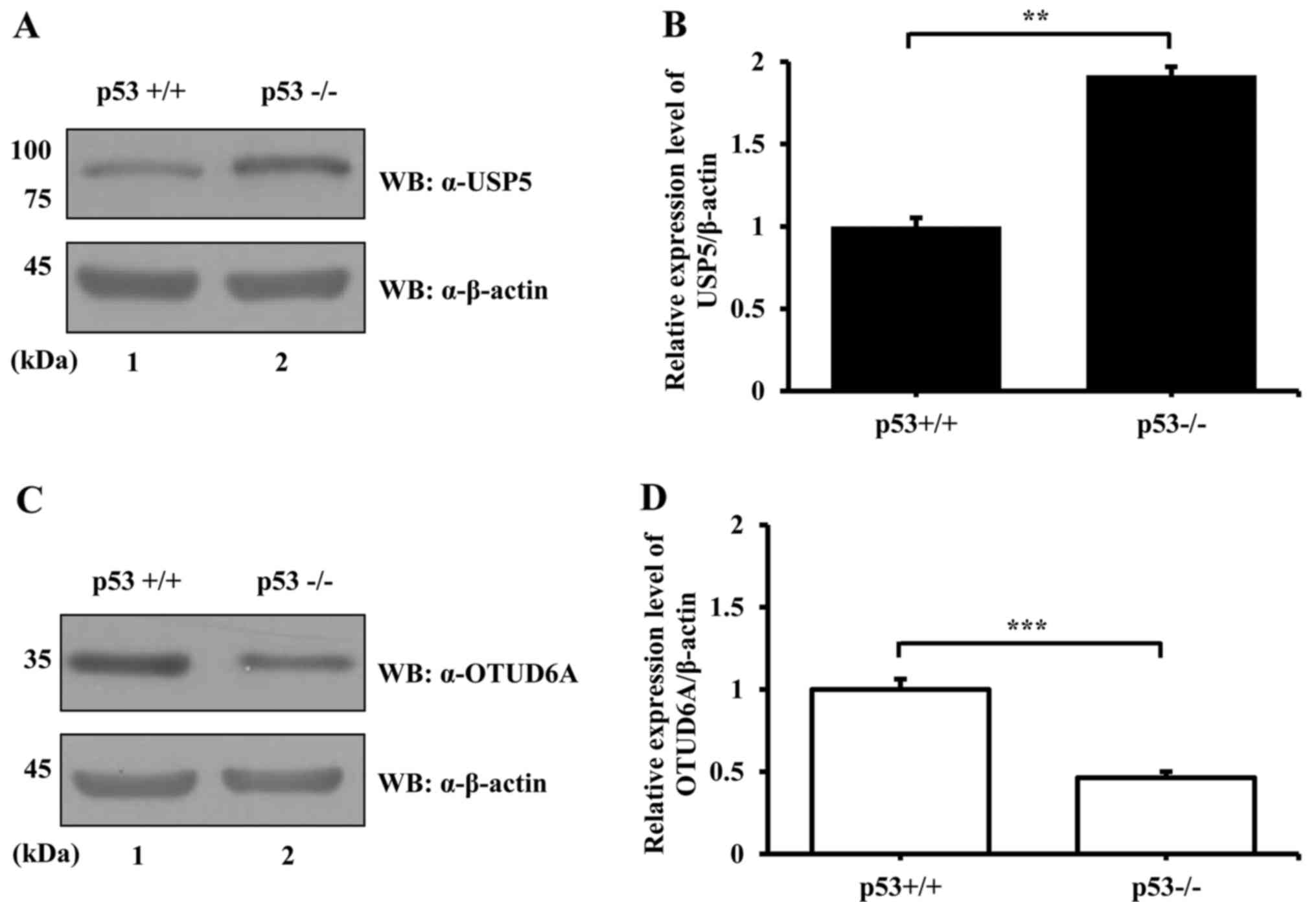
levels of USP5 and OTUD6A were examined by western blotting using
an anti-USP5 or an anti-OTUD6A antibody and lysates from
p53+/+ and p53−/− HCT116 cells. The results
demonstrated that, similar to mRNA expression, the level of USP5
protein expression in HCT116 p53+/+ cells was
significantly lower compared with expression in HCT116
p53−/− cells (Fig. 4A and
B). The protein expression level of OTUD6A in HCT116
p53+/+ was significantly higher compared with expression
in HCT116 p53−/− cells (Fig. 4C and D), which was also similar to
the mRNA expression levels. These results suggested that the
expression of USP5 and OTUD6A may be regulated by p53 at the
protein level.
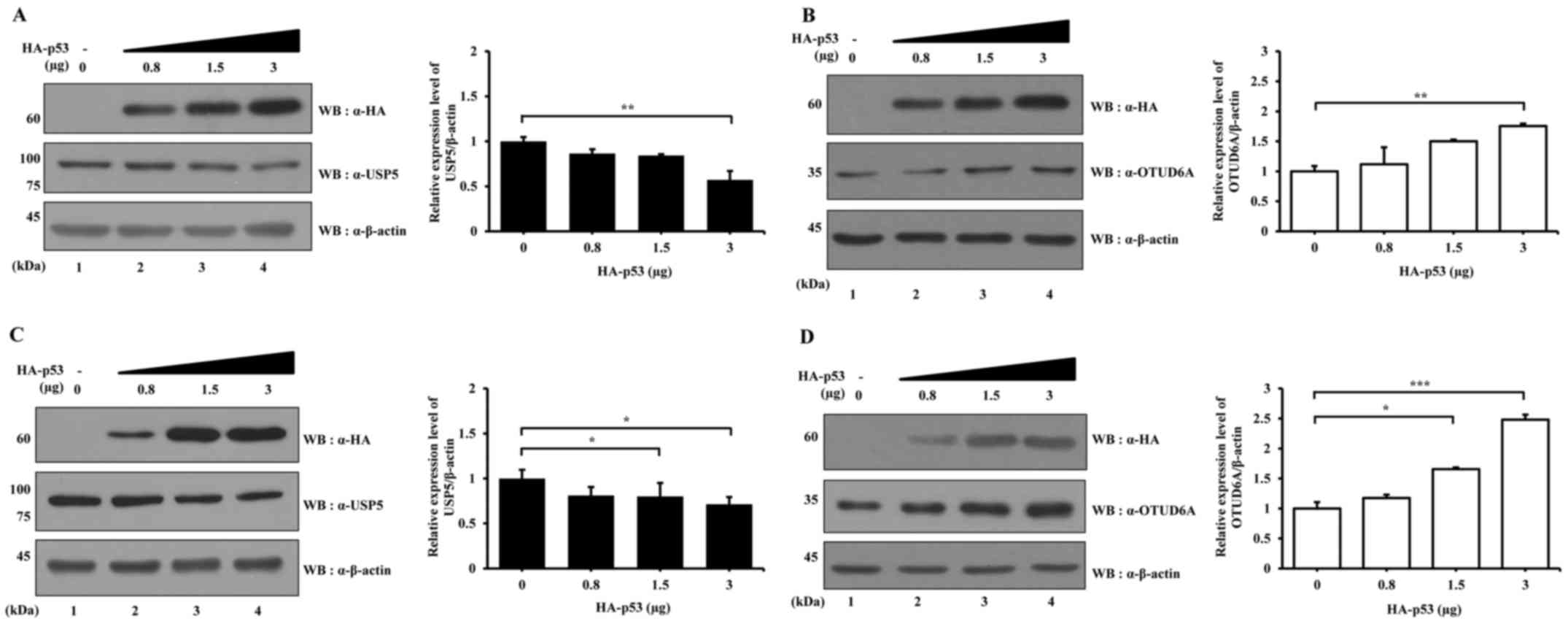
To verify the data, HA-p53 overexpression
vector was transfected into HCT116 p53+/+ cells at
several concentrations and the protein expression levels of USP5
and OTUD6A were examined. The expression level of USP5 decreased
with increasing HA-p53 concentration (Fig. 5A), whereas the expression level of
OTUD6A increased with increasing HA-p53 (Fig. 5B). In addition, the effects of
HA-p53 over-expression on USP5 and OTUD6A expression levels were
examined in HCT116 p53−/− cells. The results
demonstrated that the expression level of USP5 decreased and the
expression level of OTUD6A increased with increasing HA-p53
transfection concentration (Fig. 5C
and D, respectively).
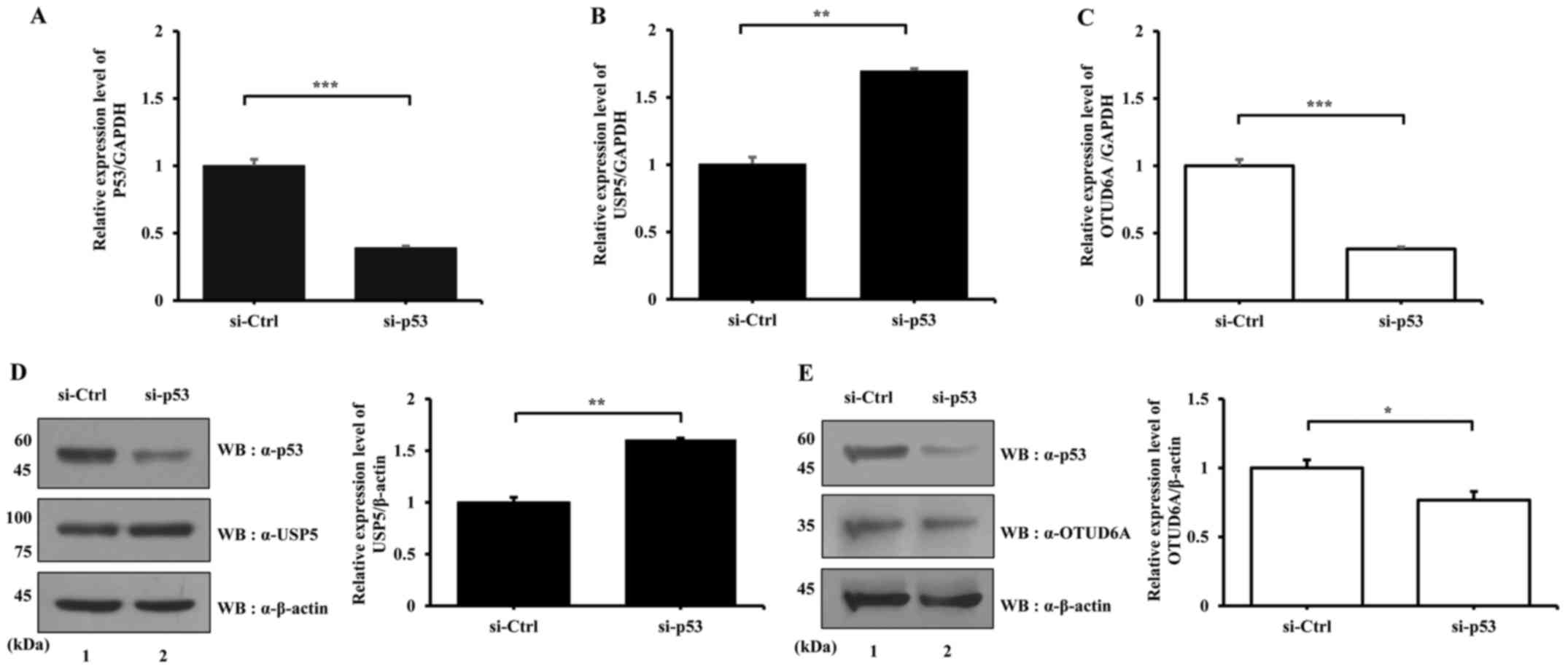
p53 knockdown affects the mRNA and
protein expression levels of USP5 and OTUD6A
The protein expression levels of USP5 and OTUD6A are
affected by overexpression of p53, as aforementioned. Subsequently,
the effects of p53 knockdown on the mRNA and protein expression
levels of USP5 and OTUD6A were determined. p53+/+ HCT116
cells transfected with si-p53 exhibited decreased p53 mRNA
expression (Fig. 6A); USP5
mRNA expression was increased and OTUD6A expression was
decreased following p53 knockdown (Fig. 6B and C, respectively). Similarly,
p53 knockdown resulted in increased USP5 and decreased
OTUD6A protein expression levels (Fig.
6D and E, respectively). Cellular stress such as UV exposure
induces p53 activation that regulates cell cycle, DNA repair, and
apoptosis (33). Therefore, the
transcription levels of USP5 and OTUD6A, which are
affected by the presence of p53 after UV exposure, were examined.
As expected, p53 activation led to the downregulation of
USP5 and upregulation of OTUD6A (data not shown).
Discussion
In eukaryotic cells, most proteins are regulated by
ubiquitination, an enzymatic process controlled by E1, E2 and E3
(34). DUBs reverse ubiquitination
by cleaving the interaction between ubiquitin and substrate
proteins (35). DUBs serve
important roles in a number of cellular processes, including cell
cycle regulation, proteasome-dependent degradation, DNA repair and
homeostasis (17,36,37).
Owing to diverse functions of DUBs in biological processes,
dysfunction of DUBs may result in human diseases, including cancer
(38).
p53 is a well studied tumor suppressor that serves a
crucial role in inducing growth arrest, apoptosis and senescence,
which aid in the prevention of oncogenic progression in stressed
cells (25). In healthy cells, p53
has a short half-life and it is expressed at a low level. Owing to
various stressors, such as DNA damage, oxidative stress and osmotic
shock, activation of p53 is induced (33,39,40).
Subsequently, the half-life of p53 is increased and p53 becomes a
transcription regulator in damaged cells. When the cells receive
low stress, p53 induces cell cycle arrest and DNA repair (41); however, when the cells experience
high stress, p53 induces apoptosis and the cells are not repaired
(41).
It is important to identify DUB-related diseases and
the rapid screening of DUB genes is required for determining
the abnormal expression of DUBs. The present study was the first,
to the best of our knowledge, to develop a method for DUB screening
using multiplex PCR. Multiplex PCR is able to amplify multiple DNA
sequences in a single PCR experiment, offering a convenient and
rapid assay to screen a set of genes simultaneously. The technique
uses DUB gene-specific primers, divided into 10 groups, to
easily amplify unique regions of DUBs ~100–500 bp long. Owing to
the advantages listed below, DUB screening through multiplex PCR
may be used as a disease diagnostic kit (42). There are several advantages of DUB
multiplex PCR. First, this method is able to detect the expression
levels of diverse DUBs at once. Second, it is suggested that
multiplex PCR may be conducted on a number of sample types,
including blood cells, various other types of cells or tissues.
Third, biomarkers may be identified through DUB screening and be
used in pathological research, which may aid in identifying the
disease state of patients and in predicting prognosis following
treatment. Furthermore, the tool may also contribute to other areas
of scientific research. By identifying DUB gene expressions
in specific samples, biological mechanisms and pathophysiology of
certain diseases can be investigated. Conversely, there are several
disadvantages of this tool. The expression levels of DUBs may be
different depending on the cell and tissue type. Expression levels
of some DUBs are too low to identify specific bands in the result
of DUB screening. Moreover, it is not possible to analyze and
compare the expression level of DUBs when they are expressed
abundantly. Although the DUBs are differentially expressed between
the control and experimental groups, the expression level of DUBs
may be seen as similar between these two groups owing to their
abundant expression levels.
In the present study, multiplex PCR was used to
identify DUBs that may be related to p53, and subsequently
investigated the putative effects of the presence or the absence of
p53 on the expression levels of two DUBs. When p53 expression was
downregulated, USP5 expression levels increased and OTUD6A
expression levels decreased. These data suggested that p53
signaling may be involved in the regulation of USP5 and OTUD6A at
the transcriptional and the translational levels. However, there
are no p53 binding sites on USP5 and OTUD6A, which
indicated that p53 does not regulate the transcription of
USP5 and OTUD6A directly (43) and it is suggested that p53 may
regulate the mediators that may serve a role in the expression of
these DUBs.
Mutations of p53 are detected in >50% of human
cancers (25); therefore,
regulating the expression of p53 may be an effective strategy for
treating cancers. The functions of normal p53 are important in
human diseases and its stability and/or functions are modulated by
diverse DUBs (21,44). USP4 deubiquitinates and negatively
modulates ubiquitinated p53 (45).
USP7 serves a key role in the p53 pathway by stabilizing p53 and
mouse double minute 2 homolog (MDM2) (12,46,47).
USP10 deubiquitinates p53 induced by E3 ubiquitin ligase MDM2 and
subsequently affects p53 localization and stabilization (48). USP11 and USP24 have also been
reported to influence DNA damage responses by deubiquitinating p53
protein (49). USP22 was
demonstrated to affect the cell cycle and cell proliferation by
controlling p53 pathway in HeLa cells (50). USP42 binds and regulates the
ubiquitination level of p53 in response to stress signal at the
early phase (51). OTUD5 also
deubiquitinates p53, leading to stabilization of p53 in response to
DNA damage (44).
DUB gene screening through the multiplex PCR
may allow for easy determination of the relationship between
p53 and USP5, as well as p53 and
OTUD6A. Results from the present study indicated that p53
downregulates the expression of USP5 and upregulates the
expression of OTUD6A. Through the present screening results,
USP5 and OTUD6A were indicated to be involved in p53
signaling. Furthermore, this screening will aid in the
identification of abnormal mechanisms of DUBs that may lead to a
number of diseases. In addition, DUBs identified through this
screening may be used as biomarkers, which are indicators of
biological processes and pathogenic processes. Biomarkers indicate
a change in expression or state of a protein, which may be
associated with increased risk or progression of a disease, or may
be used to examine the susceptibility of a disease to a certain
treatment (23,52). Biomarkers have been used for
diagnosing stoke, dementia and certain types of cancer (53–57).
Accordingly, multiplex PCR may aid in identifying abnormal
expression levels of DUBs that may indicate homeostasis disruption,
and also may aid in revealing novel mechanisms. The DUB screening
method developed in the present study may provide a cost-effective
and powerful tool for examining various expression levels of DUBs
associated with diseases or abnormal biological pathways. DUB
screening may facilitate a narrowing down of candidates that may
lead to abnormal mechanisms and cause diverse diseases. Based on
subsequent pathophysiological research, the candidates may be
quantified through qPCR. Taken together, it is suggested that DUB
screening through multiplex PCR with specific primers is
potentially useful.
Acknowledgments
We would like to thank previous and present members
of Baek Laboratory for designing primers of DUB genes and
critical comments on the manuscript.
Notes
[1]
Funding
This study was supported by the National Research
Foundation of Korea grant funded by the Ministry of Science, ICT
and Future Planning (grant no. 2016R1A2B4008635).
[2] Availability
of data and materials
Data sharing is not applicable to this article, as
no data sets were generated or analyzed during the current
study.
[3] Authors'
contributions
SYK and SKK designed the research, performed the
experiments, analyzed the data and wrote the manuscript. SYL
designed the research. KHB designed the research, wrote and edited
the manuscript. All four authors have read and approved for the
manuscript.
[4] Ethics
approval and consent to participate
Not applicable.
[5] Consent for
publication
Not applicable.
[6] Competing
interests
The authors declare that they have no competing
interests.
References
|
1
|
Wang J and Maldonado MA: The
ubiquitin-proteasome system and its role in inflammatory and
autoimmune diseases. Cell Mol Immunol. 3:255–261. 2006.PubMed/NCBI
|
|
2
|
Zhang X, Berger FG, Yang J and Lu X: USP4
inhibits p53 through deubiquitinating and stabilizing ARF-BP1. EMBO
J. 30:2177–2189. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Callis J: The ubiquitination machinery of
the ubiquitin system. Arabidopsis Book. 12:e01742014. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Kulathu Y and Komander D: Atypical
ubiquitylation - the unexplored world of polyubiquitin beyond Lys48
and Lys63 linkages. Nat Rev Mol Cell Biol. 13:508–523. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Husnjak K and Dikic I: Ubiquitin-binding
proteins: Decoders of ubiquitin-mediated cellular functions. Annu
Rev Biochem. 81:291–322. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Woelk T, Sigismund S, Penengo L and Polo
S: The ubiquitination code: A signalling problem. Cell Div.
2:112007. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Tan JM, Wong ES, Kirkpatrick DS,
Pletnikova O, Ko HS, Tay SP, Ho MW, Troncoso J, Gygi SP, Lee MK, et
al: Lysine 63-linked ubiquitination promotes the formation and
autophagic clearance of protein inclusions associated with
neurodegenerative diseases. Hum Mol Genet. 17:431–439. 2008.
View Article : Google Scholar
|
|
8
|
Nathan JA, Kim HT, Ting L, Gygi SP and
Goldberg AL: Why do cellular proteins linked to K63-polyubiquitin
chains not associate with proteasomes? EMBO J. 32:552–565. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Ikeda F and Dikic I: Atypical ubiquitin
chains: new molecular signals. 'Protein Modifications: Beyond the
Usual Suspects' review series. EMBO Rep. 9:536–542. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Iyer LM, Koonin EV and Aravind L: Novel
predicted peptidases with a potential role in the ubiquitin
signaling pathway. Cell Cycle. 3:1440–1450. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Park JJ, Lim KH and Baek KH: Annexin-1
regulated by HAUSP is essential for UV-induced damage response.
Cell Death Dis. 6:e16542015. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Lim KH, Park JJ, Gu BH, Kim JO, Park SG
and Baek KH: HAUSP-nucleolin interaction is regulated by p53-Mdm2
complex in response to DNA damage response. Sci Rep. 5:127932015.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Nijman SM, Luna-Vargas MP, Velds A,
Brummelkamp TR, Dirac AM, Sixma TK and Bernards R: A genomic and
functional inventory of deubiquitinating enzymes. Cell.
123:773–786. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Mevissen TE, Hospenthal MK, Geurink PP,
Elliott PR, Akutsu M, Arnaudo N, Ekkebus R, Kulathu Y, Wauer T, El
Oualid F, et al: OTU deubiquitinases reveal mechanisms of linkage
specificity and enable ubiquitin chain restriction analysis. Cell.
154:169–184. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Sun XX and Dai MS: Deubiquitinating enzyme
regulation of the p53 pathway: A lesson from Otub1. World J Biol
Chem. 5:75–84. 2014.PubMed/NCBI
|
|
16
|
Zhong X and Pittman RN: Ataxin-3 binds
VCP/p97 and regulates retrotranslocation of ERAD substrates. Hum
Mol Genet. 15:2409–2420. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Guterman A and Glickman MH:
Deubiquitinating enzymes are IN/(trinsic to proteasome function).
Curr Protein Pept Sci. 5:201–211. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Cope GA, Suh GS, Aravind L, Schwarz SE,
Zipursky SL, Koonin EV and Deshaies RJ: Role of predicted
metalloprotease motif of Jab1/Csn5 in cleavage of Nedd8 from Cul1.
Science. 298:608–611. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Hu HY: Editorial: Protein ubiquitination
and deubiquitination. Curr Protein Pept Sci. 13:4132012. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Lim KH, Song MH and Baek KH: Decision for
cell fate: Deubiquitinating enzymes in cell cycle checkpoint. Cell
Mol Life Sci. 73:1439–1455. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Kwon SK, Saindane M and Baek KH: p53
stability is regulated by diverse deubiquitinating enzymes. Biochim
Biophys Acta. 1868:404–411. 2017.PubMed/NCBI
|
|
22
|
Park CW and Ryu KY: Cellular ubiquitin
pool dynamics and homeostasis. BMB Rep. 47:475–482. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Henry NL and Hayes DF: Cancer biomarkers.
Mol Oncol. 6:140–146. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Edwards MC and Gibbs RA: Multiplex PCR:
Advantages, development, and applications. PCR Methods Appl.
3:S65–S75. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Wang Z and Sun Y: Targeting p53 for novel
anticancer therapy. Transl Oncol. 3:1–12. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Fridman JS and Lowe SW: Control of
apoptosis by p53. Oncogene. 22:9030–9040. 2003. View Article : Google Scholar
|
|
27
|
Dayal S, Sparks A, Jacob J, Allende-Vega
N, Lane DP and Saville MK: Suppression of the deubiquitinating
enzyme USP5 causes the accumulation of unanchored polyubiquitin and
the activation of p53. J Biol Chem. 284:5030–5041. 2009. View Article : Google Scholar :
|
|
28
|
Potu H, Peterson LF, Pal A, Verhaegen M,
Cao J, Talpaz M and Donato NJ: Usp5 links suppression of p53 and
FAS levels in melanoma to the BRAF pathway. Oncotarget.
5:5559–5569. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
30
|
Marchese FP, Grossi E, Marín-Béjar O,
Bharti SK, Raimondi I, González J, Martínez-Herrera DJ, Athie A,
Amadoz A, Brosh RM Jr, et al: A long noncoding RNA regulates sister
chromatid cohesion. Mol Cell. 63:397–407. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Kessler BM and Edelmann MJ: PTMs in
conversation: Activity and function of deubiquitinating enzymes
regulated via post-translational modifications. Cell Biochem
Biophys. 60:21–38. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Guo Y, Xiao P, Lei S, Deng F, Xiao GG, Liu
Y, Chen X, Li L, Wu S, Chen Y, et al: How is mRNA expression
predictive for protein expression? A correlation study on human
circulating monocytes. Acta Biochim Biophys Sin (Shanghai).
40:426–436. 2008. View Article : Google Scholar
|
|
33
|
Lakin ND and Jackson SP: Regulation of p53
in response to DNA damage. Oncogene. 18:7644–7655. 1999. View Article : Google Scholar
|
|
34
|
Amerik AY and Hochstrasser M: Mechanism
and function of deubiquitinating enzymes. Biochim Biophys Acta.
1695:189–207. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Reyes-Turcu FE, Ventii KH and Wilkinson
KD: Regulation and cellular roles of ubiquitin-specific
deubiquitinating enzymes. Annu Rev Biochem. 78:363–397. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Song L and Rape M: Reverse the curse - the
role of deubiquitination in cell cycle control. Curr Opin Cell
Biol. 20:156–163. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Kennedy RD and D'Andrea AD: The Fanconi
Anemia/BRCA pathway: New faces in the crowd. Genes Dev.
19:2925–2940. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Yang JM: Emerging roles of
deubiquitinating enzymes in human cancer. Acta Pharmacol Sin.
28:1325–1330. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Han ES, Muller FL, Pérez VI, Qi W, Liang
H, Xi L, Fu C, Doyle E, Hickey M, Cornell J, et al: The in vivo
gene expression signature of oxidative stress. Physiol Genomics.
34:112–126. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Kishi H, Nakagawa K, Matsumoto M, Suga M,
Ando M, Taya Y and Yamaizumi M: Osmotic shock induces G1 arrest
through p53 phosphorylation at Ser33 by activated p38MAPK without
phosphorylation at Ser15 and Ser20. J Biol Chem. 276:39115–39122.
2001. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Bieging KT, Mello SS and Attardi LD:
Unravelling mechanisms of p53-mediated tumour suppression. Nat Rev
Cancer. 14:359–370. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Pillet S, Lardeux M, Dina J, Grattard F,
Verhoeven P, Le Goff J, Vabret A and Pozzetto B: Comparative
evaluation of six commercialized multiplex PCR kits for the
diagnosis of respiratory infections. PLoS One. 8:e721742013.
View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Kaplun A, Krull M, Lakshman K, Matys V,
Lewicki B and Hogan JD: Establishing and validating regulatory
regions for variant annotation and expression analysis. BMC
Genomics. 17(Suppl 2): 3932016. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Luo J, Lu Z, Lu X, Chen L, Cao J, Zhang S,
Ling Y and Zhou X: OTUD5 regulates p53 stability by
deubiquitinating p53. PLoS One. 8:e776822013. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Li Z, Hao Q, Luo J, Xiong J, Zhang S, Wang
T, Bai L, Wang W, Chen M, Wang W, et al: USP4 inhibits p53 and
NF-κB through deubiquitinating and stabilizing HDAC2. Oncogene.
35:2902–2912. 2016. View Article : Google Scholar
|
|
46
|
Sheng Y, Saridakis V, Sarkari F, Duan S,
Wu T, Arrowsmith CH and Frappier L: Molecular recognition of p53
and MDM2 by USP7/HAUSP. Nat Struct Mol Biol. 13:285–291. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Liu X, Yang X, Li Y, Zhao S, Li C, Ma P
and Mao B: Trip12 is an E3 ubiquitin ligase for USP7/HAUSP involved
in the DNA damage response. FEBS Lett. 590:4213–4222. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Yuan J, Luo K, Zhang L, Cheville JC and
Lou Z: USP10 regulates p53 localization and stability by
deubiquitinating p53. Cell. 140:384–396. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Zhang L, Nemzow L, Chen H, Lubin A, Rong
X, Sun Z, Harris TK and Gong F: The deubiquitinating enzyme USP24
is a regulator of the UV damage response. Cell Reports. 10:140–147.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Liu YL, Zheng J, Tang LJ, Han W, Wang JM,
Liu DW and Tian QB: The deubiquitinating enzyme activity of USP22
is necessary for regulating HeLa cell growth. Gene. 572:49–56.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Hock AK, Vigneron AM, Carter S, Ludwig RL
and Vousden KH: Regulation of p53 stability and function by the
deubiquitinating enzyme USP42. EMBO J. 30:4921–4930. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Pirrone V, Mell J, Janto B and Wigdahl B:
Biomarkers of HIV Susceptibility and Disease Progression.
EBioMedicine. 1:99–100. 2014. View Article : Google Scholar
|
|
53
|
Kim K and Lee JH: Risk factors and
biomarkers of ischemic stroke in cancer patients. J Stroke.
16:91–96. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Al-Qazzaz NK, Ali SH, Ahmad SA, Chellappan
K, Islam MS and Escudero J: Role of EEG as biomarker in the early
detection and classification of dementia. Sci World J.
2014:9060382014. View Article : Google Scholar
|
|
55
|
Goossens N, Nakagawa S, Sun X and Hoshida
Y: Cancer biomarker discovery and validation. Transl Cancer Res.
4:256–269. 2015.PubMed/NCBI
|
|
56
|
Haynes HR, Camelo-Piragua S and Kurian KM:
Prognostic and predictive biomarkers in adult and pediatric
gliomas: Toward personalized treatment. Front Oncol. 4:472014.
View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Nalejska E, Mączyńska E and Lewandowska
MA: Prognostic and predictive biomarkers: Tools in personalized
oncology. Mol Diagn Ther. 18:273–284. 2014. View Article : Google Scholar : PubMed/NCBI
|