Introduction
Esophageal carcinoma (ESCA) is one of the most
common types of cancer in the digestive system which primarily
includes esophageal squamous cell carcinoma (ESCC) and esophageal
adenocarcinoma (1). According to
recent statistics, estimated new cases of and mortality from ESCA
account for a large proportion of all cancer in the USA (2). ESCA also ranks in the top ten of
cancer incidence and mortality rate worldwide, which is associated
with genetic and environmental factors (3). In addition, patients with ESCA have a
poor prognosis and high mortality rate that may be associated with
glucose-regulated protein 78 (4,5).
At present, several methods are practiced in
clinical treatment for ESCA (6).
Surgical resection is commonly used to remove cancerous tissue and
leads to an improved survival rate. Neoadjuvant chemotherapy
(without radiation therapy) provides an improved survival benefit
compared with surgery alone (7),
but the effects on local control are unclear. Tri-modality
treatment (concomitant chemotherapy and radiation therapy followed
by surgery) also has a marked effect compared with surgery alone
(5). Additionally, several
chemotherapy target agents for ESCA have been reported, including
fluoropyrimidines [5-fluorouracil (5-FU) and capecitabine],
platinum agents (cisplatin and oxaliplatin), taxanes (paclitaxel
and docetaxel), irinotecan, mitomycin-C, anthracyclines,
methotrexate, vinorelbine and gemcitabine. The combination of two
or more drugs has a more marked reaction, as it has been reported
to increase the efficacy of chemoradiotherapy in patients with ESCA
(8-10). However, intolerance of
fluoropyrimidine-based, platinum-based and taxane-based
chemotherapy has also been identified in patients, and immune
checkpoint blockade therapy remains under investigation (11). Therefore, further research is
required for more effective therapeutic methods.
Gene-associated therapy for cancers has become a
research focus in recent years (12-16).
With regard to ESCA, it was identified that the aberrant
transcription and translation of genes affect the synthesis of
associated proteins, which has an effect on the corresponding
physiological processes, and consequently influences the diagnosis
and prognosis of ESCA (17-19).
It was also identified that the chemotherapeutic mechanism of ESCA
was associated with upregulation and downregulation of genes or DNA
repair genes (20,21). In addition, gene levels and certain
signaling pathways were identified to be associated with
chemotherapy resistance (22,23).
For example, it was reported that the expression of myeloid cell
leukemia 1 in ESCA cancerous tissue with cisplatin resistance
increased in comparison with that in normal tissue (24). Let-7 was also identified to serve a
potentially positive function in patients with 5-FU resistance
(25), which indicates that the
drugs targeted in these pathways or genes are potential candidates
for improved treatment. Generally, differentially expressed genes
(DEGs) and signaling pathways may be the fundamental reason for the
development of ESCA. Thus, research into the biological
significance of target genes and their corresponding signaling
pathways is required for gene-associated chemotherapy.
However, specific drugs mapped to the target genes
and their associated pathways remain unknown. Thus, the aim of the
present study was to predict novel drug candidates of ESCA using
bioinformatics analysis, which was principally based on the
Connectivity Map (CMAP), a database that contains 1,309 Food and
Drug Administration-approved small molecules and 7,000 gene
expression profiles, and has been widely used to reveal the
mechanism of drugs (26).
Furthermore, molecular virtual docking was used in the search for
matching predicted potential drugs and screened proteins. To a
certain extent, the present study may provide a new point of
reference for chemotherapy for ESCA, with regard to its molecular
biology and the methodology.
Materials and methods
Acquisition of DEGs in ESCA
To acquire the information of gene expression levels
in ESCA, the gene expression data of ESCA were downloaded from The
Cancer Genome Atlas (TCGA) and Genotype-Tissue Expression (GTEx)
(27), and the edition of TCGA
dataset on ESCA was updated on October 13, 2017. The gene
expression data of ESCA were normalized into the transcripts per
million reads format. The differential expression analysis was
further performed on the basis of the normalized data and analysis
of variance (ANOVA), which was used for tumor vs. paired normal
samples. For the differential analysis methods of ANOVA, genes with
|log2 fold-change (FC)|≥2 and q-value <0.05 were
considered to be DEGs.
Analysis of DEGs associated with
ESCA
Functional analysis of differentially
expressed genes
WEB-based GEne SeT AnaLysis Toolkit (Webgestalt;
www.webgestalt.org) was used to explain the
function of extracted DEGs with an enrichment method of
over-representation (enrichment) analysis. Gene Ontology (GO)
enrichment analysis was performed to classify the genes into their
Biological Process (BP), Cellular Component (CC) and Molecular
Function (MF) categories. The Kyoto Encyclopedia of Genes and
Genomes (KEGG) (www.kegg.jp) and Protein Annotation
Through Evolutionary Relationships (PANTHER) (www.pantherdb.org) analyses were performed to
determine the pathways in which the DEGs were significantly
involved (28-30).
Excavation of hub genes of ESCA
For further understanding of genes that may have a
significant effect on ESCA, the protein-protein interaction (PPI)
network was then drafted via Search Tool for the Retrieval of
Interacting Genes (STRING; string-db.org). The interaction between two nodes,
which represent two genes, was confirmed using Textmining,
Experiments, Databases, Co-expression, Neighborhood, Gene Fusion
and Co-occurrence. Degrees of edges were calculated using Cytoscape
(29,31-34).
Finally, the genes with a degree of ≥30 were selected to be hub
genes of ESCA, which were considered to be potential bioactive
factors for ESCA and were further involved in the subsequent
research.
Prediction of ESCA-associated drugs and
extraction of relevant target genes
Drug prediction of ESCA based on
CMAP
CMAP (portals. broadinstitute.org/cmap) (26,28)
was selected to predict the potential drug molecules for ESCA.
Following ID conversion via Affymetrix (www.affymetrix.com/analysis/index.affx), the
upregulated and downregulated DEGs were uploaded to CMAP for online
analysis. Detailed results from CMAP were downloaded, and the first
screening drugs were further acquired following filtering for a
score of <−0.85.
Extraction of target genes for the
first screening drugs
To further screen out more significant drug
molecules and prepare for the following investigation of potential
drug-associated mechanisms in ESCA, the Search Tool for
Interactions of Chemicals (STITCH, stitch.embl.de), a
protein-compound interactions database that contains 390,000
compounds and 3.6 million proteins, was used in which the Canonical
Simplified Molecular-Input Line-Entry System (SMILES) structure of
drugs downloaded from PubChem (pubchem.ncbi.nlm.nih. gov) were
uploaded in order to search for target genes. In this way, the
second screening drugs were extracted when the drugs without
STITCH-predicted results were phased out, and their corresponding
target genes were obtained. Additionally, intersection between
predicted target genes and the extracted DEGs of ESCA was
calculated using FunRich (35) for
a preliminary understanding of their associations.
Interaction analysis of drug
candidates-target genes-ESCA- associated hub genes
To investigate the associations between the
predicted drugs and hub genes of ESCA, the target genes of drugs
and hub genes of ESCA were analyzed. First, another PPI network was
constructed to analyze the interactions between the two sources of
genes aforementioned. Connected nodes, confirmed by experiments and
databases, were identified and were considered to be associated
genes. The unconnected nodes were not included in the following
investigation and their corresponding drugs were eliminated. Up to
this point, all the extant drugs were considered to be drug
candidates of ESCA in the present study. Furthermore, genes
confirmed by PPI were then used for constructing the network of
drug candidates-target genes-ESCA-associated hub genes. Further
analysis of the selected genes was performed using the same methods
as for the aforementioned functional analysis of DEGs of ESCA.
Molecular docking between predicted
drug candidates and hub genes
To further investigate the underlying associations
between drugs and hub genes in the drug candidates-target
genes-ESCA-associated hub genes network, molecular docking was
performed using systemsDock (systemsdock.unit.oist.
jp/iddp/home/index), which is a web server for network
pharmacology-based prediction and analysis (36,37).
Selected by the second PPI analysis, protein names of the
ESCA-associated hub genes in the network were added and specified,
then the SMILES structures of drug candidates involved in the
network were also uploaded. A protein and a drug were tested each
time. The binding affinity between each hub gene and drug groups
was predicted by docking scores.
Results
DEGs of ESCA and functional analysis
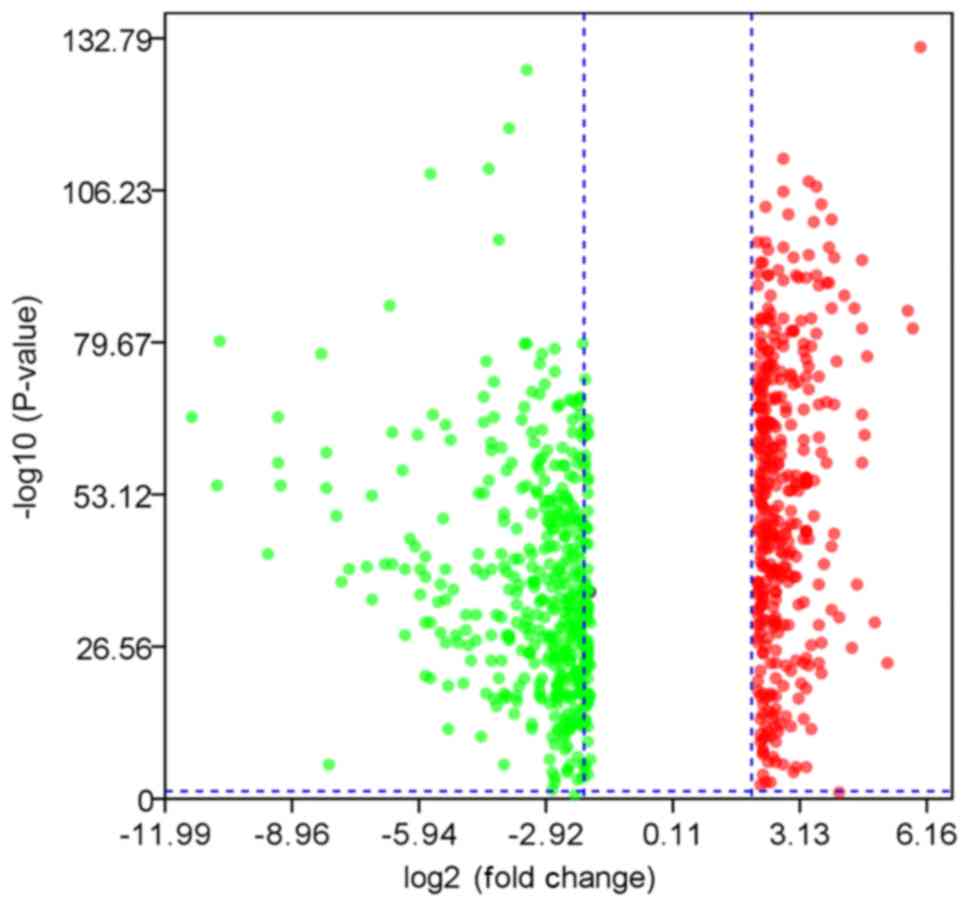
A total of 827 DEGs were downloaded from TCGA and
GTEx databases with the criteria of |log2FC|≥2 and q-value <0.05
in ANOVA, which includes 375 upregulated genes and 452
downregulated genes (Fig. 1). The
following KEGG analysis showed that DEGs were significantly
involved in 10 pathways (P<0.005), particularly in protein
digestion and absorption, extracellular matrix-receptor interaction
and complement, and coagulation cascades (Table I). A PANTHER analysis indicated
that DEGs were significantly involved in five pathways (P<0.05),
among which the plasminogen-activating cascade, blood coagulation
and integrin signaling pathway were particularly significant
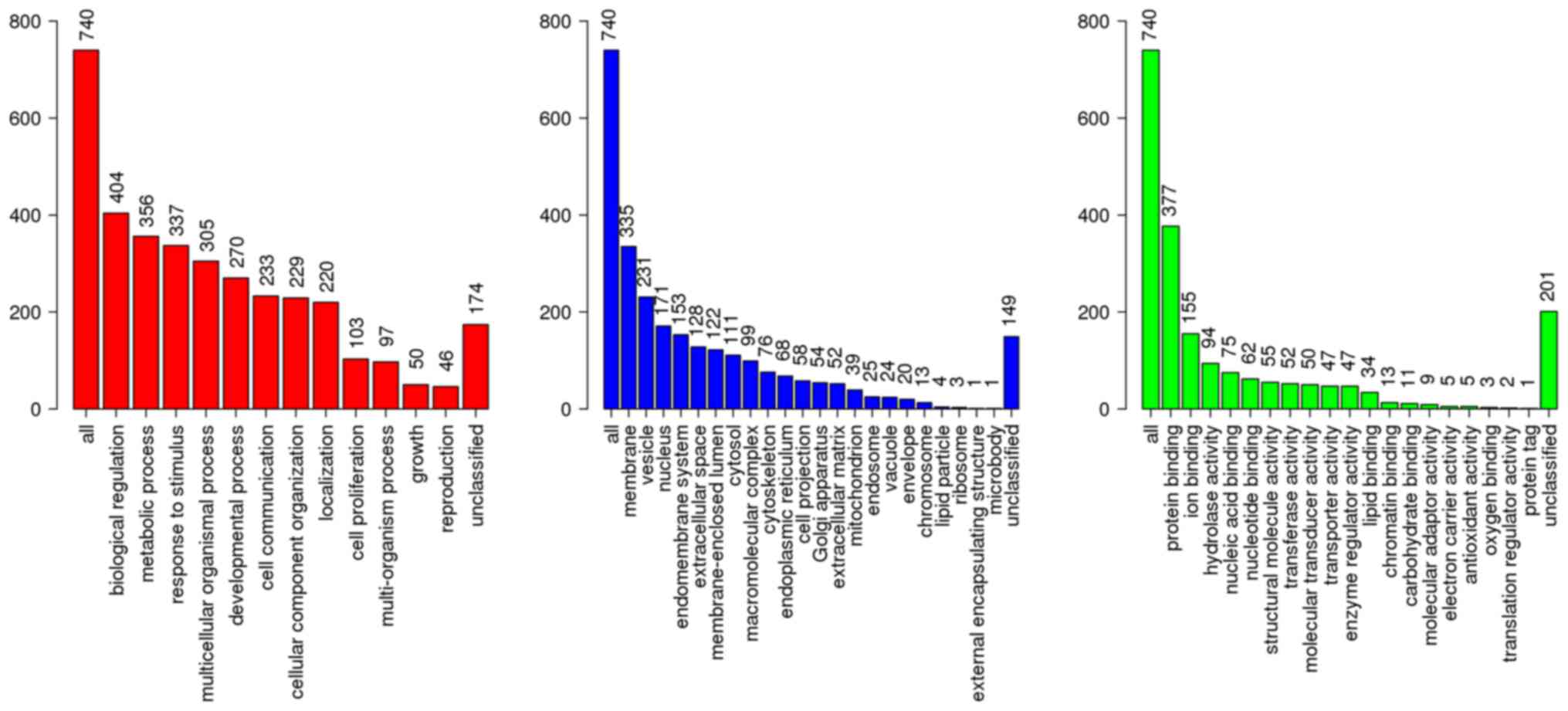
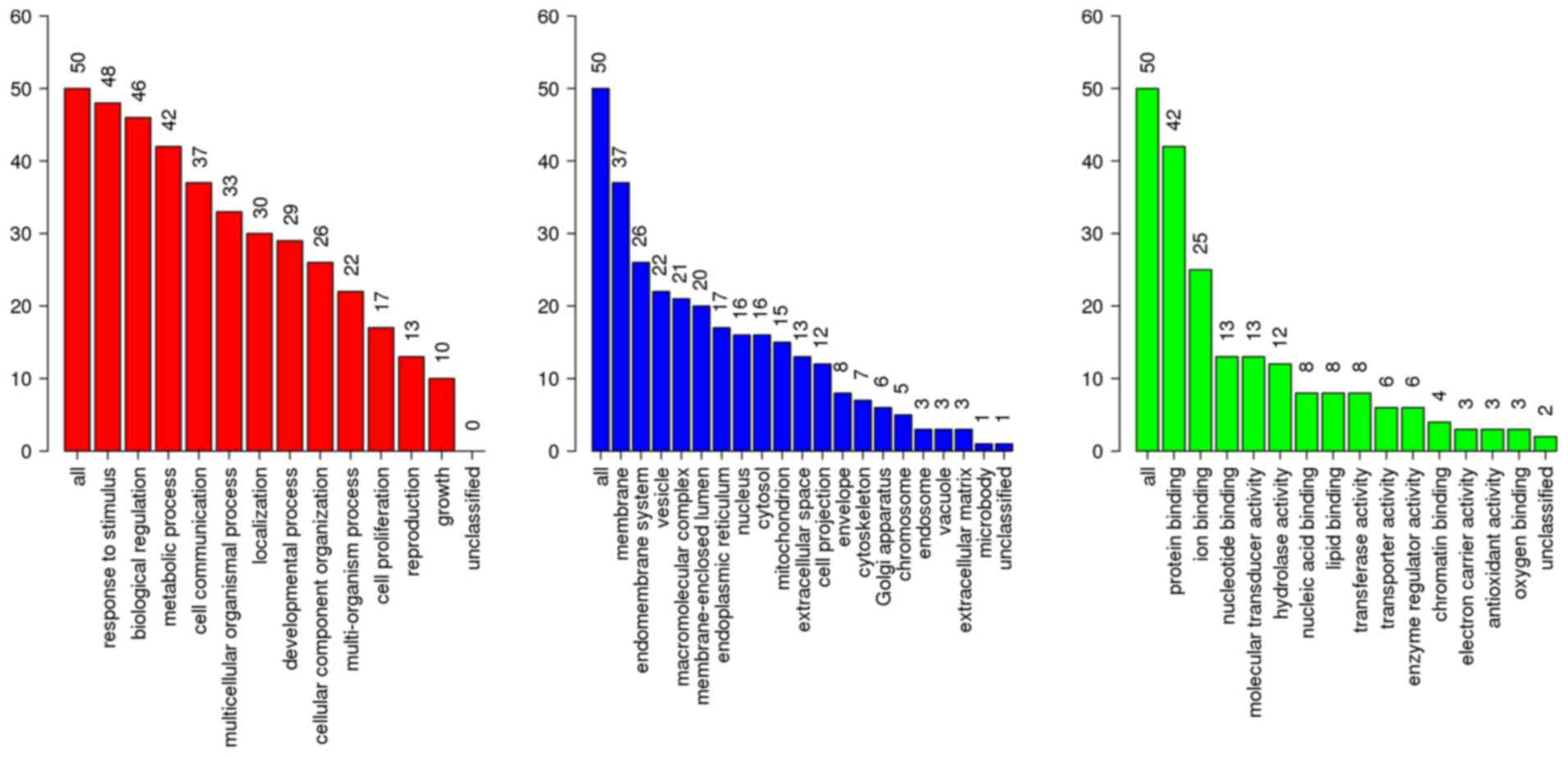
(P<0.01; Table II). Classified
by GO enrichment analysis, the DEGs were primarily involved in the
‘biological regulation’, ‘metabolic process’ and ‘response to
stimulus’ BP categories, and the ‘membrane’ and ‘vesicle’ CC
categories. They also were involved in the ‘nucleus’ CC category,
and in the ‘protein binding’, ‘ion binding’ and ‘hydrolase
activity’ MF categories (Fig. 2).
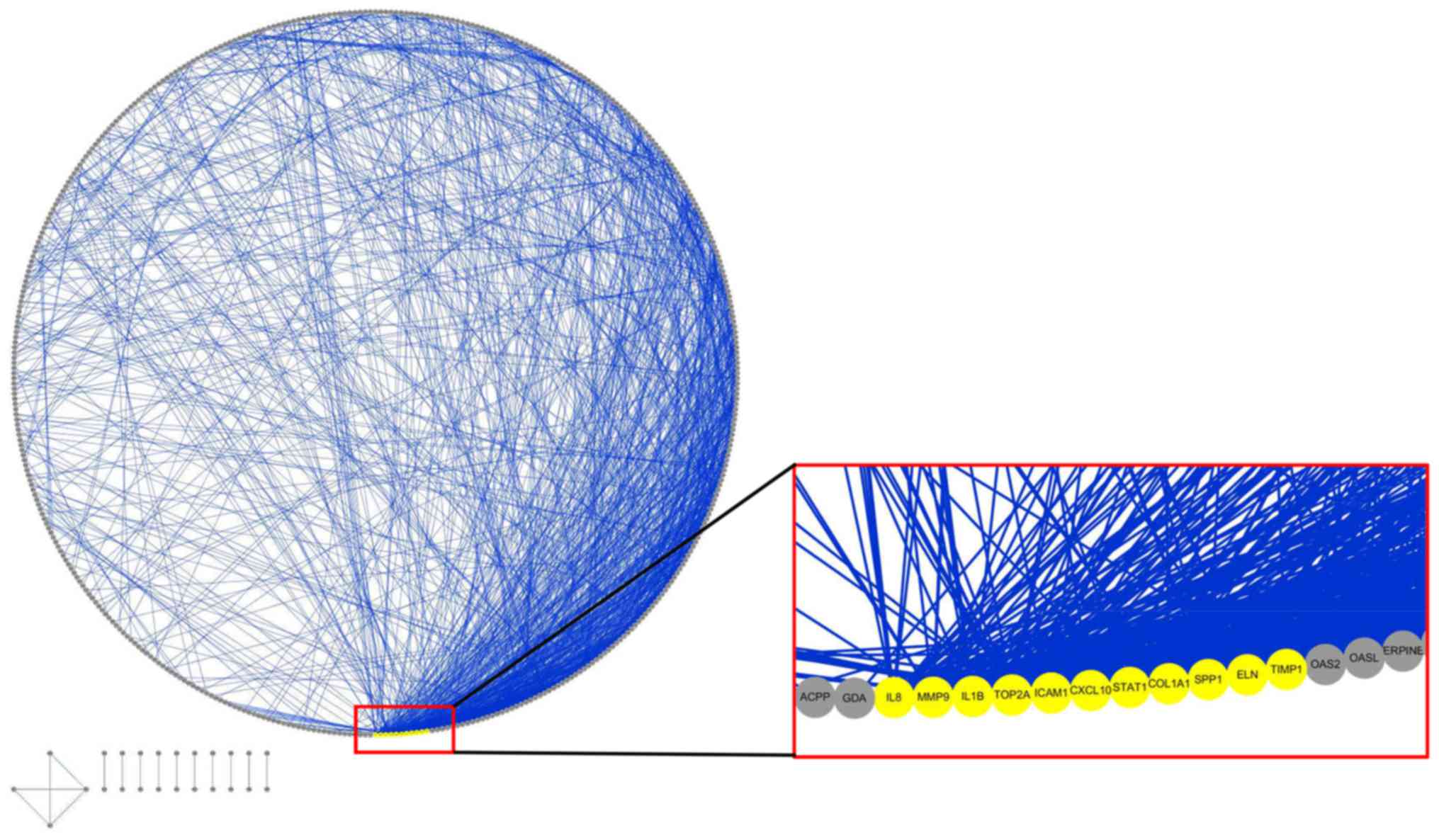
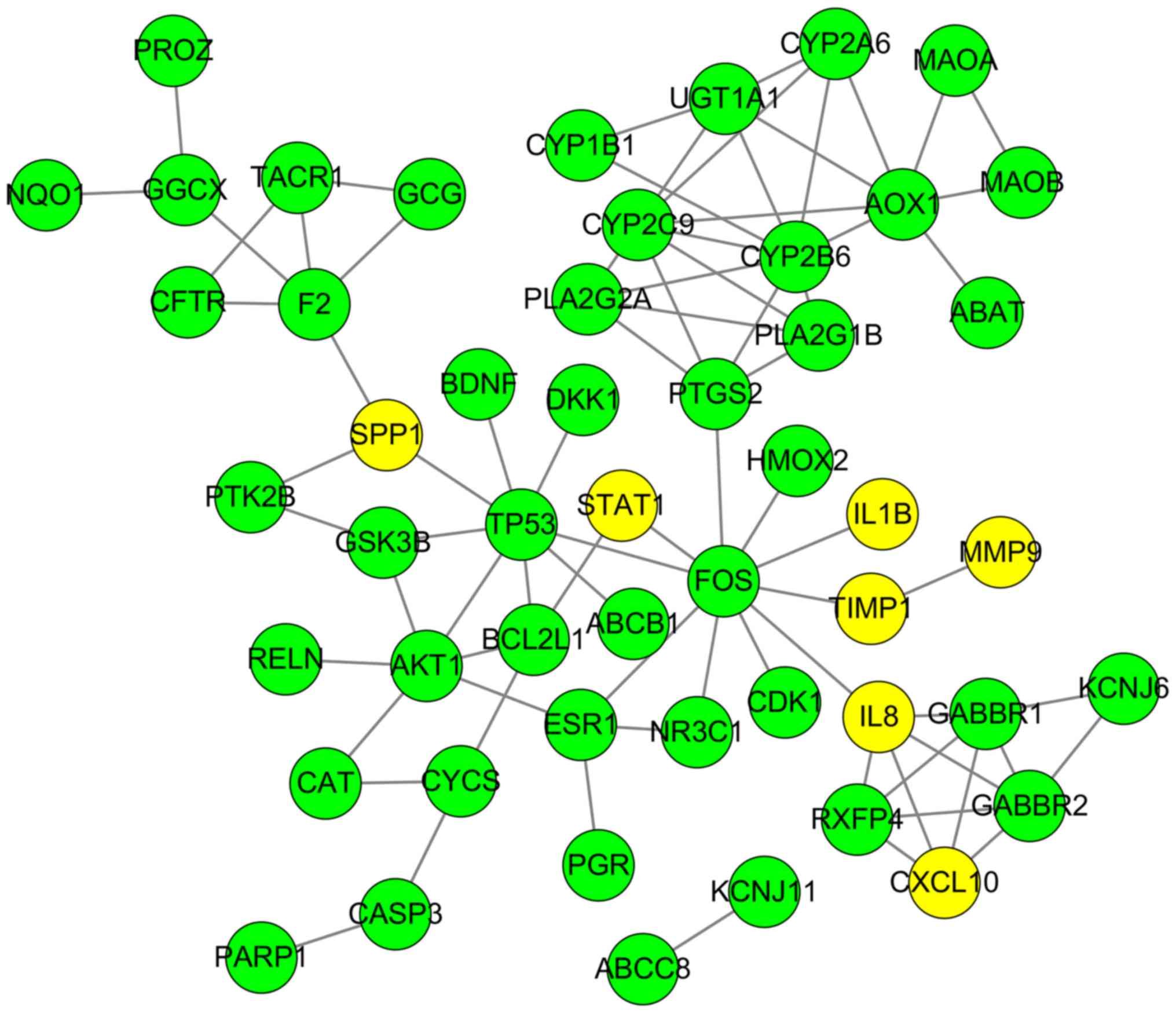
Furthermore, 11 genes, including elastin (ELN), tissue
inhibitor of matrix metalloproteinase 1 (TIMP1),
intercellular adhesion molecule 1 (ICAM1), topoisomerase 2A
(TOP2A), interleukin 1B (IL1B), collagen type I α1
(COL1A1), IL8, matrix metalloproteinase 9
(MMP9), secreted phosphoprotein 1 (SPP1), signal
transducer and activator of transcription 1 (STAT1) and CXC
chemokine ligand 10 (CXCL10), were finally determined to be
hub genes following calculation of the edges based on PPI network
analysis (all degrees ≥30) (Fig.
3).
 | Table ITwo-time Kyoto Encyclopedia of Genes
and Genomes analysis. |
Table I
Two-time Kyoto Encyclopedia of Genes
and Genomes analysis.
| Group | Gene set | Description | P-value | FDR | Overlapping
genes |
|---|
| A | hsa04974 | Protein digestion
and absorption |
2.45×10−7 |
7.41×10−5 | 10008, 10136, 1277,
1278, 1281, 1282, 1284, 1289, 1300, 2006, 477, 5644, 5645, 643834,
643847 |
| hsa04512 | ECM-receptor
interaction |
1.61×10−5 |
2.44×10−3 | 1277, 1278, 1282,
1284, 3655, 3679, 3685, 3691, 375790, 3918, 6696, 7148 |
| hsa04610 | Complement and
coagulation cascades |
5.86×10−5 |
5.92×10−3 | 1675, 2149, 2151,
5054, 5055, 5265, 5328, 5329, 713, 714, 730 |
| hsa05146 | Amebiasis |
1.20×10−4 |
9.06×10−3 | 1277, 1278, 1281,
1282, 1284, 2919, 3553, 3576, 3918, 5275, 6317, 6318 |
| hsa00590 | Arachidonic acid
metabolism |
2.01×10−4 |
1.22×10−2 | 100137049, 1571,
239, 247, 2878, 50487, 5320, 5730, 874 |
| hsa00982 | Drug metabolism:
Cytochrome P450 |
5.09×10−4 |
2.04×10−2 | 125, 131, 1571,
218, 222, 2327, 2946, 316, 54577 |
| hsa04062 | Chemokine signaling
pathway |
5.22×10−4 |
2.04×10−2 | 10563, 2919, 2921,
3576, 3579, 3627, 4067, 4283, 6358, 6362, 6363, 6364, 6366, 6373,
6772, 9560 |
| hsa04933 | AGE-RAGE signaling
pathway in diabetic complications |
5.38×10−4 |
2.04×10−2 | 1277, 1278, 1281,
1282, 1284, 3383, 3553, 3576, 5054, 5292, 6772 |
| hsa00980 | Metabolism of
xenobiotics by cytochrome P450 |
7.70×10−4 |
2.59×10−2 | 125, 131, 1571,
218, 222, 2946, 29785, 54577, 874 |
| hsa04060 | Cytokine-cytokine
receptor interaction |
1.42×10−3 |
4.30×10−2 | 10563, 2919, 2921,
3553, 3576, 3579, 3624, 3627, 4233, 4283, 51330, 53833, 6358,6362,
6363, 6364, 6366, 6373, 9560 |
| B | hsa00982 | Drug metabolism:
Cytochrome P450 |
3.84×10−7 |
1.16×10−4 | 1548, 1555, 1559,
316, 4128, 4129, 54658 |
| hsa05210 | Colorectal
cancer |
3.41×10−6 |
4.02×10−4 | 207, 2353, 2932,
54205, 7157, 836 |
| hsa04915 | Estrogen signaling
pathway |
4.40×10−6 |
4.02×10−4 | 207, 2099, 2353,
2550, 3763, 4318, 9568 |
| hsa05161 | Hepatitis B |
5.40×10−6 |
4.02×10−4 | 207, 2185, 2353,
4318, 54205, 6772, 7157, 836 |
| hsa00380 | Tryptophan
metabolism |
6.63×10−6 |
4.02×10−4 | 1545, 316, 4128,
4129, 847 |
| hsa04668 | TNF signaling
pathway |
8.30×10−6 |
4.19×10−4 | 207, 2353, 3553,
3627, 4318, 5743, 836 |
| hsa05014 | Amyotrophic lateral
sclerosis |
2.24×10−5 |
9.68×10−4 | 54205, 598, 7157,
836, 847 |
| hsa04210 | Apoptosis |
4.01×10−5 |
1.52×10−3 | 142, 207, 2353,
54205, 598, 7157, 836 |
| hsa00590 | Arachidonic acid
metabolism |
5.82×10−5 |
1.96×10−3 | 1555, 1559, 5319,
5320, 5743 |
| hsa00830 | Retinol
metabolism |
7.32×10−5 |
2.07×10−3 | 1548, 1555, 1559,
316, 54658 |
| Table IITwo-time Protein Annotation Through
Evolutionary Relationships analysis. |
Table II
Two-time Protein Annotation Through
Evolutionary Relationships analysis.
| Group | Gene set | Description | P-value | FDR | Overlapping
genes |
|---|
| A | P00050 |
Plasminogen-activating cascade |
2.39×10−7 |
2.70×10−5 | 5328, 4312, 5055,
5054, 4314, 4318, 5329 |
| P00011 | Blood
coagulation |
1.64×10−3 |
9.25×10−2 | 5328, 5265, 5055,
5054, 2149, 5329 |
| P00034 | Integrin signaling
pathway |
3.46×10−3 |
1.30×10−1 | 10095, 1277, 1281,
1282, 3685, 1278, 1284, 3691, 1289, 3655, 1300, 3679, 3918 |
| P06959 | CCKR signaling
map |
1.55×10−2 |
4.39×10−1 | 5328, 7031, 5055,
5054, 3685, 4067, 4314, 4316, 2919, 3576, 4953, 4318 |
| P00031 | Inflammation
mediated by chemokine and cytokine signaling pathway |
1.94×10−2 |
4.39×10−1 | 6366, 10095, 3627,
3576, 239, 3579, 4629, 6362, 72, 6364, 3710, 4776, 9560 |
| B | P06959 | CCKR signaling
map |
1.00×10−5 |
1.14×10−3 | 2353, 2641, 4318,
6869, 207, 5743, 836, 2932, 598, 2185 |
| P00006 | Apoptosis signaling
pathway |
1.05×10−3 |
5.93×10−2 | 2353, 7157, 207,
836, 54205, 598 |
| P00029 | Huntington
disease |
1.27×10−2 |
3.88×10−1 | 2353, 7157, 627,
207, 836 |
| P00036 | Interleukin
signaling pathway |
1.62×10−2 |
3.88×10−1 | 2353, 207, 6772,
2932 |
| P04372 | 5-Hydroxytryptamine
degradation |
1.71×10−2 |
3.88×10−1 | 4128, 4129 |
| P04397 | p53 pathway by
glucose deprivation |
2.30×10−2 |
4.34×10−1 | 7157, 207 |
| P00001 | Adrenaline and
noradrenaline biosynthesis |
3.69×10−2 |
4.78×10−1 | 4128, 4129 |
| P04393 | Ras pathway |
4.36×10−2 |
4.78×10−1 | 207, 6772,
2932 |
| P00020 | Fas signaling
pathway |
4.76×10−2 |
4.78×10−1 | 142, 836 |
ESCA-associated drugs and relevant target
genes
Upregulated and downregulated genes from the TCGA
and GTEx databases were uploaded to CMAP, then scores of the
predicted ESCA-associated drugs were obtained from the detailed
results. Drugs that had a score of <−0.85 were extracted as the
first screening drugs for further research, including verteporfin,
apigenin, cefixime, Prestwick-685, menadione, baclofen, dequalinium
chloride, butyl hydroxybenzoate, naloxone, paroxetine,
acetazolamide, gliclazide, valproic acid and metacycline (Table III). Furthermore, Prestwick-685
was categorized as clofazimine by PubChem, whereas dequalinium
chloride and butyl hydroxybenzoate were also named dequalinium and
butylparaben, respectively. On the basis of STITCH, 70 target genes
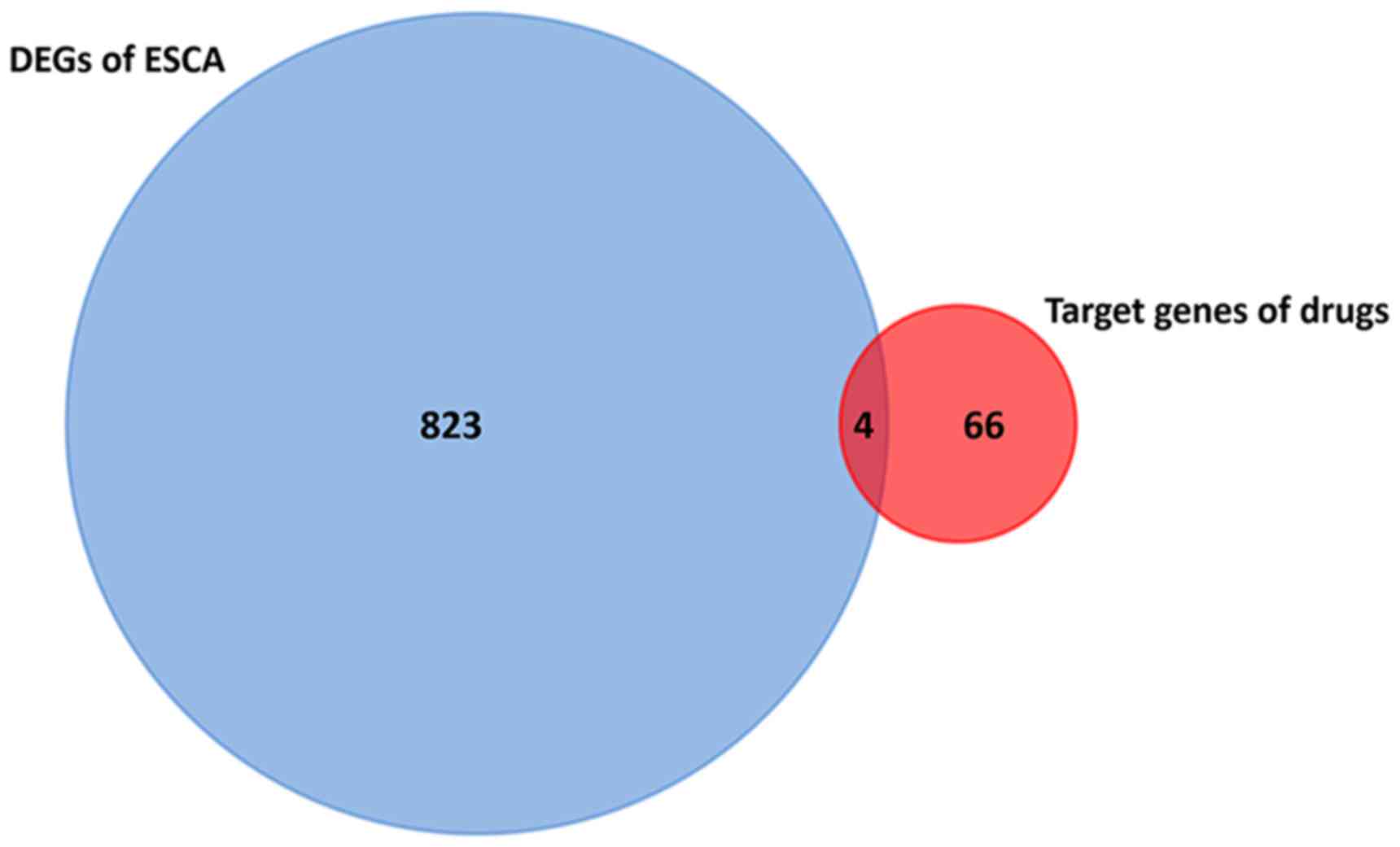
of 9 second screening drugs were acquired (Fig. 4). In addition, as Fig. 5 presents, only four gene
intersections were identified in DEGs of ESCA and target genes of
the 9 second screening drugs, which was a preliminary indication
that no significant difference in expression of the drug-associated
target genes in ESCA was identified.
| Table IIIFirst screening drugs extracted from
CMAP (scores <-0.85). |
Table III
First screening drugs extracted from
CMAP (scores <-0.85).
| No. | CMAP name | Score | STITCH | Canonical
SMILES |
|---|
| 6817 | Verteporfin | −1 | Verteporfin |
CC1=C(C2=CC3=NC(=CC4=C(C(=C(N4)C=C5C6(C(C(=CC=C6C(=N5)C=C1N2)C(=O)OC)C(=O)OC)C)C)
CCC(=O)OC)C(=C3C)CCC(=O)O)C=C |
| 4578 | Apigenin | −0.964 | Apigenin |
C1=CC(=CC=C1C2=CC(=O)C3=C(C=C(C=C3O2)OC4C(C(C(C(O4)CO)O)O)O)O)O |
| 4567 | Cefixime | −0.912 | Cefixime |
C=CC1=C(N2[C@@H]([C@@H](C2=O)NC(=O)/C(=N\OCC(=O)O)/C3=CSC(=N3)N)SC1)C(=O)O |
| 7382 | Prestwick-685 | −0.901 | Clofazimine |
CC(C)N=C1C=C2C(=NC3=CC=CC=C3N2C4=CC=C(C=C4)Cl)C=C1NC5=CC=C(C=C5)Cl |
| 4662 | Menadione | −0.896 | Menadione |
CC1=CC(=O)C2=CC=CC=C2C1=O |
| 6313 | Baclofen | −0.889 | Baclofen |
C1=CC(=CC=C1C(CC(=O)O)CN)Cl |
| 6296 | Dequalinium
chloride | −0.883 | Dequalinium |
CC1=[N+](C2=CC=CC=C2C(=C1)N)CCCCCCCCCC[N+]3=C(C=C(C4=CC=CC=C43)N)C.[Cl−].[Cl−] |
| 4647 | Butyl
hydroxybenzoate | −0.882 | Butylparaben |
CCCCOC(=O)C1=CC=C(C=C1)O |
| 4645 | Naloxone | −0.876 | Naloxone |
C=CCN1CCC23C4C(=O)CCC2(C1CC5=C3C(=C(C=C5)O)O4)O |
| 3556 | Verteporfin | −0.868 | Verteporfin |
CC1=C(C2=CC3=NC(=CC4=C(C(=C(N4)C=C5C6(C(C(=CC=C6C(=N5)C=C1N2)C(=O)OC)C(=O)OC)C)C)
CCC(=O)OC)C(=C3C)CCC(=O)O)C=C |
| 4556 | Paroxetine | −0.865 | Paroxetine |
C1CNCC(C1C2=CC=C(C=C2)F)COC3=CC4=C(C=C3)OCO4 |
| 1850 | Acetazolamide | −0.863 | Acetazolamide |
CC(=O)NC1=NN=C(S1)S(=O)(=O)N |
| 5089 | Gliclazide | −0.86 | Gliclazide |
CC1=CC=C(C=C1)S(=O)(=O)NC(=O)NN2CC3CCCC3C2 |
| 1155 | Valproic acid | −0.859 | Valproate |
CCCC(CCC)C(=O)O |
| 2901 | Metacycline | −0.855 | Methacycline |
CN(C)C1C2C(C3C(=C)C4=C(C(=CC=C4)O)C(=C3C(=O)C2(C(=O)C(=C1O)C(=O)N)O)O)O.Cl |
Network of drug candidates-target
genes-ESCA-associated hub genes
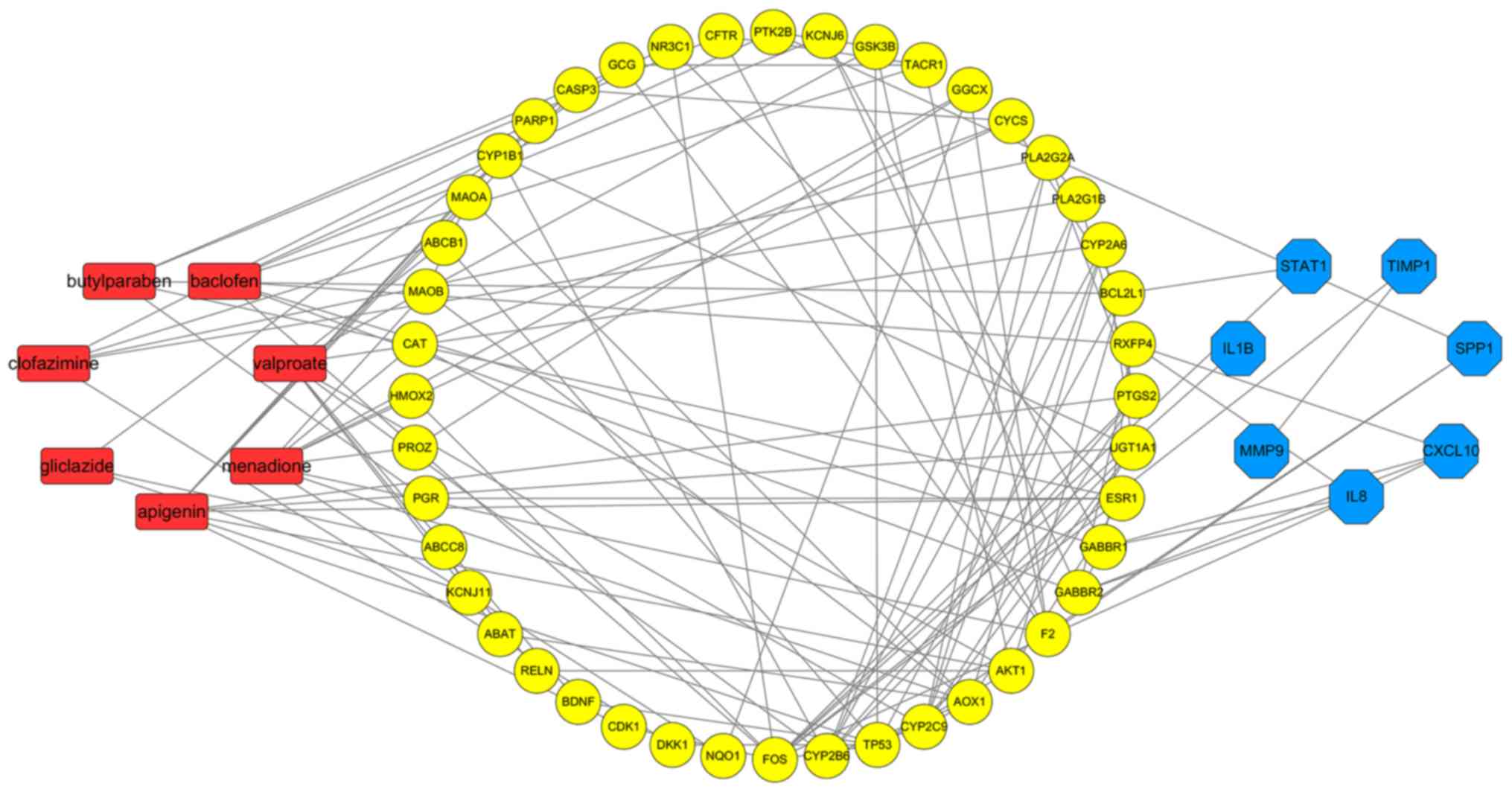
Gene association analysis
A total of 70 target genes of drugs and 11 hub genes
were united for further analysis. Interactions of the relevant
genes were identified in the second PPI network. According to
Fig. 6 (with the green circles
representing target genes of predicted drugs and yellow circles
representing the hub genes of ESCA), 51 genes were considered to be
connected and met the criteria of having been confirmed by other
experiments and databases. Acetazolamide and dequalinium were
eliminated because their target genes were considered meaningless
through PPI analysis, thus the other 7 drugs which were relevant to
the aforementioned 51 genes were consequently considered as drug
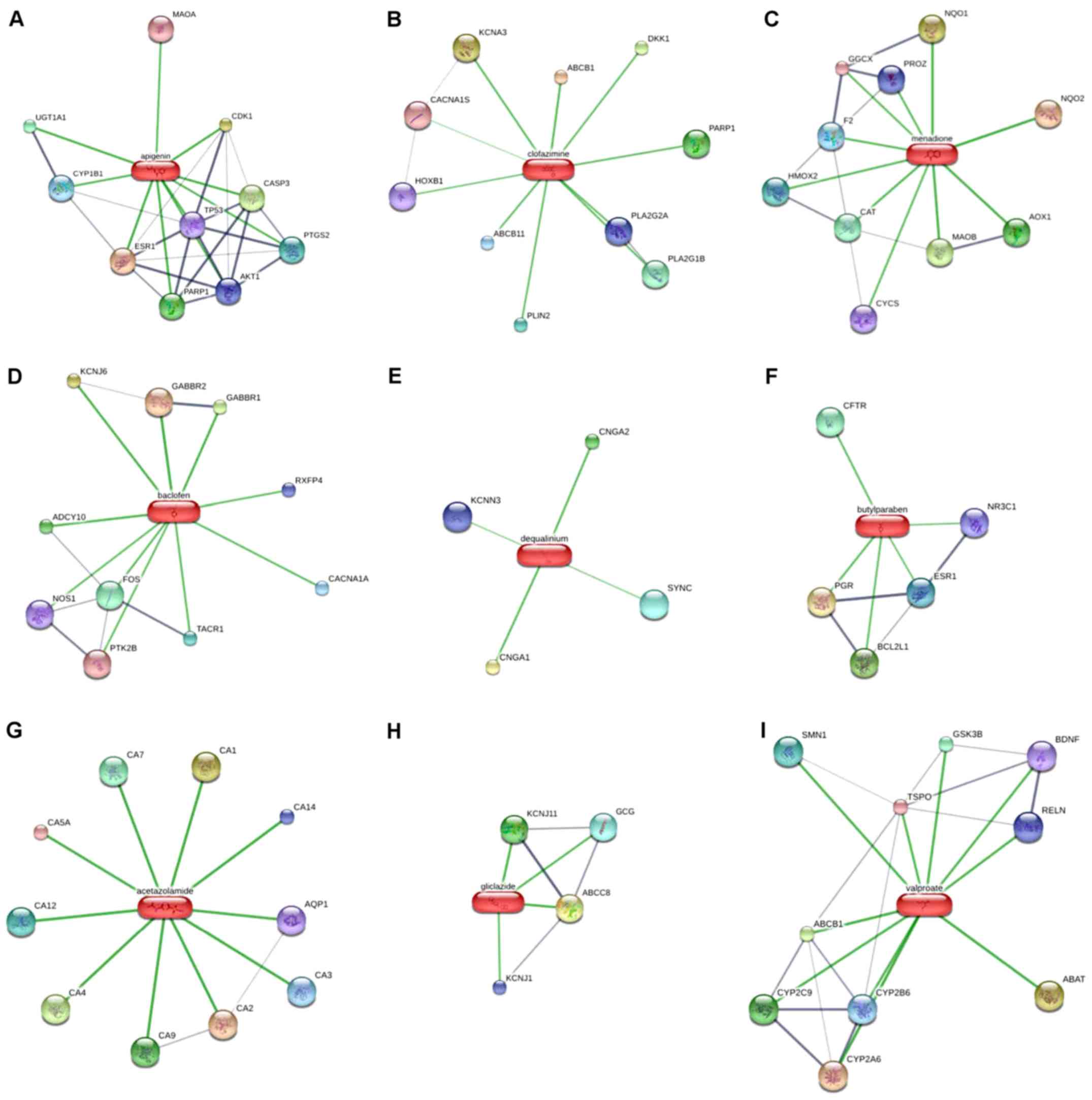
candidates of ESCA. As indicated in Fig. 7, associations among the 7 predicted
drug candidates-target genes-ESCA-associated hub genes were
revealed via network construction, in which the red oblongs
represent drug candidates, yellow circles represent 44/51 selected
genes (which were also target genes of drug candidates) and blue
polygons represent the other 7 genes also belonging to
ESCA-associated hub genes.
Functional analysis of genes
The 51 selected genes were identified to participate
principally in 10 pathways, including drug metabolism (cytochrome
P450), colorectal cancer and the estrogen signaling pathway
(Table I), on the basis of KEGG
analysis (P<0.001), as well as 9 pathways on the basis of
PANTHER analysis (P<0.05), which, significantly, included the
cholecystokinin receptor (CCKR) signaling map and apoptosis
signaling pathway (P<0.01; Table
II). GO annotation analysis indicated that the results of BP,
CC, and MF classification were almost the same as the results of
the previous analysis for DEGs of ESCA (Fig. 8). At this point, although 66/70
drug-associated target genes were not differentially expressed in
ESCA, they may be regulated by several drugs in order to affect the
pathways, including arachidonic acid metabolism, drug metabolism
(cytochrome P450) or CCKR signaling map, as well as BP, CC or MF,
particularly when regulated by the drug candidates that affect the
group of 51 connected genes. Once the drug candidate-associated
genes were directly associated with those ESCA hub genes, we
hypothesized that they would participate in some way in ESCA
pathogenesis. In such a situation, according to the results of the
present study, the predicted drugs could associate with ESCA.
Molecular docking between drug
candidates and hub genes
Docking results are presented in Table IV, among which SPP1 and
CXCL10 were eliminated because of unspecified protein
structures. The binding affinity between the predicted drug
candidates and hub genes was evaluated by the docking scores. The
score reported by the docking approach in systemsDock is a negative
logarithm of experimental dissociation/inhibition constant value
(pKd/pKi), usually ranging
between 0 and 10 (i.e., from weak to strong binding), allowing a
straightforward indication of binding strength. According to
Table IV, which lists strength by
a decreasing docking score, the molecular docking pairs with a
docking score of >5.52 were attributed to drugs including
apigenin, Prestwick-685 and menadione, which overall had the top
three scores in the CMAP results. The significant results were
plotted by respectively selecting the top three proteins for
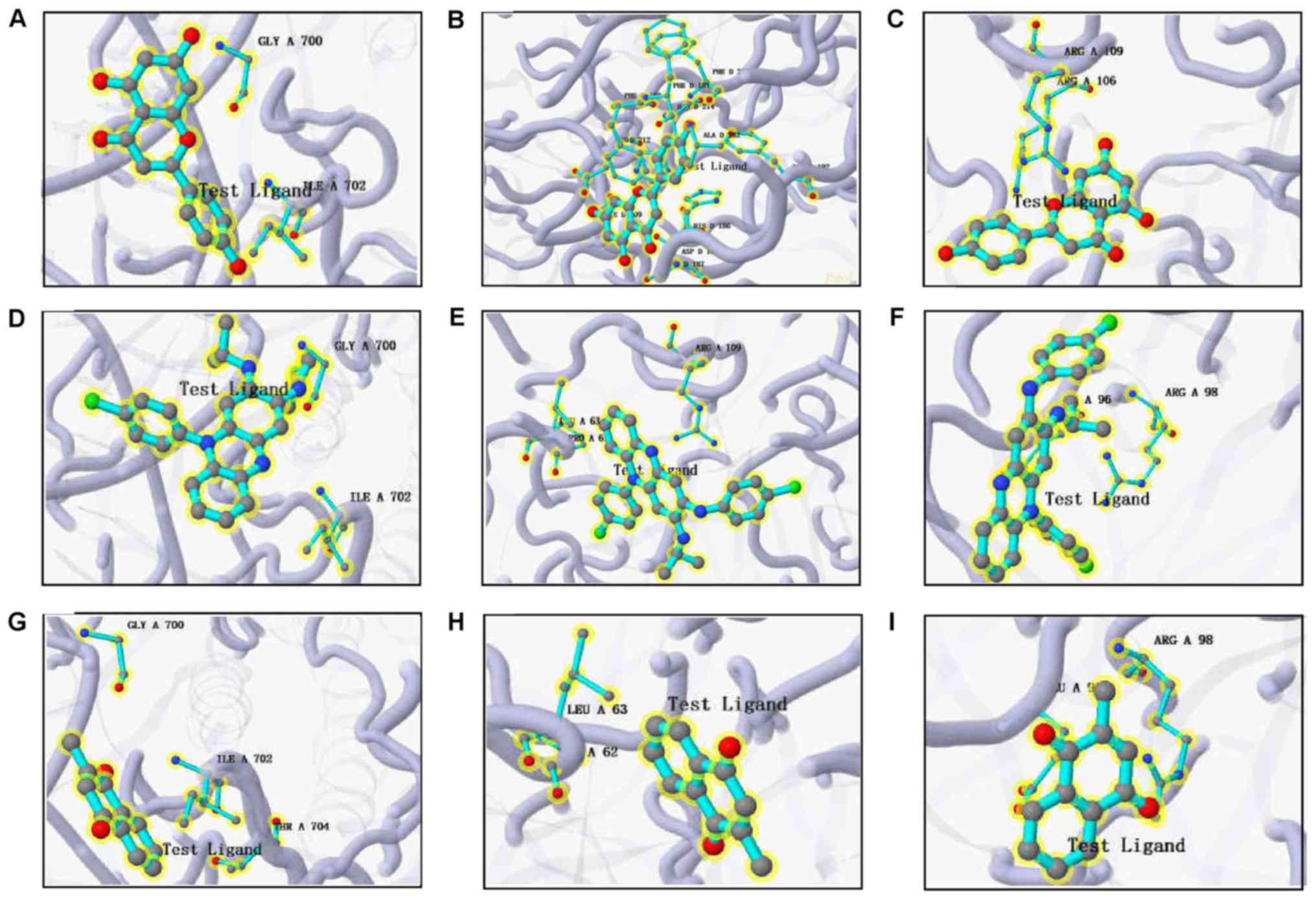
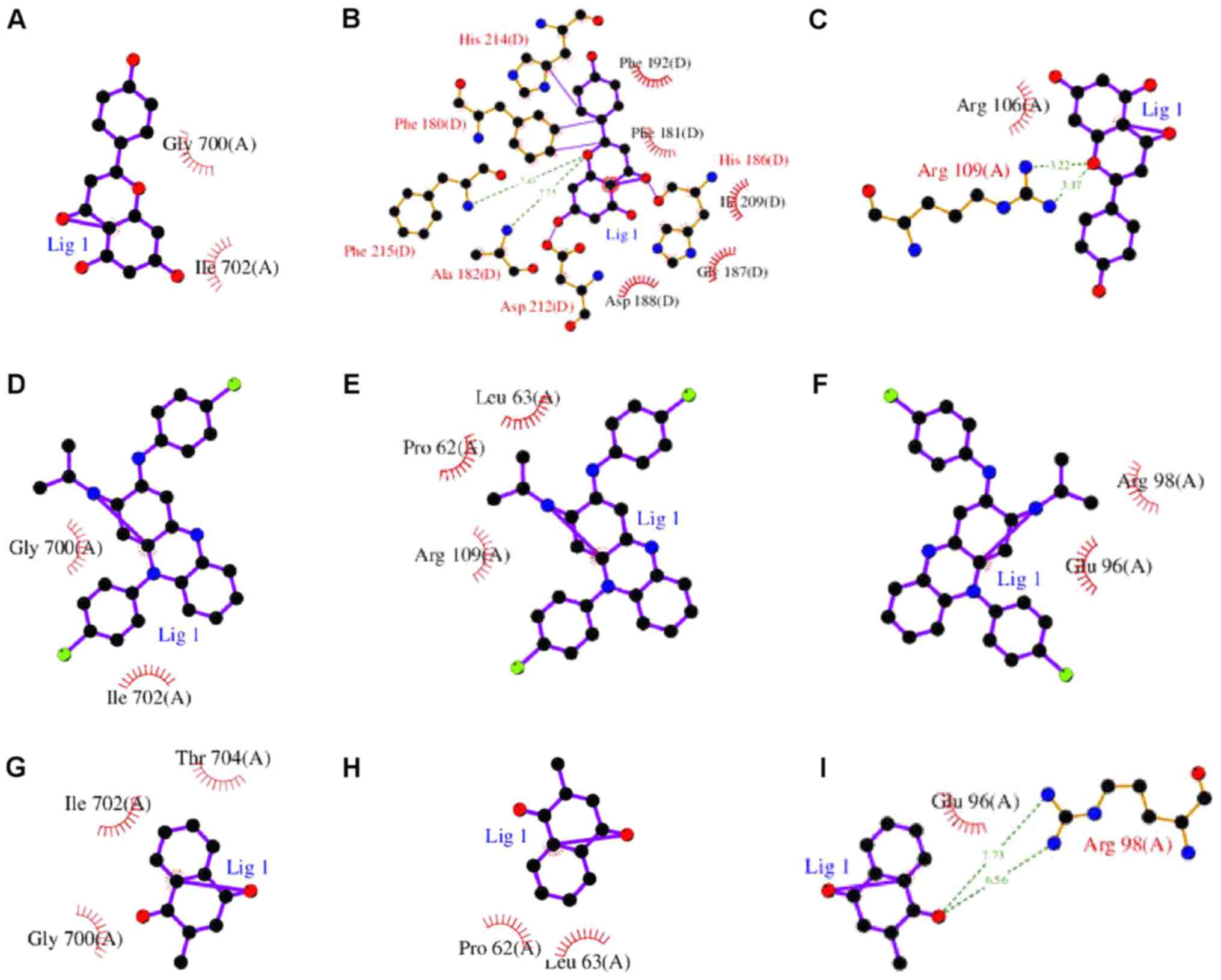
apigenin, Prestwick-685 or menadione (Figs. 9 and 10). A flow chart of the methodology used
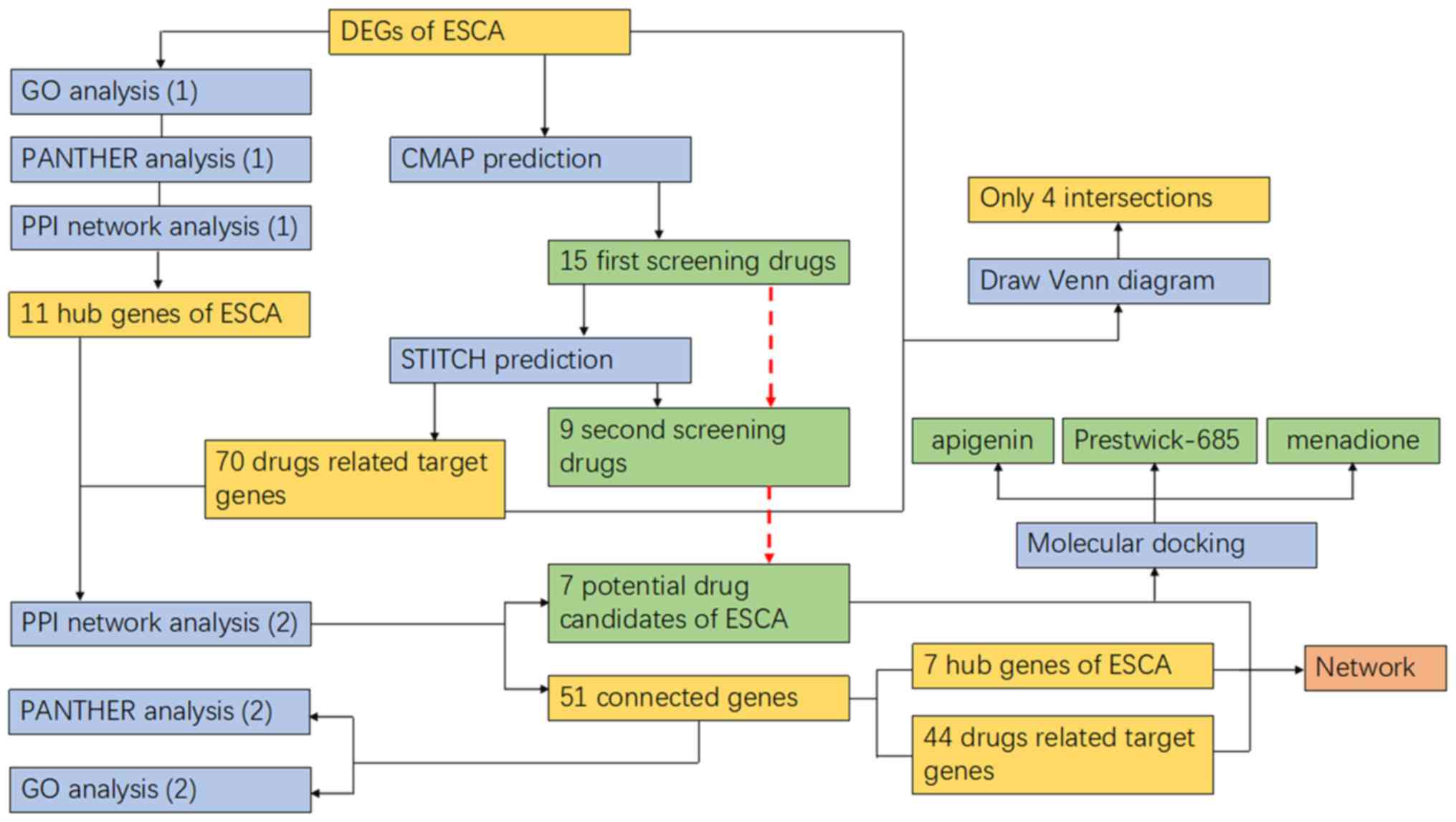
in the present study is summarized in Fig. 11.
| Table IVPredicted binding affinity between
drug candidates and hub genes. |
Table IV
Predicted binding affinity between
drug candidates and hub genes.
| Gene | PDB identifier | Test compound | Docking score
(pKd/pKi) |
|---|
| STAT1 | 1BF5 | 580043:
Apigenin | 7.866 |
| STAT1 | 1BF5 | 2794:
Prestwick-685 | 7.738 |
| MMP9 | 1ITV | 2794:
Prestwick-685 | 6.768 |
| TIMP1 | 3MA2 | 580043:
Apigenin | 6.593 |
| MMP9 | 1ITV | 580043:
Apigenin | 6.439 |
| IL1B | 2NVH | 580043:
Apigenin | 6.422 |
| STAT1 | 1BF5 | 4055:
Menadione | 6.403 |
| MMP9 | 1ITV | 4055:
Menadione | 5.612 |
| IL1B | 2NVH | 4055:
Menadione | 5.592 |
| IL8 | 1QE6 | 4055:
Menadione | 5.431 |
| IL1B | 2NVH | 2794:
Prestwick-685 | 5.417 |
| TIMP1 | 3MA2 | 4055:
Menadione | 5.191 |
| IL8 | 1QE6 | 580043:
Apigenin | 5.075 |
| IL8 | 1QE6 | 2794:
Prestwick-685 | 4.687 |
| IL8 | 1QE6 | 3475:
Gliclazide | 4.617 |
| TIMP1 | 3MA2 | 3475:
Gliclazide | 4.385 |
| TIMP1 | 3MA2 | 3121: Valproic
acid | 4.344 |
| STAT1 | 1BF5 | 3121: Valproic
acid | 4.23 |
| MMP9 | 1ITV | 3121: Valproic
acid | 4.131 |
| IL1B | 2NVH | 3121: Valproic
acid | 4.092 |
| IL8 | 1QE6 | 3121: Valproic
acid | 4.06 |
| TIMP1 | 3MA2 | 2794:
Prestwick-685 | 3.802 |
| TIMP1 | 3MA2 | 7184: Butyl
hydroxybenzoate | 3.73 |
| IL8 | 1QE6 | 7184: Butyl
hydroxybenzoate | 3.352 |
| IL1B | 2NVH | 7184: Butyl
hydroxybenzoate | 3.148 |
| STAT1 | 1BF5 | 7184: Butyl
hydroxybenzoate | 3.095 |
| IL8 | 1QE6 | 2284: Baclofen | 2.957 |
| IL1B | 2NVH | 3475:
Gliclazide | 2.894 |
| MMP9 | 1ITV | 3475:
Gliclazide | 2.871 |
| MMP9 | 1ITV | 7184: Butyl
hydroxybenzoate | 2.742 |
| TIMP1 | 3MA2 | 2284: Baclofen | 2.734 |
| STAT1 | 1BF5 | 3475:
Gliclazide | 2.695 |
| MMP9 | 1ITV | 2284: Baclofen | 2.571 |
| IL1B | 2NVH | 2284: Baclofen | 2.568 |
| STAT1 | 1BF5 | 2284: Baclofen | 2.371 |
Discussion
In the present study, 827 DEGs of ESCA were
identified, including 375 upregulated genes and 452 downregulated
genes, of which the functions were analyzed. On the basis of the
PPI network, 11 hub genes were extracted. To acquire potential
drugs for ESCA chemotherapy, the DEGs were uploaded to CMAP and 15
drugs were selected by scores. Furthermore, 9/15 drugs were
identified to target 70 genes in total. However, only four gene
intersections were identified in DEGs of ESCA and 70 target genes.
Acetazolamide and dequalinium were also eliminated by another PPI
analysis. As a result, apigenin, baclofen, Prestwick-685,
menadione, butyl hydroxybenzoate, gliclazide and valproate were
considered as the potential drug candidates, and the network of 7
predicted drug candidates-target genes-ESCA-associated hub genes
was constructed. Furthermore, apigenin, Prestwick-685 and menadione
were considered significant drug candidates by performing molecular
docking.
Thus, considering the research design, three steps
were taken to obtain the drug candidates for ESCA. First, full use
was made of the CMAP, and the first 15 screening drugs were
extracted by scores of <−0.85. CMAP is an online database that
provides an approach to identifying the connections between small
molecules, genes and diseases. It was created by the CMAP team at
the Broad Institute and the resource is publicly available
(portals.broadinstitute.org/cmap) (26). In CMAP, drug signatures are ordered
lists of genes that are stored following the profiling of 564
drug-treated instances and identified by a computer program.
Therefore, the CMAP can significantly increase the rate of drug
discovery and identify novel uses for existing drugs. For example,
Wang et al (38)
investigated candidate drugs for hepatocellular carcinoma in the
CMAP by analyzing associated subpathways. The CMAP also assisted
with the identification of similar signatures of atypical
meningiomas and provision of novel treatment methods (39). Additionally, Liu et al
(40) used CMAP to identify some
potential compounds for treating patients with B-cell chronic
lymphocytic leukemia (B-CLL) on the basis of altered pathways in
B-CLL. Similarly, in the present study, when 375 upregulated genes
and 452 downregulated genes of ESCA were uploaded as a probe set to
interrogate the CMAP, this dataset was matched with the drugs
identified using a Kolmogorov-Smirnov statistical test. The drugs
were sorted by their connectivity scores, which was the combination
of the up score and down score. The criterion of a score of
<−0.85 was also used to combine a high negative up score and a
high positive down score, indicating that the corresponding
perturbagen repressed the expression of the probe sets in the up
genes list, whereas it induced the expression of the probe sets in
the down genes list. Secondly, 6 drugs were eliminated when the
standard of STITCH was uniformly adapted. Thirdly, a new PPI
analysis was used to establish an association between target genes
predicted by the second screening drugs and DEGs of ESCA previously
obtained from TCGA and GTEx databases, of which the indicators were
artificially set with ‘experiments’ and ‘database’ for a higher
reliability. Therefore, those drugs corresponding to the certified
associated genes were finally retained as potential drug candidates
for ESCA.
The retrieval of relevant literature on drug
candidates was required to further understand their research status
and clinical application, as well as support the prediction. Among
the 7 predicted drugs, apigenin is a natural product that is
present in a number of plants. It was confirmed to have an
anticancer effect on different types of cancer (41-45).
Apigenin was once reported to induce apoptosis in ESCA cells
through destruction of the membrane structure (46). As Yan et al (47) summarized, apigenin may be an
adjuvant chemotherapeutic agent for cancer therapy by modulating
the phosphoinositide 3-kinase/protein kinase B, mitogen-activated
protein kinase/extracellular-signal-regulated kinase, Janus
kinase/signal transducer and activator of transcription, nuclear
factor-κB (NF-κB), and Wnt/β-catenin signaling pathways. In
addition, it was also reported that apigenin may have a further
function in cancer chemoprevention via the induction of apoptosis
and autophagy (48).
Baclofen is commonly used for treating spastic
movement disease (49,50). However, baclofen may help to
inhibit hiccups following the insertion of a stent for ESCC,
according to a case report (51),
and it was suggested that it may be a useful adjuvant analgesic for
decreasing cancer pain (52).
Whether baclofen has any anticancer effects or not remains unknown
owing to a lack of relevant research. Prestwick-685, also named
clofazimine in PubChem, was identified to be associated with breast
cancer (53) and lung cancer
(54). Recently, Durusu et
al (54) provided novel
evidence for the anticancer effect of clofazimine, which implies
that clofazimine also requires further investigation regarding
cancer treatment. Compared with clofazimine, it appears that
menadione, which is also known as vitamin K3, has attracted more
attention from the perspective of cancer treatment. Menadione has
been identified to exhibit significant therapeutic benefits in the
treatment of gastric cancer (55),
prostate cancer (56), oral cancer
(57) and breast cancer (58). Furthermore, the latest research
indicated that menadione may increase anticancer activity by
combining ROS-generating agents (59), but how it works in ESCA treatment
requires further investigation. Butyl hydroxybenzoate, also called
butylparaben, is often used as a flavoring additive in food.
However, it is reported to participate in a non-genomic mechanism
of activation of G-protein-coupled receptor 30 in cancerous and
non-cancerous breast cell lines through activation of the
extracellular-signal-regulated kinase 1/2 pathway; further research
into the carcinogenic action is required (60). Gliclazide is generally used for
treating diabetes (61,62), but its anticancer effect has seldom
been explored (63,64). Lastly, valproate has proved to be a
potent tool in treating cancer via enhancing doxorubicin
cytotoxicity and the efficacy of vinorelbine-cisplatin-based
chemoradiation, according to recent studies (65-68).
In summary, on the basis of the bioinformatics prediction of the
present study and aforementioned studies, apigenin, menadione and
valproate are hypothesized to have an increased possibility of
becoming the chemotherapeutics for ESCA.
Although a number of the predicted drug candidates
for ESCA have not exhibited specific anticancer effects previously,
drug repurposing was investigated via efficient bioinformatics
analysis and a structure exploration, which means that the
excavation of novel drug candidates in the present study was
primarily based on the drug repurposing. In particular, the
three-step filtering of drug candidates for ESCA executed the drug
repurposing simultaneously. In the present study, ESCA was
characterized by its DEGs, and the potential effects of drugs were
characterized by their target genes. To the extent of searching
drugs, the application of extracted DEGs was primarily utilized for
screening drugs via CMAP and posteriorly established linkages with
drug-associated target genes, which consequently made a connection
between ESCA and drugs. In line with this process, the two-time
functional analysis of two gene groups, of which one group contains
the 827 extracted DEGs of ESCA and the other group contained 51
genes that displayed in the drug candidates-target
genes-ESCA-associated hub genes network, explained the underlying
pathogenesis of ESCA and predicted the potential influence of
drugs. Results of pathway analysis coincided with the arachidonic
acid metabolism and drug metabolism (cytochrome P450), on the basis
of KEGG analysis, as well as with the CCKR signaling map, on the
basis of PANTHER analysis. According to GO analysis, the similar
results indicated that, among the drug-associated target genes, 66
unremarkable genes in ESCA, particularly those in the group of 51
connected genes, may be regulated by several drugs to affect the
BP, CC or MF and were associated with those hub genes of ESCA,
which, as a result, may influence ESCA pathogenesis.
In addition to the bioinformatics database analysis,
drug repurposing was also investigated via structure exploration.
Docking was used to screen the proteins and identify potential
targets for a query ligand (69).
Molecular docking is one of the key steps for proper computational
reconstruction of actual ligand-receptor or protein-protein
interactions, and thus is relevant to drug design and ligands
discovery (70-74). In the present study, the binding
affinity between drug molecules and proteins was determined.
Apigenin, Prestwick-685 and menadione were predicted to be
potential drugs that may affect the target genes, together with
ESCA-associated hub genes that influence pathogenesis. Higher
linking affinity in this step to hub genes in the network confirmed
apigenin, Prestwick-685 and menadione to be significant drug
candidates, as well as confirmed the CMAP results. Therefore, in
taking full advantage of databases and molecule docking, drug
repurposing of Prestwick-685 and menadione was achieved, as
apigenin has been reported to induce apoptosis in ESCA cells
(46). Thus, Prestwick-685 and
menadione were considered to be significant novel drug
candidates.
Furthermore, the biological significance of five
vital hub genes, including STAT1, matrix MMP9,
TIMP1, IL1B and IL8, for ESCA can be
explained. According to previous studies (75), STAT1 was reported to be a
tumor suppressor in ESCC by downregulating the activity of NF-κB
and STAT3, and it was associated with the substantial
downregulation of several pro-survival proteins such as B-cell
lymphoma (Bcl)-2, Bcl-extra-large and survivin, as well as
modulation of two G1 cell-cycle-regulatory proteins,
p21waf1 and cyclin D1 (76). MMP9 was also demonstrated to
be a prognostic factor in ESCA (77), and MMP9 upregulated by
protein tyrosine kinase 7 through activation of activator protein 1
and NF-κB could increase the invasive properties of ESCC cells
(78). Studies have identified
that high serum levels of TIMP1 were associated with tumor
progression and unfavorable prognosis in patients with ESCA
(79,80). For IL1B, it was only
reported that the rs16944 G>A polymorphism may contribute to
ESCC susceptibility (81). For
IL8, it is significantly upregulated during esophageal
carcinogenesis, and the IL-8/CXC chemokine receptor 2 signaling
pathway is a potential therapeutic target in EAC (82). To summarize, the significant
biological functions of STAT1, MMP9, TIMP1,
IL1B and IL8 in ESCA provided solid reasons for their
being therapeutic targets for patients with ESCA, for which the
novel drug candidates may serve certain positive functions in the
molecular docking, as predicted.
The present study had certain limitations. The
results relied heavily on the bioinformatics database and thus were
significantly influenced by the refresh rate of data. Furthermore,
drug experiments were not performed, meaning that the predicted
drugs may lack effective verification. Despite the limitations,
however, full use was made of the DEGs of ESCA in CMAP, STITCH, PPI
network and molecular docking, as well as functional analysis
including KEGG, PANTHER and GO analysis, to screen for novel drug
candidates for ESCA and have confirmed their potential therapeutic
effect at a bioinformatics level. Thus, to a certain extent, the
present study may provide a reference for the chemotherapy of ESCA,
in its molecular biology and methodology, which similar studies
have not yet reported. Furthermore, the results require further
investigation, as well as in vivo studies.
Abbreviations:
|
ESCA
|
esophageal carcinoma
|
|
DEG
|
differentially expressed genes
|
|
TCGA
|
The Cancer Genome Atlas
|
|
GTEx
|
Genotype‐Tissue Expression
|
|
PPI
|
protein‐protein interaction
|
|
CMAP
|
Connectivity Map
|
|
KEGG
|
Kyoto Encyclopedia of Genes and
Genomes
|
|
GO
|
Gene Ontology
|
|
BP
|
Biological Process
|
|
CC
|
Cellular Component
|
|
MF
|
Molecular Function
|
|
STITCH
|
Search Tool for Interactions of
Chemicals
|
|
SMILES
|
Simplified Molecular‐Input Line‐Entry
System
|
Acknowledgments
The authors thank Dr Xia Yang (Department of
Pathology, First Affiliated Hospital of Guangxi Medical University,
Nanning, China), for providing valuable suggestions on writing the
original manuscript.
Funding
The present study was supported by the Project of
Future Academic Star in Guangxi Medical University, China (grant
no. WLXSZX18004) and the Guangxi Zhuang Autonomous Region
University Student Innovative Plan, China (grant no. 2018049).
Availability of data and materials
The datasets generated and analyzed during the
current study are available in TCGA database (cancergenome.nih.gov), GTEx database (gtexportal.org/home) and CMAP database (portals.broadinstitute.org/cmap).
Authors' contributions
The study was conceived and designed by YTC, JYX, QS
and WJM. YTC, JYX and QS participated in the data mining. YTC
analyzed and interpreted data and drafted the manuscript. JYX
participated in drafting the manuscript. All authors read and
approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Ma K, Cao B and Guo M: The detective,
prognostic, and predictive value of DNA methylation in human
esophageal squamous cell carcinoma. Clin Epigenetics. 8:432016.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2018. CA Cancer J Clin. 68:7–30. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Erkizan HV, Johnson K, Ghimbovschi S,
Karkera D, Trachiotis G, Adib H, Hoffman EP and Wadleigh RG:
African-American esophageal squamous cell carcinoma expression
profile reveals dysregulation of stress response and detox
networks. BMC Cancer. 17:4262017. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Jiang S, Zhang Q, Su Y and Pan L:
Network-based differential analysis to identify molecular features
of tumorigenesis for esophageal squamous carcinoma. Molecules.
23:232018. View Article : Google Scholar
|
|
5
|
Yang XX, Ma M, Sang MX, Wang XX, Song H,
Liu ZK and Zhu SC: Radiosensitization of esophageal carcinoma cells
by knockdown of RNF2 expression. Int J Oncol. 48:1985–1996. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Ma Z, Cai H and Cui Y: Progress in the
treatment of esophageal neuroendocrine carcinoma. Tumour Biol.
39:10104283177113132017. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Vellayappan BA, Soon YY, Ku GY, Leong CN,
Lu JJ and Tey JC: Chemoradiotherapy versus chemoradiotherapy plus
surgery for esophageal cancer. Cochrane Database Syst Rev.
8:CD0105112017.PubMed/NCBI
|
|
8
|
Chen GZ, Zhu HC, Dai WS, Zeng XN, Luo JH
and Sun XC: The mechanisms of radioresistance in esophageal
squamous cell carcinoma and current strategies in radiosensitivity.
J Thorac Dis. 9:849–859. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Recio-Boiles A and Babiker HM: Cancer,
Esophageal. StatPearls [Internet]. StatPearls Publishing; Treasure
Island, FL: 2018
|
|
10
|
Sasaki Y and Kato K: Chemoradiotherapy for
esophageal squamous cell cancer. Jpn J Clin Oncol. 46:805–810.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Harada K, Mizrak Kaya D, Baba H and Ajani
JA: Immune checkpoint blockade therapy for esophageal squamous cell
carcinoma. J Thorac Dis. 10:699–702. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Cai Z, Lv H, Cao W, Zhou C, Liu Q, Li H
and Zhou F: Targeting strategies of adenovirus mediated gene
therapy and virotherapy for prostate cancer (Review). Mol Med Rep.
16:6443–6458. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Grisham RN and Iyer G: Low-grade serous
ovarian cancer: Current treatment paradigms and future directions.
Curr Treat Options Oncol. 19:542018. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Le Grazie M, Biagini MR, Tarocchi M,
Polvani S and Galli A: Chemotherapy for hepatocellular carcinoma:
The present and the future. World J Hepatol. 9:907–920. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Mayank and Jaitak V: Drug target
strategies in breast cancer treatment: Recent developments.
Anticancer Agents Med Chem. 14:1414–1427. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Michels S and Wolf J: Stratified treatment
in lung cancer. Oncol Res Treat. 39:760–766. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Jia Y, Xiao Z, Gongsun X, Xin Z, Shang B,
Chen G, Wang Z and Jiang W: CEP55 promotes the proliferation,
migration and invasion of esophageal squamous cell carcinoma via
the PI3K/Akt pathway. OncoTargets Ther. 11:4221–4232. 2018.
View Article : Google Scholar
|
|
18
|
Niyaz M, Abdurahman A, Turghun A and Awut
I: CEP3 and CEP17 DNA probe potential in the genetic diagnosis and
prognostic prediction of esophageal squamous cell cancer. Exp Ther
Med. 11:1375–1380. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Wang H, Zhou Y, Liu Q, Xu J and Ma Y:
Prognostic value of SOX2, Cyclin D1, P53, and ki-67 in patients
with esophageal squamous cell carcinoma. OncoTargets Ther.
11:5171–5181. 2018. View Article : Google Scholar
|
|
20
|
Lin S, Liu K, Zhang Y, Jiang M, Lu R,
Folts CJ, Gao X, Noble MD, Zhao T, Zhou Z, et al: Pharmacological
targeting of p38 MAP-Kinase 6 (MAP2K6) inhibits the growth of
esophageal adenocarcinoma. Cell Signal. 51:222–232. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
MacGregor TP, Carter R, Gillies RS,
Findlay JM, Kartsonaki C, Castro-Giner F, Sahgal N, Wang LM, Chetty
R, Maynard ND, et al: Translational study identifies XPF and MUS81
as predictive biomarkers for oxaliplatin-based peri-operative
chemotherapy in patients with esophageal adenocarcinoma. Sci Rep.
8:72652018. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Chang ZW, Jia YX, Zhang WJ, Song LJ, Gao
M, Li MJ, Zhao RH, Li J, Zhong YL, Sun QZ, et al:
LncRNA-TUSC7/miR-224 affected chemotherapy resistance of esophageal
squamous cell carcinoma by competitively regulating DESC1. J Exp
Clin Cancer Res. 37:562018. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Liu B, Wang C, Chen P, Cheng B and Cheng
Y: RACKI induces chemotherapy resistance in esophageal carcinoma by
upregulating the PI3K/AKT pathway and Bcl-2 expression. OncoTargets
Ther. 11:211–220. 2018. View Article : Google Scholar
|
|
24
|
Yu X, Li W, Xia Z, Xie L, Ma X, Liang Q,
Liu L, Wang J, Zhou X, Yang Y, et al: Targeting MCL-1 sensitizes
human esophageal squamous cell carcinoma cells to cisplatin-induced
apoptosis. BMC Cancer. 17:4492017. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Pang Y, Liu J, Li X, Zhang Y, Zhang B,
Zhang J, Du N, Xu C, Liang R, Ren H, et al: Nano Let 7b
sensitization of eliminating esophageal cancer stem like cells is
dependent on blockade of Wnt activation of symmetric division. Int
J Oncol. 51:1077–1088. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Lamb J, Crawford ED, Peck D, Modell JW,
Blat IC, Wrobel MJ, Lerner J, Brunet JP, Subramanian A, Ross KN, et
al: The Connectivity Map: Using gene-expression signatures to
connect small molecules, genes, and disease. Science.
313:1929–1935. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Lin P, Xiong DD, Dang YW, Yang H, He Y,
Wen DY, Qin XG and Chen G: The anticipating value of PLK1 for
diagnosis, progress and prognosis and its prospective mechanism in
gastric cancer: A comprehensive investigation based on
high-throughput data and immunohistochemical validation.
Oncotarget. 8:92497–92521. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Dang YW, Lin P, Liu LM, He RQ, Zhang LJ,
Peng ZG, Li XJ and Chen G: In silico analysis of the potential
mechanism of telocinobufagin on breast cancer MCF-7 cells. Pathol
Res Pract. 214:631–643. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
He RQ, Yang X, Liang L, Chen G and Ma J:
MicroRNA-124-3p expression and its prospective functional pathways
in hepatocellular carcinoma: A quantitative polymerase chain
reaction, gene expression omnibus and bioinformatics study. Oncol
Lett. 15:5517–5532. 2018.PubMed/NCBI
|
|
30
|
Li HM, Yang H, Wen DY, Luo YH, Liang CY,
Pan DH, Ma W, Chen G, He Y and Chen JQ: Overexpression of LncRNA
HOTAIR is associated with poor prognosis in thyroid carcinoma: A
study based on TCGA and GEO data. Horm Metab Res. 49:388–399. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Shannon P, Markiel A, Ozier O, Baliga NS,
Wang JT, Ramage D, Amin N, Schwikowski B and Ideker T: Cytoscape: A
software environment for integrated models of biomolecular
interaction networks. Genome Res. 13:2498–2504. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Gao L, Li SH, Tian YX, Zhu QQ, Chen G,
Pang YY and Hu XH: Role of downregulated miR-133a-3p expression in
bladder cancer: A bioinformatics study. OncoTargets Ther.
10:3667–3683. 2017. View Article : Google Scholar
|
|
33
|
He RQ, Wu PR, Xiang XL, Yang X, Liang HW,
Qiu XH, Yang LH, Peng ZG and Chen G: Downregulated miR-23b-3p
expression acts as a predictor of hepatocellular carcinoma
progression: A study based on public data and RT-qPCR verification.
Int J Mol Med. 41:2813–2831. 2018.PubMed/NCBI
|
|
34
|
Liang L, Wei DM, Li JJ, Luo DZ, Chen G,
Dang YW and Cai XY: Prognostic microRNAs and their potential
molecular mechanism in pancreatic cancer: A study based on The
Cancer Genome Atlas and bioinformatics investigation. Mol Med Rep.
17:939–951. 2018.
|
|
35
|
Pathan M, Keerthikumar S, Ang CS, Gangoda
L, Quek CY, Williamson NA, Mouradov D, Sieber OM, Simpson RJ, Salim
A, et al: FunRich: An open access standalone functional enrichment
and interaction network analysis tool. Proteomics. 15:2597–2601.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Hsin KY, Ghosh S and Kitano H: Combining
machine learning systems and multiple docking simulation packages
to improve docking prediction reliability for network pharmacology.
PLoS One. 8:e839222013. View Article : Google Scholar
|
|
37
|
Hsin KY, Matsuoka Y, Asai Y, Kamiyoshi K,
Watanabe T, Kawaoka Y and Kitano H: systemsDock: A web server for
network pharmacology-based prediction and analysis. Nucleic Acids
Res. 44(W1): W507–13. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Wang J, Li M, Wang Y and Liu X:
Integrating subpathway analysis to identify candidate agents for
hepatocellular carcinoma. OncoTargets Ther. 9:1221–1230. 2016.
View Article : Google Scholar
|
|
39
|
Zador Z, King AT and Geifman N: New drug
candidates for treatment of atypical meningiomas: An integrated
approach using gene expression signatures for drug repurposing.
PLoS One. 13:e01947012018. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Liu G, Hu X, Gao L and Feng Z:
Personalized drug analysis in B cell chronic lymphocytic leukemia
patients. Med Sci Monit. 23:2159–2167. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Nelson N, Szekeres K, Iclozan C, Rivera
IO, McGill A, Johnson G, Nwogu O and Ghansah T: Apigenin: Selective
CK2 inhibitor increases Ikaros expression and improves T cell
homeostasis and function in murine pancreatic cancer. PLoS One. 12.
pp. e01701972017, View Article : Google Scholar
|
|
42
|
Pal MK, Jaiswar SP, Dwivedi A, Goyal S,
Dwivedi VN, Pathak AK, Kumar V, Sankhwar PL and Ray RS: Synergistic
effect of graphene oxide coated nanotised apigenin with paclitaxel
(GO-NA/PTX): A ROS dependent mitochondrial mediated apoptosis in
ovarian cancer. Anticancer Agents Med Chem. 17:1721–1732.
2017.PubMed/NCBI
|
|
43
|
Shan S, Shi J, Yang P, Jia B, Wu H, Zhang
X and Li Z: Apigenin restrains colon cancer cell proliferation via
targeted blocking of pyruvate kinase M2-dependent glycolysis. J
Agric Food Chem. 65:8136–8144. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Vrhovac Madunić I, Madunić J, Antunović M,
Paradžik M, Garaj-Vrhovac V, Breljak D, Marijanović I and Gajski G:
Apigenin, a dietary flavonoid, induces apoptosis, DNA damage, and
oxidative stress in human breast cancer MCF-7 and MDA MB-231 cells.
Naunyn Schmiedebergs Arch Pharmacol. 391:537–550. 2018. View Article : Google Scholar
|
|
45
|
Zhou Z, Tang M, Liu Y, Zhang Z, Lu R and
Lu J: Apigenin inhibits cell proliferation, migration, and invasion
by targeting Akt in the A549 human lung cancer cell line.
Anticancer Drugs. 28:446–456. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Zhu H, Jin H, Pi J, Bai H, Yang F, Wu C,
Jiang J and Cai J: Apigenin induced apoptosis in esophageal
carcinoma cells by destruction membrane structures. Scanning.
38:322–328. 2016. View Article : Google Scholar
|
|
47
|
Yan X, Qi M, Li P, Zhan Y and Shao H:
Apigenin in cancer therapy: Anti-cancer effects and mechanisms of
action. Cell Biosci. 7:502017. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Sung B, Chung HY and Kim ND: Role of
apigenin in cancer prevention via the induction of apoptosis and
autophagy. J Cancer Prev. 21:216–226. 2016. View Article : Google Scholar
|
|
49
|
Pérez-Arredondo A, Cázares-Ramírez E,
Carrillo-Mora P, Martínez-Vargas M, Cárdenas-Rodríguez N,
Coballase-Urrutia E, Alemón-Medina R, Sampieri A III, Navarro L and
Carmona-Aparicio L: Baclofen in the therapeutic of sequele of
traumatic brain injury: Spasticity. Clin Neuropharmacol.
39:311–319. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Ertzgaard P, Campo C and Calabrese A:
Efficacy and safety of oral baclofen in the management of
spasticity: A rationale for intrathecal baclofen. J Rehabil Med.
49:193–203. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Sharma V, De A, Lamoria S and Lamba BM:
Baclofen-responsive hiccups after esophageal stenting for
malignancy-related dysphagia. Proc Bayl Univ Med Cent. 29:1502016.
View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Yomiya K, Matsuo N, Tomiyasu S, Yoshimoto
T, Tamaki T, Suzuki T and Matoba M: Baclofen as an adjuvant
analgesic for cancer pain. Am J Hosp Palliat Care. 26:112–118.
2009. View Article : Google Scholar
|
|
53
|
Koval AV, Vlasov P, Shichkova P,
Khunderyakova S, Markov Y, Panchenko J, Volodina A, Kondrashov FA
and Katanaev VL: Anti-leprosy drug clofazimine inhibits growth of
triple-negative breast cancer cells via inhibition of canonical Wnt
signaling. Biochem Pharmacol. 87:571–578. 2014. View Article : Google Scholar
|
|
54
|
Durusu IZ, Hüsnügil HH, Ataş H, Biber A,
Gerekçi S, Güleç EA and Özen C: Anti-cancer effect of clofazimine
as a single agent and in combination with cisplatin on U266
multiple myeloma cell line. Leuk Res. 55:33–40. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Lee MH, Cho Y, Kim DH, Woo HJ, Yang JY,
Kwon HJ, Yeon MJ, Park M, Kim SH, Moon C, et al: Menadione induces
G2/M arrest in gastric cancer cells by downregulation of CDC25C and
proteasome mediated degradation of CDK1 and cyclin B1. Am J Transl
Res. 8:5246–5255. 2016.
|
|
56
|
Prasad CV, Nayak VL, Ramakrishna S and
Mallavadhani UV: Novel menadione hybrids: Synthesis, anticancer
activity, and cell-based studies. Chem Biol Drug Des. 91:220–233.
2018. View Article : Google Scholar
|
|
57
|
Delwar ZM, Siden A, Cruz MH and Yakisich
JS: Menadione : Sodium orthovanadate combination eliminates and
inhibits migration of detached cancer cells. ISRN Pharmacol.
2012.307102:2012.
|
|
58
|
Yamada A, Osada S, Tanahashi T, Matsui S,
Sasaki Y, Tanaka Y, Okumura N, Matsuhashi N, Takahashi T, Yamaguchi
K, et al: Novel therapy for locally advanced triple-negative breast
cancer. Int J Oncol. 47:1266–1272. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Teixeira J, Amorim R, Santos K, Soares P,
Datta S, Cortopassi GA, Serafim TL, Sardão VA, Garrido J, Borges F,
et al: Disruption of mitochondrial function as mechanism for
anti-cancer activity of a novel mitochondriotropic menadione
derivative. Toxicology. 393:123–139. 2018. View Article : Google Scholar
|
|
60
|
Wróbel AM and Gregoraszczuk EL: Action of
methyl-, propyl- and butylparaben on GPR30 gene and protein
expression, cAMP levels and activation of ERK1/2 and PI3K/Akt
signaling pathways in MCF-7 breast cancer cells and MCF-10A
non-transformed breast epithelial cells. Toxicol Lett. 238:110–116.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Leiter LA, Shestakova MV and Satman I:
Effectiveness of gliclazide MR 60 mg in the management of type 2
diabetes: Analyses from the EASYDia trial. Diabetol Metab Syndr.
10:302018. View Article : Google Scholar
|
|
62
|
Mikov M, Danic M, Pavlovic N, Stanimirov
B, Goločorbin-Kon S, Stankov K and Al-Salami H: Potential
applications of gliclazide in treating type 1 diabetes mellitus:
Formulation with bile acids and probiotics. Eur J Drug Metab
Pharmacokinet. 43:269–280. 2018. View Article : Google Scholar
|
|
63
|
Sliwinska A, Rogalska A, Szwed M,
Kasznicki J, Jozwiak Z and Drzewoski J: Gliclazide may have an
antiapoptotic effect related to its antioxidant properties in human
normal and cancer cells. Mol Biol Rep. 39:5253–5267. 2012.
View Article : Google Scholar :
|
|
64
|
Sliwinska A, Sliwinski T, Kasznicki J and
Drzewoski J: Effect of gliclazide on nucleotide excision repair
(NER) and non-homologous DNA end joining (NHEJ) in normal and
cancer cells. J Physiol Pharmacol. 61:347–353. 2010.PubMed/NCBI
|
|
65
|
Caponigro F, Di Gennaro E, Ionna F, Longo
F, Aversa C, Pavone E, Maglione MG, Di Marzo M, Muto P, Cavalcanti
E, et al: Phase II clinical study of valproic acid plus cisplatin
and cetuximab in recurrent and/or metastatic squamous cell
carcinoma of Head and Neck-V-CHANCE trial. BMC Cancer. 16:9182016.
View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Scialdone A, Hasni MS, Damm JK,
Lennartsson A, Gullberg U and Drott K: The HDAC inhibitor valproate
induces a bivalent status of the CD20 promoter in CLL patients
suggesting distinct epigenetic regulation of CD20 expression in CLL
in vivo. Oncotarget. 8:37409–37422. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Sha S, Zhai Y, Lin C, Wang H, Chang Q,
Song S, Ren M and Liu G: A combination of valproic acid sodium
salt, CHIR99021, E-616452, tranylcypromine, and 3-Deazaneplanocin A
causes stem cell-like characteristics in cancer cells. Oncotarget.
8:53302–53312. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Wei M, Mao S, Lu G, Li L, Lan X, Huang Z,
Chen Y, Zhao M, Zhao Y and Xia Q: Valproic acid sensitizes
metformin-resistant human renal cell carcinoma cells by
upregulating H3 acetylation and EMT reversal. BMC Cancer.
18:4342018. View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Xu X, Huang M and Zou X: Docking-based
inverse virtual screening: Methods, applications, and challenges.
Biophys Rep. 4:1–16. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Abdolmaleki A, Ghasemi JB and Ghasemi F:
Computer aided drug design for multi-target drug design: SAR/QSAR,
molecular docking and pharmacophore methods. Curr Drug Targets.
18:556–575. 2017. View Article : Google Scholar
|
|
71
|
Bartuzi D, Kaczor AA, Targowska-Duda KM
and Matosiuk D: Recent advances and applications of molecular
docking to G protein-coupled receptors. Molecules. 22:222017.
View Article : Google Scholar
|
|
72
|
de Ruyck J, Brysbaert G, Blossey R and
Lensink MF: Molecular docking as a popular tool in drug design, an
in silico travel. Adv Appl Bioinform Chem. 9:1–11. 2016.PubMed/NCBI
|
|
73
|
Rosano C, Ponassi M, Santolla MF, Pisano
A, Felli L, Vivacqua A, Maggiolini M and Lappano R: Macromolecular
modelling and docking simulations for the discovery of selective
GPER ligands. AAPS J. 18:41–46. 2016. View Article : Google Scholar :
|
|
74
|
Scotti L, Mendonca Junior FJ, Ishiki HM,
Ribeiro FF, Singla RK, Barbosa Filho JM, Da Silva MS and Scotti MT:
Docking studies for multi-target drugs. Curr Drug Targets.
18:592–604. 2017. View Article : Google Scholar
|
|
75
|
Zhang Y, Molavi O, Su M and Lai R: The
clinical and biological significance of STAT1 in esophageal
squamous cell carcinoma. BMC Cancer. 14:7912014. View Article : Google Scholar : PubMed/NCBI
|
|
76
|
Zhang Y, Zhang Y, Yun H, Lai R and Su M:
Correlation of STAT1 with apoptosis and cell-cycle markers in
esophageal squamous cell carcinoma. PLoS One. 9:e1139282014.
View Article : Google Scholar : PubMed/NCBI
|
|
77
|
Klimczak-Bitner AA, Kordek R, Bitner J,
Musiał J and Szemraj J: Expression of MMP9, SERPINE1 and miR-134 as
prognostic factors in esophageal cancer. Oncol Lett. 12:4133–4138.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
78
|
Shin WS, Hong Y, Lee HW and Lee ST:
Catalytically defective receptor protein tyrosine kinase PTK7
enhances invasive phenotype by inducing MMP-9 through activation of
AP-1 and NF-κB in esophageal squamous cell carcinoma cells.
Oncotarget. 7:73242–73256. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
79
|
Juchniewicz A, Kowalczuk O, Milewski R,
Laudański W, Dzięgielewski P, Kozłowski M and Nikliński J: MMP-10,
MMP-7, TIMP-1 and TIMP-2 mRNA expression in esophageal cancer. Acta
Biochim Pol. 64:295–299. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
80
|
Kozłowski M, Laudański W, Mroczko B,
Szmitkowski M, Milewski R and Łapuć G: Serum tissue inhibitor of
metallo-proteinase 1 (TIMP-1) and vascular endothelial growth
factor A (VEGF-A) are associated with prognosis in esophageal
cancer patients. Adv Med Sci. 58:227–234. 2013. View Article : Google Scholar
|
|
81
|
Zheng L, Yin J, Wang L, Wang X, Shi Y,
Shao A, Tang W, Ding G, Liu C, Chen S, et al: Interleukin 1B
rs16944 G>A polymorphism was associated with a decreased risk of
esophageal cancer in a Chinese population. Clin Biochem.
46:1469–1473. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
82
|
Shrivastava MS, Hussain Z, Giricz O,
Shenoy N, Polineni R, Maitra A and Verma A: Targeting chemokine
pathways in esophageal adenocarcinoma. Cell Cycle. 13:3320–3327.
2014. View Article : Google Scholar : PubMed/NCBI
|