Introduction
Autophagy is the dynamic process in eukaryotes by
which dysfunctional cytoplasmic components including damaged
proteins and organelles are degraded in order to maintain neuron
homeostasis (1,2). It has been implicated in a number of
physiological processes that are important for human health and
disease. The central factors for regulating autophagy are adenosine
monophosphate (AMP) kinase (AMPK) and mammalian target of rapamycin
(mTOR). Nutrient depletion is one of the most effective
physiological inducers of autophagy.
Limited energy in the form of adenosine triphosphate
is able to active AMPK and drive autophagy. The kinase TOR (mTOR in
mammals) exerts a negative effect on autophagy; the inhibition of
mTOR activity by phosphorylating unc-51 like autophagy activating
kinase 1/2 (ULK1/2), as well as some autophagy-related (Atg)
proteins, promotes autophagy (3).
Autophagy serves an important role in cancer therapeutics. In
precancerous lesions, a previous study indicated that autophagy
enhancers may prevent the development of cancer (4). In advanced cancer, the inhibition and
enhancement of autophagy have been suggested as potential treatment
strategies (5–7).
In addition to intracellular signaling mechanisms,
an increasing body of evidence has indicated that microRNAs
(miRNAs/miRs) may serve a crucial role in the regulation of
autophagy (8–10). miRNAs are a family of small
non-coding RNA, with a length of 18–22 nucleotides, that regulate
gene expression by either triggering target mRNA degradation or the
inhibition of mRNA translation (11,12).
Multiple miRNAs has been shown to be able to regulate autophagy at
various steps. For example, during the early stages of autophagy
induction, miR-885-3p can directly modulate ULK2 expression upon
cisplatin exposure (13). Beclin
1, a critical autophagy-promoting gene, has been shown to serve a
key role in the modulation of cell survival and death in different
types of cells. Zhu et al (14) demonstrated that miR-30a regulates
autophagy by inhibiting Beclin 1 expression. In addition, miR-375
directly suppressed autophagy by inhibiting the expression of ATG7
and impaired the viability of hepatocellular carcinoma cells upon
hypoxic exposure (15). miR-376b
has been observed to inhibit starvation- and rapamycin-induced
autophagy in breast cancer cells by targeting the key autophagy
proteins, Beclin 1 and Atg4C (16). Furthermore, miR-181A blocked
starvation-and mTOR inhibition-associated autophagy by targeting
the critical autophagy protein ATG5 (17). miR-638 has also been reported to
directly target the tumor suppressor disheveled binding antagonist
of β-catenin 3 and promote autophagy, as well as malignant
phenotypes in cancer cells (18).
In addition, some other miRNAs including miR-23a (19), miR-130a (20) and miR-124 (21) are also involved in the regulation
of autophagy.
In recent years, an increasing body of evidence has
indicated that miR-92b is dysregulated in various types of cancers
(22). miR-92b promoted tumor
migration, invasion and proliferation in osteosarcoma by targeting
reversion inducing cysteine rich protein with Kazal motifs
(23). It has also been reported
to promote epithelial-mesenchymal transition in bladder cancer by
targeting disabled homolog 2-interacting protein (24). These studies revealed that miR-92b
may serve an oncogenic role in cancer. However, Zhao et al
(25) reported that miR-92b
targets mothers against decapentaplegic homolog 3 to inhibit the
migration and invasion of nasopharyngeal cancer cells. These
results demonstrated that one specific miRNA may serve various
roles in distinct genetic contexts. To date, the potential function
of miR-92b in breast cancer is still largely unknown.
In the present study, it was reported hsa-miR-92b
promote starvation- and mTOR inhibition-induced autophagy, and
inhibited the viability and invasion of breast cancer cells.
Mechanistic studies revealed that miR-92b directly targeted the
histone methyltransferase enhancer of zeste homolog 2 (EZH2), a
crucial catalyzed subunit of Polycomb repressive complex 2 (PRC2),
that regulates autophagy, proliferation and invasion in cancer
cells. In addition, miR-92b promoted autophagy, and inhibited the
viability and invasion of breast cancer cells by targeting EZH2.
The results of the present study provided a novel strategy using
miR-92b to regulate the progression of breast cancer.
Materials and methods
Plasmid constructs
miR-92b mimics, anti-miR-92b (anti-92b) and negative
control (NC) RNA (miR-NC) were purchased from Shanghai GenePharma
Co., Ltd. (Shanghai, China). Green fluorescent protein (GFP)-light
chain 3 (LC3), EZH2 short hairpin (sh)-RNA Plasmid, pCMV6
Entry-EZH2-Myc-DDK expression plasmid and control plasmid were
purchased from OriGene Technologies, Inc. (Rockville, MD, USA). For
the dual luciferase assay, the wild-type (wt) or mutant (mut) forms
of the EZH2 3′-untranslated region (UTR) were cloned into the
pGL3-promoter vector (Promega Corporation, Madison, WI, USA) using
the following primers: EZH2 wt primers, 5′-GAATCTCGAGCATCTGCTAC-3′
and 5′-CCGCTCGAGACAAGTTCAA-3′; EZH2 mut primers,
5′-ATGCAGTATGGTACATTTTTC-3′ and 5′-AAAATTCACTGGTACAAAAC-3′.
Cell culture
MCF-7 and MDA-MB-453 human breast carcinoma cells
were cultured in Dulbecco's modified Eagle's medium (DMEM; Gibco;
Thermo Fisher Scientific, Inc., Waltham, MA, USA) supplemented with
10% fetal bovine serum (FBS; Gibco; Thermo Fisher Scientific, Inc.)
and penicillin/streptomycin (Gibco; Thermo Fisher Scientific, Inc.)
in a 5% CO2-humidified incubator at 37°C. Starvation in
Earle's balanced salt solution (EBSS; Biological Industries,
Kibbutz Beit Haemek, Israel) at 37°C for 4 h or rapamycin treatment
(2.5 µM, at 37°C for 8 h; BioVision, Inc., Milpitax, CA,
USA) were used to induce autophagy.
Cell transfection
For miRNA transfection, MCF-7 and MDA-MB-453 cells
were transfected with HiPerFect Transfection Reagent (Qiagen GmbH,
Hilden, Germany). According to the manufacturer's instructions, 50
nM of miR-92b mimics, anti-92b or miR-NC were added to each well of
a 6-well plate. The sequences were as follows: miR-92b mimics,
5′-UAUUGCACUCGUCCCGGCCUCC-3′ and 5′-AGGCCGGGACGAGUGCAAUAUU-3′;
anti-92b, 5′-GGA GGCCGGGACGAGUGCAAUA-3′; and miR-NC,
5′-CAGUACUUUUGUGUAGUACAA-3′. For DNA transfection, 3 µg
plasmid DNA (GFP-LC3, EZH2 shRNA Plasmid, pCMV6 Entry-EZH2-Myc-DDK
expression plasmid and control plasmid) was transfected with the
FuGENE-HD transfection reagent (Promega Corporation) in each well
of a 6-well plate. All subsequent cell experiments were performed
at 24 h post-transfection.
GFP-LC3 analyses
For GFP-LC3 analysis, 48 h following GFP-LC3 and
miRNA co-transfection, 2×106 cells were starved for 4 h
in EBSS medium or treated with rapamycin (2.5 µM) for 8 h,
fixed for 20 min in 4% formaldehyde at 37°C, washed with PBS and
then observed using a BX3-CBH fluorescence microscope (Olympus
Corporation, Tokyo, Japan). Cells containing >10 GFP-LC3 green
fluorescing dots were classed as positive. At least 150 GFP
positive cells per condition were counted and the graphs were
plotted as a percentage of GFP-LC3 positive cells over the total
number of transfected cells.
Target gene prediction for miRNA
miRNA targets were identified using the
bioinformatics software miRanda (www.microrna.org/microrna/home.do) and TargetScan
Human version 7.1 (www.targetscan.org/vert_71/).
Western blot analysis
Total cellular proteins were harvested from cancer
cell lines. Cancer cells were transfected with 50 nM miR-92b,
anti-92b, shEZH2 or miR-NC. Cell lysates were washed in ice-cold
PBS and lysed in the radioimmunoprecipitation assay buffer (0.1%
SDS, 1% Triton, 0.5% deoxycholate, 0.5% NP40, 50 mM Tris-HCl pH
7.5, 150 mM NaCl, 50 mg/ml phenylmethylsulfonyl fluoride, 1 mg/ml
leupeptin, 1 mg/ml aprotinin, 1 mg/ml pepstatin, 50 mg/ml
Na3VO4 and 50 mg/ml NaF) with the complete
mini protease inhibitor cocktail and PhosSTOP phosphatase inhibitor
cocktail (Roche Applied Science, Penzberg, Germany). The protein
concentration of cell lysates was determined by using the Pierce
BCA Protein Assay kit (Thermo Fisher Scientific, Inc.) according to
the manufacturer's instructions; 5% bovine serum albumin (BSA;
Sigma-Aldrich; Merck KGaA, Darmstadt, Germany) was used as a
standard. Equal amounts (30 µg) of cell lysates were
separated by 10% SDS-PAGE and transferred onto a polyvinylidene
difluoride membrane (Bio-Rad Laboratories, Inc., Hercules, CA,
USA). The membranes were blocked with TBS-T (50 mM Tris-HCI, pH
8.0, 100 mM NaCl and 0.1% Tween-20) containing 5% non-fat dried
milk or 5% BSA (Sigma-Aldrich; Merck KGaA) for 1 h at room
temperature, and then probed with the following primary antibodies
overnight at 4°C: Rabbit-anti-human EZH2 antibodies (dilution,
1:1,000; cat. no. ab186006; Abcam, Cambridge, UK),
rabbit-anti-human LC3B antibodies (dilution, 1:1,000; cat. no.
ab192890; Abcam), rabbit-anti-human sequestosome 1 (SQSTM1)/p62
antibodies (dilution, 1:1,000; cat. no. GTX111393, GeneTex, Inc.,
Irvine, CA, USA) and rabbit-anti-human β-actin (dilution, 1:1,000;
cat. no. 20536-1-AP; Wuhan Sanying Biotechnology, Wuhan, China).
Following washing with TBS-Tween-20 for 15 min, the membranes were
incubated with horseradish peroxidase-conjugated secondary
anti-rabbit antibodies (dilution, 1:1,000; cat. no. 7074s; Cell
Signaling Technology, Inc., Danvers, MA, USA) for 1 h at room
temperature. Finally, the immunoreactive bands were developed with
Pierce Electro-Chemiluminescence Western blotting substrate (Thermo
Fisher Scientific, Inc.) on Blue Basic autoradiography film
(BioExpress Corporation, Kaysville, UT, USA). ImageJ software
(National Institutes of Health, Bethesda, MD, USA) was employed to
quantify protein levels.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
Extraction of total RNA, including miRNAs, from all
culture cells was performed using the TRIzol reagent (Invitrogen;
Thermo Fisher Scientific, Inc.) according to the manufacturer's
instructions. Total RNA was then reverse transcribed into cDNA
using the RevertAid First Strand cDNA Synthesis kit (Promega
Corporation) with the following temperature protocol: 25°C for 5
min, 42°C for 60 min and 70°C for 15 min. Human miRNAs were
detected with their specific primers (Guangzhou RiboBio, Co., Ltd.,
Guangzhou, China). qPCR was performed using the SYBR Green
Real-Time PCR Master Mix (Promega Corporation). The following
primers were used for qPCR: hsa-miR-92b-3p forward,
5′-TATTGCACTCGTCCCGGCCTCC-3′ and reverse, 5′-CAGTGCGTGTCGTGGAGT-3′;
U6 forward, 5′-CTCGCTTCGGCAGCACA-3′ and reverse,
5′-AACGCTTCACGAATTTGCGT-3′; EZH2 forward,
5′-CGCTTTTCTGTAGGCGATGT-3′ and reverse, 5′-TGGGTGTTGCATGAAAAGAA-3′;
GAPDH forward, 5′-CGCTGAGTACGTCGTGGAGTC-3′ and reverse,
5′-GCTGATGATCTTGAGGCTGTTGTC-3′. The 2−∆∆Cq method
(26) was used to evaluate the
relative expression levels of the indicated genes.
Dual luciferase-reporter assay
pGL3-promoter vectors containing luc-EZH2-wt or
luc-EZH2-mut were cotrans-fected using FuGENE-HD transfection
reagent (Promega Corporation) in 6-well plates with miR-92b and
Renilla luciferase into COS7 cells (The Cell Bank of Type
Culture Collection of Chinese Academy of Sciences, Shanghai,
China). Firefly and Renilla luciferase activities were
measured 48 h following transfection using the dual-luciferase
reporter assay system (Promega Corporation). The results of firefly
luciferase activity were normalized to Renilla luciferase
activity.
Transwell invasion and migration
assays
Cell migration and invasion were investigated by
Transwell assays. MCF7 and MDA-MB-453 cells were prepared following
transfection with miR-NC, miR-92b or anti-92b for 24 h,
respectively, and the MCF7 cells were also prepared following
transfection with vector, shEZH2, miR-NC, miR-92b, or
co-transfection with miR-92b and EZH2 vector. Cells
(1×105) were suspended in 200 µl DMEM without
FBS, were seeded in the upper chambers (Corning Incorporated,
Corning, NY, USA). A total of 600 µl DMEM with 10% FBS was
added to the lower chambers. The chambers were then cultured at
37°C with 5% CO2 for 24 h. For invasion assays,
Transwell chambers coated with Matrigel (BD Biosciences, Franklin
Lakes, NJ, USA) were used. Cells on the upper chamber were then
carefully removed, while the cells on the lower chamber were
stained with crystal violet at 37°C for 10 min. Cells were
subsequently counted using a light microscope (magnification, ×200)
and five random fields of view were selected to calculate cell
numbers.
Cell viability analyses
Cells were transfected with miRNAs (50 nM miR-92b
mimics, anti-92b or miR-NC), and following 24 h post-transfection,
cells were seeded into a 96-well plate (3×103/well).
Cell viability was measured using a Cell Counting Kit-8 (CCK8;
Promega Corporation) at 24, 48 or 72 h following cell seeding. The
viability of cancer cells was assessed by measuring the absorbance
at 450 nm to obtain the optical density values.
Colony formation assay
A total of 3,000 cells were seeded per well in a
6-well cell culture plate, which were then transfected with 50 nM
miR-92b mimics, anti-92b or miR-NC, respectively, using HiPerFect
Transfection Reagent. MDA-MB-453 cells were seeded in fresh 6-well
plates and maintained in DMEM containing 10% FBS for 10 days. MCF7
cells were seeded on 6-well plates with 10% FBS + DMEM for 2 weeks.
Colonies were fixed with methanol at room temperature for 15 min
and stained with crystal violet dye (0.1% w/v) for 15 min at room
temperature.
Patients and tissue specimens
A total of 30 breast cancer tissues were collected
from patients that underwent surgery between January 2015 and
October 2016 at The Fourth Clinical Hospital of Hebei Medical
University (Hebei, China). All of these patients were female, with
a median age of 48 years, and did not receive preoperative adjuvant
chemotherapy and radiotherapy. All tissues were placed in liquid
nitrogen and stored at −80°C following resection. The breast cancer
tissues were then used to evaluate the expression of miR-92b and
EZH2 via RT-qPCR, as aforementioned. The present study was approved
by the Institutional Review Boards of The Fourth Clinical Hospital
of Hebei Medical University and written informed consent was
obtained from all patients.
Statistical analysis
A two-tailed Student's t-test was used to determine
the differences between 2 groups. The associations among two or
more variables was analyzed by one-way analysis of variance with
Bonferroni's post hoc test. The correlation between miR-92b and
EZH2 mRNA expression was examined by Spearman's correlation.
P<0.05 was considered to indicate a statistically significant
difference. All analyses were performed with SPSS software (version
16.0; SPSS, Inc., Chicago, IL, USA) or GraphPad Prism (version 5;
GraphPad Software, Inc., La Jolla, CA, USA).
Results
miR-92b promotes starvation- or
rapamycin-induced autophagy in MCF-7 cells
In order to detect whether miR-92b levels were
enhanced following stress-inducing stimuli, the present study
performed an RT-qPCR analysis. The results indicated that the
miR-92b levels increased in MCF-7 cells that were starved for 4 h
(Fig. 1A). In agreement with the
starvation-associated results, treatment with rapamycin also
elevated miR-92b expression in MCF-7 cells (Fig. 1B).
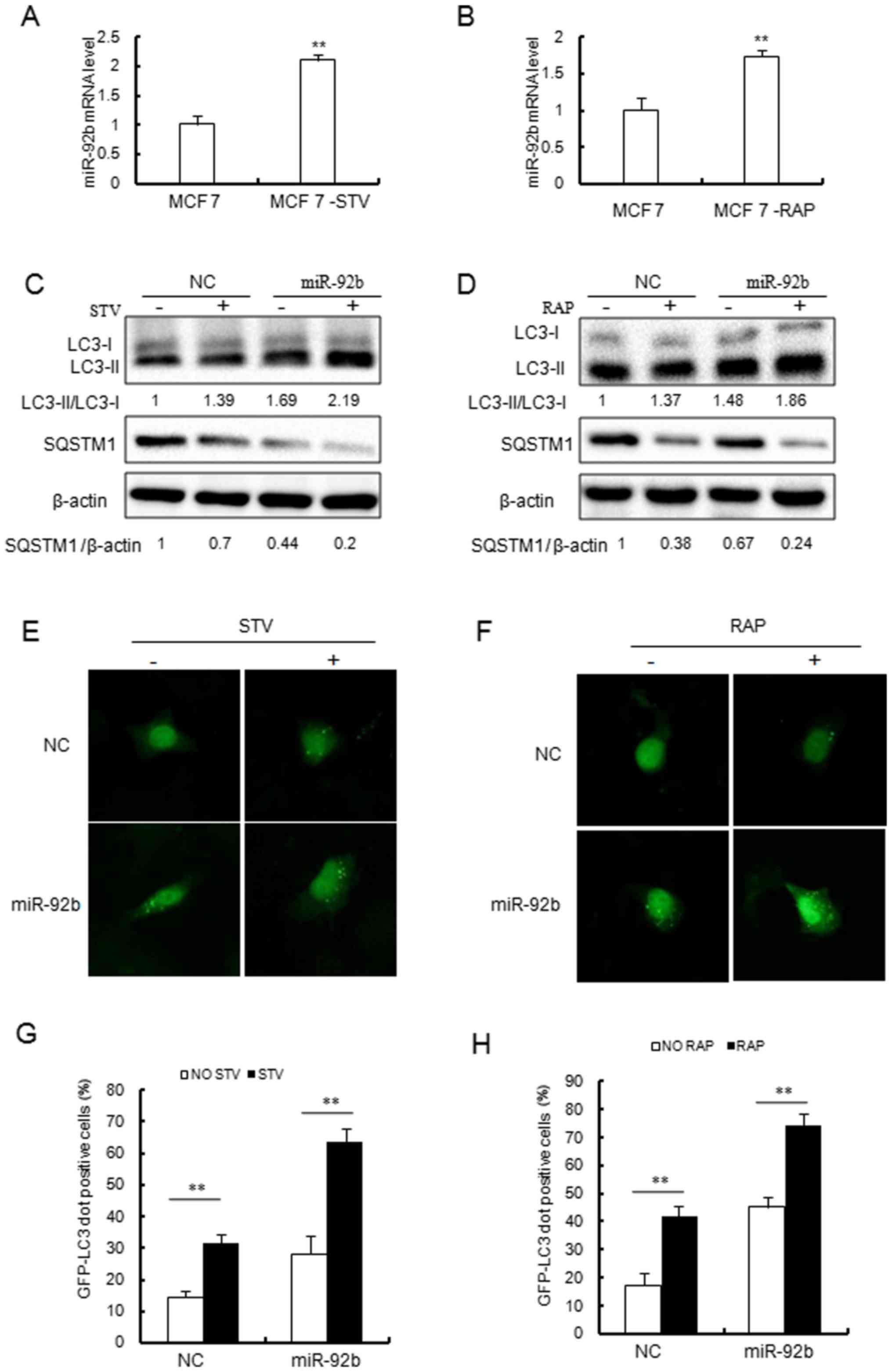 | Figure 1Overexpression of miR-92b increases
autophagic activity in MCF-7 cells. (A and B) Reverse
transcription-quantitative polymerase chain reaction analysis of
miR-92b expression in MCF-7 cells treated with (A) STV (4 h) or (B)
RAP (2.5 µM, 8 h). miR-92b expression levels were
significantly increased following STV or RAP treatment.
**P<0.01 vs. MCF 7 cells. (C and D) Immunoblotting
results indicated that miR-92b increased (C) STV and (D) RAP
induced conversion of LC3-I to LC3-II and SQSTM1 degradation in
MCF-7 cells. (E and F) miR-92b promoted STV and RAP induced GFP-LC3
dot formation in MCF-7 cells. Detection of cytoplasmic puncta
formation by soluble GFP-fused LC3 protein is a commonly used
method to follow autophagy activation using microscopy
(magnification, ×40). (G and H) Quantitative analysis of the
experiments in (E and F). **P<0.01, as indicated.
STV, starvation; RAP, rapamycin; miR, microRNA; LC3, light chain 3;
GFP, green fluorescent protein; SQSTM1, sequestosome 1; NC,
negative control. |
Further study revealed that miR-92b enhanced
autophagy following starvation in MCF-7 cells (Fig. 1C–H). As shown in Fig. 1C, overexpression of miR-92b
increased the transformation of free LC3-I to the autophagy
membrane associated LC3-II, which was induced by starvation. In
addition, miR-92b was markedly associated with starvation-induced
SQSTM1 degradation (Fig. 1C).
Overexpression of miR-92b also increased the accumulation of
starvation-induced GFP-LC3 dots (Fig.
1E and G). These results indicated that miR-92b may serve a
role in regulating starvation-induced autophagy.
Similarly, the present study evaluated whether
miR-92b overexpression also affects autophagy induced by
mTOR-inhibitor rapamycin. Consistent with the results of
starvation, rapamycin induced LC3 lipidation (Fig. 1D), SQSTM1 degradation (Fig. 1D) and GFP-LC3 dot formation
(Fig. 1F and H). Therefore, in
addition to its effect on autophagy induced by starvation, miR-92b
also promoted the autophagic activity induced by rapamycin in MCF-7
cells.
miR-92b promotes starvation- or
rapamycin-induced autophagy in MDA-MB-453 cells
In order to confirm the effect of miR-92b on
autophagy in breast cancer cells, the present study selected
MDA-MB-453 cells, which have the same low expression of miR-92b as
MCF7 cells. Starvation and treatment with the mTOR-inhibitor
rapamycin increased endogenous miR-92b expression (Fig. 2A and B). Similarly, the
overexpression of miR-92b markedly increased the lipidation of LC3
and SQSTM1 degradation (Fig. 2C and
D) in MDA-MB-453 cells. It was further observed that
autophago-some accumulation was enhanced in MDA-MB-453 cells
overexpressing miR-92b (Fig.
2E–H).
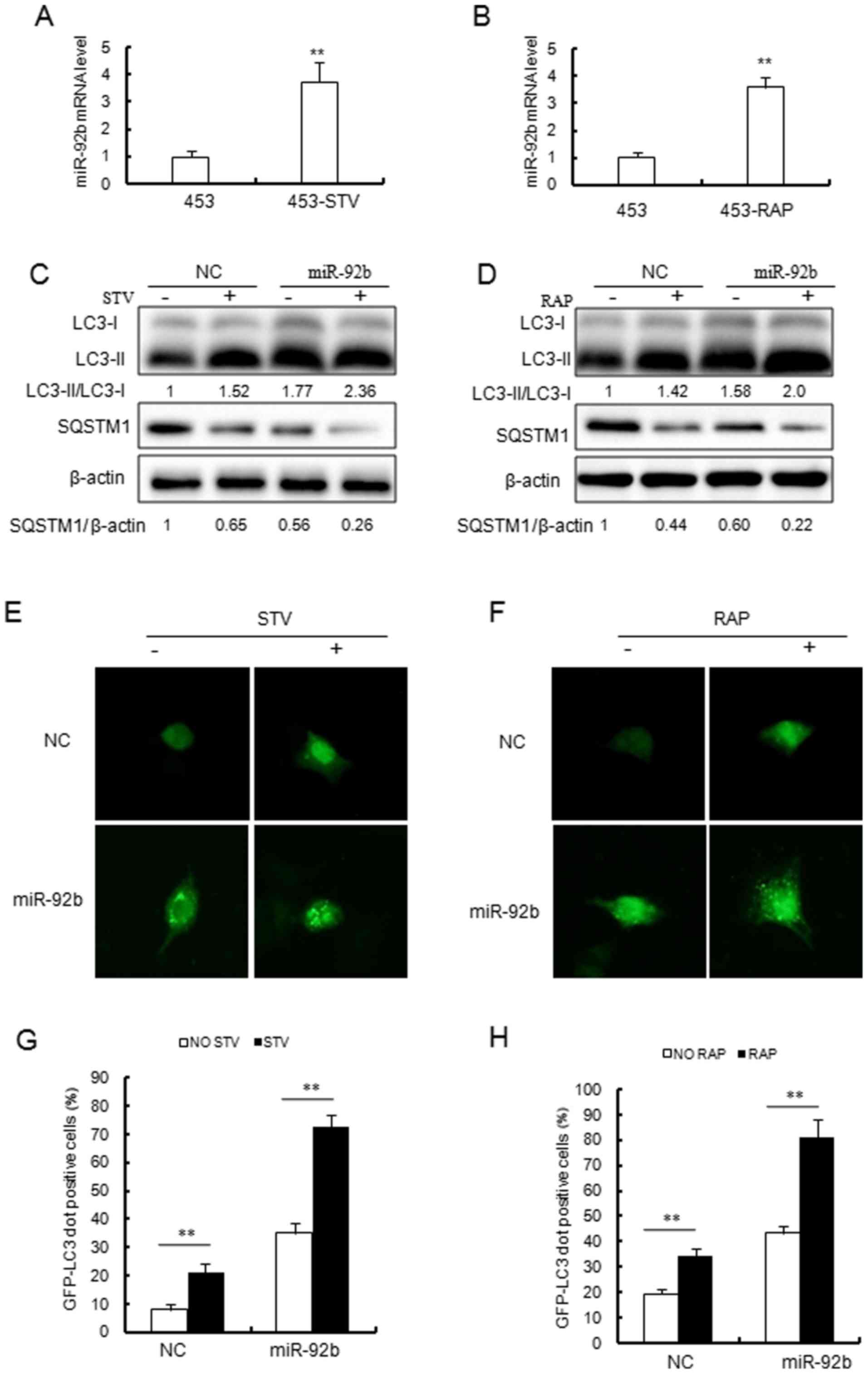 | Figure 2Overexpression of miR-92b results in
increased autophagic flux in MDA-MB-453 cells. (A) STV (4 h) and
(B) RAP (2.5 µM, 8 h) induced miR-92b expression in
MDA-MB-453 cells. miR-92b expression was detected by reverse
transcription-quantitative polymerase chain reaction.
**P<0.01 vs. 453 cells. (C and D) Immunoblotting
results indicated that miR-92b increased (C) STV and (D) RAP
induced conversion of LC3-I to LC3-II and SQSTM1 degradation in
MDA-MB-453 cells. (E and F) miR-92b promoted (E) STV and (F) RAP
induced GFP-LC3 dot formation in MDA-MB-453 cells (magnification,
×40). (G and H) Quantitative analysis of the experiments in (E and
F). **P<0.01, as indicated. STV, starvation; RAP,
rapamycin; miR, microRNA; LC3, light chain 3; GFP, green
fluorescent protein; SQSTM1, sequestosome 1; NC, negative control;
453, MDA-MB-453 cells. |
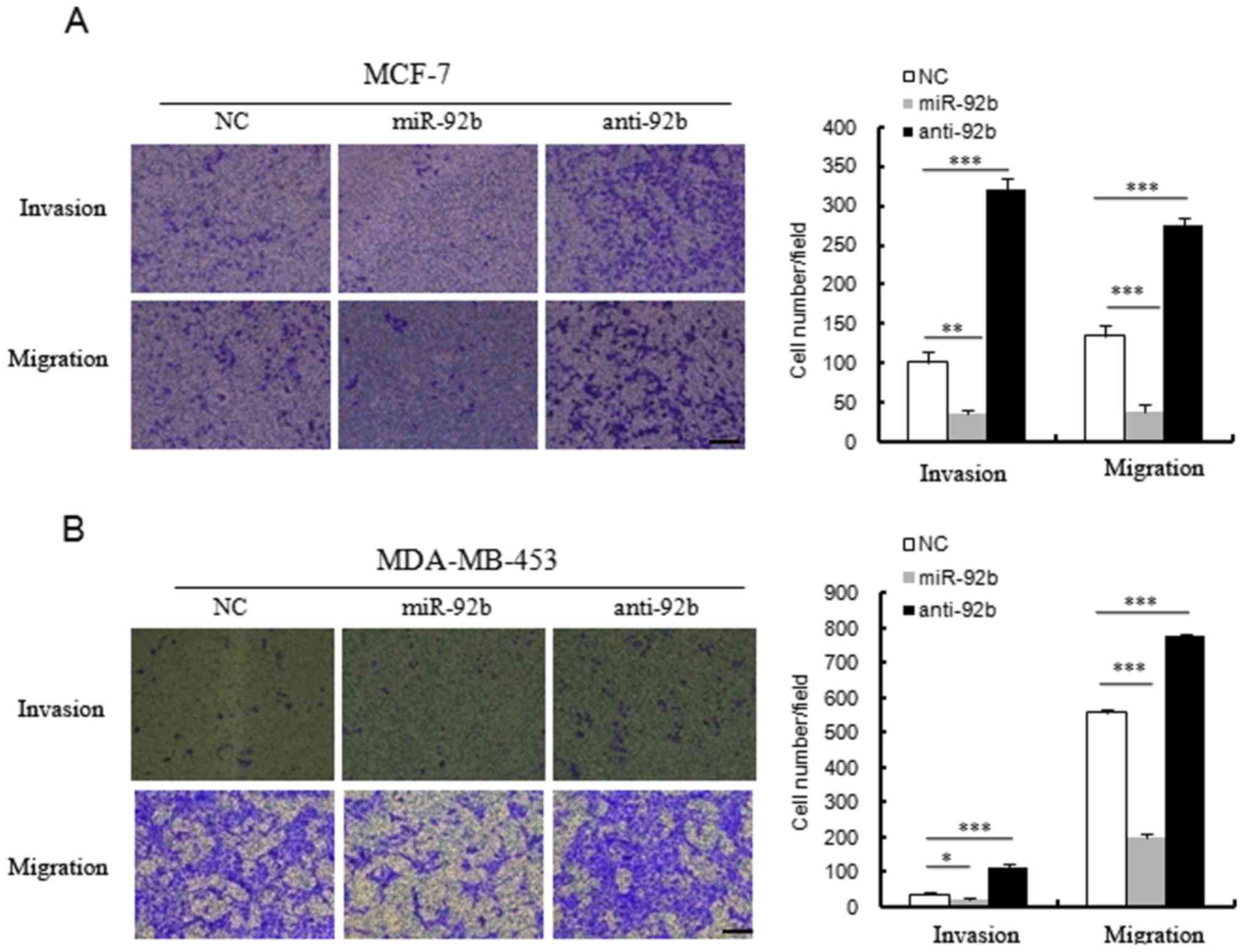
miR-92b inhibits the invasion and
migration ability of breast cancer cells
In order to examine the effect of miR-92b on the
malignant biological processes in breast cancer cells, the present
study firstly examined the role of miR-92b in the migration of MCF7
and MDA-MB-453 cells. Transwell migration assays indicated that the
number of migrating cells was reduced in miR-92b mimics-transfected
MCF7 and MDA-MB-453 cells when compared with the control group
(Fig. 3A and B). In addition, the
suppression of endogenous miR-92b with its inhibitor enhanced cell
motility when compared with control cells (Fig. 3A and B).
Furthermore, the Matrigel invasion assay system was
used to examine the effect of miR-92b on MCF7 and MDA-MB-453 cell
invasion ability. Overexpression of miR-92b decreased the invasion
ability of MCF7 and MDA-MB-453 cells (Fig. 3A and B). Consistently, transfection
with the miR-92b inhibitor resulted in the enhanced invasion
ability of these cells (Fig. 3A and
B). Therefore, these results demonstrated that miR-92b can
affect the migration and invasion ability of breast cancer
cells.
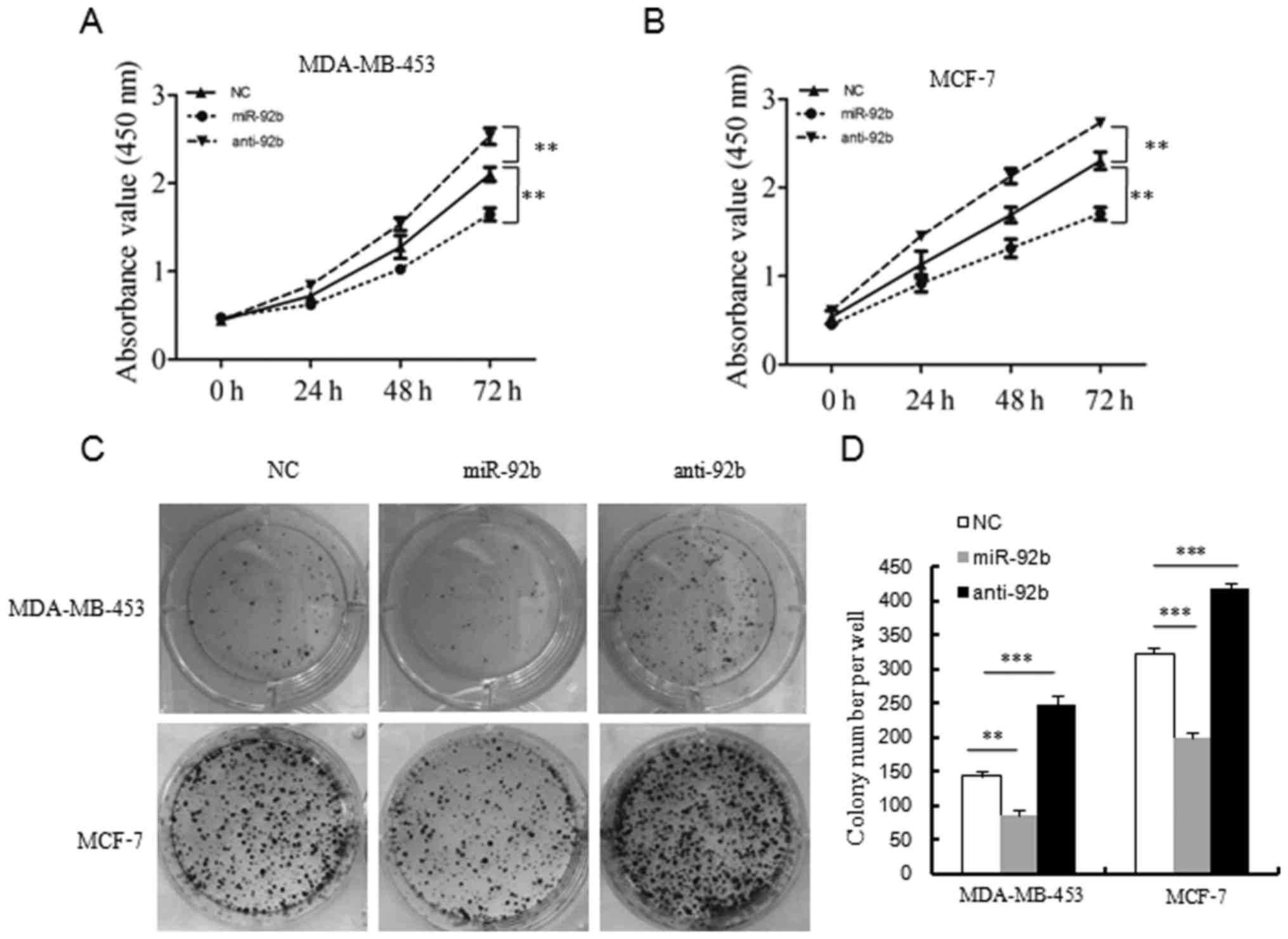
miR-92b inhibits the viability of breast
cancer cells
In order to examine the effects of miR-92b on breast
cancer cell viability, the present study performed CCK8 assays and
colony formation assays. When compared with the control groups, the
transfection-induced overexpression of miR-92b in breast cancer
cells suppressed the viability of breast cancer cells over time
(Fig. 4A and B). By contrast,
suppressing endogenous miR-92b with its inhibitor promoted the
viability of breast cancer cells (Fig.
4A and B).
In line with the cell viability assays, the colony
formation assays also demonstrated that the MCF7 and MDA-MB-453
cells transfected with miR-92b exhibited decreased colony formation
abilities in comparison with those of the control groups (Fig. 4C and D). In addition, suppressing
endogenous miR-92b with its inhibitor enhanced colony formation
(Fig. 4C and D).
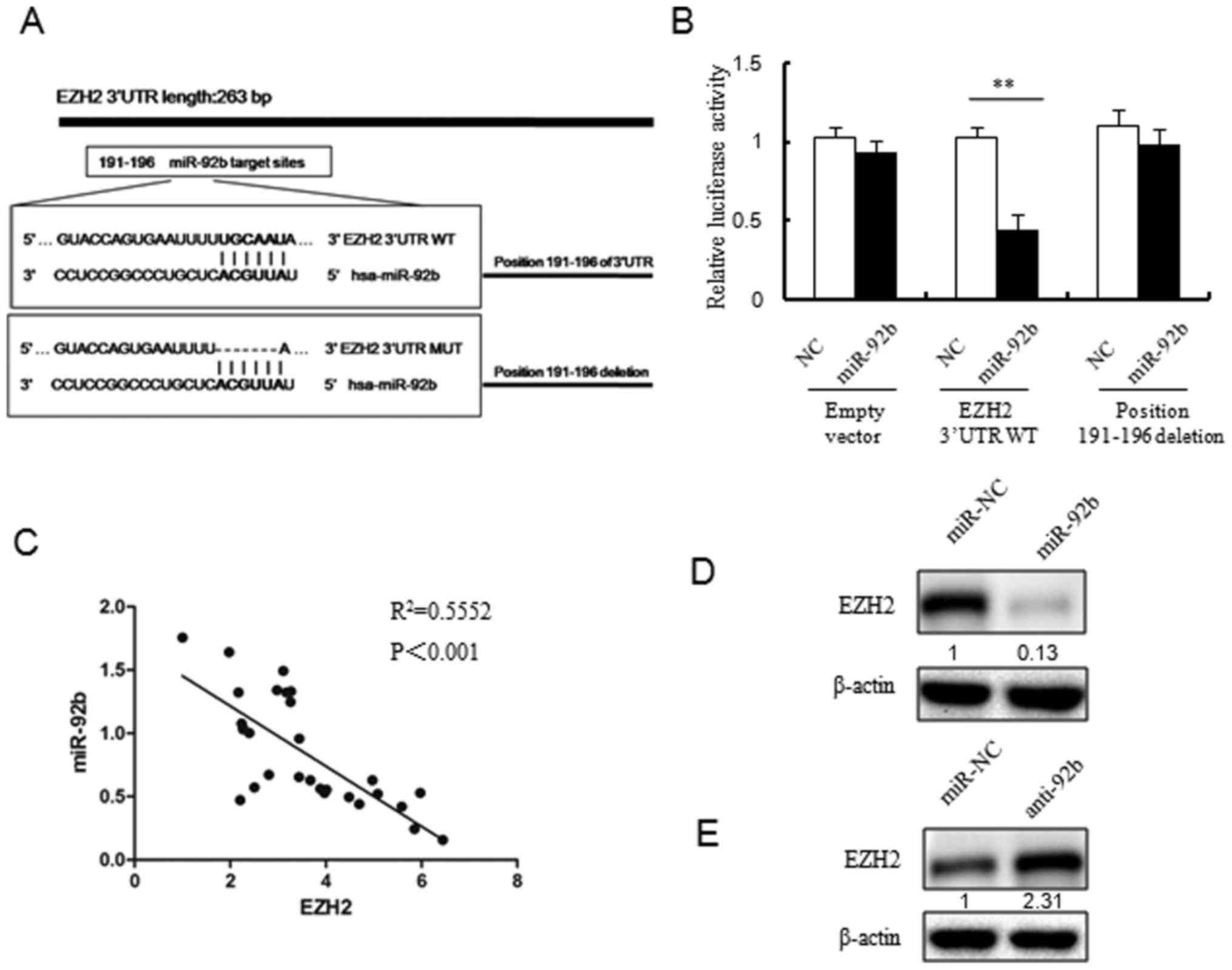
EZH2 is a direct target of miR-92b
TargetScan analysis revealed that there was a
miR-92b binding site in the 3′-UTR region of EZH2 (Fig. 5A). In order to eliminate the
individual differences caused by transfection efficiency issues,
the present study chose to use cos7 cells for the Luciferase
reporter assay due to their high transfection efficiency. The
luciferase reporter assay demonstrated that luciferase activity was
inhibited by co-transfection the wild-type 3′-UTR of EZH2 and
miR-92b mimic, and the deletion of the miR-92b target site
inhibited this decrease in luciferase activity (Fig. 5B).
Then, the expression of miR-92b and EZH2 mRNA in 30
pairs of breast cancer tissues was analyzed by RT-qPCR. Similarly,
a negative correlation was observed between miR-92b expression and
EZH2 mRNA expression in breast cancer specimens (P<0.001;
Fig. 5C).
In addition, the present study also examined EZH2
protein expression levels in MCF7 cells transfected with miR-92b
mimics, miR-92b inhibitor or miR-NC (Fig. 5D and E). The EZH2 protein levels
were reduced in MCF7 cells with miR-92b overexpression (Fig. 5D). In line with this, EZH2 protein
expression was upregulated in miR-92b inhibitor transfected MCF7
cells (Fig. 5E).
Effects of EZH2 silencing on autophagy,
viability and invasion
To evaluate the effects of EZH2 on autophagy, the
present study transfected MCF7 cells with EZH2 shRNA or Empty
vector and analyzed the levels of autophagy induced by starvation
or rapamycin treatment. As shown in Fig. 6, EZH2 silencing enhanced the
autophagy of MCF-7 cells. EZH2 shRNAs enhanced the lipidation of
LC3 and SQSTM1 degradation (Fig. 6A
and B) induced by starvation or rapamycin. Similarly, EZH2
silencing also enhanced starvation- or rapamycin-induced GFP-LC3
dot accumulation (Fig. 6C–F).
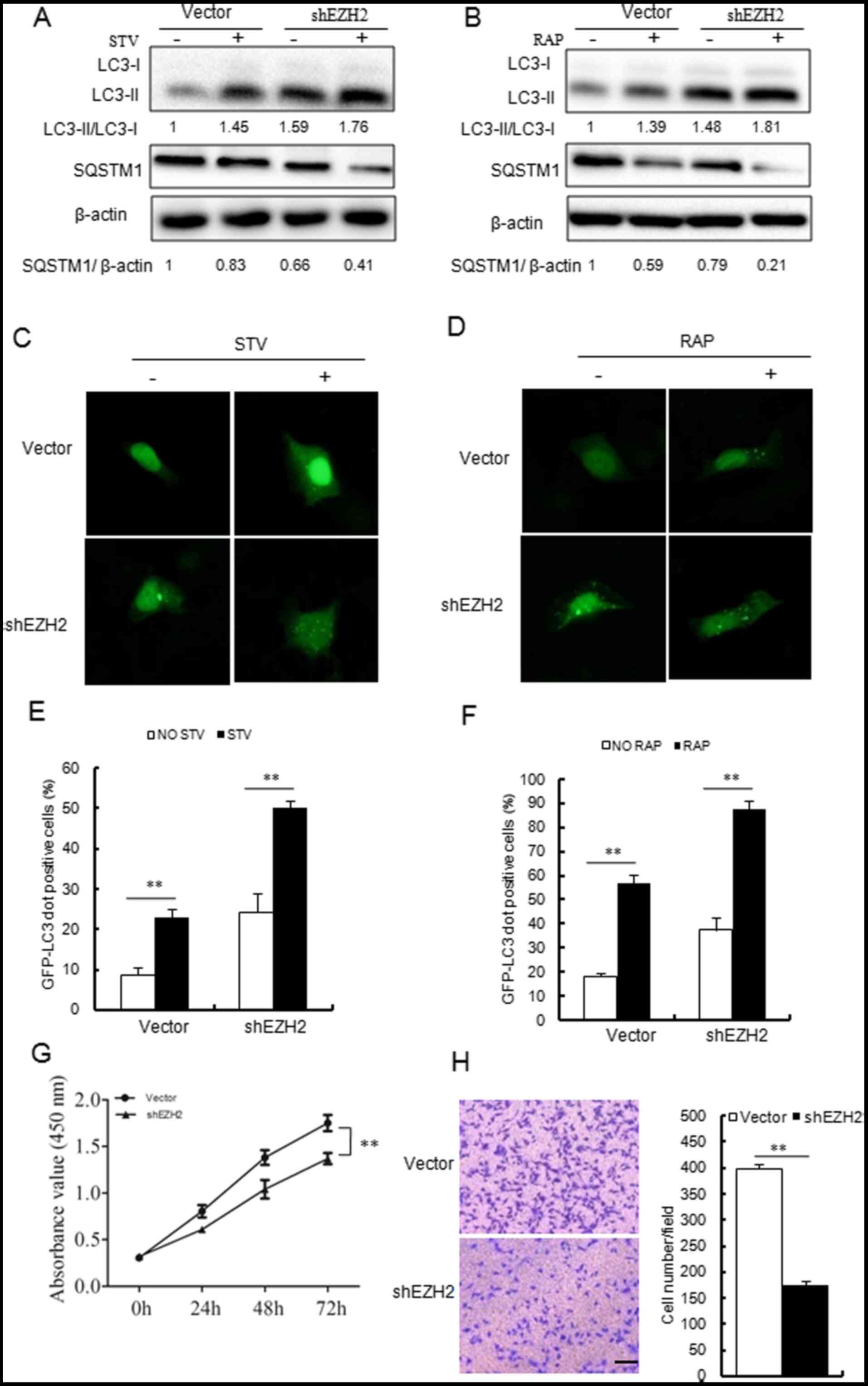 | Figure 6Silencing EZH2 expression with EZH2
shRNA promotes autophagy, and suppresses proliferation and invasion
in MCF-7 cells. (A and B) Immunoblotting results indicated that
shEZH2 increased (A) STV and (B) RAP induced conversion of LC3-I to
LC3-II and SQSTM1 degradation. (C and D) shEZH2 promoted STV and
RAP induced GFP-LC3 dot formation in MCF-7 cells (magnification,
×40). (E and F) Quantitative analyses of the experiments in panels
(C and D). (G and H) shEZH2 inhibited the proliferation and
invasion abilities of MCF-7 cells. Scale bars = 100 µm;
magnification, ×200. **P<0.01, as indicated. EZH2,
enhancer of zeste homolog 2; STV, starvation; RAP, rapamycin; miR,
microRNA; shRNA, short hairpin RNA; LC3, light chain 3; GFP, green
fluorescent protein; SQSTM1, sequestosome 1; NC, negative
control. |
The present study then examined the effects of EZH2
knockdown on breast cancer cell growth and invasion. shEZH2 plasmid
and the control empty plasmid were transfected into MCF-7 cells.
According to the results presented in Fig. 6G, cell viability was inhibited
following shEZH2 plasmid transfection. In addition, the invasion
ability of MCF7 cells was also inhibited in the shEZH2 plasmid
transfected group when compared with the vector control group
(Fig. 6H).
EZH2 is involved in miR-92b regulated
cell autophagy, viability, migration and invasion
To further verify the functional connection between
miR-92b and EZH2, MCF7 cells were cotransfected with miR-92b mimic
and EZH2 expression vector or control (Fig. 7A). Overexpression of EZH2 was
demonstrated to reverse the inhibitory effects of miR-92b on cell
autophagy, viability, migration and invasion in MCF7 cells
(Fig. 7B–D). These results
indicated that miR-92b may inhibit cell autophagy, viability and
invasion by regulating EZH2 expression.
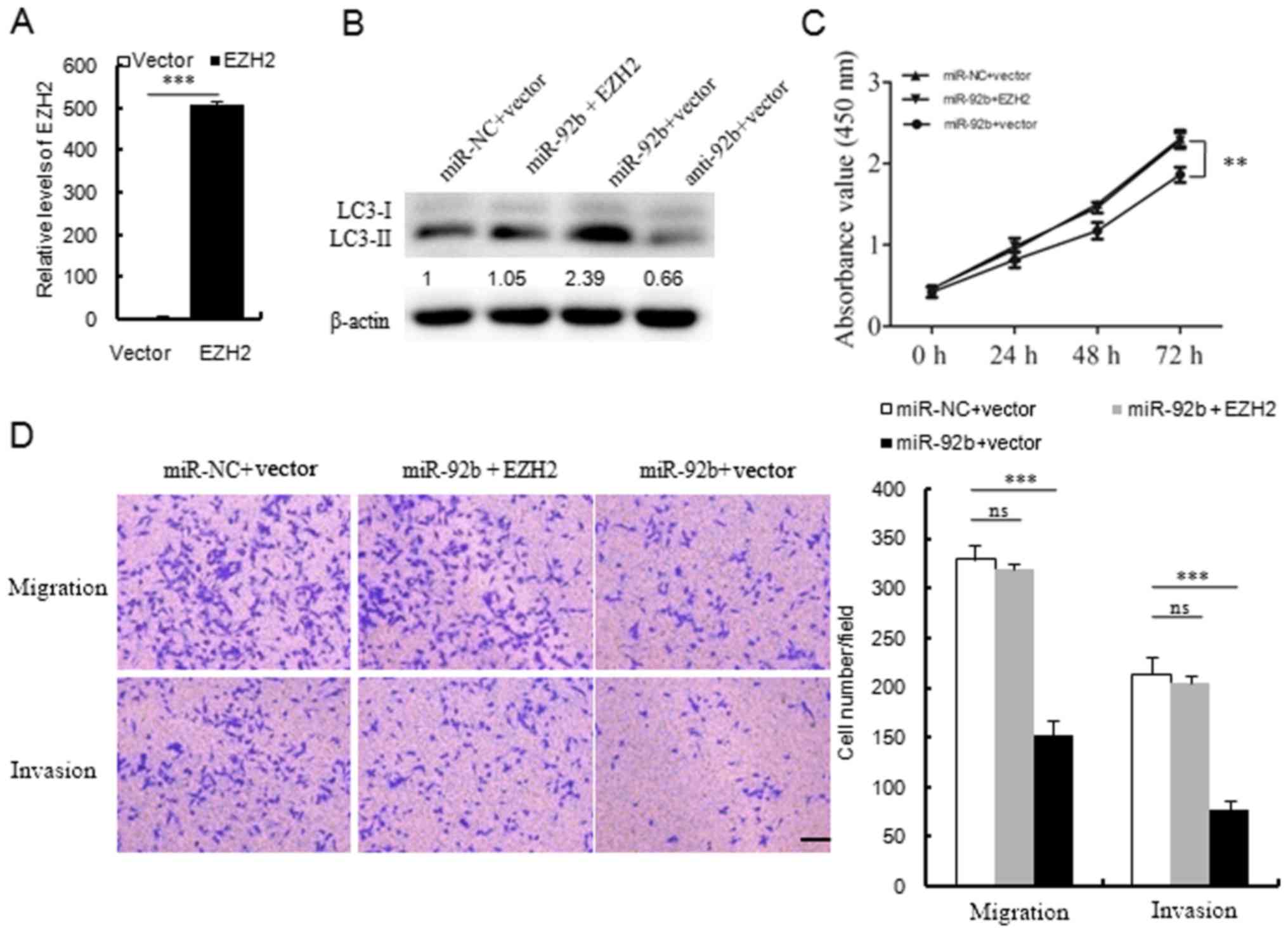 | Figure 7EZH2 is involved in miR-92b regulated
cell autophagy, proliferation and invasion. (A) Reverse
transcription-quantitative polymerase chain reaction revealed the
relative mRNA expression of EZH2 in MCF7 cells transfected with the
pCMV6 Entry-EZH2-Myc-DDK expression plasmid or the control plasmid.
(B) Western blot analyses of LC3 expression in MCF-7 cells
transfected with miR-NC plus vector, miR-92b plus EZH2, miR-92b
plus vector or anti-miR-92b (anti-92b) plus vector.
Autophagy-associated LC3-I to LC3-II conversion (LC3 lipidation)
was attenuated in anti-miR-92b plus vector overexpressing cell
extracts, when compared with miR-NC plus vector as the control
group. (C) Cell Counting Kit-8 analysis of MCF7 cells transfected
with miR-NC plus vector, miR-92b plus EZH2 and miR-92b plus vector
at the indicated times. (D) Transwell analysis of MCF-7 cells
treated with miR-NC plus vector, miR-92b plus EZH2 and miR-92b plus
vector, and the quantitative analysis is shown in the right-hand
panel. Scale bar = 100 µm; magnification, ×200.
**P<0.01 and ***P<0.001, as indicated.
EZH2, enhancer of zeste homolog 2; miR, microRNA; LC3, light chain
3; NC, negative control; ns, not significant. |
Discussion
In the present study, it was revealed that
endogenous miR-92b expression levels are elevated in response to
different autophagy inducing stimuli. Notably, miR-92b
overexpression enhanced GFP-LC3 dot formation, the lipidation of
LC3 and SQSTM1 degradation. These results were observed following
starvation and rapamycin treatment in MCF7 and MDA-MB-453 cells.
Additionally, it was detected that overexpression of miR-92b in
MCF7 and MDA-MB-453 cells decreased tumor cell viability, migration
and invasion. The present study also identified a novel miR-92b
target, EZH2, and demonstrated that shRNA-mediated knockdown of
EZH2 phenocopied the effect of miR-92b, suggesting that miR-92b may
exert its pro-autophagic function via EZH2. Taken together, these
results indicated that miR-92b may negatively regulate the
expression of EZH2, promoting autophagy, and decreasing tumor cell
viability, migration and invasion.
miR-92b is involved in the progression of the
central neural system development and the development of several
tumors (27,28). Previous reports on glioblastoma and
non-small cell lung cancer have suggested that miR-92b promoted
neoplastic cell growth and metastasis (29,30).
However, in esophageal squamous cell carcinoma, miR-92b did not
stimulate apoptosis or inhibit proliferation, though it did impair
the motility of cancer cells (31,32).
In accordance with Ma et al (31,32),
the present study observed that there was significantly reduced
miR-92b levels in human breast cancer tissues in comparison with
normal tissue specimens, and overexpression of miR-92b in breast
cancer cells decreased tumor cell proliferation, invasion and
migration. These results supported the notion that one specific
miRNA may exert a variety of effects in different genetic contexts
(33). The mechanism for miR-92b
underlying a tumors potential is rather complex, as it is mediated
by a varied range of targets that are potentially variable
depending on the type of cancer and the environmental stimuli. The
results of the present study provide further support for the
functional relevance of miR-92b in breast cancer via the
identification of the methyltransferase EZH2 as its novel target
during starvation- and rapamycin-induced autophagy.
EZH2 is the core catalytic subunit of the PRC2
complex. PRC2 methylates the lysine residue at position 27 of
histone 3 (H3K27) (34,35), which results in chromatin
compaction and epigenetic silencing of genes. EZH2 overexpression
has been reported in different types of malignancies, and was
associated with malignant tumor growth, metastasis and poor
prognosis in a variety of human malignancies (36–39).
Recently, it has been reported that EZH2 may serve an important
role in autophagy (40). Notably,
the present study demonstrated the post-transcriptional
modification of EZH2 by miR-92b and its involvement in autophagy.
It has been indicated that EZH2-mediated gene promoter methylation
can promote the malignant phenotype of different cancer cells, due
to the loss of tumor suppressor gene functions, which results in
the transformation of stem cells into cancer cells (41–43);
this may partly explain the reduced malignant properties induced by
miR-92b.
In conclusion, the present study confirmed that
miR-92b may be a crucial regulator of starvation- and
rapamycin-induced autophagy by targeting the methyltransferase
EZH2. This post-transcriptional regulation had a significant effect
on inhibiting the oncogenic phenotypes of tumor cells. In view of
these results revealing enhanced miR-92b expression and attenuated
EZH2 expression in human breast cancer cells, it was hypothesized
that the autophagy-associated miR-92b-EZH2 axis may be an appealing
target for therapeutic interventions in breast cancer.
Acknowledgments
Not applicable.
Funding
The present study was supported by the Medical
Science Research Foundation of Health and Family Planning
Commission of Hebei (grant no. 20180532), the Science Fund for
Outstanding Youth of Hebei Province (grant no. 2016206410) and the
Science and Technology Supporting Program of Hebei Province (grant
no. 152777184).
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors' contributions
FL, MS and LM performed the molecular studies and
drafted the manuscript. SL and JL collected the clinical data and
tissue samples. FL and LG performed the statistical analysis. CG
designed the study and helped to draft the manuscript. All authors
read and approved the final manuscript.
Ethics approval and consent to
participate
The use of human tissues was approved by the Ethics
Committee of the Fourth Hospital of Hebei Medical University
(Hebei, China) and written informed consent was obtained from all
patients.
Patient consent for publication
Consent for the publication of the clinical and
pathological data was obtained from all patients who were involved
in the present study.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Mizushima N: Autophagy: Process and
function. Genes Dev. 21:2861–2873. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Mizushima N, Yoshimori T and Ohsumi Y: The
role of Atg proteins in autophagosome formation. Annu Rev Cell Dev
Biol. 27:107–132. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Klionsky DJ: Autophagy: From phenomenology
to molecular understanding in less than a decade. Nat Rev Mol Cell
Biol. 8:931–937. 2007. View
Article : Google Scholar : PubMed/NCBI
|
|
4
|
Galluzzi L, Pietrocola F, Bravo-San Pedro
JM, Amaravadi RK, Baehrecke EH, Cecconi F, Codogno P, Debnath J,
Gewirtz DA, Karantza V, et al: Autophagy in malignant
transformation and cancer progression. EMBO J. 34:856–880. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Amaravadi R, Kimmelman AC and White E:
Recent insights into the function of autophagy in cancer. Genes
Dev. 30:1913–1930. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Levy JM and Thorburn A: Targeting
autophagy during cancer therapy to improve clinical outcomes.
Pharmacol Ther. 131:130–141. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Towers CG and Thorburn A: Therapeutic
targeting of autophagy. EBioMedicine. 14:15–23. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Jing Z, Han W, Sui X, Xie J and Pan H:
Interaction of autophagy with microRNAs and their potential
therapeutic implications in human cancers. Cancer Lett.
356B:332–338. 2015. View Article : Google Scholar
|
|
9
|
Fu LL, Wen X, Bao JK and Liu B:
MicroRNA-modulated autophagic signaling networks in cancer. Int J
Biochem Cell Biol. 44:733–736. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Xu J, Wang Y, Tan X and Jing H: MicroRNAs
in autophagy and their emerging roles in crosstalk with apoptosis.
Autophagy. 8:873–882. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Iorio MV and Croce CM: MicroRNAs in
cancer: Small molecules with a huge impact. J Clin Oncol.
27:5848–5856. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Bartel DP: MicroRNAs: Genomics,
biogenesis, mechanism, and function. Cell. 116:281–297. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Huang Y, Chuang AY and Ratovitski EA:
Phospho-∆Np63α/miR-885-3p axis in tumor cell life and cell death
upon cisplatin exposure. Cell Cycle. 10:3938–3947. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Zhu H, Wu H, Liu X, Li B, Chen Y, Ren X,
Liu CG and Yang JM: Regulation of autophagy by a Beclin 1-targeted
microRNA, miR-30a, in cancer cells. Autophagy. 5:816–823. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Chang Y, Yan W, He X, Zhang L, Li C, Huang
H, Nace G, Geller DA, Lin J and Tsung A: miR-375 inhibits autophagy
and reduces viability of hepatocellular carcinoma cells under
hypoxic conditions. Gastroenterology. 143:177–187.e8. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Korkmaz G, le Sage C, Tekirdag KA, Agami R
and Gozuacik D: miR-376b controls starvation and mTOR
inhibition-related autophagy by targeting ATG4C and BECN1.
Autophagy. 8:165–176. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Tekirdag KA, Korkmaz G, Ozturk DG, Agami R
and Gozuacik D: MIR181A regulates starvation- and rapamycin-induced
autophagy through targeting of ATG5. Autophagy. 9:374–385. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Ren Y, Chen Y, Liang X, Lu Y, Pan W and
Yang M: miRNA-638 promotes autophagy and malignant phenotypes of
cancer cells via directly suppressing DACT3. Cancer Lett.
390:126–136. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Guo W, Wang H, Yang Y, Guo S, Zhang W, Liu
Y, Yi X, Ma J, Zhao T, Liu L, et al: Down-regulated miR-23a
contributes to the metastasis of cutaneous melanoma by promoting
autophagy. Theranostics. 7:2231–2249. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Wang Y, Zhang X, Tang W, Lin Z, Xu L, Dong
R, Li Y, Li J, Zhang Z, Li X, et al: miR-130a upregulates mTOR
pathway by targeting TSC1 and is transactivated by NF-κB in
high-grade serous ovarian carcinoma. Cell Death Differ.
24:2089–2100. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Du X, Huo X, Yang Y, Hu Z, Botchway BOA,
Jiang Y and Fang M: miR-124 downregulates BACE 1 and alters
autophagy in APP/PS1 transgenic mice. Toxicol Lett. 280:195–205.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Triozzi PL, Achberger S, Aldrich W, Crabb
JW, Y and Singh AD: Association of tumor and plasma microRNA
expression with tumor monosomy-3 in patients with uveal melanoma.
Clin Epigenetics. 8:802016. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Zhou Z, Wang Z, Wei H, Wu S, Wang X and
Xiao J: Promotion of tumour proliferation, migration and invasion
by miR-92b in targeting RECK in osteosarcoma. Clin Sci (Lond).
130:921–930. 2016. View Article : Google Scholar
|
|
24
|
Huang J, Wang B, Hui K, Zeng J, Fan J,
Wang X, Hsieh JT, He D and Wu K: miR-92b targets DAB2IP to promote
EMT in bladder cancer migration and invasion. Oncol Rep.
36:1693–1701. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Zhao C, Zhao F, Feng H, Xu S and Qin G:
MicroRNA-92b inhibits epithelial-mesenchymal transition-induced
migration and invasion by targeting Smad3 in nasopharyngeal cancer.
Oncotarget. 8:91603–91613. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) Method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
27
|
Nowakowski TJ, Fotaki V, Pollock A, Sun T,
Pratt T and Price DJ: MicroRNA-92b regulates the development of
intermediate cortical progenitors in embryonic mouse brain. Proc
Natl Acad Sci USA. 110:7056–7061. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Zhou X, Zhu W, Li H, Wen W, Cheng W, Wang
F, Wu Y, Qi L, Fan Y, Chen Y, et al: Diagnostic value of a plasma
microRNA signature in gastric cancer: A microRNA expression
analysis. Sci Rep. 5:112512015. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Wu ZB, Cai L, Lin SJ, Lu JL, Yao Y and
Zhou LF: The miR-92b functions as a potential oncogene by targeting
on Smad3 in glioblastomas. Brain Res. 1529:16–25. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Li Y, Li L, Guan Y, Liu X, Meng Q and Guo
Q: miR-92b regulates the cell growth, cisplatin chemosensitivity of
A549 non small cell lung cancer cell line and target PTEN. Biochem
Biophys Res Commun. 440:604–610. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Ma G, Jing C, Li L, Huang F, Ding F, Wang
B, Lin D, Luo A and Liu Z: MicroRNA-92b represses
invasion-metastasis cascade of esophageal squamous cell carcinoma.
Oncotarget. 7:20209–20222. 2016.PubMed/NCBI
|
|
32
|
Ma G, Jing C, Huang F, Li X, Cao X and Liu
Z: Integrin α6 promotes esophageal cancer metastasis and is
targeted by miR-92b. Oncotarget. 8:6681–6690. 2017.
|
|
33
|
Vidigal JA and Ventura A: The biological
functions of miRNAs: Lessons from in vivo studies. Trends Cell
Biol. 25:137–147. 2015. View Article : Google Scholar :
|
|
34
|
Müller J, Hart CM, Francis NJ, Vargas ML,
Sengupta A, Wild B, Miller EL, O'Connor MB, Kingston RE and Simon
JA: Histone methyltransferase activity of a Drosophila Polycomb
group repressor complex. Cell. 111:197–208. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Czermin B, Melfi R, McCabe D, Seitz V,
Imhof A and Pirrotta V: Drosophila enhancer of Zeste/ESC complexes
have a histone H3 methyltransferase activity that marks chromosomal
Polycomb sites. Cell. 111:185–196. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Varambally S, Dhanasekaran SM, Zhou M,
Barrette TR, Kumar-Sinha C, Sanda MG, Ghosh D, Pienta KJ, Sewalt
RG, Otte AP, et al: The polycomb group protein EZH2 is involved in
progression of prostate cancer. Nature. 419:624–629. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Kleer CG, Cao Q, Varambally S, Shen R, Ota
I, Tomlins SA, Ghosh D, Sewalt RG, Otte AP, Hayes DF, et al: EZH2
is a marker of aggressive breast cancer and promotes neoplastic
transformation of breast epithelial cells. Proc Natl Acad Sci USA.
100:11606–11611. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Collett K, Eide GE, Arnes J, Stefansson
IM, Eide J, Braaten A, Aas T, Otte AP and Akslen LA: Expression of
enhancer of zeste homologue 2 is significantly associated with
increased tumor cell proliferation and is a marker of aggressive
breast cancer. Clin Cancer Res. 12:1168–1174. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Liu F, Gu L, Cao Y, Fan X, Zhang F and
Sang M: Aberrant overexpression of EZH2 and H3K27me3 serves as poor
prognostic biomarker for esophageal squamous cell carcinoma
patients. Biomarkers. 21:80–90. 2016. View Article : Google Scholar
|
|
40
|
Wei FZ, Cao Z, Wang X, Wang H, Cai MY, Li
T, Hattori N, Wang D, Du Y, Song B, et al: Epigenetic regulation of
autophagy by the methyltransferase EZH2 through an MTOR-dependent
pathway. Autophagy. 11:2309–2322. 2015. View Article : Google Scholar
|
|
41
|
Schlesinger Y, Straussman R, Keshet I,
Farkash S, Hecht M, Zimmerman J, Eden E, Yakhini Z, Ben-Shushan E,
Reubinoff BE, et al: Polycomb-mediated methylation on Lys27 of
histone H3 pre-marks genes for de novo methylation in cancer. Nat
Genet. 39:232–236. 2007. View
Article : Google Scholar : PubMed/NCBI
|
|
42
|
Tonini T, D'Andrilli G, Fucito A, Gaspa L
and Bagella L: Importance of Ezh2 polycomb protein in tumorigenesis
process interfering with the pathway of growth suppressive key
elements. J Cell Physiol. 214:295–300. 2008. View Article : Google Scholar
|
|
43
|
Widschwendter M, Fiegl H, Egle D,
Mueller-Holzner E, Spizzo G, Marth C, Weisenberger DJ, Campan M,
Young J, Jacobs I, et al: Epigenetic stem cell signature in cancer.
Nat Genet. 39:157–158. 2007. View
Article : Google Scholar : PubMed/NCBI
|