Introduction
Non-small cell lung cancer (NSCLC) remains the
leading cause of mortality from cancer, and surgery combined with
chemotherapy has increased the long-term survival rate of patients
with NSCLC (1). Chemotherapy is
effective by triggering the death of tumor cells, but the
occurrence of drug resistance can limit its efficacy (2). Therefore, seeking a therapeutic
approach for overcoming drug resistance is vital for the treatment
of NSCLC. Autophagy is a process of self-eating in cells, and can
remove toxic substances in cancer cells and is involved in cellular
degradation to maintain homeostasis (3). Studies have revealed that the ability
of autophagy to maintain homeostasis in cells has led to
chemoresistance in a number of types of cancer, including
osteosarcoma, bladder cancer and gastric cancer (4-6).
Therefore, autophagy may be a novel target for overcoming drug
resistance.
Autophagy-related protein 7 (ATG7) is an E1-like
enzyme and consists of the N-terminal domain and the C-terminal
domain (7,8). It can activate autophagy-essential
ubiquitin-like proteins ATG8 and ATG12, thus serving crucial
functions in autophagy (8,9). It has been identified that
ATG7-induced autophagy enhances the production of neural crest
cells by modifying the cell cycle (10). In addition, studies have indicated
that ATG7-dependent autophagy modulates the progression of a number
of types of cancer (11-13). ATG7 may be an important molecular
target in the study of chemo-resistance of cancer through the
involvement in different signaling pathways (14,15).
Sun et al (16) identified
that the long non-coding RNA (lncRNA) X-inactive specific
transcript knockdown decreased chemoresistance of NSCLC cells by
inhibiting autophagy, and ATG7 overexpression enhanced the
chemoresistance of A549/DDP cells, which suggested that ATG7
enhanced chemoresistance of NSCLC cells to cisplatin (DDP) by
promoting autophagy.
MicroRNAs (miRNAs/miR) are a type of small
non-coding RNAs that regulate gene expression at the
post-transcriptional level by binding to the 3′-untranslated region
in a variety of biological and pathological processes (17). It has been identified that miRNAs
serve a vital function in regulating chemoresis-tance of NSCLC
cells (18,19). Zhao et al (20) identified that miR-17 was
downregulated in A549/DDP cells compared with in A549 cells, and
decreased miR-17 expression maintained cisplatin resistance in
NSCLC. Comincini et al (21) identified that ATG7 was a potential
target of miR-17, and miR-17 was able to negatively regulate ATG7,
thus improving the sensitivity to temozolomide in human
glioblastoma cells. Therefore, miR-17/ATG7 may be involved in the
regulation of chemoresistance of NSCLC cells.
LncRNAs are a class of non-coding RNAs of >200
nucleotides. Recently, it has been revealed that several lncRNAs
regulate chemoresistance of NSCLC cells through different pathways
(22,23). Ye et al (24) identified that, compared with
adjacent normal tissues, the lncRNA bladder cancer-associated
transcript 1 (BLACAT1) was upregulated in NSCLC tissues, and
inhibition of BLACAT1 suppressed NSCLC cell proliferation,
migration and invasion (24).
Furthermore, BLACAT1 was able to promote oxaliplatin resistance of
gastric cancer via the miR-361/ATP-binding cassette transporter B1
pathway (25). Therefore, we
hypothesized that lncRNA BLACAT1 may serve a function in promoting
the chemoresistance of NSCLC cells. The bioinformatics software
LncBase (version 2) was used to predict the binding sites between
BLACAT1 and miR-17. Therefore, we hypothesized that BLACAT1
promoted autophagy through miR-17/ATG7 to enhance the
chemoresistance of NSCLC cells.
In the present study, it was identified that BLACAT1
was upregulated in cisplatin (DDP)-resistant NSCLC cells, and
confirmed the interaction of lncRNA BLACAT1 and miR-17. In
addition, lncRNA BLACAT1 facilitated autophagy through the
miR-17/ATG7 signaling pathway, thus promoting chemoresistance of
NSCLC cells.
Materials and methods
Cell culture and transfection
DDP-sensitive cell lines (A549 and H1299) and
DDP-resistant cell lines (A549/DDP and H1299/DDP) were purchased
from the Cell Bank of the Chinese Academy of Sciences (Shanghai,
China). All cell lines were cultured in RPMI-1640 medium (Gibco;
Thermo Fisher Scientific, Inc., Waltham, MA, USA) supplemented with
10% fetal bovine serum (Gibco; Thermo Fisher Scientific, Inc.), 2
µmol/l L-glutamine, 100 U/ml penicillin and 100 µg/ml
streptomycin at 37°C under humidified conditions containing 5%
CO2. In order to maintain DDP resistance of A549/DDP and
H1299/DDP cells, cells were cultured with 5 µg/ml DDP.
For BLACAT1 knockdown, short interfering RNA
targeting BLACAT1 (si-BLACAT1, 5′-CCAGTGCATGGT CCTTGACTTT-3′) was
synthesized by Shanghai GeneChem Co., Ltd. (Shanghai, China).
pcDNA-BLACAT1 was constructed by inserting BLACAT1 cDNA into
pcDNA3.1 (Invitrogen; Thermo Fisher Scientific, Inc.). miR-17 mimic
(5′-CAAAGUGCUUACAGUGCAGGUAG-3′), miR-17 inhibitor
(5′-GUUUCACGAAUGUCACGUCCAUC-3′) and scrambled miRNA control were
synthesized by Shanghai GeneChem Co., Ltd. Cells were seeded at
6×103 cells/well in a 6-well plate and cultured until
reaching 50% confluence, prior to transfection with 70 nmol
pcDNA-BLACAT1, si-BLACAT1, miR-17 mimic, miR-17 inhibitor or
negative control vectors using Lipofectamine® 2000
transfection reagent (Invitrogen; Thermo Fisher Scientific, Inc.),
according to the manufacturer’s protocol. Following transfection,
the autophagy inhibitor 3-methyladenine (3-MA; 1 mM) was used to
treat A549 cells for 2 h. The autophagy agonist rapamycin (100 nM)
was used to treat A549/DDP cells for 4 h.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
Total RNA was extracted from A549, H1299, A549/DDP
and H1299/DDP cells using TRIzol® reagent (Invitrogen;
Thermo Fisher Scientific, Inc.). A High Capacity cDNA Reverse
Transcription kit (Applied Biosystems; Thermo Fisher Scientific,
Inc.) was used for cDNA synthesis, according to the manufacturer’s
protocol. A SuperScript™ III Platinum™ SYBR™ Green One-Step qPCR
kit (Invitrogen; Thermo Fisher Scientific, Inc.) was used,
according to the manufacturer’s protocol, to determine BLACAT1 and
miR-17 expression. RT-qPCR was conducted for 35 cycles of a
denaturing phase at 94°C for 1 min, primer annealing at 60°C for
1.5 min, and an extension at 72°C for 2 min in a QuantStudio 3
Real-Time PCR System (Applied Biosystems; Thermo Fisher Scientific,
Inc.). Primers were provided as follows: BLACAT1,
5′-GACAAAGCACAAGCGAAACAAG-3′ (forward) and
5′-GGACATCTGATAGCCTGGTGAC-3′ (reverse); GAPDH,
5′-TGCACCACCAACTGCTTAGC-3′ (forward) and 5′-GGCATG
GACTGTGGTCATGAG-3′ (reverse); miR-17, 5′-CAGTAA
AGGTAAGGAGAGCTCAATCTG-3′ (forward) and 5′-CAT
ACAACCACTAAGCTAAAGAATAATCTGA-3′ (reverse); U6,
5′-CTCGCTTCGGCAGCACA-3′ (forward) and 5′-AAC GCTTCACGAATTTGCGT-3′
(reverse). The relative expression of BLACAT1 and miR-17 was
analyzed using the 2−ΔΔCq method (26).
Xenograft model in nude mice
Male nude mice (BALB/c nu/nu; 5-week-old; 16-20 g)
were purchased from the laboratory animal center of Zhengzhou
University (Zhengzhou, China), and were housed in 12-h light/12-h
dark sterile conditions (26-28°C, relative humidity 40-60%) with no
limitation to water and food. Lenti-si-control or lenti-si-BLACAT1
was synthesized by Shanghai GeneChem Co., Ltd. and transfected into
A549/DDP cells. A 100 µl volume of PBS containing
1×107 A549/DDP cells was subcutaneously injected into
the nude mice (7 mice in each group). Tumor growth was determined
using a caliper every 7 days. Mice were sacrificed at 28 days after
inoculation, and tumor tissues were collected. The animal
experiment was approved by the Ethics Committee of The First
Affiliated Hospital of Zhengzhou University.
Western blotting
Cells and tumors were collected and lysed in
radioimmunoprecipitation assay buffer (Beyotime Institute of
Biotechnology, Haimen, China). The concentration of proteins was
determined using a Bicinchoninic Acid Protein assay kit (Beyotime
Institute of Biotechnology). Equal amounts (25 g) of protein of
each sample were separated by SDS-PAGE (12% gel) and transferred
onto a polyvinylidene difluoride membrane (Invitrogen; Thermo
Fisher Scientific, Inc.). The membrane was incubated with primary
antibodies against ATG7 (1:10,000; cat. no. ab133528; Abcam,
Cambridge, MA, USA), multidrug-resistance protein 1 (MRP1; 1:50;
cat. no. ab32574; Abcam), light chain 3 (LC3; 1:3,000; cat. no.
ab51520; Abcam), Beclin 1 (1:2,000; cat. no. ab207612; Abcam) and
β-actin (1:1,000; cat. no. ab8266; Abcam), and horseradish
peroxidase-conjugated secondary antibody [goat anti-rabbit
immunoglobulin G (IgG); 1:2,000; cat. no. ab97051; Abcam]. Blots
were visualized using a ChemiDoc™ MP Imaging system (Bio-Rad
Laboratories, Inc., Hercules, CA, USA).
MTT assay
A549 or H1299 cells (6×104 cells/ml) were
seeded on a 96-well plate and incubated with various concentrations
of DDP (0, 1, 2, 4 and 8 µg/ml). A549/DDP or H1299/DDP cells
(6×104 cells/ml) were incubated with various
concentrations of DDP (0, 4, 8, 16 and 32 µg/ml). Following
incubation at 37°C for 24 h, MTT solution (20 µl, 5 mg/ml)
was added to each well in the dark prior to further incubation at
37°C for 4 h. Cells were lysed using dimethylsulfoxide (150
µl/well), and the reduction of MTT was quantified by
determining the absorbance at 540 nm against a reference wavelength
of 630 nm using a plate reader (Bio-Rad Laboratories, Inc.).
RNA immunoprecipitation (RIP)
The bioinformatics software LncBase (version 2;
diana.imis.athena-innovation. gr/DianaTools/index.php) was used to
predict the binding sites between BLACAT1 and miR-17. A RIP assay
was performed using a Magna RIP™ RNA-Binding Protein
Immunoprecipitation kit (EMD Millipore, Billerica, MA, USA). A549
cell lysate was prepared from 1.5×107 cells using 100
µl RIP lysis buffer supplemented with 0.25 µl RNase
inhibitor and 0.5 µl protease inhibitor. The cell lysate was
centrifuged at 513 × g for 15 min at 4°C, and the supernatant was
incubated with RIP buffer containing Protein A/G-Sepharose beads
conjugated with anti-Argonaute 2 (AGO2) antibody (1:5,000; cat. no.
03-110; EMD Millipore) or negative control IgG (1:5,000; cat. no.
12-371; EMD Millipore). RNA-bound protein complex was obtained, and
RT-qPCR was used to detect lncRNA BLACAT1 and miR-17 in the
precipitates, as aforementioned.
RNA pull-down
The biotin-labeled lncRNA BLACAT1 was transcribed
with the Biotin RNA Labeling mixture (Roche Diagnostics, Basel,
Switzerland) and T7 RNA polymerase (Roche Diagnostics). A549 cell
extract was prepared from 1.5×107 cells in RIP buffer,
then mixed with biotin-labeled lncRNA BLACAT1 at 4°C for 1 h. Beads
were added prior to incubation at room temperature for 1 h. Western
blotting was used to detect AGO2 in the BLACAT1 pull-down complex,
and RT-qPCR was used to detect miR-17 in the precipitates, as
aforementioned.
Statistical analysis
All experiments were performed in triplicate. All
data are expressed as the mean ± standard deviation and analyzed
using SPSS software (version 17.0; SPSS, Inc., Chicago, IL, USA).
The differences between groups were assessed by Student’s t-test or
one-way analysis of variance. P<0.05 was considered to indicate
a statistically significant difference.
Results
LncRNA BLACAT1 is upregulated in
DDP-resistant NSCLC cells
To determine the expression of BLACAT1, miR-17 and
ATG7 in NSCLC cells, RT-qPCR and western blot analysis were
performed. As presented in Fig.
1A, lncRNA BLACAT1 was upregulated and miR-17 was downregulated
in DDP-resistant cell lines (A549/DDP and H1299/DDP) compared with
in the DDP-sensitive cell lines (A549 and H1299). Protein levels of
ATG7 and MRP1 were upregulated in A549/DDP and H1299/DDP cell lines
compared with in A549 and H1299 cell lines (Fig. 1B). The levels of the
autophagy-associated proteins LC3-II/LC3-I and Beclin 1 were
investigated further, and it was identified that the LC3-II/LC3-I
ratio and Beclin 1 level were increased in A549/DDP and H1299/DDP
cell lines (Fig. 1C).
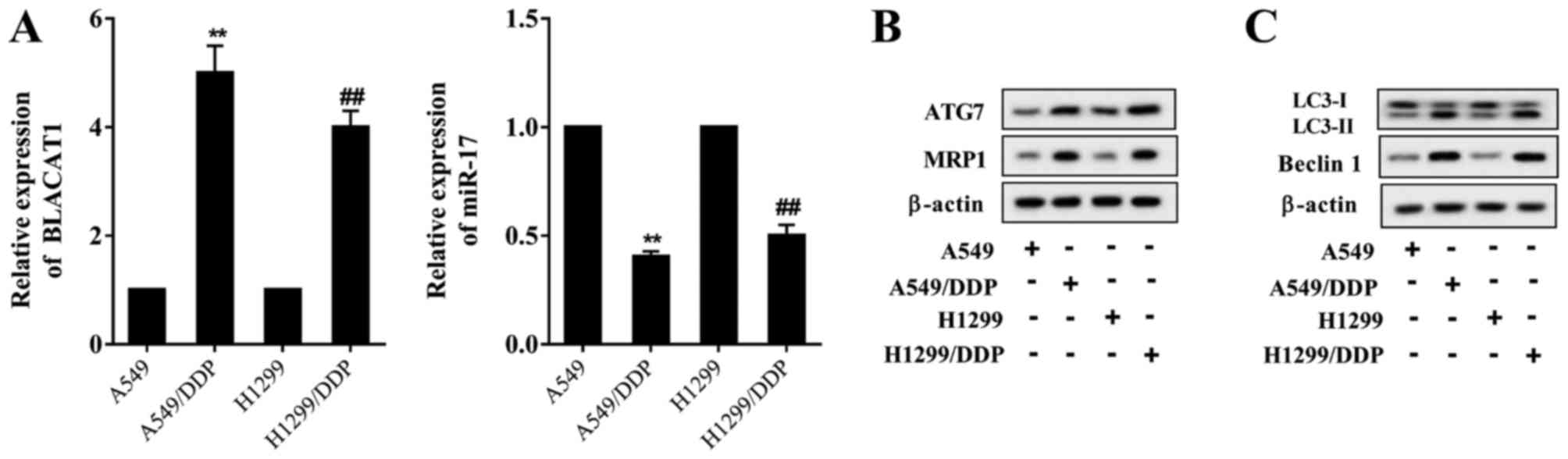 | Figure 1LncRNA BLACAT1 is upregulated in
DDP-resistant NSCLC cells. (A) Compared with the DDP-sensitive cell
lines A549 and H1299, lncRNA BLACAT1 was upregulated in the
DDP-resistant cell lines A549/DDP and H1299/DDP, and miR-17 was
downregulated in the DDP-resistant cell lines A549/DDP and
H1299/DDP. (B) Protein levels of ATG7 and MRP1 were upregulated in
the DDP-resistant cell lines A549/DDP and H1299/DDP. (C) The
autophagy-associated protein LC3-II/LC3-I ratio was increased in
the DDP-resistant cell lines A549/DDP and H1299/DDP, and the Beclin
1 level was also increased. **P<0.01 vs. A549;
##P<0.01 vs. H1299. BLACAT1, bladder
cancer-associated transcript 1; lncRNA, long non-coding RNA; DDP,
cisplatin; miR, microRNA; ATG7, autophagy-related protein 7; MRP1,
multidrug-resistance protein 1; LC3, light chain 3. |
lncRNA BLACAT1 is involved in the
viability of DDP-resistant NSCLC cells
To investigate the function of BLACAT1 in the
viability of NSCLC cells, pcDNA-BLACAT1 and si-BLACAT1 were used
for BLACAT1 overexpression and BLACAT1 inhibition, respectively.
A549 and H1299 cells were transfected with pcDNA or pcDNA-BLACAT1,
and treated with DDP (0, 1, 2, 4 and 8 g/ml) for 24 h. A549/DDP and
H1299/DDP cells were transfected with si-control or si-BLACAT1 and
treated with DDP (0, 4, 8, 16 and 32 µg/ml) for 24 h. It was
observed that cell viability of A549 and H1299 cells was
significantly increased in the pcDNA-BLACAT1 group compared with in
the pcDNA group (Fig. 2A). The
viability of A549/DDP and H1299/DDP cells was significantly
decreased in the si-BLACAT1 group compared with in the si-control
group (Fig. 2B).
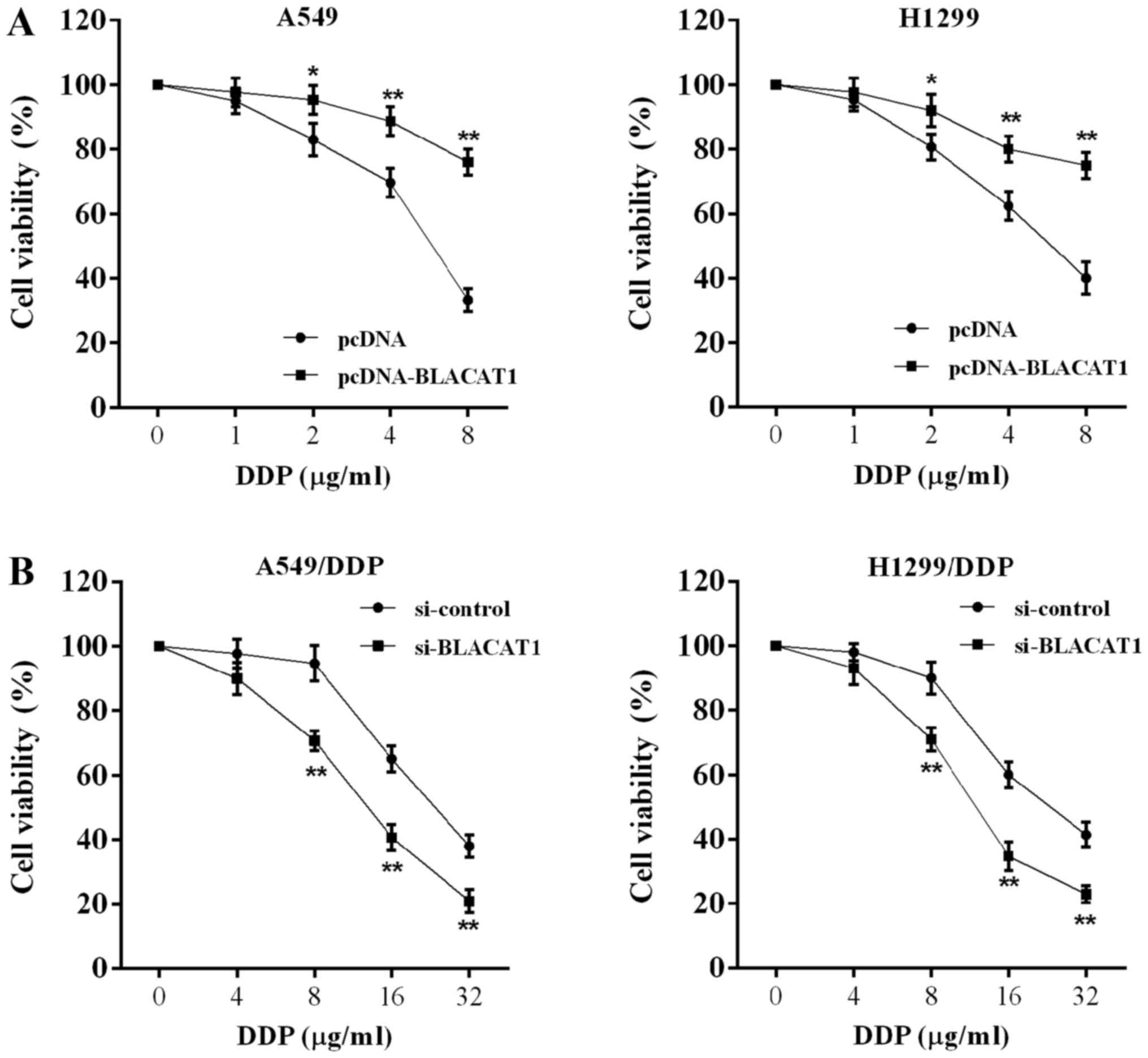 | Figure 2LncRNA BLACAT1 is involved in the
viability of DDP-resistant NSCLC cells. A549 and H1299 cells were
transfected with pcDNA or pcDNA-BLACAT1, and treated with DDP (0,
1, 2, 4 and 8 µg/ml) for 24 h. A549/DDP and H1299/DDP cells
were transfected with si-control or si-BLACAT1 and treated with DDP
(0, 4, 8, 16 and 32 µg/ml) for 24 h. (A) Viability of A549
and H1299 cells with lncRNA BLACAT1 overexpression was
significantly increased. (B) Viability of A549/DDP and H1299/DDP
cells with BLACAT1 inhibition was significantly decreased.
*P<0.05, **P<0.01 vs. pcDNA or
si-control. BLACAT1, bladder cancer-associated transcript 1;
lncRNA, long non-coding RNA; DDP, cisplatin; si, short interfering
RNA. |
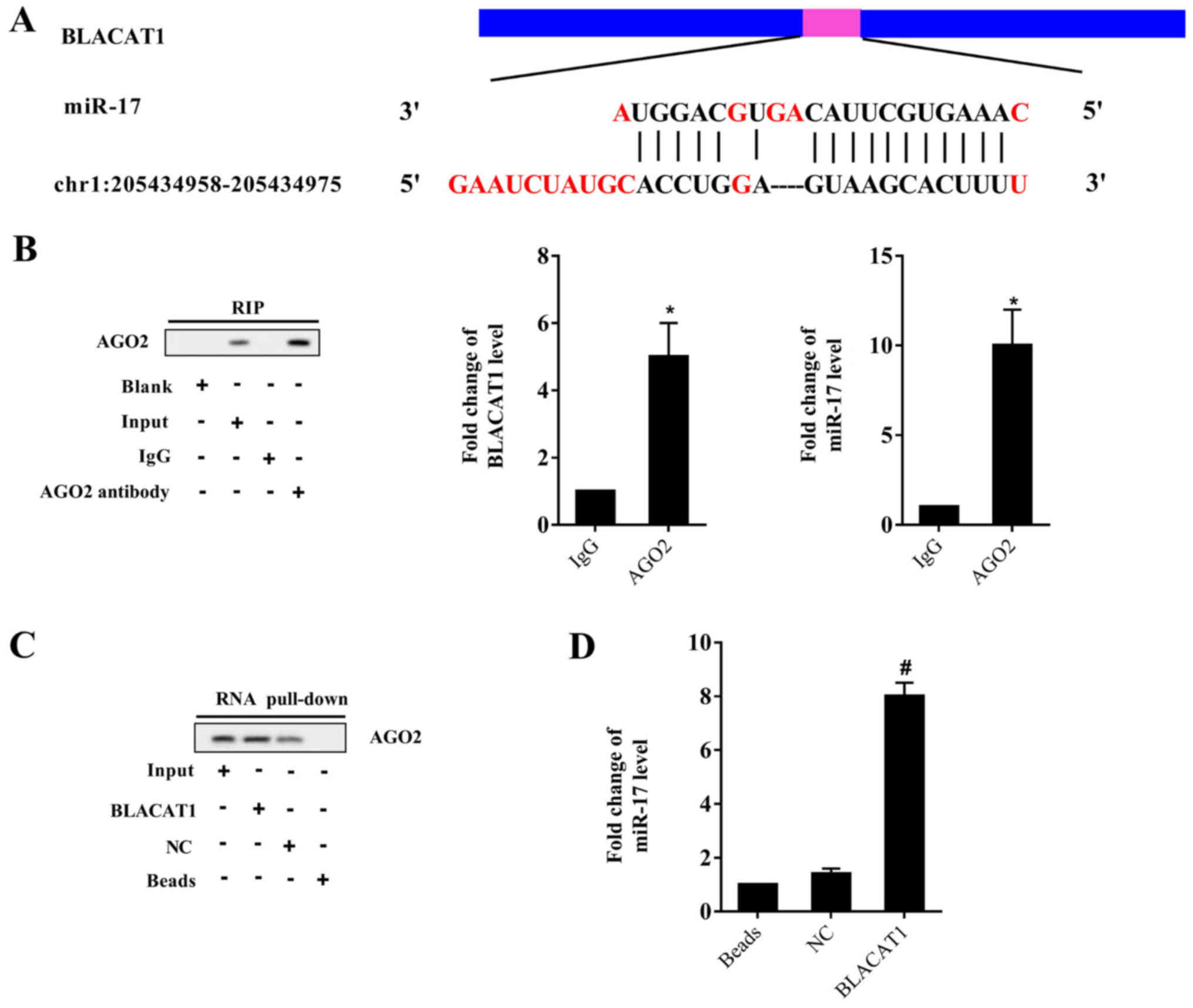
Interaction between lncRNA BLACAT1 and
miR-17
According to the bioinformatic software LncBase,
there were binding sites between BLACAT1 and miR-17 (Fig. 3A). On the basis of the opposing
expression of BLACAT1 and miR-17 in NSCLC cells in Fig. 1, we hypothesized that BLACAT1 may
directly inhibit miR-17. The RIP assay identified that BLACAT1 and
miR-17 were relatively enriched in AGO2 compared with IgG (Fig. 3B). The RNA pull-down detected AGO
in the BLACAT1 pull-down complex (Fig.
3C), and miR-17 was identified to be accumulated in the BLACAT1
pull-down complex, compared with only a slight increase in NC
(Fig. 3D). These results indicated
that BLACAT1 interacted with miR-17, and could negatively regulate
miR-17.
BLACAT1 promotes ATG7 expression through
miR-17
A previous study identified that ATG7 was a
potential target of miR-17, and miR-17 could negatively regulate
ATG7 in human glioblastoma cells (21). On the basis of the consistent
expression of BLACAT1 and ATG7, we hypothesized that BLACAT1 could
positively regulate ATG7 expression through miR-17. As presented in
Fig. 4A. miR-17 expression in A549
cells was significantly decreased following BLACAT1 overexpression,
and miR-17 mimic eliminated the inhibitory effect of BLACAT1
overexpression. The ATG7 protein level in A549 cells was
upregulated following BLACAT1 overexpression, and miR-17 mimic
eliminated the promotional effect of BLACAT1 overexpression
(Fig. 4B). Following si-BLACAT1
transfection, miR-17 expression in A549/DDP cells was significantly
increased, and miR-17 inhibitor eliminated the promotional effect
of si-BLACAT1 (Fig. 4C). The ATG7
protein level in A549/DDP cells was downregulated following
si-BLACAT1, and miR-17 inhibitor eliminated the inhibitory effect
of si-BLACAT1 (Fig. 4D).
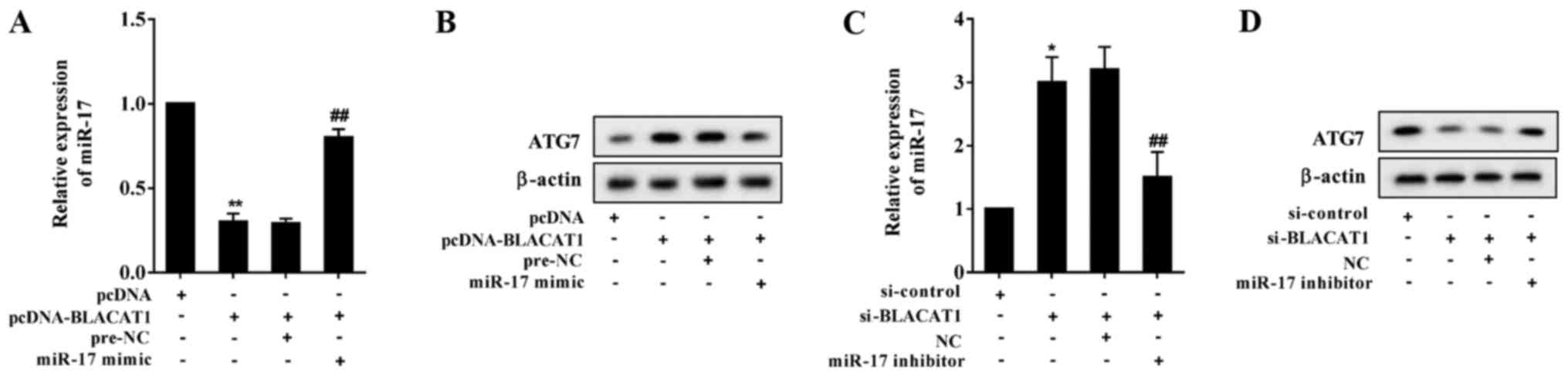 | Figure 4BLACAT1 promotes ATG7 expression
through miR-17. A549 cells were divided into pcDNA, pcDNA-BLACAT1,
pcDNA-BLACAT1+pre-NC and pcDNA-BLACAT1+miR-17 mimic groups. (A)
miR-17 expression in A549 cells was significantly decreased
following BLACAT1 overexpression, and miR-17 mimic eliminated the
inhibitory effect of BLACAT1 overexpression. **P<0.01
vs. pcDNA; ##P<0.01 vs. pcDNA-BLACAT1+pre-NC. (B)
ATG7 protein level in A549 cells was upregulated following BLACAT1
overexpression, and miR-17 mimic eliminated the promotional effect
of BLACAT1 overexpression. A549/ DP cells were divided into
si-control, si-BLACAT1, si-BLACAT1+NC and si-BLACAT1+miR-17
inhibitor groups. (C) miR-17 expression in A549/ DP cells was
significantly increased following si-BLACAT1 treatment, and miR-17
inhibitor eliminated the promotional effect of si-BLACAT1.
*P<0.05 vs. si-control; ##P<0.01 vs.
si-BLACAT1+NC. (D) ATG7 protein level in A549/DDP cells was
downregulated following si-BLACAT1 treatment, and miR-17 inhibitor
eliminated the inhibitory effect of si-BLACAT1. BLACAT1, bladder
cancer-associated transcript 1; ATG7, autophagy-related protein 7;
miR, microRNA; NC, negative control; si, short interfering RNA. |
BLACAT1 facilitates autophagy through
miR-17
To investigate whether BLACAT1 activated autophagy
through miR-17, the LC3-I/LC3-II ratio and Beclin 1 expression were
determined. It was identified that treatment with autophagy
inhibitor 3-methyladenine (3-MA) suppressed autophagy of A549
cells. BLACAT1 overexpression promoted the conversion of LC3-I into
LC3-II, and facilitated Beclin 1 expression in A549 cells, whereas
miR-17 mimic eliminated the promotional effect of BLACAT1
overexpression (Fig. 5A).
Following treatment with the autophagy agonist rapamycin, autophagy
of A549/DDP cells was promoted. si-BLACAT1 inhibited the conversion
of LC3-I into LC3-II, and suppressed Beclin 1 expression in
A549/DDP cells, whereas miR-17 inhibitor eliminated the inhibitory
effect of si-BLACAT1 (Fig. 5B).
These results indicated that BLACAT1 upregulated
autophagy-associated proteins to activate autophagy through
inhibiting miR-17.
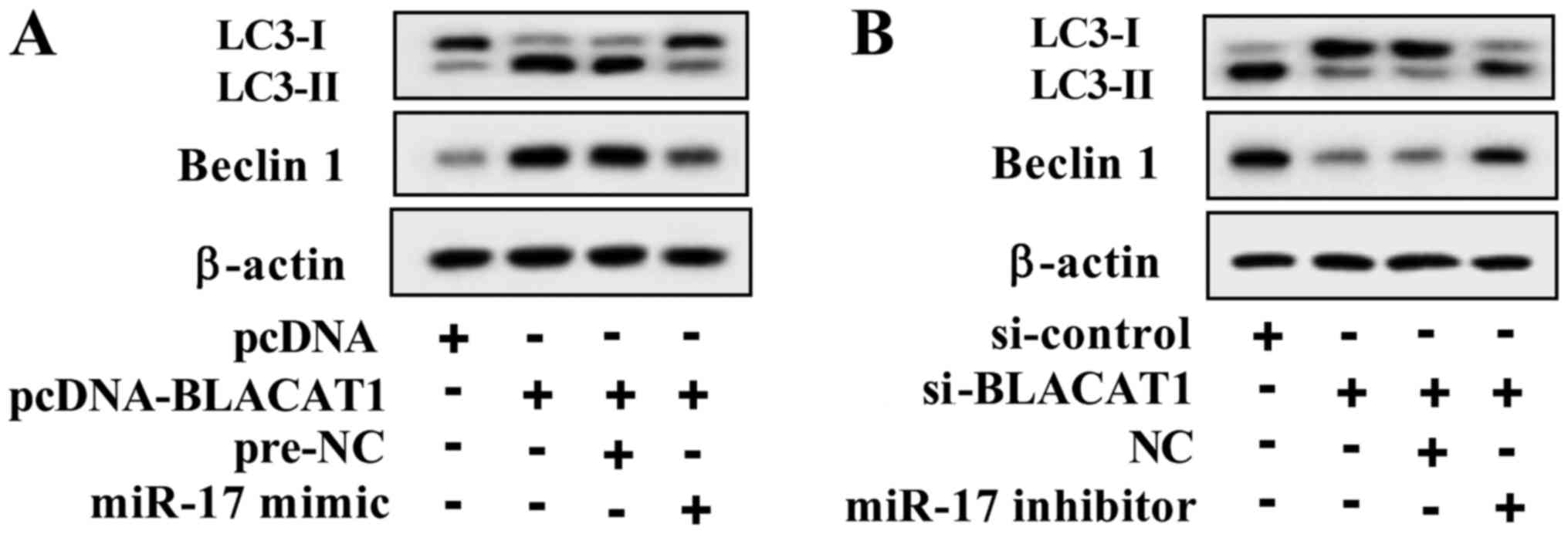 | Figure 5BLACAT1 facilitates autophagy through
miR-17. (A) A549 cells were transfected with pcDNA, pcDNA-BLACAT1,
pcDNA-BLACAT1+pre-NC and pcDNA-BLACAT1+miR-17 mimic groups. The
autophagy inhibitor 3-methyladenine (1 mM) was used to treat A549
cells for 2 h. BLACAT1 overexpression promoted the conversion of
LC3-I into LC3-II, and facilitated Beclin 1 expression, whereas
miR-17 mimic eliminated the promotional effect of BLACAT1
overexpression. (B) A549/DDP cells were transfected with
si-control, si-BLACAT1, si-BLACAT1+NC and si-BLACAT1+miR-17
inhibitor. The autophagy agonist rapamycin (100 nM) was used to
treat A549/DDP cells for 4 h. si-BLACAT1 inhibited the conversion
of LC3-I into LC3-II, and suppressed Beclin 1 expression, whereas
miR-17 inhibitor eliminated the inhibitory effect of si-BLACAT1.
BLACAT1, bladder cancer-associated transcript 1; miR, microRNA;
LC3, light chain 3; si, short interfering RNA; NC, negative
control; DDP, cisplatin. |
BLACAT1 promotes chemoresistance of NSCLC
cells through miR-17
The function of BLACAT1 in the chemoresistance of
NSCLC cells was investigated further. Following stimulation with
DDP, BLACAT1 overexpression promoted chemoresistance of A549 cells
and increased MRP1 expression in A549 cells, and miR-17 mimic
eliminated the promotional effect of BLACAT1 overexpression
(Fig. 6A). Following stimulation
with DDP, si-BLACAT1 inhibited chemoresistance of A549/DDP cells
and decreased MRP1 expression, and miR-17 inhibitor eliminated the
inhibitory effect of si-BLACAT1 (Fig.
6B).
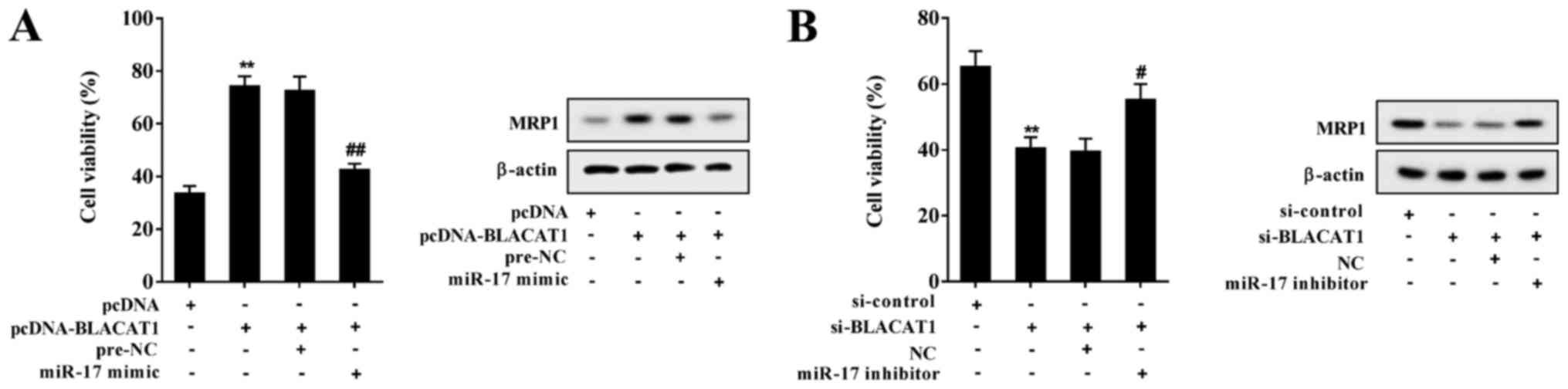 | Figure 6BLACAT1 promotes chemoresistance of
non-small cell lung cancer cells through miR-17. (A) A549 cells
were transfected with pcDNA, pcDNA-BLACAT1, pcDNA-BLACAT1+pre-NC
and pcDNA-BLACAT1+miR-17 mimic. A549 cells were stimulated with 8
µg/ml DDP. BLACAT1 overexpression promoted chemoresistance
of A549 cells and increased MRP1 expression, and miR-17 mimic
eliminated the promotional effect of BLACAT1 overexpression.
**P<0.01 vs. pcDNA; ##P<0.01 vs.
pcDNA-BLACAT1+pre-NC. (B) A549/DDP cells were transfected with
si-control, si-BLACAT1, si-BLACAT1+NC and si-BLACAT1+miR-17
inhibitor. A549/DDP cells were stimulated with 16 µg/ml DDP.
si-BLACAT1 inhibited chemoresistance of A549/DDP cells and
decreased MRP1 expression, and miR-17 inhibitor eliminated the
inhibitory effect of si-BLACAT1. **P<0.01 vs.
si-control; #P<0.05 vs. si-BLACAT1+NC. BLACAT1,
bladder cancer-associated transcript 1; NC, negative control; DDP,
cisplatin; MRP1, multidrug-resistant protein 1; miR, microRNA; si,
short interfering RNA. |
BLACAT1 inhibition ameliorates
chemoresistance of NSCLC in vivo
A549/DDP cells were transfected with
lenti-si-control or lenti-si-BLACAT1, and the effect of BLACAT1 on
chemo-resistance in NSCLC in vivo was observed. Tumor
volumes in mice of the lenti-si-BLACAT1 group were lower compared
with those in the lenti-si-control group (Fig. 7A). As presented in Fig. 7B, BLACAT1 inhibition suppressed
BLACAT1 and ATG7 expression, and promoted miR-17 expression of
tumor tissues in the xenograft mouse model. Furthermore, BLACAT1
inhibition suppressed conversion of LC3-I into LC3-II, and
inhibited Beclin 1 and MRP1 expression (Fig. 7C), which suggested that BLACAT1
inhibition ameliorated autophagy and chemoresistance in NSCLC.
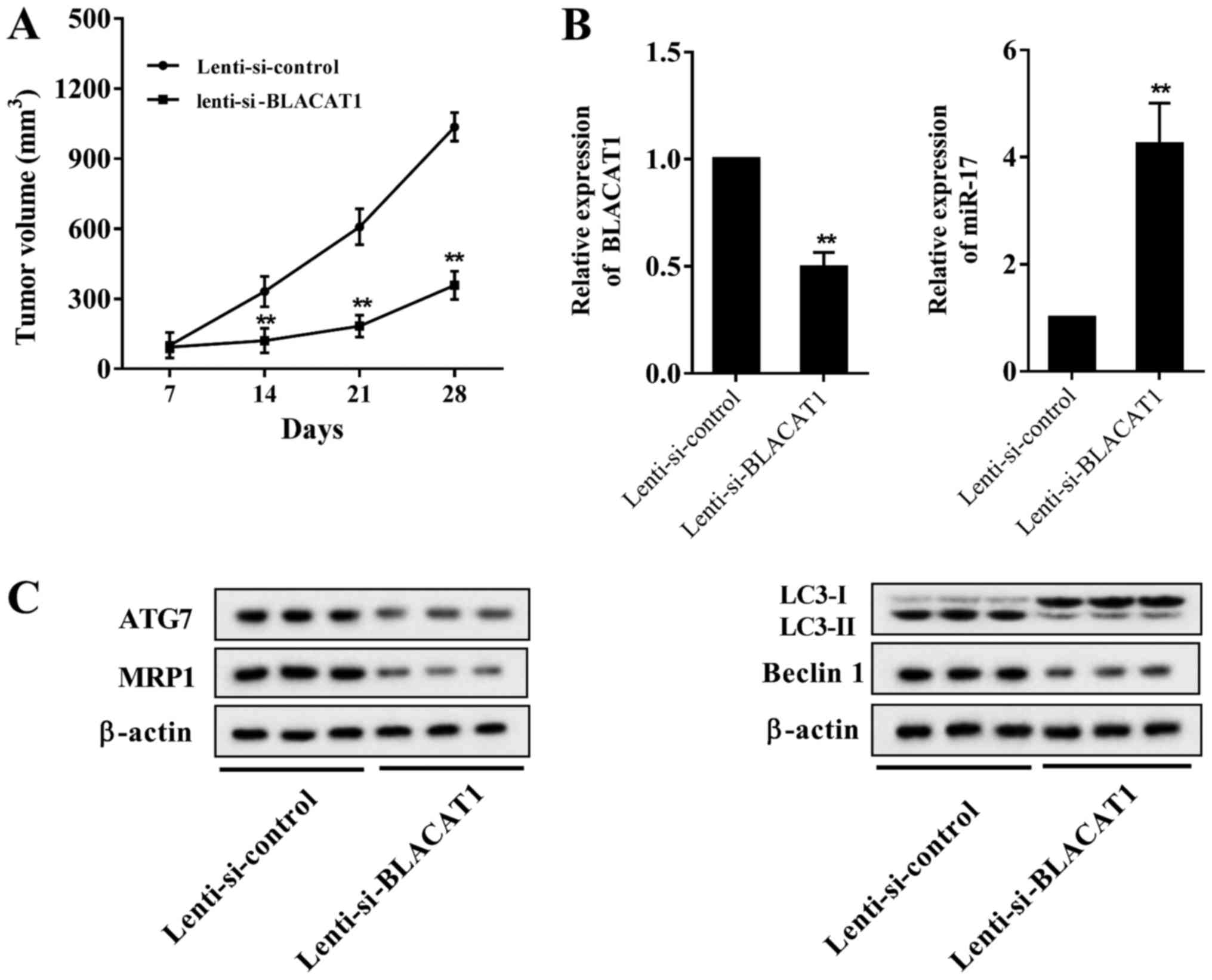 | Figure 7BLACAT1 inhibition ameliorates
chemoresistance of non-small cell lung cancer cells in vivo.
Lenti-si-control or lenti-si-BLACAT1 was transfected into A549/DDP
cells. A total of 1×107 A549/DDP cells was
subcutaneously injected into male nude mice (BALB/c nu/nu,
5-week-old). Tumor growth was measured using a caliper every 7
days. Mice were sacrificed 28 days after inoculation, and tumor
tissues were collected. (A) Tumor volumes in mice of
lenti-si-BLACAT1 group were lower compared with those of the
lenti-si-control group. (B) BLACAT1 inhibition suppressed BLACAT1
and ATG7 expression, and promoted miR-17 expression. (C) BLACAT1
inhibition suppressed the conversion of LC3-I into LC3-II, and
inhibited Beclin 1 and MRP1 expression. **P<0.01 vs.
lenti-si-control. BLACAT1, bladder cancer-associated transcript 1;
si, short interfering RNA; DDP, cisplatin; LC3, light chain 3; miR,
microRNA; ATG7, autophagy 7; MRP1, multidrug-resistance protein
1. |
Discussion
In the present study, it was identified that lncRNA
BLACAT1, ATG7, MRP1 LC3-II/LC3-I and Beclin 1 were significantly
upregulated in DDP-resistant NSCLC cells, whereas miR-17 was
downregulated in DDP-resistant NSCLC cells. Inhibition of BLACAT1
significantly decreased the viability of A549/DDP and H1299/DDP
cells following treatment with DDP. Furthermore, it was identified
that BLACAT1 interacted with miR-17, and BLACAT1 negatively
regulated miR-17. Inhibition of BLACAT1 decreased the LC3-II/LC3-I
ratio and Beclin 1 expression, and decreased the viability of
A549/DDP cells. In vivo experiments indicated that
inhibition of BLACAT1 suppressed autophagy in NSCLC.
LncRNAs exert an important function in a variety of
cancer types, and previous studies have identified that lncRNAs
regulate the chemoresistance of NSCLC cells (16,23,27).
lncRNA BLACAT1, originally called linc-UBC1, was first identified
in bladder cancer in 2013, and was considered as a negative
prognostic factor for metastasis to lymph nodes (28). Later, it was identified to be
abnormally expressed in colorectal cancer, NSCLC, cervical cancer,
etc. (24,29). Previous research has identified
that lncRNA BLACAT1 was increased in NSCLC tissues and cell lines,
and inhibition of BLACAT1 suppressed NSCLC cell proliferation and
migration (24). lncRNA BLACAT1 is
also involved in drug resistance of cancer (25); however, the underlying molecular
mechanism of BLACAT1 in regulating the chemoresistance of NSCLC
cells remains unclear. In the present study, it was identified that
lncRNA BLACAT1 was upregulated in DDP-resistant NSCLC cells, which
was consistent with results of previous research (24).
A further aim of the present study was to identify
the signaling pathway mediated by lncRNA BLACAT1 in regulating the
chemoresistance of NSCLC cells. It was identified that miR-17 was
downregulated in DDP-resistant NSCLC cells, which was consistent
with results of a previous study (20). Furthermore, miR-17 is involved in
modulating the cell viability and chemoresistance in a number of
types of cancer, such as pancreatic and breast cancer (30,31).
According to the bioinformatics software, there were binding sites
between BLACAT1 and miR-17. Therefore, lncRNA BLACAT1 may be
involved in chemoresistance of NSCLC cells via miR-17. Using RIP
and RNA pull-down assays, the interaction between BLACAT1 and
miR-17 was confirmed. Following transfection with pcDNA-BLACAT1 or
si-BLACAT1, miR-17 expression was markedly decreased in A549 cells
or increased in A549/DDP cells, respectively, which indicated that
BLACAT1 negatively regulated miR-17.
Research has focused on the function of autophagy in
chemoresistance of cancer cells, but the function of autophagy in
the response of cancer cells to chemoresistance is complex and
remains unclear (32,33). ATG7, as an autophagy-associated
protein, promoted autophagy and enhanced chemoresistance of cancer
cells (14,34). On the basis of the negative
regulation of miR-17 on ATG7 (21), we hypothesized that lncRNA BLACAT1
may promote autophagy thus to enhance chemo-resistance of NSCLC
cells via miR-17/ATG7. In the present study, it was identified that
the protein level of ATG7 was upregulated in DDP-resistant NSCLC
cells, and BLACAT1 positively regulated ATG7. Furthermore,
si-BLACAT1 suppressed autophagy-associated protein and
multidrug-resistance-associated protein expression in DDP-resistant
NSCLC cells, which indicated that inhibition of BLACAT1 could
suppress autophagy and chemoresistance of NSCLC cells.
In conclusion, lncRNA BLACAT1 was upregulated in
DDP-resistant NSCLC cells, and promoted autophagy and
chemoresistance of NSCLC cells. The interaction between BLACAT1 and
miR-17 was investigated, and identified the effect of
BLACAT1/miR-17/ATG7 pathway in chemoresistance of NSCLC cells,
which provided potential targets for the treatment of NSCLC.
Funding
The present study was supported by the Youth
Innovation Fund of The First Affiliated Hospital of Zhengzhou
University (grant no. YNQN 2017170) and the Science and Technology
Key Program of Henan Province (grant no. 162102310196).
Availability of data and materials
The datasets used and analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors’ contributions
FXH and LJM designed the study. FXH and HJC drafted
the paper. FXH, HJC, FXZ, ZYG and PFS conducted the experiments.
QP, YL, XD, YHH and CZ analyzed the data. FXH, HJC and LJM revised
the paper.
Ethics approval and consent to
participate
The study was approved by the Ethics Committee of
The First Affiliated Hospital of Zhengzhou University.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Acknowledgments
Not applicable.
References
|
1
|
Sapalidis K, Zarogoulidis P, Pavlidis E,
Laskou S, Katsaounis A, Koulouris C, Giannakidis D, Mantalovas S,
Huang H, Bai C, et al: Aerosol Immunotherapy with or without
Cisplatin for metastatic lung cancer non-small cell lung cancer
disease: In vivo Study. A more efficient combination J Cancer.
9:1973–1977. 2018.
|
|
2
|
Bach DH, Kim D, Bae SY, Kim WK, Hong JY,
Lee HJ, Rajasekaran N, Kwon S, Fan Y, Luu TT, et al: Targeting
Nicotinamide N-Methyltransferase and miR-449a in EGFR-TKI-Resistant
Non-Small-Cell Lung Cancer Cells. Mol Ther Nucleic Acids.
11:455–467. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Boya P, Reggiori F and Codogno P: Emerging
regulation and functions of autophagy. Nat Cell Biol. 15:713–720.
2013. View
Article : Google Scholar : PubMed/NCBI
|
|
4
|
Kim M, Jung JY, Choi S, Lee H, Morales LD,
Koh JT, Kim SH, Choi YD, Choi C, Slaga TJ, et al: GFRA1 promotes
cisplatin-induced chemoresistance in osteosarcoma by inducing
autophagy. Autophagy. 13:149–168. 2017. View Article : Google Scholar :
|
|
5
|
Mani J, Vallo S, Rakel S, Antonietti P,
Gessler F, Blaheta R, Bartsch G, Michaelis M, Cinatl J, Haferkamp
A, et al: Chemoresistance is associated with increased
cytoprotective autophagy and diminished apoptosis in bladder cancer
cells treated with the BH3 mimetic (-)-Gossypol (AT-101). BMC
Cancer. 15:2242015. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Kim TW, Lee SY, Kim M, Cheon C, Jang BH,
Shin YC and Ko SG: DSGOST regulates resistance via activation of
autophagy in gastric cancer. Cell Death Dis. 9:6492018. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Hong SB, Kim BW, Lee KE, Kim SW, Jeon H,
Kim J and Song HK: Insights into noncanonical E1 enzyme activation
from the structure of autophagic E1 Atg7 with Atg8. Nat Struct Mol
Biol. 18:1323–1330. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Noda NN, Satoo K, Fujioka Y, Kumeta H,
Ogura K, Nakatogawa H, Ohsumi Y and Inagaki F: Structural basis of
Atg8 activation by a homodimeric E1, Atg7. Mol Cell. 44:462–475.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Yamaguchi M, Satoo K, Suzuki H, Fujioka Y,
Ohsumi Y, Inagaki F and Noda NN: Atg7 Activates an
Autophagy-Essential Ubiquitin-like Protein Atg8 through Multi-Step
Recognition. J Mol Biol. 430:249–257. 2018. View Article : Google Scholar
|
|
10
|
Wang G, Chen EN, Liang C, Liang J, Gao LR,
Chuai M, Münsterberg A, Bao Y, Cao L and Yang X: Atg7-Mediated
Autophagy Is Involved in the Neural Crest Cell Generation in Chick
Embryo. Mol Neurobiol. 55:3523–3536. 2018. View Article : Google Scholar
|
|
11
|
Gonzalez Y, Aryal B, Chehab L and Rao VA:
Atg7- and Keap1-dependent autophagy protects breast cancer cell
lines against mitoquinone-induced oxidative stress. Oncotarget.
5:1526–1537. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Zhang Hu J, Mei L, Jiang Z, Yi Y, Liu Y,
Meng L, Zhou Y, Zeng L, Wu JH, et al: Interaction of E3 ubiquitin
ligase MARCH7 with long noncoding RNA MALAT1 and autophagy-related
protein ATG7 promotes autophagy and invasion in ovarian cancer.
Cell Physiol Biochem. 47:654–666. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Mandelbaum J, Rollins N, Shah P, Bowman D,
Lee JY, Tayber O, Bernard H, LeRoy P, Li P, Koenig E, et al:
Identification of a lung cancer cell line deficient in
atg7-dependent autophagy. Autophagy. Jun 19–2015.Epub ahead of
print. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Desai S, Liu Z, Yao J, Patel N, Chen J, Wu
Y, Ahn EE, Fodstad O and Tan M: Heat shock factor 1 (HSF1) controls
chemoresistance and autophagy through transcriptional regulation of
autophagy-related protein 7 (ATG7). J Biol Chem. 288:9165–9176.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Xu N, Zhang J, Shen C, Luo Y, Xia L, Xue F
and Xia Q: Cisplatin-induced downregulation of miR-199a-5p
increases drug resistance by activating autophagy in HCC cell.
Biochem Biophys Res Commun. 423:826–831. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Sun W, Zu Y, Fu X and Deng Y: Knockdown of
lncRNA-XIST enhances the chemosensitivity of NSCLC cells via
suppression of autophagy. Oncol Rep. 38:3347–3354. 2017.PubMed/NCBI
|
|
17
|
Fabian MR, Sonenberg N and Filipowicz W:
Regulation of mRNA translation and stability by microRNAs. Annu Rev
Biochem. 79:351–379. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Filipska M, Skrzypski M, Czetyrbok K,
Stokowy T, Stasiłojć G, Supernat A, Jassem J, Żaczek AJ and Bigda
J: MiR-192 and miR-662 enhance chemoresistance and invasiveness of
squamous cell lung carcinoma. Lung Cancer. 118:111–118. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Wei X, Shen X, Ren Y and Hu W: The roles
of micrornas in regulating chemotherapy resistance of non-small
cell lung cancer. Curr Pharm Des. 23:5983–5988. 2018. View Article : Google Scholar
|
|
20
|
Zhao J, Fu W, Liao H, Dai L, Jiang Z, Pan
Y, Huang H, Mo Y, Li S, Yang G, et al: The regulatory and
predictive functions of miR-17 and miR-92 families on cisplatin
resistance of non-small cell lung cancer. BMC Cancer. 15:7312015.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Comincini S, Allavena G, Palumbo S, Morini
M, Durando F, Angeletti F, Pirtoli L and Miracco C: microRNA-17
regulates the expression of ATG7 and modulates the autophagy
process, improving the sensitivity to temozolomide and low-dose
ionizing radiation treatments in human glioblastoma cells. Cancer
Biol Ther. 14:574–586. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Hu B, Zhang H, Wang Z, Zhang F, Wei H and
Li L: LncRNA CCAT1/miR-130a-3p axis increases cisplatin resistance
in non-small-cell lung cancer cell line by targeting SOX4. Cancer
Biol Ther. 18:974–983. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Wang P, Chen D, Ma H and Li Y: LncRNA
SNHG12 contributes to multidrug resistance through activating the
MAPK/Slug pathway by sponging miR-181a in non-small cell lung
cancer. Oncotarget. 8:84086–84101. 2017.PubMed/NCBI
|
|
24
|
Ye JR, Liu L and Zheng F: Long noncoding
RNA bladder cancer associated transcript 1 promotes the
proliferation, migration, and invasion of nonsmall cell lung cancer
through sponging miR-144. DNA Cell Biol. 36:845–852. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Wu X, Zheng Y, Han B and Dong X: Long
noncoding RNA BLACAT1 modulates ABCB1 to promote oxaliplatin
resistance of gastric cancer via sponging miR-361. Biomed
Pharmacother. 99:832–838. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) Method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
27
|
Zhang W, Cai X, Yu J, Lu X, Qian Q and
Qian W: Exosome-mediated transfer of lncRNA RP11 838N2.4 promotes
erlotinib resistance in non-small cell lung cancer. Int J Oncol.
53:527–538. 2018.PubMed/NCBI
|
|
28
|
He W, Cai Q, Sun F, Zhong G, Wang P, Liu
H, Luo J, Yu H, Huang J and Lin T: linc-UBC1 physically associates
with polycomb repressive complex 2 (PRC2) and acts as a negative
prognostic factor for lymph node metastasis and survival in bladder
cancer. Biochim Biophys Acta. 1832.1528–1537. 2013.
|
|
29
|
Zhang Su J, Han E, Yin L, Liu D, He Z,
Zhang X, Lin Y, Lin F, Mao QP, et al: Long noncoding RNA BLACAT1
indicates a poor prognosis of colorectal cancer and affects cell
proliferation by epigenetically silencing of p15. Cell Death Dis.
8:e26652017. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Wang Gu J, Zhang D, Zhu J, Li Y, Chen Y,
Shi H, Wang M, Shen X, Deng BX, et al: GFRα2 prompts cell growth
and chemoresistance through down-regulating tumor suppressor gene
PTEN via Mir-17-5p in pancreatic cancer. Cancer Lett. 380:434–441.
2016. View Article : Google Scholar
|
|
31
|
Ao X, Nie P, Wu B, Xu W, Zhang T, Wang S,
Chang H and Zou Z: Decreased expression of microRNA-17 and
microRNA-20b promotes breast cancer resistance to taxol therapy by
upregu-lation of NCOA3. Cell Death Dis. 7:e24632016. View Article : Google Scholar
|
|
32
|
Notte A, Leclere L and Michiels C:
Autophagy as a mediator of chemotherapy-induced cell death in
cancer. Biochem Pharmacol. 82:427–434. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Wu WK, Coffelt SB, Cho CH, Wang XJ, Lee
CW, Chan FK, Yu J and Sung JJ: The autophagic paradox in cancer
therapy. Oncogene. 31:939–953. 2012. View Article : Google Scholar
|
|
34
|
Piya S, Kornblau SM, Ruvolo VR, Mu H,
Ruvolo PP, McQueen T, Davis RE, Hail N Jr, Kantarjian H, Andreeff
M, et al: Atg7 suppression enhances chemotherapeutic agent
sensitivity and overcomes stroma-mediated chemoresistance in acute
myeloid leukemia. Blood. 128:1260–1269. 2016. View Article : Google Scholar : PubMed/NCBI
|