Stress is a constellation of events that begins with
a stressor, leading to stress perception and subsequently resulting
in stress response (1). Stress can
impair the balance or homeostatic state of an organism
physiologically or psychologically when exposed to extrinsic or
intrinsic adverse forces (2). It
is well known that stress is ubiquitous and unavoidable in life.
For certain individuals, stress can serve as a stimulant under
certain conditions and as a burden under others. In addition,
stress serving as a stimulant in certain individuals may be a
burden for others, which indicates that there exists a varying
degree of stress susceptibility among individuals (3,4).
Stress occurs when an organism perceives a disruption, or a threat
of disruption, of homeostasis. Factors that can induce stress
include, among others, social circumstances, physical environment,
hypoxia, emotional state, pain, glucose deprivation, accidents and
chronic anxiety (5).
Stress-related symptoms include negative emotions or altered mood
and behavior, such as anxiety, irritability, anger, startle
response, hostility and depression, which can negatively affect
various organs of the human body (6). Studies have preliminarily revealed
that differences in the stress experienced by different individuals
can be due to genetic and environmental factors (7,8). It
is also generally believed that moderate stress, such as routine or
regular exercise or physical activity, can enhance the body’s
immune response and reduce the risk of cancer occurrence,
progression and mortality (9,10),
while chronic stress can affect a number of physiological functions
and lead to several diseases, including cancer (11-13)
(Table I). Stressors can result in
mood and anxiety disorders, including depression, anxiety and
irritability, which may be associated with malignancies (14,15).
In order to elucidate the underlying mechanisms
linking stress with tumorigenesis and metastasis, it is necessary
to establish effective animal models. At present, the most common
animal stress models are those developed for chronic restraint
stress, maternal separation, and dietary and environmental stress.
Tumor metastasis is a complex process that consists of
proliferation/angiogenesis, detachment/invasion,
embolism/circulation and evasion of immune system surveillance. The
present review attempted to classify and summarize the mechanisms
underlying the role of stress in tumorigenesis and cancer
progression.
Animal models mimicking the pattern of human
diseases serve a key role in understanding the effects of stress on
cancer. To study this effect, multiple physiological stress models
have been utilized to promote fear or anxiety in rodents, including
the following: i) Social isolation model, where laboratory rodents,
which are highly social creatures, are housed individually in cages
for extended periods to elicit loneliness; ii) restraint stress
model, in which the animals are immobilized or confined to small
spaces; and iii) intimidation-induced stress model, which involves
placing rodents into the cage of another animal. When categorized
by type, stressors are classified as physical (trauma/injury and
exercise/exhaustion), cognitive (anxiety and depression), a
combination of physical and cognitive (fire-fighting on a 24-h
shift) or chemical (environmental toxins and diet). When
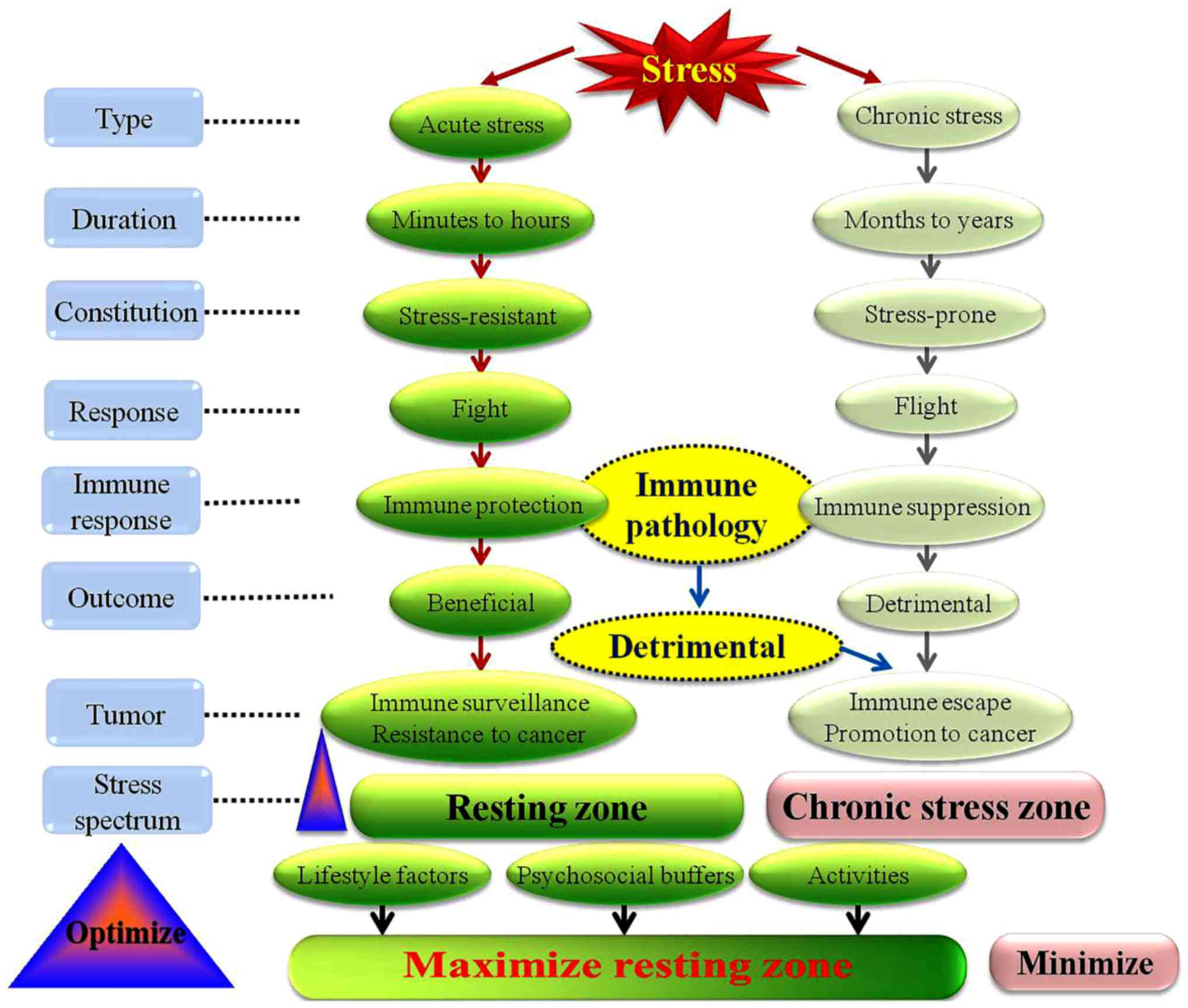
categorized by duration, stressors are classified as acute (minutes
to hours), and chronic (months to years) (16,17).
At present, the chronic mild stress (CMS) model is
one of the most widely used stress models, allowing for a
combination of a large variety of stressors with different
numbers/lengths of intervals and the measurement of different
behaviors as a response to rewards (18). Specifically, the CMS model consists
of immobilization, forced swimming, noise, hypothermia, social
isolation, resident/intruder aggression, maternal deprivation,
stroboscopic illumination, cage titling, and food, water or sleep
deprivation (19). Meanwhile, the
chronic restraint stress animal model is one of the most common
immobilizations, in which the duration of restraint has ranged from
1 to 12 h daily (20,21). Generally speaking, these stress
models are evaluated based on three major criteria: i) Construct
validity, where the experimental conditions are replicating causes
of human diseases; ii) face validity, where the symptoms observed
in diseased animals are consistent with clinical observations; and
iii) predictive validity, where the animal responds to the
treatment currently used in the clinic (22). However, no studies have
systematically evaluated the validity or the efficiency of the CMS
model, as researchers have often chosen a particular combination of
stressors with timing based on previous practical experience and/or
the particular requirements of their experiments (18). Stress can be harmful when it is
chronic or long lasting, but a fact that is often overlooked is
that a stress response can have salubrious adaptive effects in the
short run, and that short-term stress may enhance cellular immunity
and increase early resistance to cancer (23,24).
The timing or duration of the stress models is an important factor
that requires consideration. Therefore, it is essential to
distinguish between acute and chronic stress in the following
discussion (Fig. 1).
Effects of stress on the neuroendocrine system. In
the fast-paced society of today, the pressure on individuals is
high, and it often manifests itself in the form of anxiety,
tension, insomnia and depression, all of which can lead to chronic
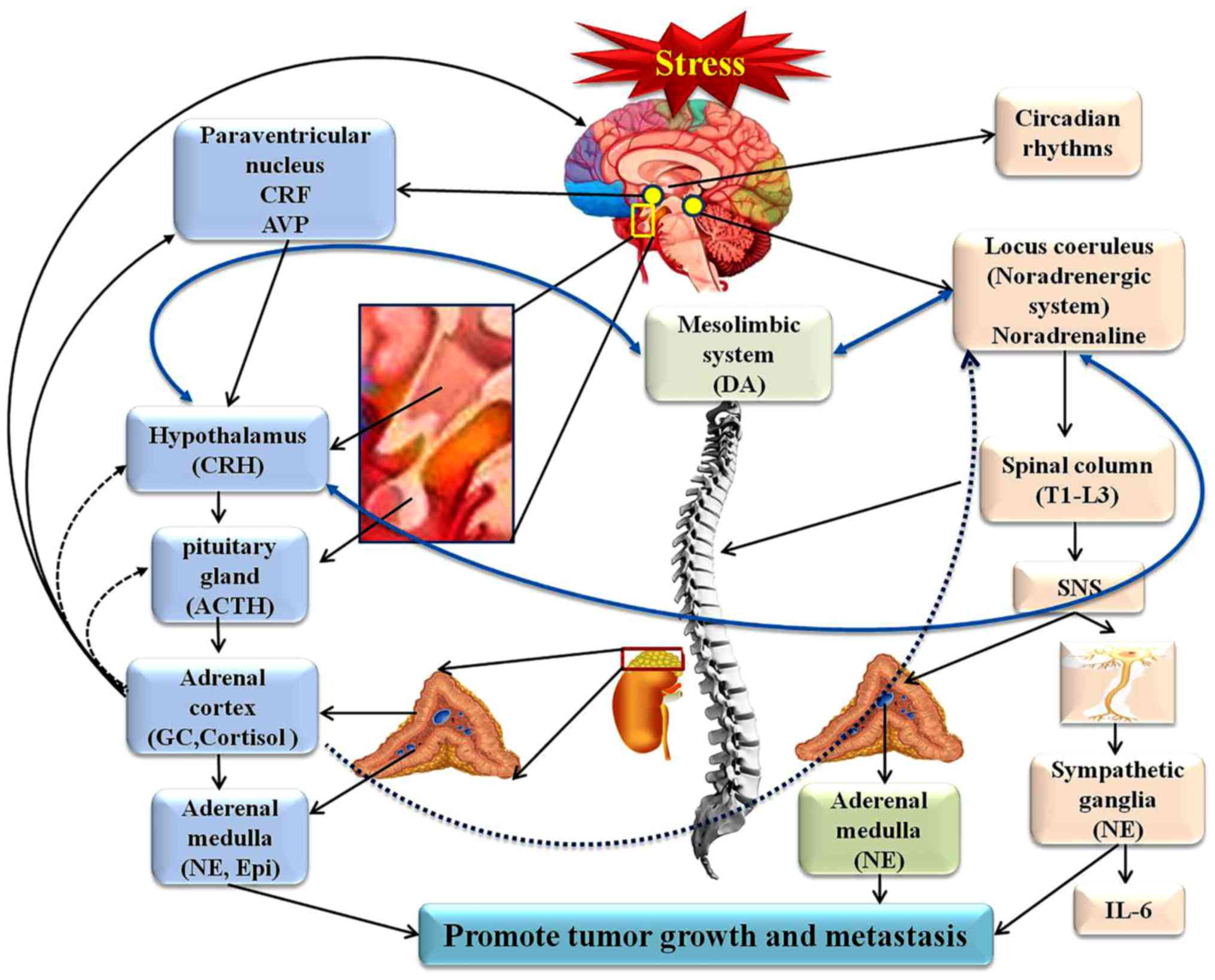
stress. The hypothalamic-pituitary-adrenal (HPA) axis and the
sympathetic nervous system (SNS) are the two branches of the
neuroendocrine system that govern the response to stress (25,26).
Corticotrophin-releasing hormone (CRH) and arginine vasopressin
(AVP), released from the hypothalamus, can activate the pituitary
gland to release adrenocorticotropic hormone (ACTH), encephalin and
endorphin (END), and the adrenal cortex to release ACTH-induced
glucocorticoid (GC) (3,27). The release of CRH is under
excitatory input from the amygdala and inhibitory input from the
hippocampus (28). Meanwhile, the
secretion of CRH and AVP is characterized by a precise circadian
rhythm that can be disrupted by imposed stressors, and a circadian
rhythm disorder can have the same detrimental effects as chronic
stress (29,30). Stress can also result in increased
secretion of catecholamines, including norepinephrine (NE) and
epinephrine (E), which can enhance alertness and physiological
functions, and elicit the fight-or-flight response (31,32).
GC serves an important role in the treatment and chemo-resistance
of tumors, and catecholamine can promote tumor growth and
metastasis (33,34). Adolescent chronic stress can cause
HPA hypo-responsiveness and depression-like behavior (35). Psychosocial stressors in cancer can
result in the dysregulation of the HPA axis, and vice versa
(36). Restraint stress
facilitates cancer angiogenesis and metastasis by releasing β-END,
prolactin, increasing concentrations of circulating catecholamine
and GC, and increasing tumor-associated macrophage infiltration
into the primary tumor (37,38).
Psychological stress may attenuate antiangiogenic therapy,
primarily through activating β-adrenergic signaling to promote
tumor angiogenesis (39), and
β-blockers or behavioral therapies can limit skeletal metastasis of
breast cancer cells (40).
Dopamine (DA) is also a catecholamine hormone, which can stabilize
tumor blood vessels to block the effects of chronic stress on tumor
vasculature, as the depletion of DA under chronic stress conditions
creates a permissive microenvironment for tumor growth (41,42).
Notably, studies have revealed that DA acts through five types of
DA receptors; DA type-1 receptor overexpression is associated with
advanced breast cancer and a poor prognosis (43), but DA type-2 receptor has been
found to inhibit tumor growth (44).
Furthermore, since the lymphatic system and the
pancreas are innervated by fibers of the SNS and have receptors for
SNS neurotransmitters, chronic stress-induced SNS activity can
increase pancreatic cancer growth, lymphatic vessel contraction and
lymphocyte output into the lymphatic circulation. These processes
may affect tumor lymphatic dissemination and cancer progression
(45,46). Chronic restraint stress can
attenuate the levels and function of p53 proteins, and promote the
growth of human xenograft tumors, which is mediated by GC elevation
during chronic restraint stress (47). A study has shown that exposure to
chronic psychological stress may lead to significant changes in the
proteomic profile of tumors (48).
It is noteworthy that cytokines are potent activators of the
central stress response and have regulatory effects on the HPA axis
by forming a feedback loop through which the immune/inflammatory
system communicates with the brain (49). The duration and magnitude of
stress-induced increases in NE, Epi and GC have significant effects
on immune cell redistribution and function (50,51)
(Fig. 2). However, several studies
have indicated that β-adrenergic receptor blockers can reverse the
effects of chronic stress on cancer progression and abrogate drug
resistance, which merits further investigation as a novel strategy
for cancer treatment (52-54).
Effects of stress on immune function. Stress exerts
pleiotropic effects on the immune system, affecting the innate and
adaptive immune responses (55).
Stress-induced immune responses can be categorized as
immune-protective, immune-pathological and immune-inhibitory.
Notably, stress can be categorised as good or bad, based on the
duration of the biological stress response (11). It is well known that acute stress
can enhance immune function, whereas chronic stress can suppress
it, as well as increasing the susceptibility to cancer (1). Stress can have beneficial and harmful
effects, depending on the type of immune response; factors that
determine the effects of stress on immune function include duration
(acute or chronic), endogenous versus synthetic GCs and time of
stressor (at early stages or late stages of the immune response)
(1,56) (Fig.
1). It has been reported that chronic stress can affect
individual components of the cellular immune system and
downregulate the cellular immune response (57,58),
which manifests in the form of a significant decrease in body
weight and lymphatic organs (spleen, thymus and axillary lymph
nodes) and a significant increase in the apoptotic cell count in
all lymphatic organs (59).
Chronic stress may influence the immune function and promote tumor
growth by depressing T-cell-mediated immunity and reducing the
lymphocyte count, which may depend on toll-like receptor 9 and
β-arrestin 2 (60,61). Cytotoxic T lymphocytes (CTLs) are
capable of secreting cytokines, such as interferon-γ (IFN-γ), and
other effector molecules that serve a role in immune surveillance
against tumor cells and the eradication of cancer stem cells
(62), but NE and GC can decrease
the number of CTLs or impair their function to attenuate their
antitumor effect (63). However, a
previous study found that psychological stress could augment immune
response (64). With regards to
the effect on non-specific cellular immunity, previous studies have
demonstrated that chronic stress can weaken natural killer (NK)
cell function and inhibit their activity, promoting tumor
progression (65). Stress can also
stimulate macrophages or monocytes to secrete interleukin (IL)-1,
IL-6 and tumor necrosis factor (TNF)-α, but reduce the secretion of
IL-2, IFN-γ and major histocompatibility complex (MHC), which may
aid tumor cells to evade immune surveillance (66,67).
Tumor associated-macrophages (TAMs) are one of the components of
the tumor microenvironment in which stress can increase tumor
infiltration by macrophages, and macrophage infiltration can
mediate stress-enhanced metastasis in primary breast cancer
(68). GC can induce macrophage
phenotype changes from M1 to M2 to promote tumor progression
(69). Daily restraint stress can
also lead to increased monocyte chemoattractant protein-1 (MCP-1)
expression and infiltration of cluster of differentiation
(CD)14+ and CD68+ cells. In addition, the
elevation of peripheral blood monocytes and TAMs has been
associated with a worse progression-free survival time in patients
with ovarian cancer (70). The
matricellular protein thrombospondin-1 response to stress via the
cluster of differentiation 47 (CD47) and thrombospondin-1/CD47
signaling pathways serves an important role in tumor angiogenesis
(71). The upregulated CD47
protein on the surface of cancer cells can combine with signal
regulatory protein-α, located on the surface of macrophages, in
order to avoid phagocytosis. The expression of β2-microglobulin (a
component of MHC class I molecules) in cancer cells directly
protects them from phagocytosis, which is mediated by the
inhibitory receptor leukocyte immunoglobulin-like receptor B1,
whose expression is upregulated on the surface of macrophages
(72). Dendritic cells (DCs) are
specialized antigen-presenting cells that have a direct cytotoxic
effect on tumor cells (73), but
restraint stress compromises the suppressor function of regulatory
T cells (Tregs) and alters DCs to contribute to intestinal
inflammation (74). A short-term,
stress-induced increase in IFN-γ, macrophage inflammatory
protein-3α, TNF-α, MCP-1, IL-1α, IL-1β and IL-6 can enhance the
immunization phase of cell-mediated immunity (75,76).
Nevertheless, chronic stress can lead to a reduction in the number
of cytokines, including IFN-γ, IL-2 and IL-12, due to the
inhibitory effect of GC, NE and E, which have an important
antitumor function (77,78). Chronic stress-induced
neuroendocrine changes have been found to suppress the immune
response, including NK cell cytotoxicity, phagocytosis,
inflammatory cytokine production and cytotoxic T-cell activity,
compromising the most important effectors of the immune response
against tumors (66). It is worth
highlighting that, as the largest organ and the body’s first line
of defence, cutaneous cell-mediated immunity serves an important
role in the elimination of immune-responsive tumors such as
squamous cell carcinoma (24).
Short-term stress not only enhances primary cutaneous immune
responses, but also augments secondary or recall responses in the
skin (79). In addition, the
lymphatic system serves an important role in immune function, while
also contributing to tumor cell invasion and metastasis. Studies
have found that chronic stress can promote tumor cell dissemination
by remodeling the lymph vasculature (80,81).
Effects of stress on the gut microbiota. The gut
microbiota serves an important role in maintaining gut homeostasis
(82). The most common bacterial
phyla in the gut include the Firmicutes, Bacteroidetes,
Actinobacteria and Proteobacteria, and the composition of the gut
microbiota is highly sensitive to chronic stress (83,84).
Furthermore, stress during the perinatal period can markedly
influence the microbiota, leading to long-lasting immunological
aberrations (85). It has been
demonstrated that microbiota dysbiosis is closely associated with
the occurrence, development and treatment of cancer (86,87).
Certain bioactive substances of the microbiota metabolite
production have also been found to be involved in carcinogenesis
(88). The influence of the gut
microbiota on tumorigenesis and development is mainly through
several methods that include direct contact with the tumor
(89), affecting the tumor cells
by regulating body metabolism indirectly (90,91)
and promoting tumor progression by regulating the immune system
(92,93). Studies performed on a restraint
stress mouse model suggested that commensal microbiota can affect
the postnatal development of the HPA stress response (94), and that gut dysbiosis is associated
with brain dysfunction and stress-related behavior, including
anxiety and depression (95,96).
It was shown that the absence of the gut microbiota enhanced
anxiety-like behavior and neuroendocrine response to acute stress
in rats (97). Chronic stress can
not only cause an imbalance in and disorders of the gut microbiota,
but also behavioral, cognitive and biochemical aberrations, which
may be involved in the microbiotagut-brain axis. A study revealed
that Lactobacillus helveticus NS8 can improve chronic
restraint stress-induced behavioral and cognitive dysfunction
(98). Rifaximin can alter the
bacterial population in the ileum of rats and lead to a relative
abundance of Lactobacillus, which can prevent intestinal
abnormalities and visceral hyperalgesia in response to chronic
psychological stress (99). A
study showed that probiotic treatment attenuated the HPA response
to acute stress (100). Notably,
a recent study demonstrated that dietary bioactive compounds and
probiotics could reduce the risk of colon cancer by shaping
functional gut microbiota (101).
In addition, resident gut bacteria can affect patient response to
cancer immunotherapy, and maintaining a healthy gut flora could
assist patients in their fight against cancer (102). Based on the aforementioned
literature, we tentatively conclude that there may be an
association of mutual causality between tumorigenesis and gut
microbiota imbalance.
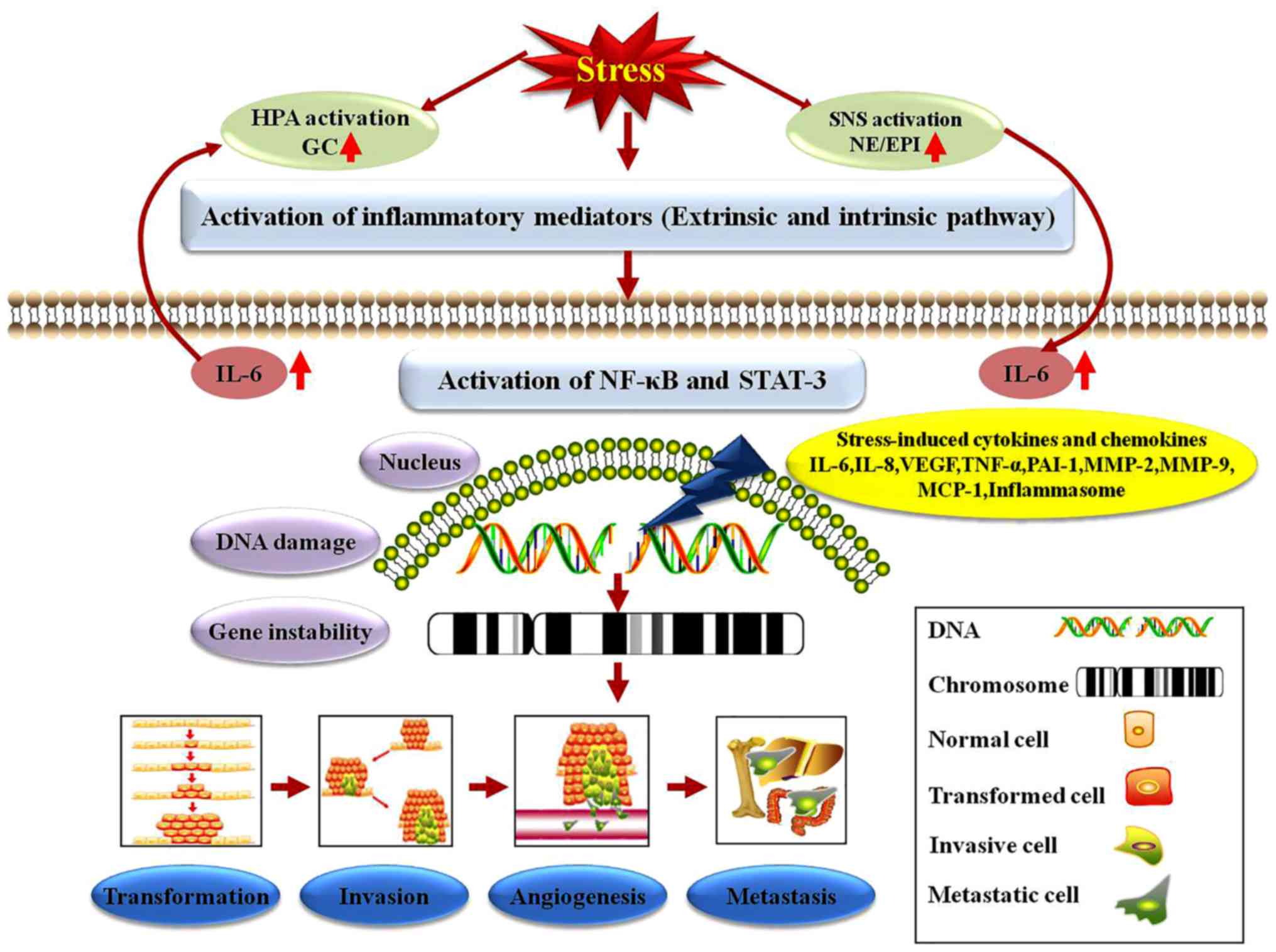
Effects of stress on inflammation. Approximately 25%
of all cancer types are associated with chronic inflammations of
broad origin (103). Just as
stress can be divided into acute and chronic according to its
beneficial or detrimental effects on the body, so can inflammation.
It has been shown that chronic inflammatory processes affect all
stages of tumor development, as well as the efficacy of therapy,
particularly in gastric, hepatic and colorectal cancer (104,105). There are two major signaling
pathways underlying cancer-related inflammation: The transcription
factor nuclear factor-κB (NF-κB) and the signal transducer and
activator of transcription 3 signaling pathways, which can be
activated by the majority of cancer risk factors, including stress,
diet, infectious agents and environmental pollutants (106). Studies have also revealed that
inflammatory cells can be recruited by stress to tumor sites,
increasing the formation of blood vessels (107). Meanwhile, stress-inducible
inflammatory factors and genes, including IL-6, IL-8 and vascular
endothelial growth factor, are increased in the circulation
following stressor exposure. Stress can lead to metastatic invasion
and metabolic syndrome by the activation of the HPA axis and SNS,
respectively, which are characterized by an increased production of
IL-6, TNF-α, llasminogen activator inhibitor-1 and
metalloproteinase-2 and -9 (108). Reciprocally, adipose-derived IL-6
may further stimulate the HPA axis, forming a deleterious vicious
cycle (109). Chronic
psychological stress can induce vascular inflammation via the TNF-α
and p38/c-Jun N-terminal kinase pathways and increase the
expression of inflammatory molecules, including mRNA and proteins
such as TNF-α, C-reactive protein, MCP-1, macrophage migration
inhibitory factor and intercellular adhesion molecule-1 (110). Psychological stress-derived
prolactin has been shown not only to induce IL-6 and IL-23
production by DCs, the former of which serve a critical role in
altering the phenotypes of Tregs, but also to alter Treg
properties, leading to intestinal inflammation (75). Chronic restraint stress has been
shown to result in a marked decrease in CD4 T cell numbers and
intracellular IFN-γ expression, while increasing IL-4 production.
It was also found that, in chronically stressed mice, treatment
with 4-methylhistamine (4-MeH) agonist was able to restore the
immune response, particularly via the production of Th1 cytokines.
Stimulation of the histamine 4 receptor with 4-MeH modulates the
effects of chronic stress on the Th1/Th2 cytokine balance (111). The sympathetic and neuroendocrine
responses to psychosocial stress have been shown to have a
significant impact on cancer, partly through the regulation of
inflammatory mediators (112).
Psychological stress increases extracellular adenosine triphosphate
(ATP), IL-1β and TNF-α in the hippocampus, and activates the
inflammasomes via the release of ATP and the stimulation of the
purinergic type 2X7 receptor (16). Inflammasomes are multiprotein
complexes that operate as platforms for the activation of caspase-1
and can be categorized based on their main constituent as either
NLR family pyrin domain containing 1 (NLRP1), NLRP3, NLR family
CARD domain containing 4, NLRP6 or absent in melanoma 2. The
activation of inflammasomes can lead to the conversion of inactive
inflammation mediators to active ones (IL-1β and IL-18), and,
subsequently, the active inflammation mediators are secreted to the
cell exterior to modulate cell function in an autocrine or
paracrine manner. This process may mechanistically explain the link
between inflammasome activation and tumorigenesis, angiogenesis or
metastasis (113,114). Furthermore, the external IL-1β
can initiate self-reinforcing feedback loops to further perpetuate
its existence through the IL-1R-MyD88-NF-κB pathway by inflammasome
activators (115). Based on the
aforementioned findings, the inhibition of inflammasomes or
neutralization of their products can have profound effects on
carcinogenesis and tumor progression. The stress hormones NE and
Epi can enhance IL-8 expression and thereby mediate the effects of
stress on the growth and metastasis of ovarian cancer (116). IL-8 gene silencing with liposomal
small interfering RNA incorporated in
1,2-dioleoyl-sn-glycero-3-phosphocholine has been shown to decrease
tumor growth and angiogenesis in ovarian cancer. In addition, the
increase of pro-inflammatory cytokines has been associated with
irritability, insomnia and fatigue, which, in turn, are associated
with cancer (117) (Fig. 3).
Effects of stress on chemo-resistance (multidrug
resistance). At present, chemotherapy is one of the main treatment
strategies for malignant tumors. However, a number of
chemotherapeutic approaches fail due to intrinsic or acquired drug
resistance, particularly multidrug resistance. Multiple studies
have focused on the mechanisms underlying multidrug resistance in
cancer, including DNA damage repair (118) and abnormalities in the expression
and function of transporters of the ATP-binding cassette
superfamily. P-glycoprotein is the member of this mechanism that
has been most frequently reported in association with tumor drug
resistance, cancer stem cell, epithelial-mesenchymal transitions
and hypoxia (119,120). A study on mice showed that
psychological stress reduced the antitumor effects of
chemotherapeutic drugs and induced chemo-resistance in breast
cancer by upregulating multidrug resistance protein 1 via
adrenergic stimulation (121).
Stress hormones induced by restraint stress, including cortisol and
epinephrine, can decrease the efficacy of paclitaxel in
triple-negative breast cancer through the induction of DNA damage
and ATR serine/threonine kinase and p21 expression (122). Adrenaline can induce
chemo-resistance in HT-29 colon adenocarcinoma cells by
upregulating the ATP binding cassette subfamily B member 1 gene
expression via α2-adrenergic receptors (123), as well as cisplatin resistance
through the activation of the NF-κB pathway and subsequent
induction of miR-155 (124). One
study showed that miR-155 is responsible for the drug resistance in
breast cancer cells, by targeting fork-head box O3 (125). Meanwhile, injections of
epinephrine or immobilization stress can counteract the antitumor
effects of phosphatidylinositol-4,5-bisphosphate 3-kinase
inhibitors on prostate cancer xenografts in mice (126). A dynamic network model of
apoptosis regulation in prostate cancer indicated that
psychological stress could trigger a synergism pattern switch in
drug combination therapy (127).
However, it is important to note that chemotherapy itself may
generate or deteriorate psychological stress, which is known to be
accompanied by chronic elevation of plasma catecholamine,
conversely leading to chemo-resistance (121). Psychological stress has been
found to attenuate the anti-angiogenic efficacy of sunitinib
through the activation of β-adrenergic signaling and the promotion
of tumor angiogenesis, which, however, can be improved by the use
of β-blockers (39). Stress
hormones have also been shown to promote resistance to epidermal
growth factor receptor (EGFR) inhibitor in non-small cell lung
cancer, which can be abrogated by combinations of β-blockers and
EGFR tyrosine kinase inhibitors (53). Furthermore, certain gut microbiota,
such as Escherichia coli strains, can decrease the efficacy
of chemotherapeutic agent gemcitabine by metabolizing and
deactivating the active form of the drug (128).
Stress is one of the fundamental survival mechanisms
in nature; it begins with a stressor, leading to stress perception
and subsequently resulting in a stress response. Stress can be
divided into acute and chronic, mainly according to its respective
salubrious or detrimental effects on the health of an individual.
Several studies have demonstrated that stress, particularly chronic
stress, serves an important role in tumorigenesis and metastasis.
Relevant and effective animal models are essential for the study of
the effects of stress on cancer and the underlying mechanisms. The
induction of CMS or chronic restraint stress has been widely used
for the development of experimental models. The stress-induced
persistent activation of the HPA axis and SNS has been shown to
result in a cascade reaction leading to cross-talk among the
neuroendocrine system, the immune system, gut microbiota and
inflammation. Further elucidation of the influence of the
interactions among these factors in mediating stress-associated
effects on tumorigenesis and metastasis is imperative. Novel
approaches to the prevention and blockage of the harmful effects of
stress on tumorigenesis and metastasis are required.
Stress and stress-associated disorders have become
prevalent in modern societies due to the fast-paced nature of
contemporary lifestyles. Due to their prevalence, such disorders
are often concealed, which has detrimental effects to health.
Studies have demonstrated that exposure to stress during critical
periods in human development is sufficient for it to have severe,
long-term consequences (35,129). In addition, chronic stress may
lead to epigenetic heritable modifications, suggesting a possible
propagation across generations (55). It is noteworthy that stress may
exist universally during the diagnosis and subsequent treatment
process in patients with cancer (130). On a positive note, it has been
shown that effective intervention in cancer patients presenting
with stress can improve their immune function and physical
activity, and social support can modulate cancer-related pathways
and improve the levels of certain biomarkers associated with a
better prognosis and longer survival (131). Studies have also revealed that
disturbances in mood, anxiety and irritability may precede the
appearance of a medical disorder, and that stress-prone
personalities or unfavorable coping mechanisms and negative
emotional responses are associated with a higher incidence, poorer
survival and higher mortality rates in patients with cancer
(132). Dhabhar (11) proposed the stress spectrum model
and concept, so as to reconcile the potentially beneficial effect
of stress with the harmful; according to the stress spectrum model,
one can keep fit by minimizing chronic stress, maximizing the
resting zone of low/no stress and optimizing the acute stress
response.
In the research findings discussed within the
present review, it was demonstrated that effective stress
prevention and management are plausible and imperative, and that
the primary, secondary and tertiary prevention strategies should
receive adequate recognition. In particular, moderate exercise, a
healthy diet, high-quality sleep and emotional management are key
aspects that can be improved in the lives of patients with
stress-induced cancer, so that they benefit from them. Regular
exercise can assist in keeping the short-term stress response
well-oiled, finely-tuned and ready for fight-or-flight (11), as well as improving the physical
and emotional well-being of an individual. Furthermore, regular
exercise has also been shown to reduce the risk for breast cancer
recurrence (133). The type,
intensity, duration and frequency of exercise should be tailored to
the constitution an individual. A number of traditional methods of
exercise, including five-animal boxing, eight-section brocade,
changing tendon exercise, meditation and yoga, may contribute to
successful stress management.
The present review found that diet and gut
microbiota play an important role in tumorigenesis and metastasis.
Therefore, closer attention should be paid to dietary strategies,
and particularly the maintenance of healthy-eating habits.
Furthermore, developing effective microbial agents is necessary and
promising with regard to the reduction of cancer risks. An
association has been identified between lack of sleep or sleep
disturbances such as insomnia, fatigue, obstructive sleep apnea and
restless legs syndrome, and depression or anxiety (134). The latter are also common among
cancer patients, and often comprise the symptoms that lead to the
diagnosis of certain types of cancer (135). In addition, circadian cortisol
rhythm disruptions serve as an important indicator and/or mediator
of the deleterious effects of chronic stress (1). In view of this, future studies are
warranted to better understand the mechanisms underlying the
effects of stress on tumorigenesis and metastasis.
The present review was supported by the National
Natural Science Foundation of China (grant nos. 81573764 and
81774095), the Program for Outstanding Medical Academic Leader,
Shanghai Academic/Technology Research Leader (grant no.
16XD1403600), the Shanghai Rising-Star Program (grant no.
16QA1403700) and the Municipal Human Resources Development Program
for Outstanding Leaders in Medical Disciplines in Shanghai (grant
no. 2017BR031).
Not applicable.
QL and YW put forward the conception and design of
the manuscript, and ZZZ and YW were major contributors in writing
the manuscript. All authors read and approved the final
manuscript.
Not applicable.
Not applicable.
The authors declare that they have no competing
interests.
The authors would like to thank Professor Junfeng
Zhang (Duke University, Durham, NC, USA) for the revision of the
original manuscript.
|
1
|
Dhabhar FS and McEwen BS: Acute stress
enhances while chronic stress suppresses cell-mediated immunity in
vivo: A potential role for leukocyte trafficking. Brain Behav
Immun. 11:286–306. 1997. View Article : Google Scholar
|
|
2
|
Amin SN, El-Aidi AA, Ali MM, Attia YM and
Rashed LA: Modification of hippocampal markers of synaptic
plasticity by memantine in animal models of acute and repeated
restraint stress: Implications for memory and behavior.
Neuromolecular Med. 17:121–136. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Charmandari E, Tsigos C and Chrousos G:
Endocrinology of the stress response. Annu Rev Physiol. 67:259–284.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Gunnar M and Quevedo K: The neurobiology
of stress and development. Annu Rev Psychol. 58:145–173. 2007.
View Article : Google Scholar
|
|
5
|
Krizanova O, Babula P and Pacak K: Stress,
catecholaminergic system and cancer. Stress. 19:419–428. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Hering D, Lachowska K and Schlaich M: Role
of the sympathetic nervous system in stress-mediated cardiovascular
disease. Curr Hypertens Rep. 17:802015. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Dhabhar FS, McEwen BS and Spencer RL:
Stress response, adrenal steroid receptor levels and
corticosteroid-binding globulin levels - a comparison between
Sprague-Dawley, Fischer 344 and Lewis rats. Brain Res. 616:89–98.
1993. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Dhabhar FS, McEwen BS and Spencer RL:
Adaptation to prolonged or repeated stress - comparison between rat
strains showing intrinsic differences in reactivity to acute
stress. Neuroendocrinology. 65:360–368. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Ambarish V, Chandrashekara S and Suresh
KP: Moderate regular exercises reduce inflammatory response for
physical stress. Indian J Physiol Pharmacol. 56:7–14.
2012.PubMed/NCBI
|
|
10
|
Clague J and Bernstein L: Physical
activity and cancer. Curr Oncol Rep. 14:550–558. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Dhabhar FS: Effects of stress on immune
function: The good, the bad, and the beautiful. Immunol Res.
58:193–210. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Lagraauw HM, Kuiper J and Bot I: Acute and
chronic psychological stress as risk factors for cardiovascular
disease: Insights gained from epidemiological, clinical and
experimental studies. Brain Behav Immun. 50:18–30. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Muffly LS, Hlubocky FJ, Khan N, Wroblewski
K, Breitenbach K, Gomez J, McNeer JL, Stock W and Daugherty CK:
Psychological morbidities in adolescent and young adult blood
cancer patients during curative-intent therapy and early
survivorship. Cancer. 122:954–961. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Cohen L, Cole SW, Sood AK, Prinsloo S,
Kirschbaum C, Arevalo JM, Jennings NB, Scott S, Vence L, Wei Q, et
al: Depressive symptoms and cortisol rhythmicity predict survival
in patients with renal cell carcinoma: Role of inflammatory
signaling. PLoS One. 7:e423242012. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Shan T, Ma J, Ma Q, Guo K, Guo J, Li X, Li
W, Liu J, Huang C, Wang F, et al: β2-AR-HIF-1α: A novel regulatory
axis for stress-induced pancreatic tumor growth and angiogenesis.
Curr Mol Med. 13:1023–1034. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Iwata M, Ota KT, Li XY, Sakaue F, Li N,
Dutheil S, Banasr M, Duric V, Yamanashi T, Kaneko K, et al:
Psychological stress activates the inflammasome via release of
adenosine triphosphate and stimulation of the purinergic type 2X7
receptor. Biol Psychiatry. 80:12–22. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Agarwal SK and Marshall GD Jr: Stress
effects on immunity and its application to clinical immunology.
Clin Exp Allergy. 31:25–31. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Yin X, Guven N and Dietis N: Stress-based
animal models of depression: Do we actually know what we are doing?
Brain Res. 1652:30–42. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Heinrichs SC and Koob GF: Application of
experimental stressors in laboratory rodents. Curr Protoc Neurosci.
Chapter 8: Unit8.4. 2006. View Article : Google Scholar
|
|
20
|
Zorzet S, Perissin L, Rapozzi V and
Giraldi T: Restraint stress reduces the antitumor efficacy of
cyclophosphamide in tumor-bearing mice. Brain Behav Immun.
12:23–33. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Nukina H, Sudo N, Aiba Y, Oyama N, Koga Y
and Kubo C: Restraint stress elevates the plasma interleukin-6
levels in germ-free mice. J Neuroimmunol. 115:46–52. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Willner P: The validity of animal models
of predisposition to depression. Behav Pharmacol. 13:169–188. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Glaser R and Kiecolt-Glaser JK:
Stress-induced immune dysfunction: Implications for health. Nat Rev
Immunol. 5:243–251. 2005. View
Article : Google Scholar : PubMed/NCBI
|
|
24
|
Dhabhar FS, Saul AN, Daugherty C, Holmes
TH, Bouley DM and Oberyszyn TM: Short-term stress enhances cellular
immunity and increases early resistance to squamous cell carcinoma.
Brain Behav Immun. 24:127–137. 2010. View Article : Google Scholar
|
|
25
|
Eng JW, Kokolus KM, Reed CB, Hylander BL,
Ma WW and Repasky EA: A nervous tumor microenvironment: The impact
of adrenergic stress on cancer cells, immunosuppression, and
immunotherapeutic response. Cancer Immunol Immunother.
63:1115–1128. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Frick LR, Rapanelli M, Bussmann UA, Klecha
AJ, Arcos ML, Genaro AM and Cremaschi GA: Involvement of thyroid
hormones in the alterations of T-cell immunity and tumor
progression induced by chronic stress. Biol Psychiatry. 65:935–942.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Volpi S, Rabadan-Diehl C and Aguilera G:
Vasopressinergic regulation of the hypothalamic pituitary adrenal
axis and stress adaptation. Stress. 7:75–83. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Meltzer HY, Lowy MT and Koenig JI: The
hypothalamic-pituitary-adrenal axis in depression. Adv Biochem
Psychopharmacol. 43:165–182. 1987.PubMed/NCBI
|
|
29
|
Chrousos GP: Ultradian, circadian, and
stress-related hypothalamic-pituitary-adrenal axis activity - a
dynamic digital-to-analog modulation. Endocrinology. 139:437–440.
1998. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Sephton S and Spiegel D: Circadian
disruption in cancer: A neuroendocrine-immune pathway from stress
to disease? Brain Behav Immun. 17:321–328. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
McEwen BS: Physiology and neurobiology of
stress and adaptation: Central role of the brain. Physiol Rev.
87:873–904. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Moreno-Smith M, Lutgendorf SK and Sood AK:
Impact of stress on cancer metastasis. Future Oncol. 6:1863–1881.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Gündisch S, Boeckeler E, Behrends U,
Amtmann E, Ehrhardt H and Jeremias I: Glucocorticoids augment
survival and proliferation of tumor cells. Anticancer Res.
32:4251–4261. 2012.PubMed/NCBI
|
|
34
|
Wang HM, Liao ZX, Komaki R, Welsh JW,
O’Reilly MS, Chang JY, Zhuang Y, Levy LB, Lu C and Gomez DR:
Improved survival outcomes with the incidental use of beta-blockers
among patients with non-small-cell lung cancer treated with
definitive radiation therapy. Ann Oncol. 24:1312–1319. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Wulsin AC, Wick-Carlson D, Packard BA,
Morano R and Herman JP: Adolescent chronic stress causes
hypothalamo-pituitary-adrenocortical hypo-responsiveness and
depression-like behavior in adult female rats.
Psychoneuroendocrinology. 65:109–117. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Bortolato B, Hyphantis TN, Valpione S,
Perini G, Maes M, Morris G, Kubera M, Köhler CA, Fernandes BS,
Stubbs B, et al: Depression in cancer: The many biobehavioral
pathways driving tumor progression. Cancer Treat Rev. 52:58–70.
2017. View Article : Google Scholar
|
|
37
|
Zhao L, Xu J, Liang F, Li A, Zhang Y and
Sun J: Effect of chronic psychological stress on liver metastasis
of colon cancer in mice. PLoS One. 10:e01399782015. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Xie H, Li C, He Y, Griffin R, Ye Q and Li
L: Chronic stress promotes oral cancer growth and angiogenesis with
increased circulating catecholamine and glucocorticoid levels in a
mouse model. Oral Oncol. 51:991–997. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Liu J, Deng GH, Zhang J, Wang Y, Xia XY,
Luo XM, Deng YT, He SS, Mao YY, Peng XC, et al: The effect of
chronic stress on anti-angiogenesis of sunitinib in colorectal
cancer models. Psychoneuroendocrinology. 52:130–142. 2015.
View Article : Google Scholar
|
|
40
|
Elefteriou F: Chronic stress, sympathetic
activation and skeletal metastasis of breast cancer cells. Bonekey
Rep. 4:6932015. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Moreno-Smith M, Lu C, Shahzad MM, Pena GN,
Allen JK, Stone RL, Mangala LS, Han HD, Kim HS, Farley D, et al:
Dopamine blocks stress-mediated ovarian carcinoma growth. Clin
Cancer Res. 17:3649–3659. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Moreno-Smith M, Lee SJ, Lu C, Nagaraja AS,
He G, Rupaimoole R, Han HD, Jennings NB, Roh JW, Nishimura M, et
al: Biologic effects of dopamine on tumor vasculature in ovarian
carcinoma. Neoplasia. 15:502–510. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Borcherding DC, Tong W, Hugo ER, Barnard
DF, Fox S, LaSance K, Shaughnessy E and Ben-Jonathan N: Expression
and therapeutic targeting of dopamine receptor-1 (D1R) in breast
cancer. Oncogene. 35:3103–3113. 2016. View Article : Google Scholar
|
|
44
|
Peters MA, Walenkamp AM, Kema IP, Meijer
C, de Vries EG and Oosting SF: Dopamine and serotonin regulate
tumor behavior by affecting angiogenesis. Drug Resist Updat.
17:96–104. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Barbieri A, Palma G, Rosati A, Giudice A,
Falco A, Petrillo A, Petrillo M, Bimonte S, Di Benedetto M,
Esposito G, et al: Role of endothelial nitric oxide synthase (eNOS)
in chronic stress-promoted tumour growth. J Cell Mol Med.
16:920–926. 2012. View Article : Google Scholar
|
|
46
|
Partecke LI, Speerforck S, Käding A,
Seubert F, Kühn S, Lorenz E, Schwandke S, Sendler M, Kessler W,
Trung DN, et al: Chronic stress increases experimental pancreatic
cancer growth, reduces survival and can be antagonised by
beta-adrenergic receptor blockade. Pancreatology. 16:423–433. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Feng Z, Liu L, Zhang C, Zheng T, Wang J,
Lin M, Zhao Y, Wang X, Levine AJ and Hu W: Chronic restraint stress
attenuates p53 function and promotes tumorigenesis. Proc Natl Acad
Sci USA. 109:7013–7018. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Gao J, Gao G, Zhang Y and Wang F:
Proteomic analysis of human epithelial ovarian cancer xenografts in
immunodeficient mice exposed to chronic psychological stress. Sci
China Life Sci. 54:112–120. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Turnbull AV and Rivier CL: Regulation of
the hypothalamic-pituitary-adrenal axis by cytokines: Actions and
mechanisms of action. Physiol Rev. 79:1–71. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Schwab CL, Fan R, Zheng Q, Myers LP,
Hebert P and Pruett SB: Modeling and predicting stress-induced
immunosuppression in mice using blood parameters. Toxicol Sci.
83:101–113. 2005. View Article : Google Scholar
|
|
51
|
Dhabhar FS, Malarkey WB, Neri E and McEwen
BS: Stress-induced redistribution of immune cells - from barracks
to boulevards to battlefields: A tale of three hormones - Curt
Richter Award winner. Psychoneuroendocrinology. 37:1345–1368. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Barbieri A, Bimonte S, Palma G, Luciano A,
Rea D, Giudice A, Scognamiglio G, La Mantia E, Franco R, Perdonà S,
et al: The stress hormone norepinephrine increases migration of
prostate cancer cells in vitro and in vivo. Int J Oncol.
47:527–534. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Nilsson MB, Sun H, Diao L, Tong P, Liu D,
Li L, Fan Y, Poteete A, Lim SO, Howells K, et al: Stress hormones
promote EGFR inhibitor resistance in NSCLC: Implications for
combinations with β-blockers. Sci Transl Med. 9:92017. View Article : Google Scholar
|
|
54
|
Kim-Fuchs C, Le CP, Pimentel MA,
Shackleford D, Ferrari D, Angst E, Hollande F and Sloan EK: Chronic
stress accelerates pancreatic cancer growth and invasion: A
critical role for beta-adrenergic signaling in the pancreatic
microenvironment. Brain Behav Immun. 40:40–47. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Nagaraja AS, Sadaoui NC, Dorniak PL,
Lutgendorf SK and Sood AK: SnapShot: Stress and Disease. Cell
Metab. 23:388–388.e1. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Dhabhar FS: Enhancing versus suppressive
effects of stress on immune function: Implications for
immunoprotection and immunopathology. Neuroimmunomodulation.
16:300–317. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Glaser R, MacCallum RC, Laskowski BF,
Malarkey WB, Sheridan JF and Kiecolt-Glaser JK: Evidence for a
shift in the Th-1 to Th-2 cytokine response associated with chronic
stress and aging. J Gerontol A Biol Sci Med Sci. 56:M477–M482.
2001. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Ochoa CE, Mirabolfathinejad SG, Ruiz VA,
Evans SE, Gagea M, Evans CM, Dickey BF and Moghaddam SJ:
Interleukin 6, but not T helper 2 cytokines, promotes lung
carcinogenesis. Cancer Prev Res (Phila). 4:51–64. 2011. View Article : Google Scholar
|
|
59
|
Divyashree S, Sarjan HN and Yajurvedi HN:
Effects of long-term chronic stress on the lymphoid organs and
blood l. Can J Zool. 94:137–143. 2015. View Article : Google Scholar
|
|
60
|
Frick LR, Arcos ML, Rapanelli M, Zappia
MP, Brocco M, Mongini C, Genaro AM and Cremaschi GA: Chronic
restraint stress impairs T-cell immunity and promotes tumor
progression in mice. Stress. 12:134–143. 2009. View Article : Google Scholar
|
|
61
|
Li H, Zhao J, Chen M, Tan Y, Yang X,
Caudle Y and Yin D: Toll-like receptor 9 is required for chronic
stress-induced immune suppression. Neuroimmunomodulation. 21:1–7.
2014. View Article : Google Scholar :
|
|
62
|
Lakshmi Narendra B, Eshvendar Reddy K,
Shantikumar S and Ramakrishna S: Immune system: a double-edged
sword in cancer. Inflamm Res. 62:823–834. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Hunzeker JT, Elftman MD, Mellinger JC,
Princiotta MF, Bonneau RH, Truckenmiller ME and Norbury CC: A
marked reduction in priming of cytotoxic CD8+ T cells
mediated by stress-induced glucocorticoids involves multiple
deficiencies in cross-presentation by dendritic cells. J Immunol.
186:183–194. 2011. View Article : Google Scholar
|
|
64
|
Kour K and Bani S: Augmentation of immune
response by chicoric acid through the modulation of CD28/CTLA-4 and
Th1 pathway in chronically stressed mice. Neuropharmacology.
60:852–860. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Lutgendorf SK, Sood AK, Anderson B, McGinn
S, Maiseri H, Dao M, Sorosky JI, De Geest K, Ritchie J and Lubaroff
DM: Social support, psychological distress, and natural killer cell
activity in ovarian cancer. J Clin Oncol. 23:7105–7113. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Reiche EM, Nunes SO and Morimoto HK:
Stress, depression, the immune system, and cancer. Lancet Oncol.
5:617–625. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Nakatani Y, Amano T and Takeda H:
Corticosterone suppresses the proliferation of RAW264.7 macrophage
cells via glucocor-ticoid, but not mineralocorticoid, receptor.
Biol Pharm Bull. 36:592–601. 2013. View Article : Google Scholar
|
|
68
|
Sloan EK, Priceman SJ, Cox BF, Yu S,
Pimentel MA, Tangkanangnukul V, Arevalo JM, Morizono K, Karanikolas
BD, Wu L, et al: The sympathetic nervous system induces a
metastatic switch in primary breast cancer. Cancer Res.
70:7042–7052. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Schmieder A, Michel J, Schönhaar K, Goerdt
S and Schledzewski K: Differentiation and gene expression profile
of tumor-associated macrophages. Semin Cancer Biol. 22:289–297.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Armaiz-Pena GN, Gonzalez-Villasana V,
Nagaraja AS, Rodriguez-Aguayo C, Sadaoui NC, Stone RL, Matsuo K,
Dalton HJ, Previs RA, Jennings NB, et al: Adrenergic regulation of
monocyte chemotactic protein 1 leads to enhanced macrophage
recruitment and ovarian carcinoma growth. Oncotarget. 6:4266–4273.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Roberts DD, Miller TW, Rogers NM, Yao M
and Isenberg JS: The matricellular protein thrombospondin-1
globally regulates cardiovascular function and responses to stress
via CD47. Matrix Biol. 31:162–169. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Barkal AA, Weiskopf K, Kao KS, Gordon SR,
Rosental B, Yiu YY, George BM, Markovic M, Ring NG, Tsai JM, et al:
Engagement of MHC class I by the inhibitory receptor LILRB1
suppresses macrophages and is a target of cancer immunotherapy. Nat
Immunol. 19:76–84. 2018. View Article : Google Scholar :
|
|
73
|
Hanke N, Alizadeh D, Katsanis E and
Larmonier N: Dendritic cell tumor killing activity and its
potential applications in cancer immunotherapy. Crit Rev Immunol.
33:1–21. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Wu W, Sun M, Zhang HP, Chen T, Wu R, Liu
C, Yang G, Geng XR, Feng BS, Liu Z, et al: Prolactin mediates
psychological stress-induced dysfunction of regulatory T cells to
facilitate intestinal inflammation. Gut. 63:1883–1892. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Dhabhar FS, Satoskar AR, Bluethmann H,
David JR and McEwen BS: Stress-induced enhancement of skin immune
function: A role for gamma interferon. Proc Natl Acad Sci USA.
97:2846–2851. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
76
|
Dhabhar FS and Viswanathan K: Short-term
stress experienced at time of immunization induces a long-lasting
increase in immunologic memory. Am J Physiol Regul Integr Comp
Physiol. 289:R738–R744. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
77
|
Levi B, Benish M, Goldfarb Y, Sorski L,
Melamed R, Rosenne E and Ben-Eliyahu S: Continuous stress disrupts
immunostimulatory effects of IL-12. Brain Behav Immun. 25:727–735.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
78
|
Lopes RP, Grassi-Oliveira R, de Almeida
LR, Stein LM, Luz C, Teixeira AL and Bauer ME: Neuroimmunoendocrine
interactions in patients with recurrent major depression, increased
early life stress and long-standing posttraumatic stress disorder
symptoms. Neuroimmunomodulation. 19:33–42. 2012. View Article : Google Scholar
|
|
79
|
Dhabhar FS and McEwen BS: Stress-induced
enhancement of antigen-specific cell-mediated immunity. J Immunol.
156:2608–2615. 1996.PubMed/NCBI
|
|
80
|
Stacker SA, Williams SP, Karnezis T,
Shayan R, Fox SB and Achen MG: Lymphangiogenesis and lymphatic
vessel remodelling in cancer. Nat Rev Cancer. 14:159–172. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
81
|
Le CP, Nowell CJ, Kim-Fuchs C, Botteri E,
Hiller JG, Ismail H, Pimentel MA, Chai MG, Karnezis T, Rotmensz N,
et al: Chronic stress in mice remodels lymph vasculature to promote
tumour cell dissemination. Nat Commun. 7:106342016. View Article : Google Scholar : PubMed/NCBI
|
|
82
|
Boleij A and Tjalsma H: Gut bacteria in
health and disease: A survey on the interface between intestinal
microbiology and colorectal cancer. Biol Rev Camb Philos Soc.
87:701–730. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
83
|
O’Toole PW: Gut microbiota and aging.
Science. 350:1214–1215. 2015. View Article : Google Scholar
|
|
84
|
Rook GAW, Raison CL and Lowry CA:
Microbiota, immuno-regulatory old friends and psychiatric
disorders. Adv Exp Med Biol. 817:319–356. 2014. View Article : Google Scholar
|
|
85
|
Penders J, Gerhold K, Stobberingh EE,
Thijs C, Zimmermann K, Lau S and Hamelmann E: Establishment of the
intestinal microbiota and its role for atopic dermatitis in early
childhood. J Allergy Clin Immunol. 132:601–607.e8. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
86
|
Gagnière J, Raisch J, Veziant J, Barnich
N, Bonnet R, Buc E, Bringer MA, Pezet D and Bonnet M: Gut
microbiota imbalance and colorectal cancer. World J Gastroenterol.
22:501–518. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
87
|
Paul B, Barnes S, Demark-Wahnefried W,
Morrow C, Salvador C, Skibola C and Tollefsbol TO: Influences of
diet and the gut microbiome on epigenetic modulation in cancer and
other diseases. Clin Epigenetics. 7:1122015. View Article : Google Scholar : PubMed/NCBI
|
|
88
|
Berni Canani R, Di Costanzo M and Leone L:
The epigenetic effects of butyrate: Potential therapeutic
implications for clinical practice. Clin Epigenetics. 4:42012.
View Article : Google Scholar : PubMed/NCBI
|
|
89
|
Yamamoto ML, Maier I, Dang AT, Berry D,
Liu J, Ruegger PM, Yang JI, Soto PA, Presley LL, Reliene R, et al:
Intestinal bacteria modify lymphoma incidence and latency by
affecting systemic inflammatory state, oxidative stress, and
leukocyte genotoxicity. Cancer Res. 73:4222–4232. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
90
|
Maslowski KM, Vieira AT, Ng A, Kranich J,
Sierro F, Yu D, Schilter HC, Rolph MS, Mackay F, Artis D, et al:
Regulation of inflammatory responses by gut microbiota and
chemoattractant receptor GPR43. Nature. 461:1282–1286. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
91
|
Garrett WS: Cancer and the microbiota.
Science. 348:80–86. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
92
|
Iida N, Dzutsev A, Stewart CA, Smith L,
Bouladoux N, Weingarten RA, Molina DA, Salcedo R, Back T, Cramer S,
et al: Commensal bacteria control cancer response to therapy by
modulating the tumor microenvironment. Science. 342:967–970. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
93
|
Vétizou M, Pitt JM, Daillère R, Lepage P,
Waldschmitt N, Flament C, Rusakiewicz S, Routy B, Roberti MP, Duong
CP, et al: Anticancer immunotherapy by CTLA-4 blockade relies on
the gut microbiota. Science. 350:1079–1084. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
94
|
Sudo N, Chida Y, Aiba Y, Sonoda J, Oyama
N, Yu XN, Kubo C and Koga Y: Postnatal microbial colonization
programs the hypothalamic-pituitary-adrenal system for stress
response in mice. J Physiol. 558:263–275. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
95
|
Luna RA and Foster JA: Gut brain axis:
Diet microbiota interactions and implications for modulation of
anxiety and depression. Curr Opin Biotechnol. 32:35–41. 2015.
View Article : Google Scholar
|
|
96
|
Naseribafrouei A, Hestad K, Avershina E,
Sekelja M, Linløkken A, Wilson R and Rudi K: Correlation between
the human fecal microbiota and depression. Neurogastroenterol
Motil. 26:1155–1162. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
97
|
Crumeyrolle-Arias M, Jaglin M, Bruneau A,
Vancassel S, Cardona A, Daugé V, Naudon L and Rabot S: Absence of
the gut microbiota enhances anxiety-like behavior and
neuroendocrine response to acute stress in rats.
Psychoneuroendocrinology. 42:207–217. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
98
|
Liang S, Wang T, Hu X, Luo J, Li W, Wu X,
Duan Y and Jin F: Administration of Lactobacillus helveticus NS8
improves behavioral, cognitive, and biochemical aberrations caused
by chronic restraint stress. Neuroscience. 310:561–577. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
99
|
Xu D, Gao J, Gillilland M III, Wu X, Song
I, Kao JY and Owyang C: Rifaximin alters intestinal bacteria and
prevents stress-induced gut inflammation and visceral hyperalgesia
in rats. Gastroenterology. 146:484–96.e4. 2014. View Article : Google Scholar :
|
|
100
|
Ait-Belgnaoui A, Durand H, Cartier C,
Chaumaz G, Eutamene H, Ferrier L, Houdeau E, Fioramonti J, Bueno L
and Theodorou V: Prevention of gut leakiness by a probiotic
treatment leads to attenuated HPA response to an acute
psychological stress in rats. Psychoneuroendocrinology.
37:1885–1895. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
101
|
Seidel DV, Azcárate-Peril MA, Chapkin RS
and Turner ND: Shaping functional gut microbiota using dietary
bioactives to reduce colon cancer risk. Semin Cancer Biol.
46:191–204. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
102
|
Gopalakrishnan V, Spencer CN, Nezi L,
Reuben A, Andrews MC, Karpinets TV, Prieto PA, Vicente D, Hoffman
K, Wei SC, et al: Gut microbiome modulates response to anti-PD-1
immunotherapy in melanoma patients. Science. 359:97–103. 2018.
View Article : Google Scholar
|
|
103
|
Balkwill FR and Mantovani A:
Cancer-related inflammation: Common themes and therapeutic
opportunities. Semin Cancer Biol. 22:33–40. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
104
|
Hanahan D and Weinberg RA: Hallmarks of
cancer: The next generation. Cell. 144:646–674. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
105
|
Elinav E, Nowarski R, Thaiss CA, Hu B, Jin
C and Flavell RA: Inflammation-induced cancer: Crosstalk between
tumours, immune cells and microorganisms. Nat Rev Cancer.
13:759–771. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
106
|
Aggarwal BB, Vijayalekshmi RV and Sung B:
Targeting inflammatory pathways for prevention and therapy of
cancer: short-term friend, long-term foe. Clin Cancer Res.
15:425–430. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
107
|
Thaker PH, Han LY, Kamat AA, Arevalo JM,
Takahashi R, Lu C, Jennings NB, Armaiz-Pena G, Bankson JA, Ravoori
M, et al: Chronic stress promotes tumor growth and angiogenesis in
a mouse model of ovarian carcinoma. Nat Med. 12:939–944. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
108
|
Armaiz-Pena GN, Cole SW, Lutgendorf SK and
Sood AK: Neuroendocrine influences on cancer progression. Brain
Behav Immun. 30(Suppl): S19–S25. 2013. View Article : Google Scholar
|
|
109
|
Kyrou I, Tsigos C, Seedorf K and Ferré P:
Stress hormones: Physiological stress and regulation of metabolism.
Curr Opin Pharmacol. 9:787–793. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
110
|
Lu XT, Liu YF, Zhao L, Li WJ, Yang RX, Yan
FF, Zhao YX and Jiang F: Chronic psychological stress induces
vascular inflammation in rabbits. Stress. 16:87–98. 2013.
View Article : Google Scholar
|
|
111
|
Ahmad SF, Zoheir KM, Ansari MA, Korashy
HM, Bakheet SA, Ashour AE and Attia SM: Stimulation of the
histamine 4 receptor with 4-methylhistamine modulates the effects
of chronic stress on the Th1/Th2 cytokine balance. Immunobiology.
220:341–349. 2015. View Article : Google Scholar
|
|
112
|
Powell ND, Tarr AJ and Sheridan JF:
Psychosocial stress and inflammation in cancer. Brain Behav Immun.
30(Suppl): S41–S47. 2013. View Article : Google Scholar
|
|
113
|
Zitvogel L, Kepp O, Galluzzi L and Kroemer
G: Inflammasomes in carcinogenesis and anticancer immune responses.
Nat Immunol. 13:343–351. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
114
|
Müzes G and Sipos F: Inflammasome,
inflammation and cancer: An interrelated pathobiological triad.
Curr Drug Targets. 16:249–257. 2015. View Article : Google Scholar
|
|
115
|
Dunn JH, Ellis LZ and Fujita M:
Inflammasomes as molecular mediators of inflammation and cancer:
Potential role in melanoma. Cancer Lett. 314:24–33. 2012.
View Article : Google Scholar
|
|
116
|
Shahzad MM, Arevalo JM, Armaiz-Pena GN, Lu
C, Stone RL, Moreno-Smith M, Nishimura M, Lee JW, Jennings NB,
Bottsford-Miller J, et al: Stress effects on FosB- and
interleukin-8 (IL8)-driven ovarian cancer growth and metastasis. J
Biol Chem. 285:35462–35470. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
117
|
Cosci F, Fava GA and Sonino N: Mood and
anxiety disorders as early manifestations of medical illness: A
systematic review. Psychother Psychosom. 84:22–29. 2015. View Article : Google Scholar
|
|
118
|
Casorelli I, Bossa C and Bignami M: DNA
damage and repair in human cancer: Molecular mechanisms and
contribution to therapy-related leukemias. Int J Environ Res Public
Health. 9:2636–2657. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
119
|
Higgins CF: Multiple molecular mechanisms
for multidrug resistance transporters. Nature. 446:749–757. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
120
|
Abraham J, Salama NN and Azab AK: The role
of P-glycoprotein in drug resistance in multiple myeloma. Leuk
Lymphoma. 56:26–33. 2015. View Article : Google Scholar
|
|
121
|
Su F, Ouyang N, Zhu P, Ouyang N, Jia W,
Gong C, Ma X, Xu H and Song E: Psychological stress induces
chemoresistance in breast cancer by upregulating mdr1. Biochem
Biophys Res Commun. 329:888–897. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
122
|
Reeder A, Attar M, Nazario L, Bathula C,
Zhang A, Hochbaum D, Roy E, Cooper KL, Oesterreich S, Davidson NE,
et al: Stress hormones reduce the efficacy of paclitaxel in triple
negative breast cancer through induction of DNA damage. Br J
Cancer. 112:1461–1470. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
123
|
Yao H, Duan Z, Wang M, Awonuga AO,
Rappolee D and Xie Y: Adrenaline induces chemoresistance in HT-29
colon adenocar-cinoma cells. Cancer Genet Cytogenet. 190:81–87.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
124
|
Pu J, Bai D, Yang X, Lu X, Xu L and Lu J:
Adrenaline promotes cell proliferation and increases
chemoresistance in colon cancer HT29 cells through induction of
miR-155. Biochem Biophys Res Commun. 428:210–215. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
125
|
Kong W, He L, Coppola M, Guo J, Esposito
NN, Coppola D and Cheng JQ: MicroRNA-155 regulates cell survival,
growth, and chemosensitivity by targeting FOXO3a in breast cancer.
J Biol Chem. 285:17869–17879. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
126
|
Hassan S, Karpova Y, Baiz D, Yancey D,
Pullikuth A, Flores A, Register T, Cline JM, D’Agostino R Jr,
Danial N, et al: Behavioral stress accelerates prostate cancer
development in mice. J Clin Invest. 123:874–886. 2013.PubMed/NCBI
|
|
127
|
Sun X, Bao J, Nelson KC, Li KC, Kulik G
and Zhou X: Systems modeling of anti-apoptotic pathways in prostate
cancer: Psychological stress triggers a synergism pattern switch in
drug combination therapy. PLOS Comput Biol. 9:e10033582013.
View Article : Google Scholar : PubMed/NCBI
|
|
128
|
Geller LT, Barzily-Rokni M, Danino T,
Jonas OH, Shental N, Nejman D, Gavert N, Zwang Y, Cooper ZA, Shee
K, et al: Potential role of intratumor bacteria in mediating tumor
resistance to the chemotherapeutic drug gemcitabine. Science.
357:1156–1160. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
129
|
van Bodegom M, Homberg JR and Henckens
MJAG: Modulation of the hypothalamic-pituitary-adrenal axis by
early life stress exposure. Front Cell Neurosci. 11:872017.
View Article : Google Scholar : PubMed/NCBI
|
|
130
|
Antoni MH: Psychosocial intervention
effects on adaptation, disease course and biobehavioral processes
in cancer. Brain Behav Immun. 30(Suppl): S88–S98. 2013. View Article : Google Scholar
|
|
131
|
Saxton JM, Scott EJ, Daley AJ, Woodroofe
M, Mutrie N, Crank H, Powers HJ and Coleman RE: Effects of an
exercise and hypocaloric healthy eating intervention on indices of
psychological health status, hypothalamic-pituitary-adrenal axis
regulation and immune function after early-stage breast cancer: A
randomised controlled trial. Breast Cancer Res. 16:R392014.
View Article : Google Scholar : PubMed/NCBI
|
|
132
|
Chida Y, Hamer M, Wardle J and Steptoe A:
Do stress-related psychosocial factors contribute to cancer
incidence and survival? Nat Clin Pract Oncol. 5:466–475. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
133
|
Dieli-Conwright CM and Orozco BZ: Exercise
after breast cancer treatment: Current perspectives. Breast Cancer
(Dove Med Press). 7:353–362. 2015.
|
|
134
|
Nota JA and Coles ME: Shorter sleep
duration and longer sleep onset latency are related to difficulty
disengaging attention from negative emotional images in individuals
with elevated transdiagnostic repetitive negative thinking. J Behav
Ther Exp Psychiatry. 58:114–122. 2018. View Article : Google Scholar
|
|
135
|
Guzman-Marin R and Avidan AY: Sleep
disorders in patients with cancer. J Community Support Oncol.
13:148–155. 2015. View Article : Google Scholar : PubMed/NCBI
|