Introduction
Epithelial-to-mesenchymal transition (EMT) is a
multi-step process that converts epi thelial cells into mesenchymal
cells, which is mediated by series of transcription factors (TFs)
(1). EMT has been linked to
epithelial tumor characteristics, including the metastasis and
invasiveness of gastric cancer (GC) cells (2-5).
E-cadherin is an important EMT TF and epithelial factor that
contributes to cell adhesions and cell junctions between adjacent
cells (6). However, the molecular
mechanisms of EMT require further exploration to reveal its
association with the early metastasis of tumor cells.
Long non-coding RNA HOX transcript antisense
intergenic RNA (HOTAIR) is a functional RNA that does not encode a
protein, and was first identified by Rinn et al (7) in 2007. Long non-coding RNA HOTAIR is
transcribed from one of the chromosomal loci, the HOXC locus, which
contains the clustered HOX genes (8). HOTAIR is associated with different
types of cancer, including breast cancer, gastrointestinal tumors,
colorectal cancer and pancreatic cancer, and the overexpression of
HOTAIR has also been shown to be associated with a poor prognosis
and with increased metastasis (9-11).
HOTAIR promotes EMT in various types of cancer through different
mechanisms. It has previously been identified that HOTAIR regulates
glioblastoma (GBM) invasion by increasing the expression of
β-catenin (12). A previous study
also demonstrated that HOTAIR serves as a modular scaffold for the
polycomb repressive complex 2 (PRC2) complex, which consists of
EZH2, SUZ12 and EED, and the LSD1/CoREST/REST complex, both of
which are histone modification complexes (13). The methyltransferase EZH2 catalyzes
H3K27 tri-methylation and induces the epigenetic silencing of gene
expression (14). Based on this
mechanism, HOTAIR epigenetically represses numerous factors,
including microRNA (miRNA or miR)-200 (15) and miR-34a (16), which contribute to the GC cell-EMT
process. Notably, long intergenic non-coding RNA HOTAIR is
associated with EMT in multiple cancer types, including GC, through
different mechanisms, but most importantly, through the epigenetic
regulation of gene expression.
Epigenetics refers to the regulation of genetic
function and expression, which is mediated through non-DNA-encoded
mechanisms (17). An important
type of epigenetic modification is histone modification, which is a
post-translational modification mediated by histone-modifying
enzymes, including methylation and acetylation. Histone lysine
methylation includes mono-, di- and tri-methylation at lysine
residues. The methylation event is catalyzed by a group of lysine
methyltransferases (KMTs). Notably, histone acetylation requires
histone acetyltransferases (HATs) (18,19).
Histone modifications regulate transcriptional activation or
repression. For example, H3K27me3 is associated with gene
repression, and H3K27ac is a hallmark of transcriptionally active
genes (20,21). It is important to understand the
association between histone modification and the mechanisms of
tumorigenesis, which may provide new insight into effective
treatment strategies and may improve the prognosis of epithelial
tumors.
Recent studies have suggested that HOTAIR is
involved in the development of GC and the overexpression of HOTAIR
is positively associated with a poor clinical outcome (22-24)
In this study, we explored the association between HOTAIR and EMT,
particularly as regards the effects on E-cadherin expression. As
HOTAIR targets PRC2 (EZH2) to catalyze H3K27me3, we examined
whether HOTAIR influences the acetylation of H3K27 by indirectly
interacting with HAT. We attempted to connect the HOTAIR-mediated
antagonistic switch between histone H3K27 methylation and
acetylation at the E-cadherin promoter with the expression of
E-cadherin. Our study provides new insight into the mechanisms
through which HOTAIR regulates EMT and promotes the carcinogenesis
and progression of GC.
Materials and methods
Cell culture
The human gastric cancer cell lines, MGC-803 and
SGC-7901 were purchased from China Academia Sinica (Shanghai,
China). All cells were cultured in DMEM and RPMI-1640 medium
supplemented with 10% FBS (HyClone, Logan, UT, USA) and were
maintained in a humidified atmosphere with 5% CO2 at
37˚C.
Lentiviral infection and transient
transfection
Lentiviruses containing the HOTAIR overexpression
sequence (Lenti-HOTAIR), a HOTAIR inhibitor sequence (Lenti-HOTAIR
si) and a negative control sequence (Lenti-NC and NC) were obtained
from Shanghai GenePharma (Shanghai, China). For the transient
transfection of cells, the tumor cells were transfected with
negative control siRNA oligonucleotides against EZH2 and SUZ12
using X-treme GENE siRNA transfection reagent (Roche, Shanghai,
China) according to the manufacturer’s instructions. The siRNA
sequences were as follows: si-EZH2, 5′GUCGCAACGGACCAGUUAA-3′; and
si-SUZ12, 5′-GCTTACGTTTACTGGTTTCTT-3′. Subsequent experiments were
then carried out 48 h following transfection.
Western blot analysis
RIPA lysis buffer was used to extract the total
protein, and the Nuclear and Cytoplasmic Protein Extraction kit
(Beyotime Biotechnology, Shanghai, China) was used to extract the
nuclear protein, according to the manufacturer’s instructions.
Equal amounts of protein per lane (30 µg) were separated by
10 and 15% SDS-polyacrylamide gel and transferred onto PVDF
membranes. The membranes were blocked in 5% skim milk at 37˚C for 1
h and then incubated with a specific antibody at 4˚C overnight. The
antibodies used in this study were as follows: H3K4me3 (ab8580),
H3K4ac (ab240195), H3K27me2 (ab24684), H3K27me3 (ab6002), HEK27ac
(ab45173) and β-catenin (ab16051) antibodies (Abcam, Cambridge, UK;
1 µg/ml); EZH2 (5246), SUZ12 (3737), CBP (7389), E-cadherin
(14472), N-cadherin (13116), snail (3879), slug (9585), twist
(46702), H3K4me1 (5326), H3K4me2 (9725) and H3K27me1 (84932) (Cell
Signaling Technology, Danvers, MA, USA; 1 µg/ml); H3
(AF0009; Beyotime Biotechnology; 0.2 µg/ml); GAPDH (TA-08;
Zhongshan Bio Corporation, Beijing, China; 0.2 µg/ml). They
were then incubated with their respective secondary antibodies
(ZB-2305, ZB2306; Zhongshan Bio Corporation; 1:2,000 dilution). The
membranes were stripped, and the bands were visualized using the
SuperSignal protein detection kit (Pierce/Thermo Fisher Scientific,
Waltham, MA, USA) following normalization with the density of GAPDH
and H3. ImageJ 1.41 software (National Institutes of Health,
Bethesda, MD, USA) was employed to quantify protein expression.
RNA isolation and RT-qPCR
TRIzol reagent (Invitrogen/Thermo Fisher Scientific)
was used to extract the RNA from the gastric cancer cells. Equal
amounts of RNA were converted into cDNA using HOTAIR and CDH1
primers by using a PrimeScript RT Reagent kit (Takara Bio, Inc.,
Otsu, Japan). Quantitative PCR (qPCR) was performed using a Reverse
Transcription System (Promega, Madison, WI, USA) on a CFX96
Real-Time PCR Detection System (Bio-Rad Laboratories, Hercules, CA,
USA) according to the instructions of the manufacturer. Each sample
was analyzed in triplicate. The primers used were as follows:
HOTAIR forward, 5′-AAATATGGCGGCGTCTACACGGA-3′ and reverse,
5′-TCCAGAACCCTCTGACATTTGCCT-3′; CDH1 forward,
5′-ACGCCGAGAGCTACACGTTC-3′ and reverse, 5′-AGGCTGTCCTTTGTCGACCG-3′;
GAPDH forward, 5′-CCGGGAAACTGTGGCGTGATGG-3′ and reverse,
5′-AGGTGGAGGAGTGGGTGTCGCTGTT-3′. Relative expression was calculated
using the 2−ΔΔCq method (25). The PCR conditions were as follows:
35 cycles of 95˚C for 30 sec, 56˚C for 30 sec, 72˚C for 90 sec, and
a final extension at 72˚C for 5 min.
Immunofluorescence
The MGC-803 cells were transfected with the
Lenti-HOTAIR and Lenti-HOTAIR si, NC and Lenti-NC as a control to
detect the amounts of H3K27me3/ac in the nuclei, and images were
captured using a microscope (Olympus IX81; Olympus Corporation).
The antibodies used were as follows: H3K27me3 (ab6002; 5
µg/ml), H3K27ac (ab45173; 5 µg/ml) (both from Abcam)
and E-cadherin (14472; Cell Signaling Technology; 5 µg/ml).
DAPI was used to dye the nuclei into blue.
Proliferation assay
The transfected and control cells were plated in
96-well plates at a cell density of 104 cells/well with
6 replicate wells. The cells were incubated at 37°C for an
additional 4 h by adding 10 µl of CCK-8 (5 g/l; Sigma, St.
Louis, MO, USA) into each well. The optical density (OD) was
measured at 450 nm at 0, 24, 48, 72, 96 and 120 h, and the data
were expressed as a percentage of the control, which are derived
from triplicate samples.
Cell invasion analysis
Transwell invasion assays were performed using
Matrigel-coated Transwell membranes (BD Biosciences, San Jose, CA,
USA). The Matrigel solidified after a 30-min incubation at 37°C and
post-transfection, the cells (105) in 100 µl of
serum-free RPMI-1640 medium were added to the upper compartment of
the chamber. As a chemoattractant, 20% FBS in RPMI-1640 medium was
added to the lower chambers of the wells. The cells were then
incubated at 37°C in 5% CO2 for 48 h, and the lower
cells were stained with crystal violet (Zhongshan Bio Corporation)
for 5 min at room temperature. The average number of cells per
field was calculated by counting cells in 6 random fields per well
under the microscope at ×200 magnification (Olympus
Corporation).
Chromatin immunoprecipitation (ChIP)
To determine whether H3K27me3/ac binds to the
E-cadherin promoter, ChIP was performed using the EZ-ChIP kit
(Millipore, Billerica, MA, USA) according to the manufacturer’s
instructions. Cross-linked chromatin was extracted from the gastric
cancer cells by adding ChIP lysis buffer (1% SDS, 10 mM EDTA, 50 mM
Tris-HCl pH 8.0) and sonicated into 200-1,000 fragments. The
chromatin was immunoprecipitated by the addition of anti-H3K27ac
(ab45173, 1 µg/ml) and anti-H3K27me3 (ab6002, 1
µg/ml) antibodies, and a negative control IgG (ab171870,
1 µg/ml) (both from Abcam), which was incubated overnight at
4°C with Protein G agarose. The Protein G
agarose-antibody/chromatin complex was resuspended in wash buffer
and centrifuged (70 × g, 3 min) to collect the protein/DNA complex,
which were reversed to obtain free DNA. qPCR was conducted
according to the method described above. Input sample represents
the amount of chromatin used in the ChIP, the results of each
antibody sample represent the percent of Input. The primers used
were as follows: CDH1a forward, 5′-CCCTATGACTGCACCCCTTG-3′ and
reverse, 5′-GCAGATAACAGACCCCGCAT-3′; CDH1b forward,
5′-CAAATGGCCCTGGGAGGTTC-3′ and reverse,
5′AGCCCTCTTGGCCCAATACA-3′.
The ENCODE Histone Modification Tracks embedded in
the UCSC Genome Browser (University of California Santa Cruz, Santa
Cruz, CA, USA) was used to find the promoter.
Animal experiments
A total of 12 BALB/c-A nude female mice at 6 weeks
of age and weighing 8 g were purchased from the Animal Center of
the Cancer Institute, Chinese Academy of Medical Science and bred
at Tianjin Institute of Biological Engineering. The maintenance
conditions for the mice were as follows: Temperature, 26-28°C;
humidity, 40-60%; the food and water were bacteria-free without any
special requirements. The mice were randomly assigned to 2 groups
(6 mice a group), and were separately subcutaneously injected with
107 MGC-803 cells transfected with Lenti-NC and
Lenti-HOTAIR si in 100 µl of PBS. The tumor volumes and the
weight of the mice were measured every 2 days, and the tumor weight
was surveyed at the endpoint of the study. At 25 days post-tumor
implantation, the xenograft samples were collected and subjected to
immunohistochemistry (IHC). Animal experiments were approved by the
Ethics Committee of Tianjin Medical University General
Hospital.
IHC
For staining, 5-µm-thick sections were cut
and dried, deparaffinized, and immunostaining was performed on the
sections using the avidin-biotin-complex method. Primary antibodies
specific for H3K27me3 (ab6002), H3K27ac (ab45173) (Abcam; 1:100
dilution) and E-cadherin (14472) (Cell Signaling Technology; 1:50
dilution) were diluted in PBS with 0.1% Tween-20 and incubated
overnight at 4°C. A secondary antibody (PV9003; Zhongshan Bio
Corp.; 1:100 dilution) was added to the slides at 37°C for 1 h,
followed by avidin-biotin complex solution for an additional 1 h.
Protein expression was detected by coloration with diaminobenzidine
(DAB) buffer for 5 min, and the sections were counterstained with
hematoxylin for 5 min at room temperature. All images were captured
using a microscope (Olympus Corporation).
Statistical analysis
All statistical analyses were performed using SPSS
version 16.0 (SPSS, Inc., Chicago, IL, USA) or GraphPad 5.0
(GraphPad Software, Inc., La Jolla, CA, USA) software. Data are
expressed as the means ± standard deviation; the experiments were
repeated 3 times. The statistical significance of differences
between groups was determined using a Student’s t-test, and one-way
ANOVA was used to determine the differences among at least 2
groups. P-values <0.05 were considered to indicate statistically
significant differences.
Results
HOTAIR knockdown inhibits the malignant
properties of GC cells
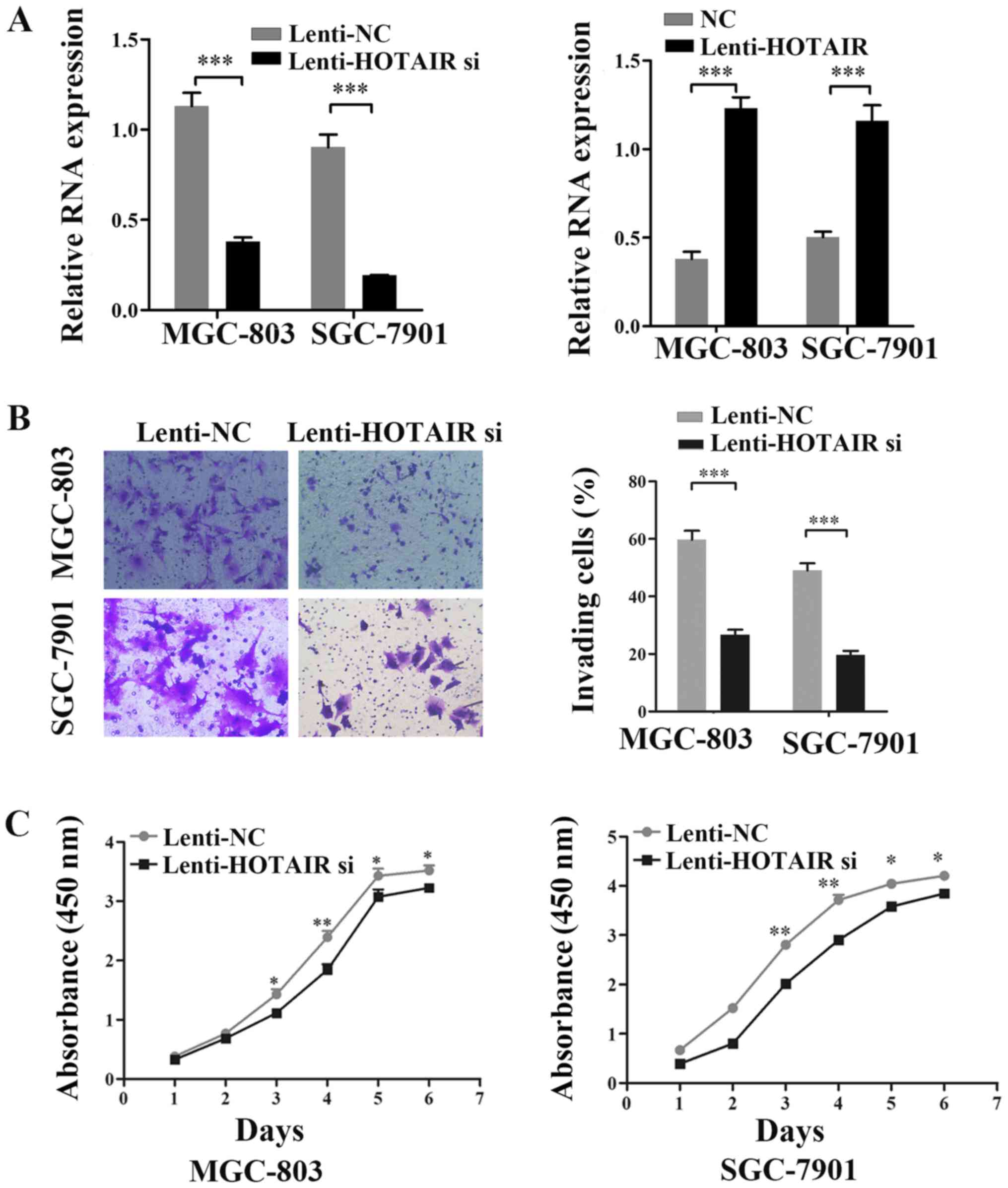
Based previous results regarding HOTAIR (23), in this study, we examined the
effect of HOTAIR on the malignant properties of GC cells. HOTAIR
expression was significantly knocked down or overexpressed
following lentiviral infection detected by qPCR (Fig. 1A). The results of Transwell assays
revealed that the inhibition of HOTAIR decreased MGC-803 and
SGC-7901 cell invasion (Fig. 1B).
In addition, the CCK-8 assay was used to observe the proliferation
of MGC-803 and SGC-7901 cells following HOTAIR knockdown in
vitro. We estimated the proliferation rate of the GC cells at
0, 24, 48, 72, 96 and 120 h and compared this with that of the
negative control. The results indicated that HOTAIR knockdown
suppressed the growth of the GC cells (Fig. 1C). Taken together, these results
indicate that HOTAIR is associated with the malignant properties of
GC cells.
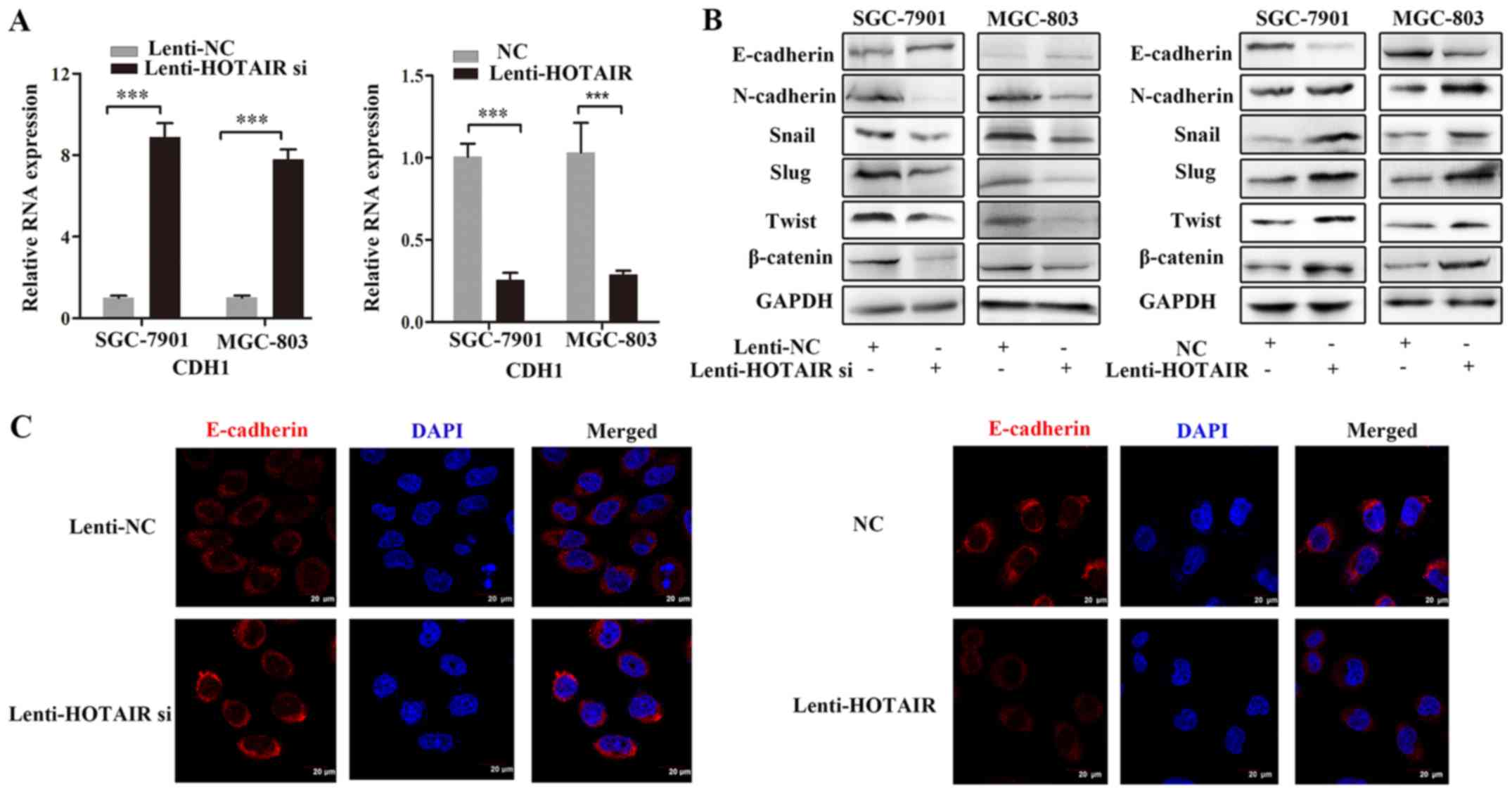
HOTAIR negatively regulates E-cadherin
expression
Since the malignant properties of cancer cells are
associated with EMT (2), we
examined the effects of HOTAIR on EMT processes in GC cells.
Previous research revealed that HOTAIR is involved in GC cell ECM
function (16). Consequently, we
detected the of mRNA and protein expression levels of EMT-related
TFs following HOTAIR knockdown or overexpression. The expression
levels of the epithelial marker, E-cadherin (also known as CDH1),
in cells in which HOTAIR was knocked down were increased compared
with the control cells. By contrast, the expression levels of
mesenchymal markers, including N-cadherin, Snail, Slug, Twist and
β-catenin, were decreased in the GC cells in which HOTAIR was
knocked down. However, in the HOTAIR-overexpressing cells, the
expression levels of EMT markers exhibited the opposite effects
(Fig. 2A and B). We then performed
immunofluorescence staining to evaluate E-cadherin-related
morphological changes induced by HOTAIR knockdown or
overexpression. As shown in Fig.
2C, the Lenti-HOTAIR si-infected cells exhibited an epithelial
morphology, which was characterized by the increased expression of
E-cadherin. By contrast, the cells infected with Lenti-HOTAIR
exhibited a mesenchymal morphology, as indicated by the decreased
expression of E-cadherin in the cytoplasm. The E-cadherin protein
is a hallmark of EMT and is crucial for maintaining an epithelial
phenotype (26). These results
suggested that HOTAIR may promote GC cell metastasis by negatively
regulating E-cadherin expression at the mRNA and protein level.
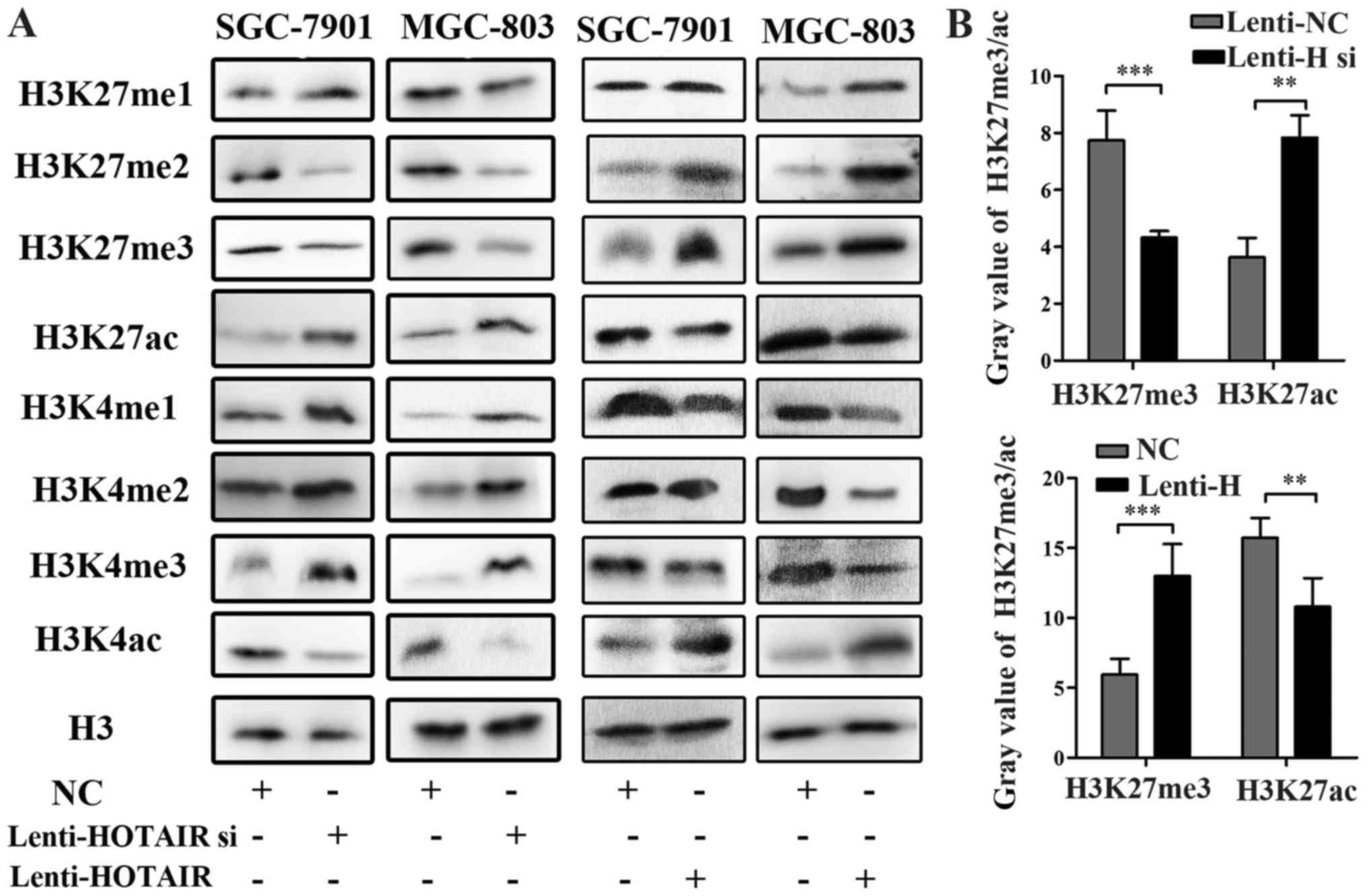
HOTAIR regulates histone methylation and
the acetylation of H3K4 and H3K27
Previous research has indicated that long intergenic
non-coding RNA HOTAIR serves as a scaffold for linking a histone
methylase and a demethylase, which separately affects histone H3
lysine 27 methylation and lysine 4 demethylation (12). It has also been reported that the
loss of H3K27me3 results in increased levels of H3K27ac, which are
specifically dependent on PRC2 activity (27). It is believed that the methylation
of H3K27 mediates transcriptional repression. However, the
methylation and acetylation of H3K4, and the acetylation of H3K27
are regarded as transcriptionally active markers (28). Therefore, in this study, we
performed western blot analysis to explore the molecular mechanisms
through which HOTAIR regulates transcription in SGC-7901 and
MGC-803 GC cells. The expression levels of H3K4me1/2/3 and H3K27ac
were increased, whereas those of H3K4ac and H3K27me1/2/3 were
decreased following infection with Lenti-HOTAIR si. By contrast,
infection with Lenti-HOTAIR increased the H3K4ac and H3K27me1/2/3
expression levels, and the decreased H3K4me1/2/3 and H3K27ac
expression levels (Fig. 3A). The
results of western blot analysis indicated that HOTAIR regulated
the methylation and acetylation of H3K4/27 and mediated the
transcriptional repression or activation. Furthermore, we analyzed
the gray value of H3K27me3/ac. The results indicated that HOTAIR
may regulate histone H3K27 methylation and acetylation using an
antagonistic approach (Fig.
3B).
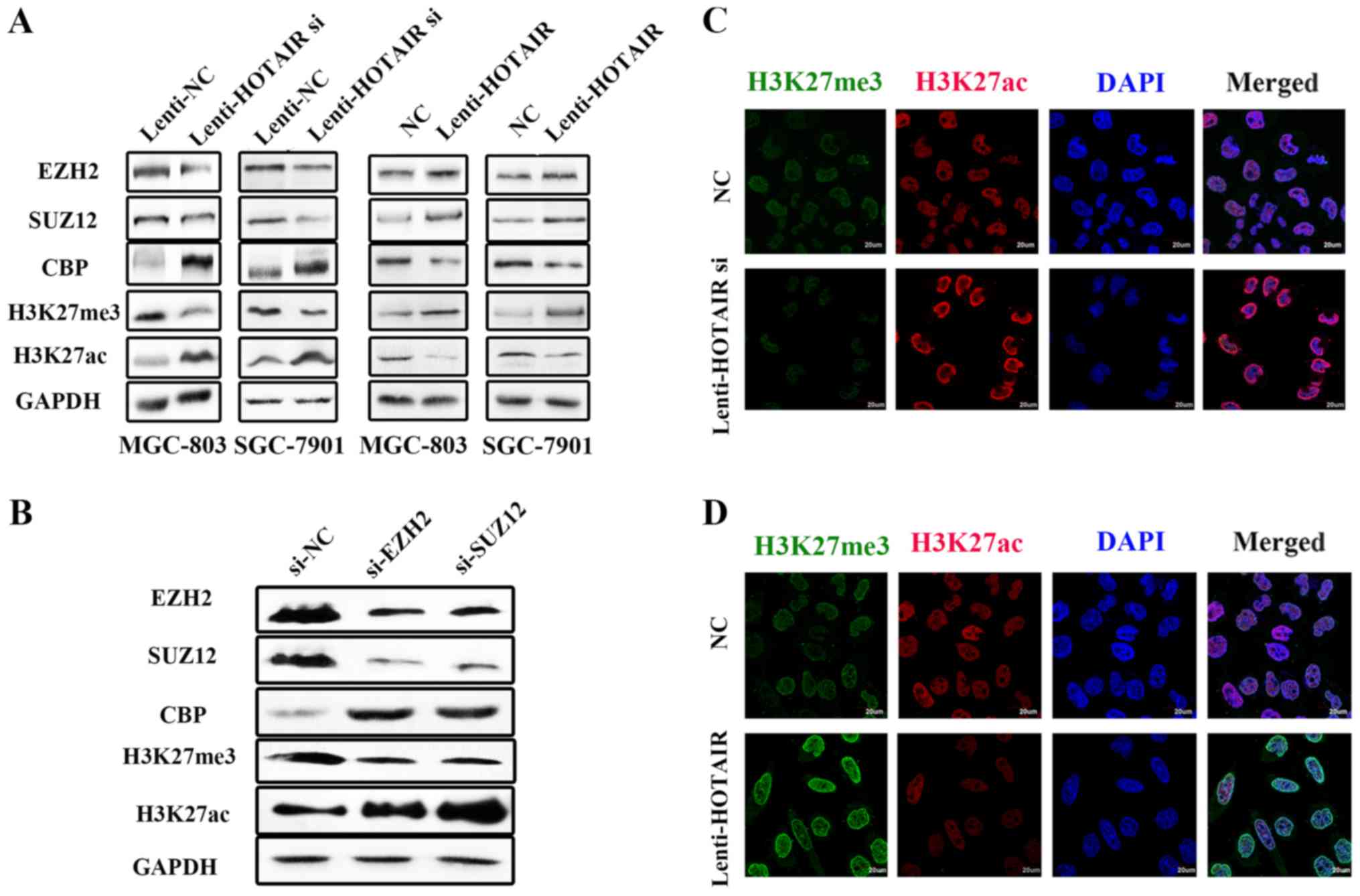
HOTAIR is associated with an antagonistic
switch between histone H3K27 methylation and acetylation
The above-mentioned findings demonstrated that H3K27
methylation and acetylation were regulated by HOTAIR in opposing
ways. Subsequently, we aimed to identify the histone-modifying
enzymes that may be involved in HOTAIR-associated H3K27 methylation
and acetylation. It is known that HOTAIR targets PRC2, which
consists of H3K27 methyltransferase EZH2, SUZ12 and EED (29). In this study, the GC cell lines,
SGC-7901 and MGC-803, were infected with Lenti-HOTAIR si or
Lenti-HOTAIR. Western blot analysis was then used to examine the
expression levels of EZH2 and SUZ12 and the results revealed that
these were not markedly altered by HOTAIR knockdown or
overexpression. However, the expression levels of acetyltransferase
CBP increased with Lenti-HOTAIR si infection and decreased with the
overexpression of HOTAIR. Additionally, the expression levels of
H3K27me3 were decreased, whereas the expression levels of H3K27ac
were increased by Lenti-HOTAIR si infection. The opposite effects
were observed as regards the expression levels of these factors
with HOTAIR overexpression (Fig.
4A). To further explore the association between the PRC2
complex and CBP, the expression levels of EZH2, SUZ12, CBP,
H3K27me3 and H3K27ac were detected following the inhibition of EZH2
and SUZ12 expression. The results were similar to those observed
with Lenti-HOTAIR si infection (Fig.
4B). Following this, immunofluorescence assays were performed
to further verify the HOTAIR-mediated antagonistic switch between
histone H3K27 methylation and acetylation in GC cells (Fig. 4C and D). All the results mentioned
above indicated that the antagonistic switch of methylation and
acetylation of H3K27 were specifically dependent on PRC2 activity,
which was induced by HOTAIR. Conversely, acetyltransferase CBP was
considered to play an important role in histone H3K27ac without the
existence of the PRC2 complex.
Epigenetic regulation of E-cadherin
expression by the methylation and acetylation of H3K27
To examine the underlying mechanisms through which
E-cadherin transcription is suppressed by HOTAIR-mediated H3K27
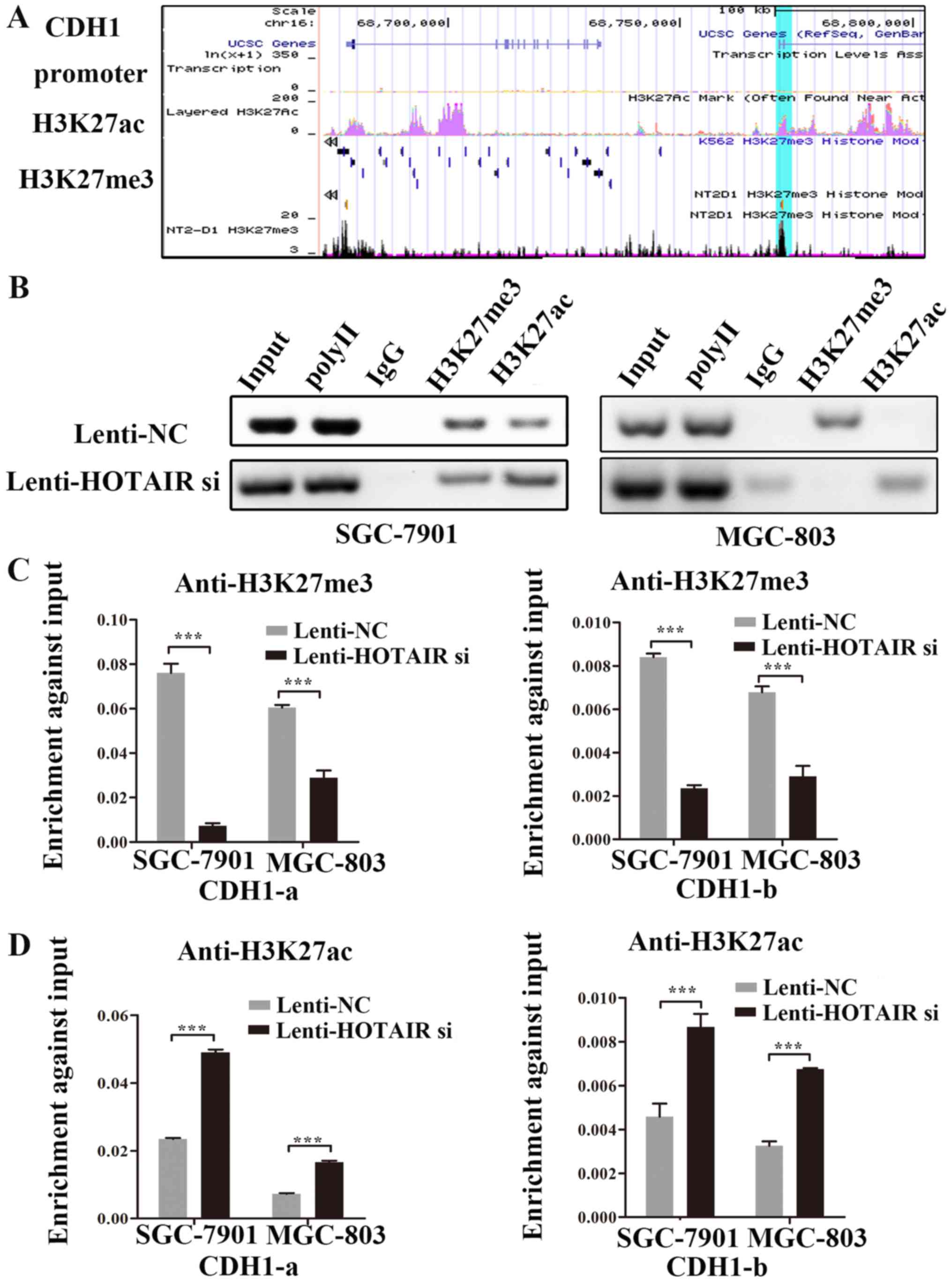
methylation and acetylation, UCSC Genome Browser was used to
identify H3K27me3 and H3K27ac enrichment peaks in the E-cadherin
promoter region (Fig. 5A). ChIP
was performed to obtain specific DNA, including the DNA of the CDH1
promoter binding to H3K27me3 and H3K27ac using agarose gel
electrophoresis (Fig. 5B). The
results suggested that H3K27me3 and H3K27ac were enriched in the
promoter region of the CDH1 gene. In addition, qPCR analysis
revealed that HOTAIR knockdown decreased the binding of H3K27me3 to
the E-cadherin promoter in GC cells (Fig. 5C); however, HOTAIR knockdown
induced the recruitment of H3K27ac to the E-cadherin binding site
at the promoter of E-cadherin (Fig.
5D). Therefore, the knockdown of HOTAIR appears to relieve the
repressive histone marker, H3K27me3, and induces a switch to the
active marker, acetylated H3K27, at the promoter of E-cadherin.
Taken together, these data provide evidence of a HOTAIR-mediated
competition between H3K27me3 and H3K27ac in regulating CDH1
expression.
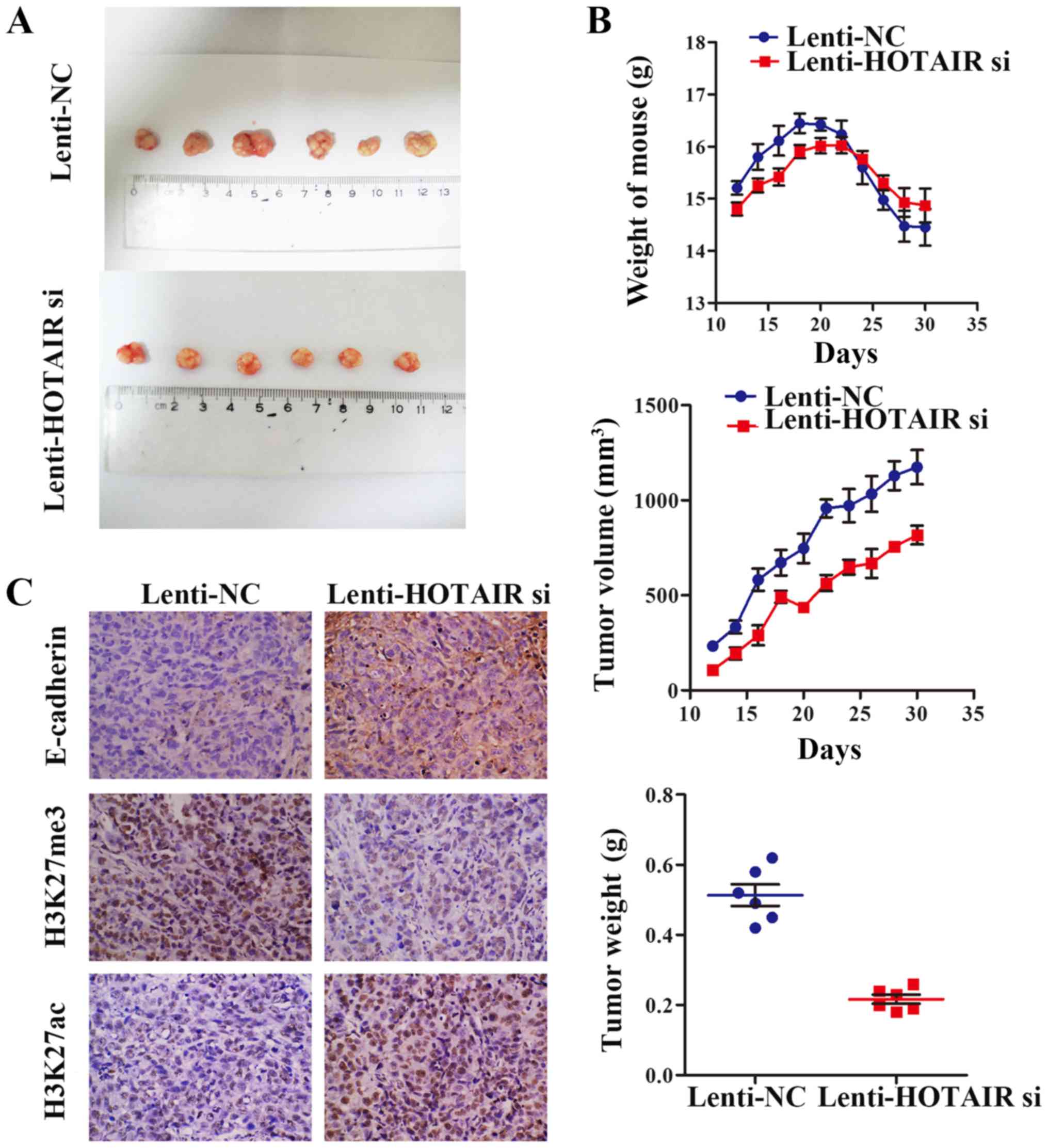
HOTAIR knockdown inhibits GC growth in
vivo
To further verify the role of HOTAIR in GC cells and
to determine the therapeutic potential of HOTAIR knockdown, we
established a subcutaneous tumor model of GC using MGC-803 GC
cells. Nude mice were subcutaneously injected with MGC-80 cells
that were infected with Lenti-NC or Lenti-HOTAIR si. From 10 days
post-injection, the weight of the mice and tumor volume were
measured every 2 days for a total of 25 days. The tumor weight was
measured after the mice were sacrificed (Fig. 6A and B). Compared with the nude
mice injected with Lenti-NC-infected MGC-803 cells, the mice
bearing tumors derived from MGC-803 cells in which HOTAIR was
knocked down exhibited suppressed GC tumor growth. In addition, IHC
was performed to evaluate the pathological changes in each of the
GC orthotopic tumors (Fig. 6C).
Compared with the Lenti-NC tumors, treatment with Lenti-HOTAIR si
significantly elevated E-cadherin expression. Furthermore, the
methylation of H3K27 was decreased and the acetylation of H3K27 was
increased following infection with Lenti-HOTAIR si, which was in
line with our results in vitro.
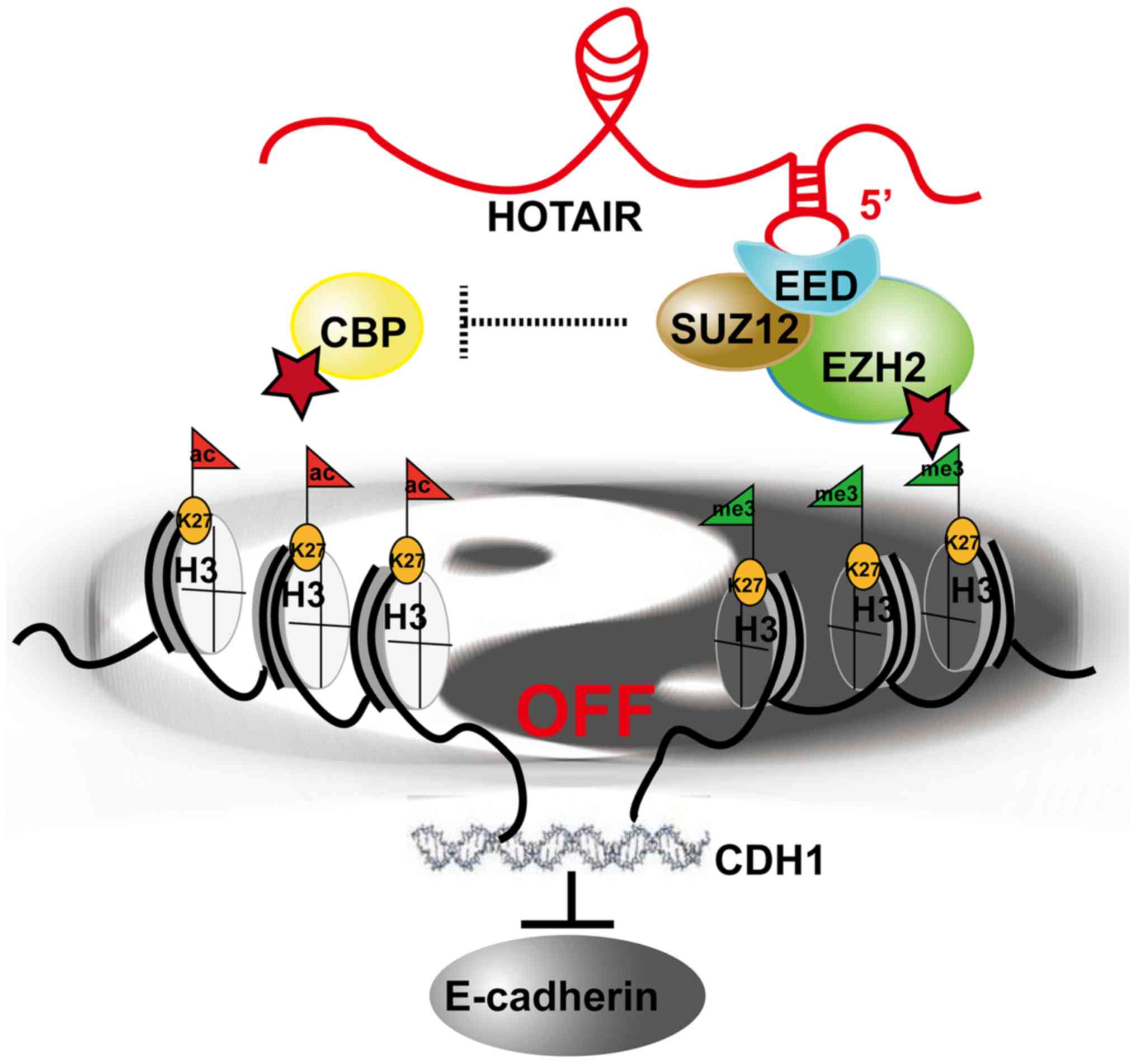
Taken together, the findings of this study indicate
that HOTAIR promotes the EMT of gastric cancer by inhibiting the
expression of E-cadherin through an antagonistic switch of histone
H3K27 acetylation to methylation at the E-cadherin promoter
(Fig. 7).
Discussion
Since the introduction of HOTAIR by Rinn et
al (7) in 2007, numerous
studies have been conducted regarding HOTAIR and the associated
mechanisms of carcinogenesis and tumor progression. It has been
reported that overexpression of long non-coding RNA HOTAIR is
associated with various types of cancer (30-33),
which suggests that HOTAIR functions as an oncogene in a variety of
human cancer types. Previous studies have revealed that, compared
with adjacent non-tumor tissues, HOTAIR is upregulated in GC tumor
tissues and is also associated with diffuse type GC, venous
invasion and other clinicopathological factors, including TNM stage
and lymph node metastasis (11,34,35).
In this study, as shown in Fig. 1,
we performed Transwell assays to detect the invasion and metastasis
of GC cells. Cell proliferation was also evaluated by CCK-8 assay.
Notably, the results in vivo had the same trend as those
in vitro. All the results indicated that the overexpression
of HOTAIR promoted the development of GC. These findings suggest
that HOTAIR plays an important role in the aggressiveness and
metastasis of GC, and may serve as a novel biomarker for the
metastasis or prognosis of GC.
HOTAIR serves as a modular scaffold of histone
modification complexes, with the 5′ domain binding the PRC2 and the
3′ domain binding the LSD1/CoREST/REST complex (12). The methyltransferase EZH2 is a
catalytic subunit of PRC2 that can epigenetically silence
downstream gene transcription by the addition of three methyl
groups to lysine 27 of histone 3 (36). However, the demethylase LSD1
performs the enzymatic demethylation of H3K4me2. Despite the
influence of the HOTAIR-mediated methylation of histone 3, lysine
27 and lysine 4, H3K4/27 acetylation is significantly altered in an
opposite manner compared with methylation. In this study, the
results of western blot analysis in Fig. 3 demonstrated that the knockdown of
HOTAIR decreased H3K27 methylation and increased H3K27 acetylation,
whereas H3K4 methylation was increased and the acetylation of H3K4
was decreased. Therefore, we attempted to explore the underlying
mechanisms of HOTAIR and histone acetylation. Previous research has
indicated that histone H3 lysine 27 methylation and acetylation act
as an antagonistic switch in regulating gene expression. In
addition, the loss of SUZ12, a subunit of PRC2, results in a
significant increase of global H3K27ac, which requires the activity
of acetyltransferase CBP (37).
Accordingly, in this study, we performed western blot analysis to
examine the levels of EZH2, SUZ12, CBP, H3K27me3 and H3K27ac
following infection with Lenti-HOTAIR si, Lenti-HOTAIR, si-EZH2 and
si-SUZ12. The results in shown in Fig.
4 indicated a potential mechanism with regard to the switch
between H3K27 methylation and acetylation. HOTAIR induced the
methylation of H3K27 by recruiting EZH2, and the acetylation of
H3K27 was decreased in the presence of SUZ12, which may inhibit the
binding of CBP and H3K27ac. Thus, HOTAIR mediates a switch between
the acetylation and tri-methylation of H3K27, which are associated
with the transcriptional activation and repression of target genes,
respectively.
EMT is associated with multiple cancer cells
(38). During the metastasis and
invasion of GC cells, the cells lose the function of epithelial
factors, such as E-cadherin, and increase the expression of
mesenchymal markers, including Twist, Snail, Slug, N-cadherin and
β-catenin (39). E-cadherin is
considered an important epithelial marker and its loss of function
may contribute to cancer progression by increasing proliferation,
invasion and/or metastasis. Multiple studies have reported that
HOTAIR is negatively associated with the expression of E-cadherin
in cancer cells (30,31,40).
Therefore, we performed experiments to detect the levels of CDH1
and E-cadherin in GC cells. The results indicated that E-cadherin
may be the pivotal marker that promotes HOTAIR-mediated GC cell
migration and metastasis. Recent evidence has linked EMT with
epigenetic modifications (41-43),
particularly histone modifications. It has been proposed that the
E-cadherin promoter, EZH2, and H3K27me3 have binding sites that can
result in the silencing of E-cadherin. As a result, we performed
ChIP assay in the present study. The results indicated that HOTAIR
increased the binding of H3K27me3 instead of H3K27ac at the
E-cadherin promoter, which causes a switch from the transcriptional
activation to the transcriptional repression of E-cadherin.
In this study, we validated that HOTAIR promoted EMT
by epigenetically silencing E-cadherin expression in GC. Long
non-coding RNA HOTAIR and histone modification are critical factors
during carcinogenesis and tumor progression. In addition to the
previously mentioned mechanisms of HOTAIR and EMT, we propose a new
regulation of EMT by HOTAIR through an competitive switch of
histone H3K27 acetylation and methylation at the E-cadherin
promoter, which differs from the traditional model of HOTAIR
concerning PRC2 and H3K27me3. This finding provides new insight
into HOTAIR and epigenetic modification. However, the exact
mechanism of PRC2, CBP and H3K27ac requires further exploration.
Additionally, the association between HOTAIR and other EMT-TFs,
particularly mesothelial factors, is not explained by the
HOTAIR-mediated switch of histone acetylation and methylation and
therefore requires further exploration.
In conclusion, the findings of the present study
indicate that HOTAIR promotes EMT in GC by switching between the
acetylation and the tri-methylation of H3K27, which is associated
with the change of transcriptional activation to inhibition of
E-cadherin. Taken together, these findings indicate a novel
mechanism involving HOTAIR, histone modification and EMT (Fig. 7).
Acknowledgments
The authors would like to thank the members of the
Laboratory of Neuro-Oncology, Tianjin Neurological Institute for
their technical assistance.
Funding
This study was supported by funds from the China
National Natural Scientific Found (nos. 81372703 and 81172356).
Availability of data and materials
All data generated or analyzed during this study are
included in this published article or are available from the
corresponding author on reasonable request.
Authors’ contributions
YS and RW performed the molecular studies. LWL and
XL performed the animal experiments. YFW and QXW provided
experimental technical support and performed the statistical
analysis. QYZ designed the study and helped to draft the
manuscript. All authors read and approved the final manuscript.
Ethics approval and consent to
participate
Animal experiments were approved by the Ethics
Committee of Tianjin Medical University General Hospital.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
De Craene B and Berx G: Regulatory
networks defining EMT during cancer initiation and progression. Nat
Rev Cancer. 13:97–110. 2013. View
Article : Google Scholar : PubMed/NCBI
|
|
2
|
Thiery JP, Acloque H, Huang RY and Nieto
MA: Epithelial-mesenchymal transitions in development and disease.
Cell. 139:871–890. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Huang L, Wu RL and Xu AM:
Epithelial-mesenchymal transition in gastric cancer. Am J Transl
Res. 7:2141–2158. 2015.
|
|
4
|
Yan Y, Zhang J, Li JH, Liu X, Wang JZ, Qu
HY, Wang JS and Duan XY: High tumor-associated macrophages
infiltration is associated with poor prognosis and may contribute
to the phenomenon of epithelial-mesenchymal transition in gastric
cancer. Onco Targets Ther. 9:3975–3983. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Peng Z, Wang CX, Fang EH, Wang GB and Tong
Q: Role of epithelial-mesenchymal transition in gastric cancer
initiation and progression. World J Gastroenterol. 20:5403–5410.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Bedi U, Mishra VK, Wasilewski D, Scheel C
and Johnsen SA: Epigenetic plasticity: A central regulator of
epithelial-to-mesenchymal transition in cancer. Oncotarget.
5:2016–2029. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Rinn JL, Kertesz M, Wang JK, Squazzo SL,
Xu X, Brugmann SA, Goodnough LH, Helms JA, Farnham PJ, Segal E, et
al: Functional demarcation of active and silent chromatin domains
in human HOX loci by noncoding RNAs. Cell. 129:1311–1323. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Marchese FP and Huarte M: Long non-coding
RNAs and chromatin modifiers: Their place in the epigenetic code.
Epigenetics. 9:21–26. 2014. View Article : Google Scholar :
|
|
9
|
Kogo R, Shimamura T, Mimori K, Kawahara K,
Imoto S, Sudo T, Tanaka F, Shibata K, Suzuki A, Komune S, et al:
Long noncoding RNA HOTAIR regulates polycomb-dependent chromatin
modification and is associated with poor prognosis in colorectal
cancers. Cancer Res. 71:6320–6326. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Amicone L, Citarella F and Cicchini C:
Epigenetic regulation in hepatocellular carcinoma requires long
noncoding RNAs. Biomed Res Int. 2015.1–10. 2015. View Article : Google Scholar
|
|
11
|
Hajjari M and Salavaty A: HOTAIR: An
oncogenic long non-coding RNA in different cancers. Cancer Biol
Med. 12:1–9. 2015.PubMed/NCBI
|
|
12
|
Zhou X, Ren Y, Zhang J, Zhang C, Zhang K,
Han L, Kong L, Wei J, Chen L, Yang J, et al: HOTAIR is a
therapeutic target in glioblastoma. Oncotarget. 6:8353–8365.
2015.PubMed/NCBI
|
|
13
|
Tsai MC, Manor O, Wan Y, Mosammaparast N,
Wang JK, Lan F, Shi Y, Segal E and Chang HY: Long noncoding RNA as
modular scaffold of histone modification complexes. Science.
329:689–693. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Baylin SB and Ohm JE: Epigenetic gene
silencing in cancer - a mechanism for early oncogenic pathway
addiction? Nat Rev Cancer. 6:107–116. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Ning X, Shi Z, Liu X, Zhang A, Han L,
Jiang K, Kang C and Zhang Q: DNMT1 and EZH2 mediated methylation
silences the microRNA-200b/a/429 gene and promotes tumor
progression. Cancer Lett. 359:198–205. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Liu YW, Sun M, Xia R, Zhang EB, Liu XH,
Zhang ZH, Xu TP, De W, Liu BR and Wang ZX: LincHOTAIR
epigenetically silences miR34a by binding to PRC2 to promote the
epithelial-to-mesenchymal transition in human gastric cancer. Cell
Death Dis. 6:e1802–e1811. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Margueron R and Reinberg D: Chromatin
structure and the inheritance of epigenetic information. Nat Rev
Genet. 11:285–296. 2010. View
Article : Google Scholar : PubMed/NCBI
|
|
18
|
Pradeepa MM, Grimes GR, Kumar Y, Olley G,
Taylor GC, Schneider R and Bickmore WA: Histone H3 globular domain
acetylation identifies a new class of enhancers. Nat Genet.
48:681–686. 2016. View
Article : Google Scholar : PubMed/NCBI
|
|
19
|
Greer EL and Shi Y: Histone methylation: A
dynamic mark in health, disease and inheritance. Nat Rev Genet.
13:343–357. 2012. View
Article : Google Scholar : PubMed/NCBI
|
|
20
|
Li KK, Luo C, Wang D, Jiang H and Zheng
YG: Chemical and biochemical approaches in the study of histone
methylation and demethylation. Med Res Rev. 32:815–867. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Kim YW and Kim A: Characterization of
histone H3K27 modifications in the β-globin locus. Biochem Biophys
Res Commun. 405:210–215. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Xu ZY, Yu QM, Du YA, Yang LT, Dong RZ,
Huang L, Yu PF and Cheng XD: Knockdown of long non-coding RNA
HOTAIR suppresses tumor invasion and reverses
epithelial-mesenchymal transition in gastric cancer. Int J Biol
Sci. 9:587–597. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Hajjari M, Behmanesh M, Sadeghizadeh M and
Zeinoddini M: Up-regulation of HOTAIR long non-coding RNA in human
gastric adenocarcinoma tissues. Med Oncol. 30:670–673. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Fang XY, Pan HF, Leng RX and Ye DQ: Long
noncoding RNAs: Novel insights into gastric cancer. Cancer Lett.
356:357–366. 2015. View Article : Google Scholar
|
|
25
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−Delta Delta C(T)) methods. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
26
|
Perl AK, Wilgenbus P and Dahl U: Acausal
role for E-cadherin in the transition from adenomato carcinoma.
Nature. 392:190–193. 1998. View
Article : Google Scholar : PubMed/NCBI
|
|
27
|
Pasini D, Malatesta M, Jung HR,
Walfridsson J, Willer A, Olsson L, Skotte J, Wutz A, Porse B,
Jensen ON, et al: Characterization of an antagonistic switch
between histone H3 lysine 27 methylation and acetylation in the
transcriptional regulation of Polycomb group target genes. Nucleic
Acids Res. 38:4958–4969. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Latham JA and Dent SY: Cross-regulation of
histone modifications. Nat Struct Mol Biol. 14:1017–1024. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Gupta RA, Shah N, Wang KC, Kim J, Horlings
HM, Wong DJ, Tsai MC, Hung T, Argani P, Rinn JL, et al: Long
non-coding RNA HOTAIR reprograms chromatin state to promote cancer
metastasis. Nature. 464:1071–1076. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Battistelli C, Cicchini C, Santangelo L,
Tramontano A, Grassi L, Gonzalez FJ, de Nonno V, Grassi G, Amicone
L and Tripodi M: The Snail repressor recruits EZH2 to specific
genomic sites through the enrollment of the lncRNA HOTAIR in
epithelial-to-mesenchymal transition. Oncogene. 10:1–14. 2016.
|
|
31
|
Liu L, Xu Z, Zhong L, Wang H, Jiang S,
Long Q, Xu J and Guo J: Enhancer of zeste homolog 2 (EZH2) promotes
tumour cell migration and invasion via epigenetic repression of
E-cadherin in renal cell carcinoma. BJU Int. 117:351–362. 2016.
View Article : Google Scholar
|
|
32
|
Wu Y, Zhang L, Zhang L, Wang Y, Li H, Ren
X, Wei F, Yu W, Liu T, Wang X, et al: Long non-coding RNA HOTAIR
promotes tumor cell invasion and metastasis by recruiting EZH2 and
repressing E-cadherin in oral squamous cell carcinoma. Int J Oncol.
46:2586–2594. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Zhang JX, Han L, Bao ZS, Wang YY, Chen LY,
Yan W, Yu SZ, Pu PY, Liu N, You YP, et al Chinese Glioma
Cooperative Group: HOTAIR, a cell cycle-associated long noncoding
RNA and a strong predictor of survival, is preferentially expressed
in classical and mesenchymal glioma. Neuro Oncol. 15:1595–1603.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Pan W, Liu L, Wei J, Ge Y, Zhang J, Chen
H, Zhou L, Yuan Q, Zhou C and Yang M: A functional lncRNA HOTAIR
genetic variant contributes to gastric cancer susceptibility. Mol
Carcinog. 55:90–96. 2016. View Article : Google Scholar
|
|
35
|
Zhao W, Dong S, Duan B, Chen P, Shi L, Gao
H and Qi H: HOTAIR is a predictive and prognostic biomarker for
patients with advanced gastric adenocarcinoma receiving
fluorouracil and platinum combination chemotherapy. Am J Transl
Res. 7:1295–1302. 2015.PubMed/NCBI
|
|
36
|
Chase A and Cross NC: Aberrations of EZH2
in cancer. Clin Cancer Res. 17:2613–2618. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Tie F, Banerjee R, Stratton CA,
Prasad-Sinha J, Stepanik V, Zlobin A, Diaz MO, Scacheri PC and
Harte PJ: CBP-mediated acetylation of histone H3 lysine 27
antagonizes Drosophila Polycomb silencing. Development.
136:3131–3141. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Christiansen JJ and Rajasekaran AK:
Reassessing epithelial to mesenchymal transition as a prerequisite
for carcinoma invasion and metastasis. Cancer Res. 66:8319–8326.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Yang J and Weinberg RA:
Epithelial-mesenchymal transition: At the crossroads of development
and tumor metastasis. Dev Cell. 14:818–829. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Zha L, Cao Q, Cui X, Li F, Liang H, Xue B
and Shi H: Epigenetic regulation of E-cadherin expression by the
histone demethylase UTX in colon cancer cells. Med Oncol. 33:21–31.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Wang Y and Shang Y: Epigenetic control of
epithelial-to-mesenchymal transition and cancer metastasis. Exp
Cell Res. 319:160–169. 2013. View Article : Google Scholar
|
|
42
|
Wang R, Chen X, Xu T, Xia R, Han L, Chen
W, De W and Shu Y: MiR-326 regulates cell proliferation and
migration in lung cancer by targeting phox2a and is regulated by
HOTAIR. Am J Cancer Res. 6:173–186. 2016.PubMed/NCBI
|
|
43
|
Wolff EM, Liang G and Jones PA: Mechanisms
of Disease: Genetic and epigenetic alterations that drive bladder
cancer. Nat Clin Pract Urol. 2:502–510. 2005. View Article : Google Scholar
|