Introduction
Lung cancer is one of the most frequently diagnosed
types of cancer and the leading cause of cancer-associated
mortality worldwide (1). Non-small
cell lung cancer (NSCLC), which includes squamous cell carcinoma
and adenocarcinoma, is a highly metastatic subset of lung cancer
(2-4). At present, typical treatments for
NSCLC include surgery, radiotherapy and chemotherapy; however,
despite advances in these techniques, the median survival period of
patients with late-stage and metastatic NSCLC remains
unsatisfactory (5-7). As such, developing novel effective
treatments for NSCLC is of utmost importance.
MicroRNAs (miRNAs or miRs) are a class of small
non-coding RNAs of 20-25 nucleotides in length (8). miRNAs are able to bind to the
3′-untranslated region (3′-UTR) of target genes, leading to mRNA
degradation or suppressing the translation of target mRNAs
(9-11). It is considered that ~33% of human
genes associated with tumorigenesis, embryonic development and
inflammation are regulated by miRNAs (12-15).
Increasing evidence indicates that tumor cell proliferation,
migration and invasion are directly governed by miRNAs, suggesting
that miRNAs may be promising therapeutic targets for tumor
treatment (16,17).
miR-98-5p, a member of the let-7 family of miRNAs,
is associated with a number of types of cancer, including lung
cancer, colorectal cancer and breast cancer (18-21).
It has been reported that miR-98-5p enhances radiosensitivity and
chemosensitivity, while it inhibits proliferation, migration and
invasion in NSCLC (22-24). However, the inherent molecular
mechanisms through which miR-98-5p affects NSCLC progression remain
largely unknown. As such, in this study, we focused on transforming
growth factor beta receptor 1 (TGFBR1), which is one of the most
established target genes of miR-98-5p. TGFBR1 is involved in the
regulation of cellular processes, including motility,
differentiation, adhesion, division and apoptosis (25-28).
TGFBR1 also plays an important role in tumor progression, with its
activation or overexpression often observed in various types of
cancer. For example, TGFBR1 has been shown to enhance the migration
and invasion of MCF-7 breast cancer cells and to promote the
invasion and metastasis of colorectal cancer (29-31).
Furthermore, TGFBR1 plays a role in tumor invasion and metastasis
by mediating epithelial-to-mesenchymal transition (EMT) (31). Taken together, these data suggest
that TGFBR1 inhibition may serve as a potential target for cancer
therapy.
The aim of the present study was to investigate the
function of miR-98-5p in NSCLC progression and explore the
interaction between miR-98-5p and TGFBR1. The results of our study
demonstrate that miR-98-5p attenuates tumor growth and metastasis
by suppressing TGFBR1 and EMT signaling in NSCLC.
Materials and methods
Clinical tissue samples
Lung cancer specimens and adjacent normal tissues
(n=70) were obtained from patients suffering from NSCLC who had not
received any pre-operative radiotherapy and chemotherapy at Wujin
People’s Hospital of Changzhou (Changzhou, China) from January,
2010 to May, 2013. The patients’ sex distribution (male, n=39;
female, n=31) and age distribution in years (≥60, n=38; <60,
n=32; age range, 27 to 79 years; mean age, 58.88±12.65 years) were
recorded. The diagnosis of lung cancer was performed by our
surgeons and pathologists. The collected tissues were stored at
−80°C in liquid nitrogen until use. Tissues and information of
patients were obtained with written informed consent. The
procedures used in this study were approved by the Human Ethics
Committee at the Wujin People’s Hospital of Changzhou. All the
experiments were conducted according to the approved
guidelines.
Cell lines and animals
The human NSCLC A549 (no. CCL-185), H1299 (no.
CRL-5803), ANIP-973 (no. CS-0168) and GLC-82 (no. ZY-H065) cell
lines (American Type Culture Collection, Manassas, VA, USA) were
cultured in DMEM (Gibco/Thermo Fisher Scientific, Inc., Waltham,
MA, USA) supplemented with 10% fetal bovine serum (FBS) and
antibiotics at 37°C in a humidified atmosphere containing 5%
CO2.
A total of 10 male NOD/SCID mice (6-8 weeks old)
were purchased from HFK Bio-Technology Co. (Beijing, China). Mice
were housed under specific pathogen-free (SPF) conditions and were
quarantined for 1 week prior to treatment. The animals were housed
in a controlled environment with a temperature of 20-22°C, relative
humidity of 50-60% and 12 h light/dark cycles, and were provided
with access to food and water ad libitum. All animal
procedures were conducted in accordance with the protocol approved
by the Institutional Animal Care and Treatment Committee of Wujin
People’s Hospital of Changzhou. All animals were treated humanely
throughout the experimental period.
RNA preparation and reverse
transcription-quantitative PCR (RT-qPCR)
Total RNA was extracted from the collected tissues
or cells using TRIzol reagent (Invitrogen/Thermo Fisher Scientific,
Inc.) according to the manufacturer’s instructions. RNA was
converted into cDNA using a Reverse Transcription kit (Thermo
Fisher Scientific, Inc.). PCR was then performed using Taq
polymerase (Takara Bio, Inc., Otsu, Japan). Stem-loop primers
(Guangzhou RiboBio Co., Ltd., Guangzhou, China) were used to detect
miRNAs. U6 small nucleolar RNA and GAPDH were used for
normalization. The relative expression levels were calculated using
the 2−ΔΔCt method (32)
(CFX manager software 3.1; Bio-Rad Laboratories, Inc., Hercules,
CA, USA). The primers used were as follows: hsa-miR-98-5p forward,
5′-ACACTCCCUAUACAACUUAC-3′ and reverse,
5′-GGGAAAGUAGUGAGGCCTCAGA-3′; TGFBR1 forward,
5′-TCGTCTGCATCTCACTCAT-3′ and reverse, 5′-GATAAATCTCTGCCTCACG-3′;
and GAPDH forward, 5′-TCTCTGCTCCTCCTGTTC-3′ and reverse,
5′-GGTTGAGCACAGGGTACTTTATTGA-3′. Quantitative PCR was carried out
using SYBR Premix Ex Taq II (Tli RNaseH Plus; 2X; 6 µl), 0.5
µl PCR forward primer (10 µM), 0.5 µl PCR
reverse primer (10 µM), 1 µl cDNA solution, 3
µl RNase-free dH2O, total was 10 µl. The
cycling conditions for PCR were as follows: Stage 1: 95°C, 30 sec;
repeat: 1; stage 2: 95°C, 5 sec, 60°C, 34 sec; repeat: 40; stage 3:
dissociation.
Transfection
Hsa-miR-98-5p mimics, hsa-miR-98-5p inhibitors and
the corresponding negative controls were purchased from Obio
Technology (Shanghai, China) and transfected into the A549, H1299,
ANIP-973 and GLC-82 cells using Lipofectamine® 2000
(Invitrogen/Thermo Fisher Scientific, Inc.) according to the
manufacturer’s instructions. At 48 h following transfection, the
cells were collected for use in further experiments.
Cell viability and congenic survival
Cell viability was assessed using an MTT assay. The
cells were seeded into 96-well plates at a density of 2,000
cells/well. After 12 h, the cells were transfected with miR-98-5p
mimics or miR-NC. At 24, 48, 72 or 96 h following transfection, 20
µl MTT solution (5 mg/ml; Amresco, Inc., Framingham, MA,
USA) was added to each well followed by incubation at 37°C for 4 h.
The cells were subsequently treated with 200 µl DMSO to
dissolve the precipitated crystals. Finally, the absorbance of each
well was measured using a microplate reader (EL10A; Biobase, Jinan,
China) at 570 nm. All experiments were performed 3 times. Data are
presented as the means ± standard deviation (SD).
The cells were seeded in 6-well plates (500
cells/well) and incubated at 37°C for 12 h. Following transfection
with miR-98-5p mimics or miR-NC, the cells were further incubated
at 37°C for 6-8 days. The cells were fixed with methanol for 10 min
and stained with 0.1% crystal violet at room temperature for 30
min. Images were subsequently captured using a microscope (Olympus,
Tokyo, Japan).
Cell migration and invasion assay
The cell migratory and invasive abilities were
investigated using a 24-well plate with 8-µm pore size
Boyden chamber inserts. For the migration assays, 4×104
cells were seeded into the upper chamber without a coated membrane.
For the invasion assays, 1×105 cells were seeded into
the upper chamber with a Matrigel-coated membrane. The cells in the
upper chamber were transfected with miR-98-5p mimics or inhibitors
and suspended in 200 µl serum-free DMEM. The control groups
were transfected with corresponding negative controls. A total of
800 µl DMEM supplemented with 10% FBS was added to the lower
chamber. The chambers were incubated for 24 h at 37°C and the
non-migrated and non-invasive cells remaining in the upper chamber
were removed using a cotton swab. Migrated or invaded cells in the
lower chamber were fixed with methanol and stained with 0.1%
crystal violet at room temperature for 20 min. Finally, the cells
were imaged by the microscope (Olympus) and counted using
high-power magnifications.
Wound healing assay
The cells were seeded in 6-well plates
(5×105 cells/well) and incubated at 37°C for 12 h. When
the cells reached 90% confluence, a scratch wound was made using a
10-µl pipette tip. The cells were subsequently washed with
PBS to remove cell debris and transfected with miR-98-5p mimics or
inhibitors. The control groups were transfected with corresponding
negative controls. The cells were incubated with DMEM supplemented
with 1% FBS for 24 h, following which images were captured under a
microscope (Olympus).
Western blot analysis
Western blot assays were performed to measure the
expression of target proteins. Total cellular proteins were
extracted from the cultured cells using lysis buffer [20 mM Tris
(pH7.4), 150 mM NaCl, 5 mM EDTA, 50 mM NaF and 0.1% NP-40].
Proteins were separated by SDS-PAGE (10%, w/v) and transferred onto
PVDF membranes, which were then blocked using 5% skim milk for 2 h
at room temperature. The membranes were subsequently incubated with
target primary antibodies against the following: MMP-9 [13667S;
Cell Signaling Technology (CST), Danvers, MA, USA], vimentin
(10R-1903; Fitzgerald Industries International, Acton, MA, USA),
α-smooth muscle actin (α-SMA; 19245; CST), TGFBR1 (70R-36484) and
β-actin (10R-10263) (both from Fitzgerald Industries International)
at 4°C overnight. The dilution used for MMP-9, vimentin, α-SMA and
TGFBR1 was 1:100, and the dilution used for β-actin was 1:1,000.
Following incubation with horseradish peroxidase-labeled secondary
antibodies (A0216; Byotime, Shanghai, China) which were diluted at
1:1,000 at 37°C for 2 h, the protein bands were visualized using a
chemiluminescence (ECL) detection system and images were
captured.
Dual-luciferase reporter assay
The 3′-UTR of wild-type (WT) and mutant TGFBR1 were
amplified from human genomic DNA and individually inserted into
pmiR-RB-REPORT™ luciferase vectors (Obio, Shanghai, China). The
A549 cells were co-transfected with 200 ng of mutant or WT
pmiR-RB-REPORT™ plasmid and 100 ng of miR-98-5p mimics or miR-NC
Following incubation for 36 h, and luciferase activity was measured
with a Dual-Luciferase Reporter Assay System (Promega Corporation,
Madison, WI, USA) according to the manufacturer’s instructions.
In vivo anti-tumor evaluation
A549 cells suspensions (1×107 cells; 200
µl) were subcutaneously injected into the right flanks of
NOD/SCID mice to establish a xenograft NSCLC model. After 6 days,
tumor volume reached ~200 mm3, and the mice were
randomly divided into 2 groups as follows: The control (n=5) and
miR-98-5p mimic (n=5) group. A total of 5 µg/mouse plasmid
was administered every 3 days. The treatment was terminated when
mice in the control group became moribund (at day 21). Mice were
sacrificed, and tumors were harvested, weighed and imaged. Finally,
tumors were collected for further hematoxylin and eosin (H&E)
and immunohistochemical evaluation. Tissues were collected, fixed
in 4% paraformaldehyde, embedded in paraffin and sectioned (5
µm thickness). After dewaxing and rehydration, the sections
were stained with H&E at room temperature for 10 sec and imaged
under a light microscope (Olympus). For immunohistochemistry, the
sections were respectively incubated with primary rat anti-mouse
antibody Ki-67 (9449; CST; dilution: 1:400), MMP-9 (13667S; CST;
dilution: 1:325) and TGFBR1 (70R-36484; Fitzgerald Industries
International; dilution: 1:200), and then treated with secondary
antibody biotinylated goat anti-rat immunoglobulin (Abcam,
Cambridge, MA, USA). The sections were incubated with
streptavidin-peroxidase and DAB solution (Solarbio, Beijing, China)
at 37°C for 20 min to visualize the biotinylated goat anti-rat
immunoglobulin. The sections were then treated with hematoxylin at
room temperature for 10 sec to stain the nuclei of the tumor cells.
Finally, the sections were imaged and examined under a light
microscope (Olympus).
In vivo anti-metastasis assessment
A total of 1×106 of A549 stable
luciferase reporter cells suspended in 100 µl PBS were
injected into NOD/SCID mice via the tail vein to establish a model
of pulmonary metastasis. At 3 days after inoculation, 10 mice were
randomly divided into the control and miR-98-5p mimics groups (n=5)
and treated with 5 µg miR-98-5p mimic plasmids or control
plasmids, respectively, every 3 days. At 5, 10 and 15 days
following inoculation, mice were administered with D-luciferin (3
mg/mouse) for analysis. Bioluminescence was then detected from lung
metastatic tumors was detected using an IVIS Lumina In Vivo Imaging
System (Perkin-Elmer, Waltham, MA, USA). Finally, the lungs were
harvested from each mouse to count the metastatic nodes and fixed
with 4% paraformaldehyde for further H&E evaluation.
Statistical analysis
The results are presented as the means ± SD.
Statistical analysis was performed using SPSS 17.0 software (SPSS
Inc., Chicago, IL, USA). The significance among multiple groups was
evaluated using one-way ANOVA followed by Newman-Keuls (SNK)
t-test. Significant differences between 2 groups (parametric) were
analyzed using a Student’s t-test, and significant differences
between 2 groups (non-parametric) were analyzed by the Mann-Whitney
U test. A value of P<0.05 was considered to indicate a
statistically significant difference.
Results
miR-98-5p is downregulated in NSCLC and
is associated with progression
Lung cancer progression is a complex process
associated with the activation of oncogenes and the silencing of
tumor suppressor genes (32-34).
It has been reported that miRNA dysregulation is significantly
associated with tumorigenesis and cancer progression (35). In this study, to confirm the
biological function of miR-98-5p in NSCLC, we first detected its
expression in 70 human primary lung tumor tissues and adjacent
normal lung tissues by RT-PCR. The results revealed that miR-98-5p
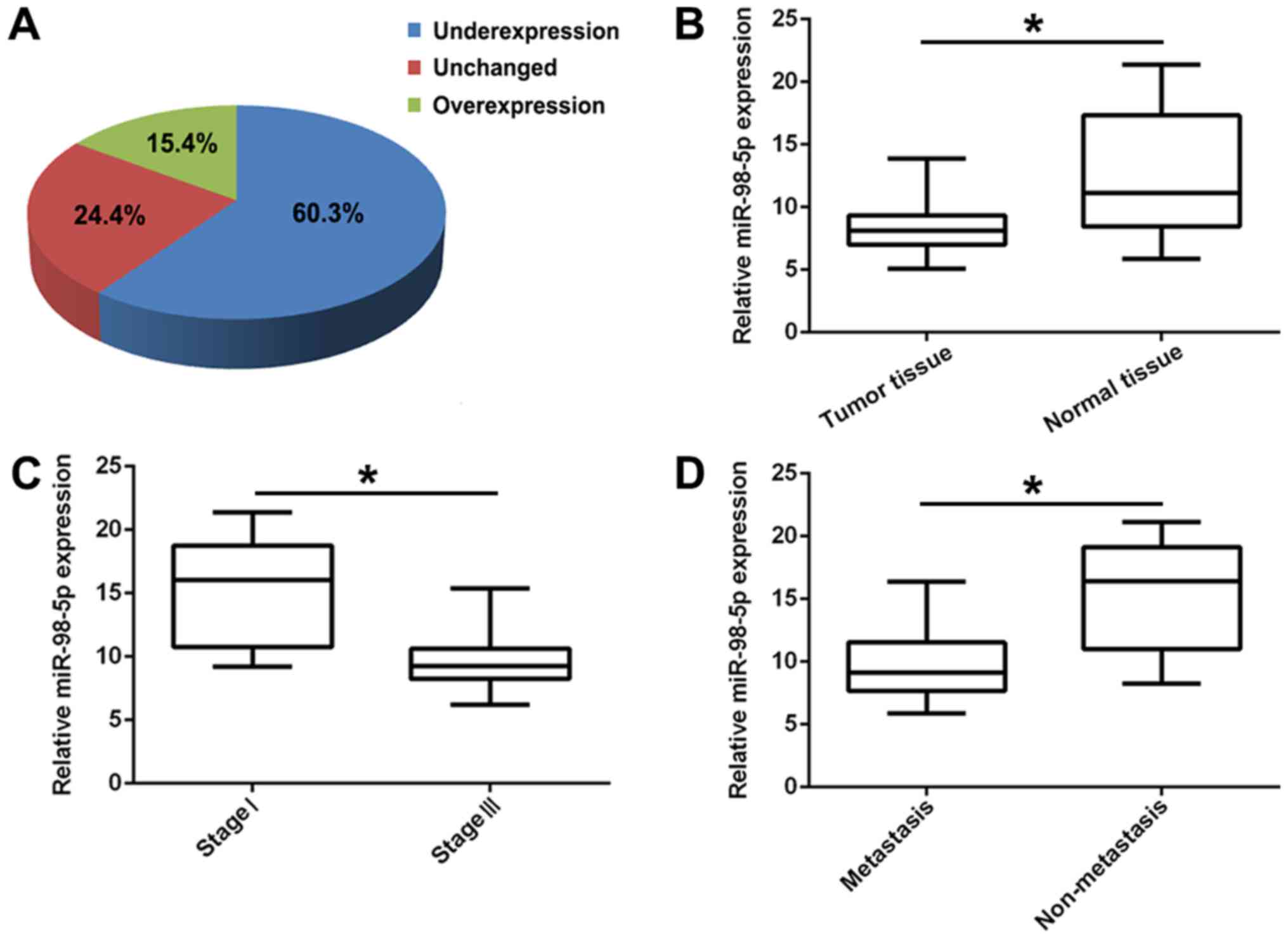
expression was significantly downregulated in the tumor tissues
(60.3%) compared with in the controls (Fig. 1A and B; P<0.01). In addition,
the expression of miR-98-5p in patients with advanced NSCLC (stage
III; n=41) was significantly decreased compared with that in
patients with the early stage NSCLC (stage I; n=29; P<0.05;
Fig. 1C). Furthermore, compared
with non-metastatic NSCLC (n=25), miR-98-5p expression was
distinctly downregulated in metastatic NSCLC (n=45; P<0.05,
Fig. 1D). These results suggest
that miR-98-5p is downregulated in NSCLC and is associated with
NSCLC progression.
miR-98-5p overexpression inhibits NSCLC
tumorigenesis in vitro
In order to investigate the biological role of
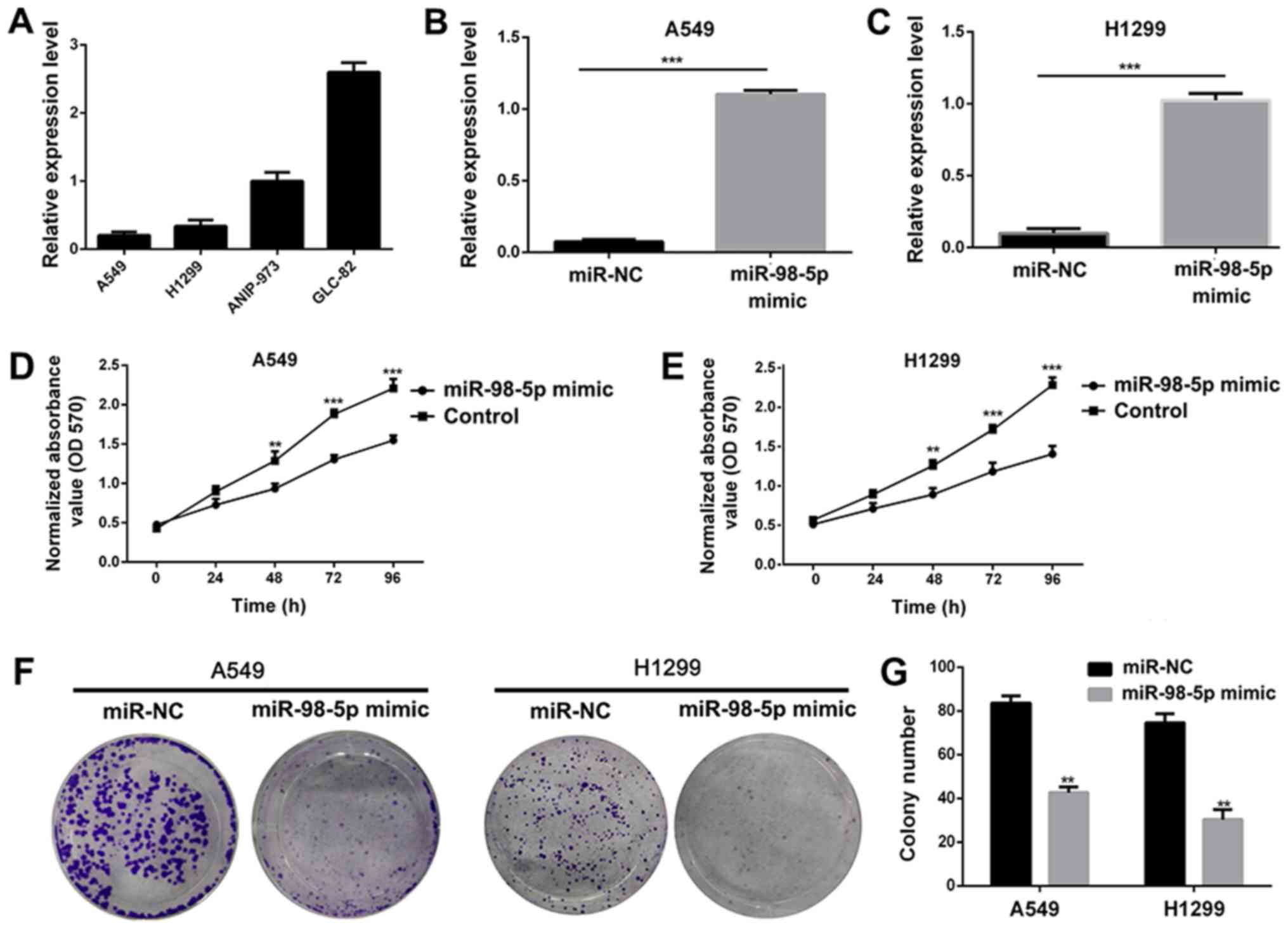
miR-98-5p in NSCLC, we measured the expression of miR-98-5p in 5
different NSCLC cell lines. It was observed that miR-98-5p
expression was downregulated in the A549 and H1299 cells, but was
increased in the ANIP-973 and GLC-82 cells (Fig. 2A). We thus transfected the A549 and
H1299 cells with plasmid miR-98-5p mimics to restore miR-98-5p
expression, and the results revealed that miR-98-5p expression was
significantly increased following transfection (P<0.001;
Fig. 2B and C). Following
transfection with miR-98-5p mimics, the viability (Fig. 2D and E) and colony formation
abilities (Fig. 2F and G) of the
A549 and H1299 cells were significantly suppressed, indicating an
anti-proliferative effect of miR-98-5p mimics. A number of studies
have demonstrated that miRNA dysregulation is associated with tumor
migration and invasion (33,34),
and we thus used miR-98-5p mimics to restore the expression level
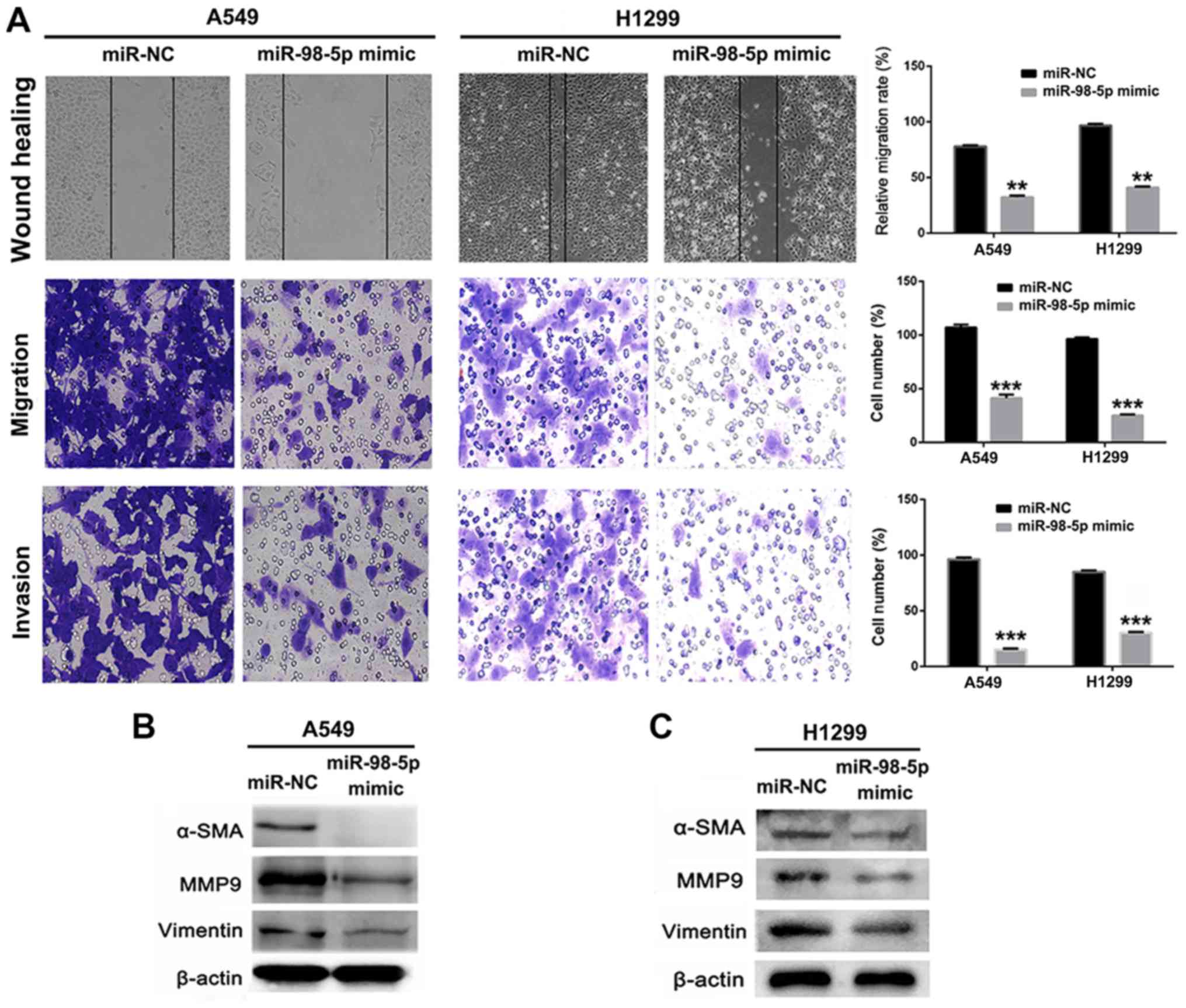
of miR-98-5p in the A549 and H1299 cells. The results of a wound
healing assay revealed that the migration of the A549 and H1299
cells was suppressed following transfection with miR-98-5p mimic
(P<0.05; Fig. 3A). Furthermore,
Transwell assays revealed that the migration and invasion of A549
and H1299 cells were distinctly suppressed by transfection with
miR-98-5p mimics (P<0.001). The expression of proteins
associated with tumor metastasis and EMT was measured using western
blotting following treatment with miR-98-5p mimics. As shown in
Fig. 3B and C, the expression of
MMP-9, α-SMA and vimentin was significantly downregulated in the
A549 and H1299 cells following transfection with miR-98-5p mimics.
These results suggested that the restoration of miR-98-5p
expression significantly suppressed the viability, proliferation,
migration and invasion of both the A549 and H1299 cells, acting as
a tumor suppressor in vitro.
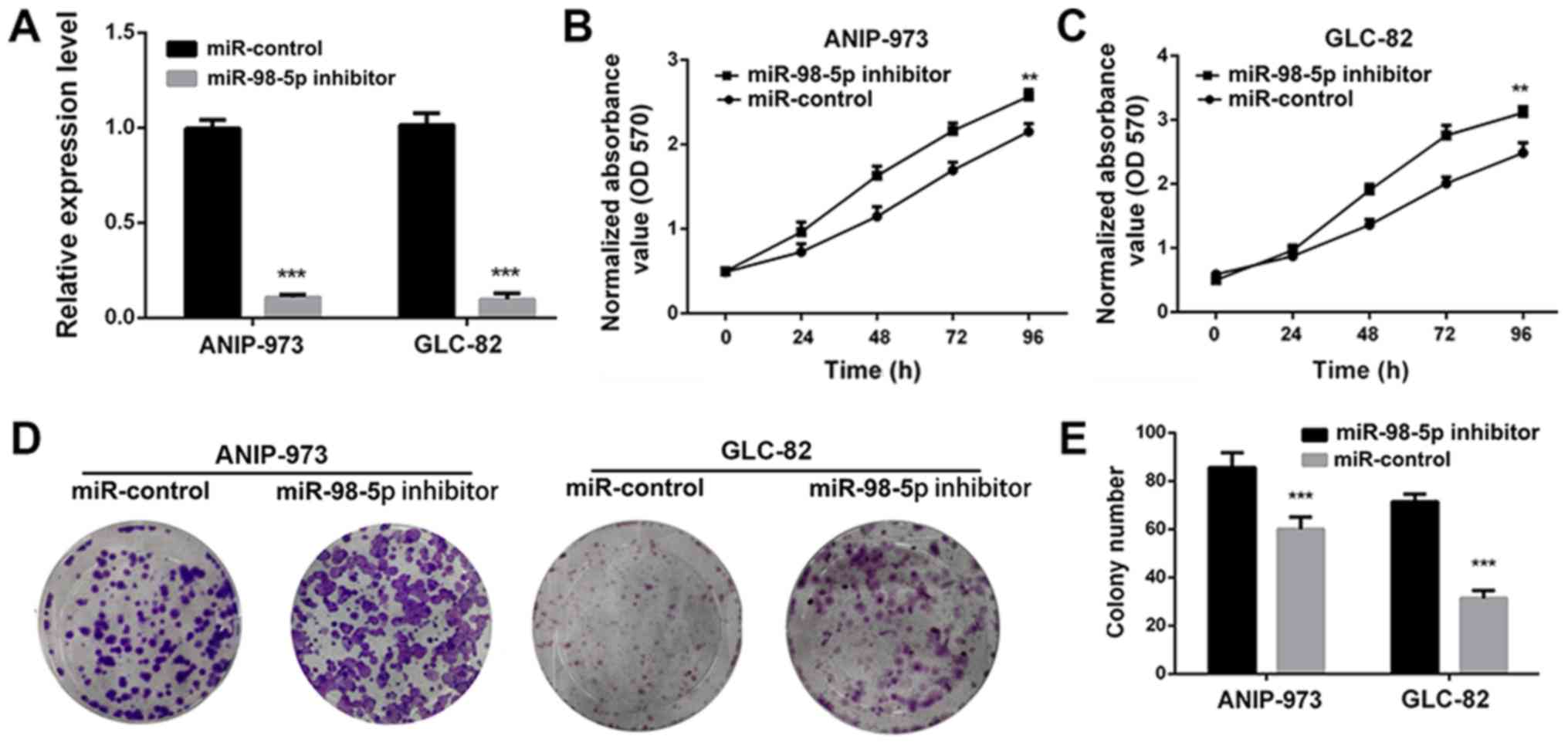
To further confirm that miR-98-5p acts as a tumor
suppressor in NSCLC, we utilized a plasmid miR-98-5p inhibitor.
Compared with the miR-controls, transfection with miR-98-5p
inhibitors significantly decreased the expression of miR-98-5p in
both the ANIP-973 and GLC-82 cells (Fig. 4A; P<0.001). When miR-98-5p
expression was downregulated, the viability (Fig. 4B and C) and colony formation
ability (Fig. 4D and E) of the
ANIP-973 and GLC-82 cells were increased. Furthermore, cell
migration and invasion were investigated following the knockdown of
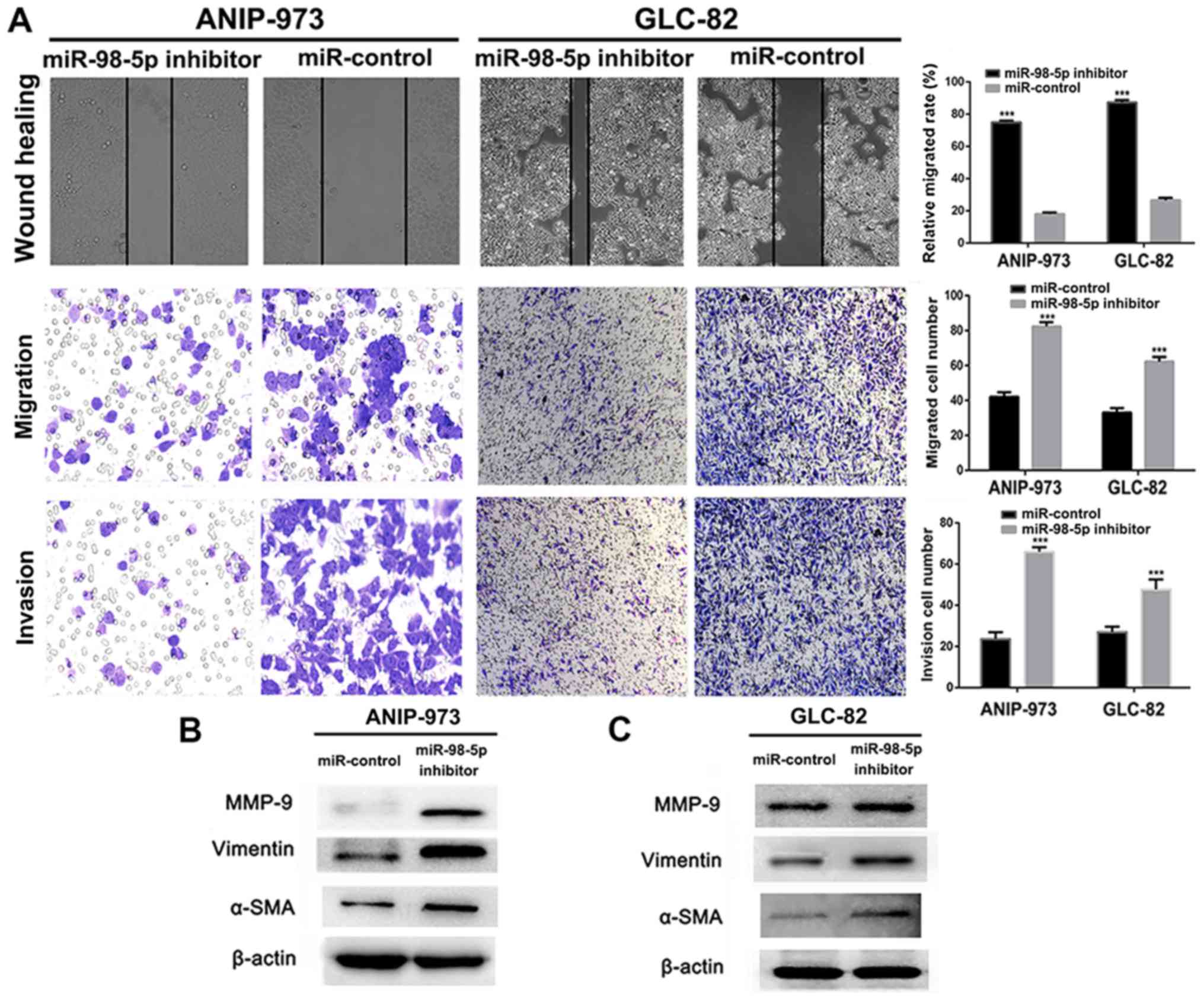
miR-98-5p. It was demonstrated that the mobility of the ANIP-973
and GLC-82 cells was increased following miR-98-5p downregulation
(Fig. 5A). In addition, the
migration and invasion of the ANIP-973 and GLC-82 cells were
distinctly enhanced following transfection with miR-98-5p
inhibitors. When the ANIP-973 and GLC-82 cells were transfected
with the miR-98-5p inhibitors, the expression levels of
metastasis-associated proteins (MMP-9, α-SMA and vimentin) were
upregulated, which was in agreement with the wound healing and
Transwell assay results (Fig. 5B and
C). Taken together, these results suggest that miR-98-5p
downregulation enhances the viability, proliferation, migration and
invasion of NSCLC cells; in other words, miR-98-5p inhibits NSCLC
tumorigenesis in vitro.
TGFBR1 is a target of miR-98-5p
In order to further investigate the mechanisms
through which miR-98-5p affects NSCLC cell progression, we used the
computational prediction program TargetScan (www.targetscan.org) to predict target genes. In total,
60 genes were simultaneously predicted by the TargetScan databases,
and TGFBR1 was detected as a candidate gene related to NSCLC based
on its associated Gene Ontology (GO) terms and also harbors
miR-98-5p binding sites, suggesting that TGFBR1, which stimulates
tumor cell proliferation, migration and invasion via the TGF-β/Smad
signaling pathway (35-38), was identified as a potential target
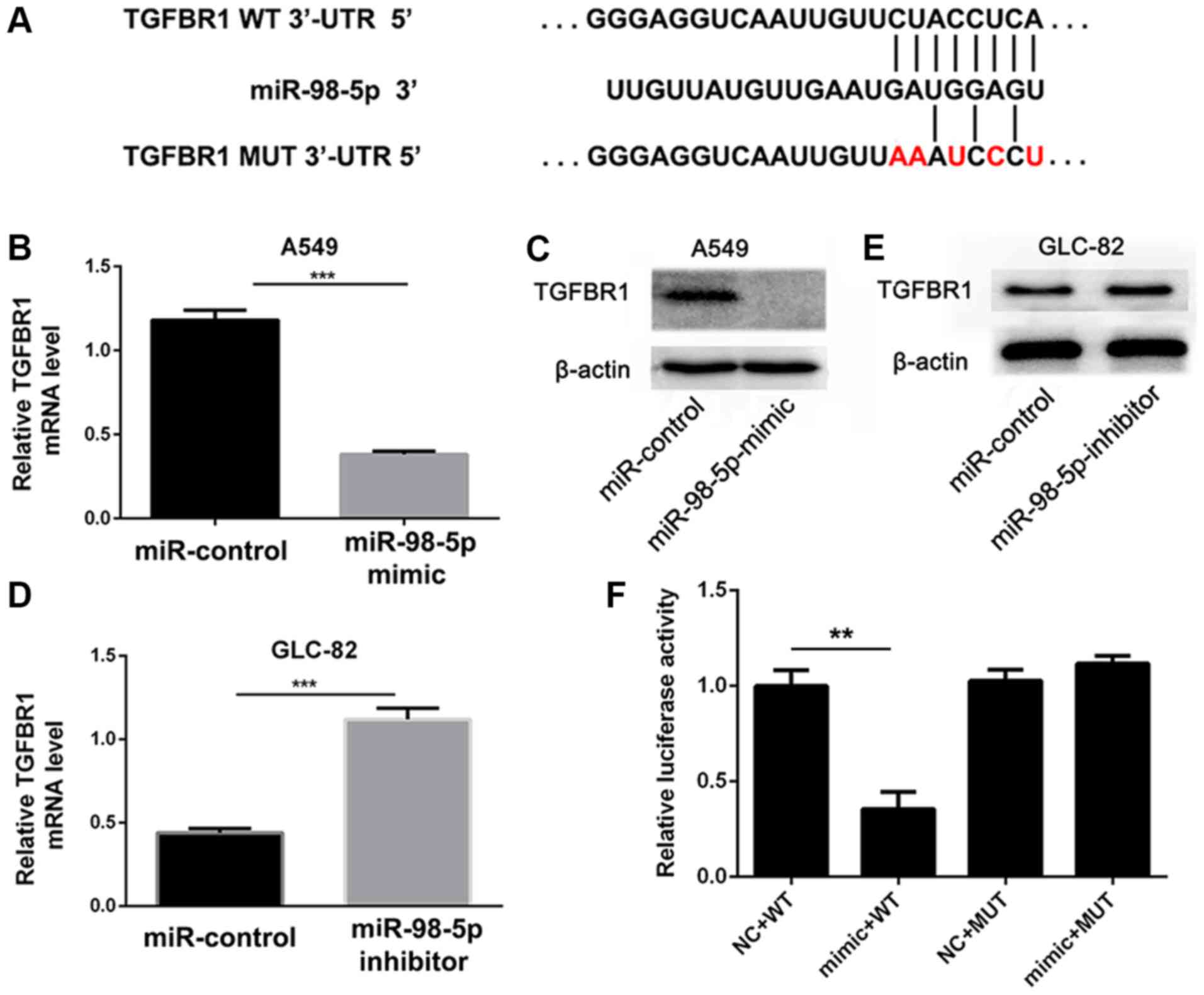
of miR-98-5p (Fig. 6A).
To confirm that TGFBR1 is a downstream target of
miR-98-5p, we transfected the A549 cells with miR-98-5p mimics and
found that TGFBR1 expression was significantly decreased
(P<0.001; Fig. 6B and C).
Conversely, TGFBR1 expression was upregulated in the GLC-82 cells
following miR-98-5p knockdown (P<0.001; Fig. 6D and E). A dual-luciferase reporter
assay was also performed to further verify that miR-98-5p regulates
TGFBR1. The WT TGFBR1 3′-UTR (pmiR-RB-REPORT™-TGFBR1-3′-UTR-WT) and
mutated TGFBR1-3′-UTR (pmiR-RB-REPORT™-TGFBR1-3′-UTR-MUT) were
cloned and co-transfected with miR-98-5p or miR-NC into A549 cells.
As shown in Fig. 6F, the
luciferase activity of pmiR-RB-REPORT™-TGFBR1-3′-UTR-WT was
distinctly inhibited by miR-98-5p mimics compared with miR-NC.
However, no significant differences were observed in the luciferase
activity of pmiR-RB-REPORT™-TGFBR1-3′-UTR-MUT in the presence of
miR-98-5p mimics or miR-NC. These results suggest that miR-98-5p
directly targets the 3′-UTR of TGFBR1.
miR-98-5p suppresses tumor growth in
vivo
Based on the anti-proliferative effects of miR-98-5p
in vitro, these effects were further assessed using a
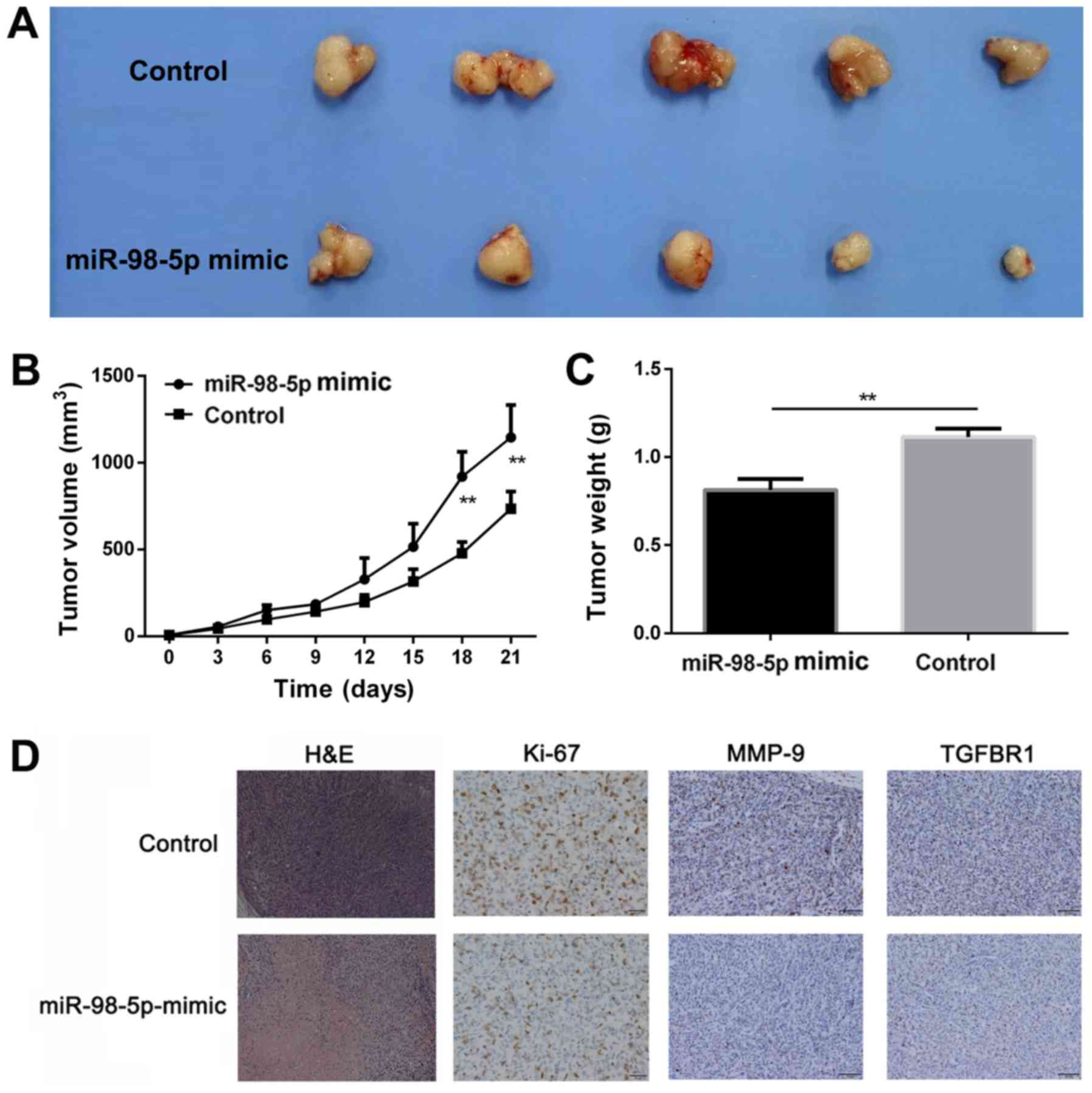
xenograft NSCLC mouse model. Compared with the control group, mice
in the miR98-5p mimics group exhibited a significantly slower tumor
growth (P<0.01; Fig. 7A and B).
Tumor weight was considerably reduced in the miR-98-5p mimic group
(0.81±0.091 g) compared with the control group (1.13±0.082 g;
P<0.01; Fig. 7C), indicating
that miR-98-5p exerted a significant anti-tumor effect.
Pathological analysis was performed to evaluate the anti-tumor
effects of miR-98-5p (Fig. 7D).
H&E staining of the tumor sections revealed preferable
anti-proliferative effects in the miR-98-5p mimics group. The
number of Ki-67-positive cells in the tumor sections was
considerably lower in the miR-98-5p mimic group. Furthermore,
TGFBR1 expression was upregulated in the control group compared
with the miR-98-5p mimic group. The expression of MMP-9 was
decreased in the miR-98-5p mimics group, which is in agreement with
the results of immunohistochemical TGFBR1 staining. Taken together,
these results suggest that miR-98-5p exerts significant anti-tumor
effects in vivo.
miR-98-5p suppresses tumor metastasis in
vivo
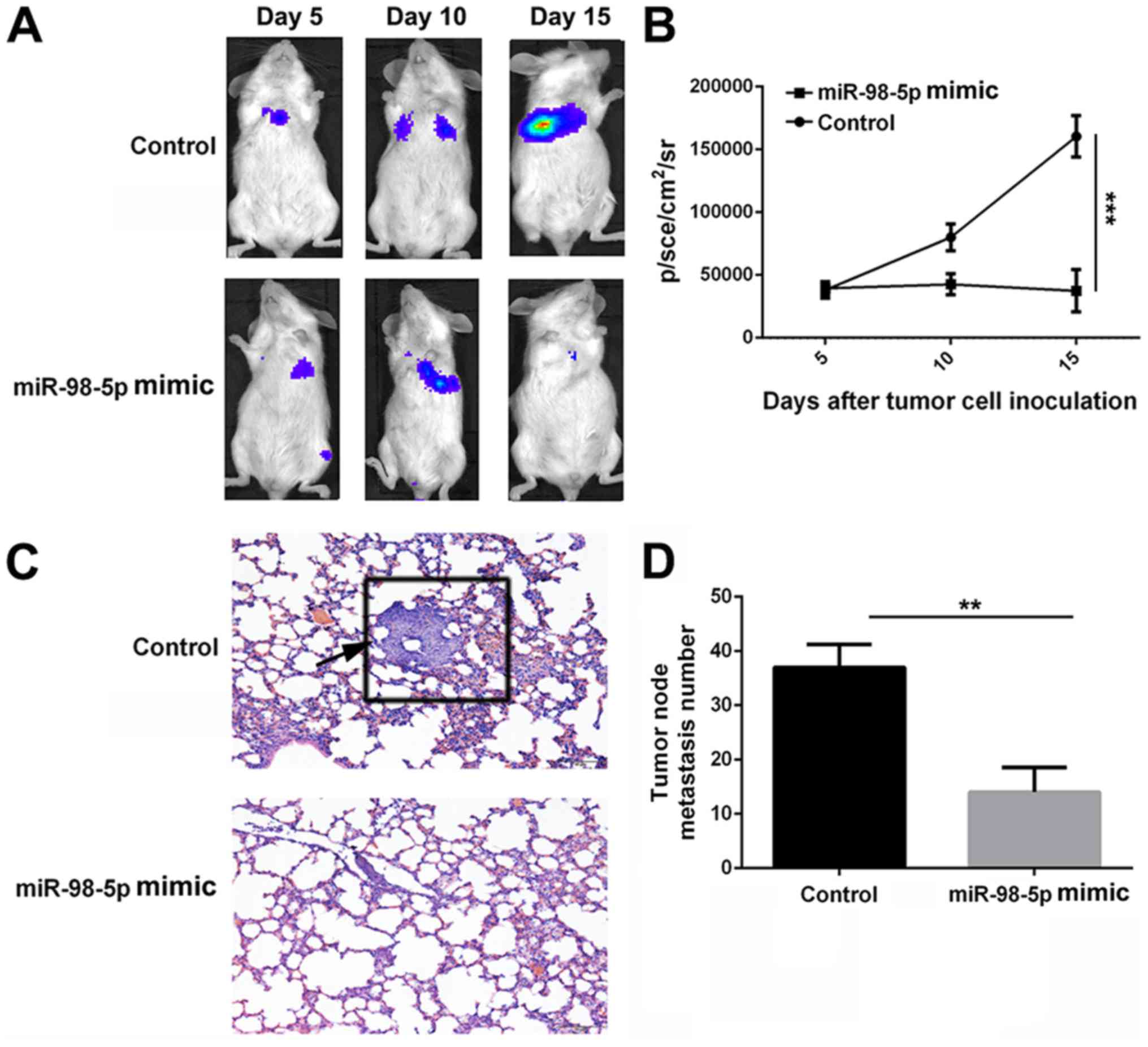
A pulmonary metastasis model of A549-luciferase
cells in NOD/SCID mice was used to evaluate the anti-metastatic
effect of miR-98-5p mimics. Bioluminescence at the lung site, which
indicates metastatic tumor cells, increased rapidly in the control
group following implantation. However, bioluminescence remained
largely unchanged in the miR-98-5p mimic treatment group. At 15
days after tumor cell implantation, bioluminescence was
significantly lower in the miR-98-5p mimic group (37475±16967.87
p/sec/cm2/sr) compared with the control group
(160432±16443.39 p/sec/cm2/sr; P<0.001). The
harvested lung tissues were subjected to H&E staining and
metastatic nodes were counted. As shown in Fig. 8D, the number of metastatic nodes in
the miR-98-5p group was decreased compared with the control group.
Furthermore, H&E staining of the lung tissue sections revealed
fewer tumor nodes in the miR-98-5p group compared with the control
group (Fig. 8C). These results
suggest that miR-98-5p is able to effectively inhibit tumor
metastasis in vivo.
Discussion
Lung cancer is a complex process which is related to
the activation of oncogenes and the silencing of tumor suppressor
genes (39-41). It has been reported that miRNAs are
significantly associated with tumorigenesis, progression and
metastasis (42). Moreover, the
disrupted expression of miRNAs is commonly found in human NSCLC. In
this study, the role of miR-98-5p in lung cancer was investigated
and the underlying molecular mechanisms were examined.
A previous study revealed that miR-98 expression was
markedly decreased both in lung cancer tissues and in immortal
cancer cell lines (18). In this
study, we first identified miR-98-5p was an anti-tumor miRNA in
NSCLC that was significantly downregulated in tumor tissues in
comparison with normal tissues. The expression levels of miR-98-5p
in advanced NSCLC were notably lower than those in tissues from
patients with early stages of disease. Furthermore, compared with
non-metastatic NSCLC, the miR-98-5p expression levels in metastatic
NSCLC were distinctly downregulated. Thus, it was suggested from
the above results that miR-98-5p was downregulated in NSCLC and was
associated with progression in NSCLC, in agreement with previous
studies (18,43).
An increasing number of studies have indicated that
miR-98 suppresses the proliferation, migration and invasion of
various tumor cells. For example, miR-98 has been shown to inhibit
the proliferation, invasion and migration, and promote the
apoptosis of breast cancer cells by binding to HMGA2 (44). Cai et al also reported that
SNHG16 contributed to breast cancer cell migration by competitively
binding miR-98 (45). In the
current study, we evaluated the biological roles of miR-98-5p in
NSCLC in vitro. We demonstrated that transfection with
miR-98-5p mimics suppressed the proliferation, colony formation and
migration of NSCLC cells. However, the decreased expression of
miR-98-5p promoted tumor cell proliferation, colony formation and
migration. Various studies have indicated that disrupted levels of
miRNAs are closely associated with tumor migration and invasion
(16,17). In this study, to investigate the
anti-metastatic mechanisms of action of miR-98-5p, we used western
blot analysis to demonstrate that miR-98-5p impairs the metastasis
of tumor cells via the EMT process. In brief, the restoration of
the expression levels of miR-98-5p which resulted from transfection
with miR-98-5p mimics significantly suppressed the viability,
proliferation, migration and invasion of NSCLC cells, that is to
say, miR-98-5p is a tumor suppressor and inhibits NSCLC
tumorigenesis in vitro.
To further elucidate the underlying mechanisms of
action of miR-98-5p as regards the development of NSCLC, we carried
out in vitro experiments using NSCLC cell lines and
certified that TGFBR1 was a downstream target of miR-98-5p. Due to
the excellent anti-proliferative efficiency of miR-98-5p in
vitro, we utilized it for further anti-tumor assessment in a
xenograft NSCLC mouse model. Mice in the miR-98-5p mimics group
exhibited a significantly slower tumor growth rate, and the tumor
weight of the mice in the miR-98-5p mimic group was considerably
less than that of the control group. Pathological analysis revealed
that preferable anti-proliferative efficiency was found in the
miR-98-5p mimics group. Therefore, the above results demonstrated
that miR-98-5p exerted efficient anti-tumor effects in vivo.
On account of the considerable anti-metastatic efficacy of
miR-98-5p mimics in vitro, we exploited a pulmonary
metastasis model of A549 reporter luciferase cells in NOD/SCID
mouse to evaluate the anti-metastatic efficacy of the miR-98-5p
mimic. The metastatic tumor cells which were located in the lung
site were distinctly decreased in the miR-98-5p-transfected group
in comparison with the control group, indicating that miR-98-5p
effectively inhibited tumor metastasis in vivo.
In conclusion, the results of the present study
suggest that miR-98-5p, which is downregulated in NSCLC and is
associated with tumor progression, effectively inhibits the
proliferation, migration and invasion of NSCLC cells in
vitro by targeting TGFBR1. In addition, miR-98-5p was observed
to have considerable anti-tumor efficacy in an A548 subcutaneous
xenograft tumor mouse model. Furthermore, miR-98-5p significantly
suppressed tumor metastasis in a mouse model of pulmonary
metastasis. Thus, miR-98-5p and TGFBR1 may be potential therapeutic
candidates and targets for NSCLC treatment.
Acknowledgments
Not applicable.
Funding
This study was financially supported by Natural
Science Foundation of Jiangsu Province (BK20151175).
Availability of data and materials
All the data generated or analyzed during this study
is included in this published article or are available from the
corresponding author on reasonable request.
Authors’ contributions
FJ and QW conceived and designed the study. QY, YC
and XZ acquired and analyzed the data. WL and QL interpreted the
data and wrote the manuscript. FJ, QY and QW critically revised the
manuscript. All authors have read and approved the final
manuscript.
Ethics approval and consent to
participate
Tissues and information of patients were obtained
with written informed consent. Procedures involving patients in
this study were approved by the Human Ethics Committee at the Wujin
People’s Hospital of Changzhou. All the animal experiments in this
study were performed according to the National Institutes of Health
(Bethesda, MD, USA) guidelines and were approved by the
Institutional Animal Care and Treatment Committee of Wujin People’s
Hospital of Changzhou.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Tsao AS, Scagliotti GV, Bunn PA Jr,
Carbone DP, Warren GW, Bai C, de Koning HJ, Yousaf-Khan AU,
McWilliams A, Tsao MS, et al: Scientific Advances in Lung Cancer
2015. J Thorac Oncol. 11:613–638. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Gupta GP and Massagué J: Cancer
metastasis: Building a framework. Cell. 127:679–695. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Hsiao SH, Chung CL, Chou YT, Lee HL, Lin
SE and Liu HE: Identification of subgroup patients with stage
IIIB/IV non-small cell lung cancer at higher risk for brain
metastases. Lung Cancer. 82:319–323. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Torre LA, Bray F, Siegel RL, Ferlay J,
Lortet-Tieulent J and Jemal A: Global cancer statistics, 2012. CA
Cancer J Clin. 65:87–108. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Ardizzoni A, Tiseo M, Boni L, Di Maio M,
Buffoni L, Belvedere O, Grossi F, D’Alessandro V, de Marinis F,
Barbera S, et al: Randomized phase III PITCAP trial and
meta-analysis of induction chemotherapy followed by thoracic
irradiation with or without concurrent taxane-based chemotherapy in
locally advanced NSCLC. Lung Cancer. 100:30–37. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Zhao Z, Su Z, Zhang W, Luo M, Wang H and
Huang L: A randomized study comparing the effectiveness of
microwave ablation radioimmunotherapy and postoperative adjuvant
chemoradiation in the treatment of non-small cell lung cancer. J
BUON. 21:326–332. 2016.PubMed/NCBI
|
|
7
|
Hall RD, Le TM, Haggstrom DE and Gentzler
RD: Angiogenesis inhibition as a therapeutic strategy in non-small
cell lung cancer (NSCLC). Transl Lung Cancer Res. 4:515–523.
2015.PubMed/NCBI
|
|
8
|
John B, Enright AJ, Aravin A, Tuschl T,
Sander C and Marks DS: Human MicroRNA targets. PLoS Biol.
2:e3632004. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Tutar L, Tutar E, Özgür A and Tutar Y:
Therapeutic Targeting of microRNAs in Cancer: Future Perspectives.
Drug Dev Res. 76:382–388. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Sethi S, Ali S, Sethi S and Sarkar FH:
MicroRNAs in personalized cancer therapy. Clin Genet. 86:68–73.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Monroig PC, Chen L, Zhang S and Calin GA:
Small molecule compounds targeting miRNAs for cancer therapy. Adv
Drug Deliv Rev. 81:104–116. 2015. View Article : Google Scholar
|
|
12
|
Kane NM, Thrasher AJ, Angelini GD and
Emanueli C: Concise review: MicroRNAs as modulators of stem cells
and angiogenesis. Stem Cells. 32:1059–1066. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Vira D, Basak SK, Veena MS, Wang MB, Batra
RK and Srivatsan ES: Cancer stem cells, microRNAs, and therapeutic
strategies including natural products. Cancer Metastasis Rev.
31:733–751. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Sun X, Jiao X, Pestell TG, Fan C, Qin S,
Mirabelli E, Ren H and Pestell RG: MicroRNAs and cancer stem cells:
The sword and the shield. Oncogene. 33:4967–4977. 2014. View Article : Google Scholar
|
|
15
|
Liu Y, Li M, Zhang G and Pang Z:
MicroRNA-10b overexpression promotes non-small cell lung cancer
cell proliferation and invasion. Eur J Med Res. 18:412013.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Ambros V: The functions of animal
microRNAs. Nature. 431:350–355. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Bartel DP: MicroRNAs: Genomics,
biogenesis, mechanism, and function. Cell. 116:281–297. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Ni R, Huang Y and Wang J: miR-98 targets
ITGB3 to inhibit proliferation, migration, and invasion of
non-small-cell lung cancer. Onco Targets Ther. 8:2689–2697.
2015.PubMed/NCBI
|
|
19
|
Jiang P, Wu X, Wang X, Huang W and Feng Q:
NEAT1 upregulates EGCG-induced CTR1 to enhance cisplatin
sensitivity in lung cancer cells. Oncotarget. 7:43337–43351.
2016.PubMed/NCBI
|
|
20
|
Siragam V, Rutnam ZJ, Yang W, Fang L, Luo
L, Yang X, Li M, Deng Z, Qian J, Peng C, et al: MicroRNA miR-98
inhibits tumor angiogenesis and invasion by targeting activin
receptor-like kinase-4 and matrix metalloproteinase-11. Oncotarget.
3:1370–1385. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Jinushi T, Shibayama Y, Kinoshita I,
Oizumi S, Jinushi M, Aota T, Takahashi T, Horita S, Dosaka-Akita H
and Iseki K: Low expression levels of microRNA-124-5p correlated
with poor prognosis in colorectal cancer via targeting of SMC4.
Cancer Med. 3:1544–1552. 2014. View
Article : Google Scholar : PubMed/NCBI
|
|
22
|
Yang G, Zhang X and Shi J: MiR-98 inhibits
cell proliferation and invasion of non-small cell carcinoma lung
cancer by targeting PAK1. Int J Clin Exp Med. 8:20135–20145.
2015.
|
|
23
|
Chen X, Xu Y, Liao X, Liao R, Zhang L, Niu
K, Li T, Li D, Chen Z, Duan Y, et al: Plasma miRNAs in predicting
radiosensitivity in non-small cell lung cancer. Tumour Biol.
37:11927–11936. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Zhou H, Huang Z, Chen X and Chen S: miR-98
inhibits expression of TWIST to prevent progression of non-small
cell lung cancers. Biomed Pharmacother. 89:1453–1461. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Pasche B, Pennison MJ, Jimenez H and Wang
M: TGFBR1 and cancer susceptibility. Trans Am Clin Climatol Assoc.
125:300–312. 2014.PubMed/NCBI
|
|
26
|
Wang H, Zhang Q, Wang B, Wu W, Wei J, Li P
and Huang R: miR-22 regulates C2C12 myoblast proliferation and
differentiation by targeting TGFBR1. Eur J Cell Biol. 97:257–268.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Yu F, Chen B, Fan X, Li G, Dong P and
Zheng J: Epigenetically-Regulated MicroRNA-9-5p Suppresses the
Activation of Hepatic Stellate Cells via TGFBR1 and TGFBR2. Cell
Physiol Biochem. 43:2242–2252. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Cheng R, Dang R, Zhou Y, Ding M and Hua H:
MicroRNA-98 inhibits TGF-β1-induced differentiation and collagen
production of cardiac fibroblasts by targeting TGFBR1. Hum Cell.
30:192–200. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Rosman DS, Phukan S, Huang CC and Pasche
B: TGFBR1*6A enhances the migration and invasion of MCF-7 breast
cancer cells through RhoA activation. Cancer Res. 68:1319–1328.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Zhou R, Huang Y, Cheng B, Wang Y and Xiong
B: TGFBR1*6A is a potential modifier of migration and invasion in
colorectal cancer cells. Oncol Lett. 15:3971–3976. 2018.PubMed/NCBI
|
|
31
|
Mody HR, Hung SW, Pathak RK, Griffin J,
Cruz-Monserrate Z and Govindarajan R: miR-202 diminishes TGFβ
receptors and attenuates TGFβ1-Induced EMT in pancreatic cancer.
Mol Cancer Res. 15:1029–1039. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
33
|
Lee YS and Dutta A: MicroRNAs in cancer.
Annu Rev Pathol. 4:199–227. 2009. View Article : Google Scholar :
|
|
34
|
Berindan-Neagoe I, Monroig PC, Pasculli B
and Calin GA: MicroRNAome genome: A treasure for cancer diagnosis
and therapy. CA Cancer J Clin. 64:311–336. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Akhurst RJ and Hata A: Targeting the TGFβ
signalling pathway in disease. Nat Rev Drug Discov. 11:790–811.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Bierie B and Moses HL: Tumour
microenvironment: TGFbeta: the molecular Jekyll and Hyde of cancer.
Nat Rev Cancer. 6:506–520. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Sethi N, Dai X, Winter CG and Kang Y:
Tumor-derived JAGGED1 promotes osteolytic bone metastasis of breast
cancer by engaging notch signaling in bone cells. Cancer Cell.
19:192–205. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Kang Y, He W, Tulley S, Gupta GP,
Serganova I, Chen CR, Manova-Todorova K, Blasberg R, Gerald WL and
Massagué J: Breast cancer bone metastasis mediated by the Smad
tumor suppressor pathway. Proc Natl Acad Sci USA. 102:13909–13914.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Attili I, Karachaliou N, Bonanno L,
Berenguer J, Bracht J, Codony-Servat J, Codony-Servat C, Ito M and
Rosell R: STAT3 as a potential immunotherapy biomarker in
oncogene-addicted non-small cell lung cancer. Ther Adv Med Oncol.
Apr 2–2018.Epub ahead of print. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Reis H, Metzenmacher M, Goetz M, Savvidou
N, Darwiche K, Aigner C, Herold T, Eberhardt WE, Skiba C, Hense J,
et al: MET expression in advanced non-small-cell lung cancer:
Effect on clinical outcomes of chemotherapy, targeted therapy, and
immunotherapy. Clin Lung Cancer. 19:e441–e463. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Yang H, Yan L, Sun K, Sun X, Zhang X, Cai
K and Song T: LncRNA BCAR4 increases viability, invasion and
migration non-small cell lung cancer cells by targeting
glioma-associated oncogene 2 (GLI2). Oncol Res. Apr 3–2018.Epub
ahead of print. View Article : Google Scholar
|
|
42
|
Ambros V: MicroRNA pathways in flies and
worms: Growth, death, fat, stress, and timing. Cell. 113:673–676.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Liu W, Xiao P, Wu H, Wang L, Kong D and Yu
F: MicroRNA-98 plays a suppressive role in non-small cell lung
cancer through inhibition of SALL4 protein expression. Oncol Res.
25:975–988. 2017. View Article : Google Scholar
|
|
44
|
Wang MJ, Zhang H, Li J and Zhao HD:
microRNA-98 inhibits the proliferation, invasion, migration and
promotes apoptosis of breast cancer cells by binding to HMGA2.
Biosci Rep. 38:BSR201805712018. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Cai C, Huo Q, Wang X, Chen B and Yang Q:
SNHG16 contributes to breast cancer cell migration by competitively
binding miR-98 with E2F5. Biochem Biophys Res Commun. 485:272–278.
2017. View Article : Google Scholar : PubMed/NCBI
|