Pancreatic cancer is a highly aggressive and fatal
malignancy, with stably high incidence and mortality for the past
30 years globally (1-4). Patients with pancreatic
adenocarcinoma account for ~85% of all patients with pancreatic
cancer and have a poor prognosis, compared with patients with other
common solid tumors (2,5-9).
Pancreatic cancer is primarily treated through surgical resection,
whereas postoperative patients and patients with advanced disease
are frequently subjected to adjuvant therapies with radiotherapy,
chemotherapy, and a combination of radiotherapy and chemotherapy
that are associated with high recurrence rates and poor treatment
effectiveness (10). The recent
development of and research into molecular targeted drugs have
great significance for the early diagnosis and prognosis of
malignant tumors, and have ushered a new era of tumor therapy
(11-15). However, the overall treatment
effect of these drugs remains poor, and further investigation is
required (16).
Autophagy is a process by which eukaryotic cells
undergo self-degradation of intracellular damaged macromolecular
proteins and organelles (17,18).
Autophagy at the physiological level is necessary to maintain the
stability of the internal environment, and the occurrence of a
variety of diseases, including inflammation, tumors and
degenerative diseases, is frequently accompanied by an abnormal
autophagic level (19-25). Currently, well-characterized,
autophagy-associated proteins include Beclin 1,
microtubule-associated-protein-1-light-chain-3 (LC3) and P62, and
well-studied pathways include mechanistic target of rapamycin
(mTOR) and phosphoinositide 3-kinase (PI3K) pathways (26,27).
mRNA is a single-stranded RNA that is transcribed
using one of the DNA chains as the template and carries the genetic
information that guides protein synthesis. Thus, mRNA occupies an
important position in the heredity dogma (39). mRNA serves a key role in various
human diseases and is involved in autophagy pathways (40,41);
for example, signal transducer and activator of transcription 3
could inhibit autophagy in pancreatic cancer cells (42) and ubiquitin specific peptidase 1
regulates autophagy by targeting Unc-51 like autophagy activating
kinase 1 (ULK1) (43). Non-coding
RNA (ncRNA) refers to RNA that does not encode a protein and
includes ribosomal RNA (rRNA), transfer RNA (tRNA), small nuclear
RNA (snRNA), long ncRNA (lncRNA), microRNA (miRNA) and circular RNA
(circRNA) (44-50). The involvement of lncRNAs and
miRNAs in cancer has been studied intensively, whereas the
potential significance of circRNAs in tumors has attracted
attention over the last two years (51). The post-transcriptional levels of
circRNAs serve a notable regulatory role in gene expression
(52). Furthermore, the sponge
effect of circRNAs on miRNAs is an important mechanism (53). circRNAs can contain multiple miRNA
binding sites, and attract miRNAs effectively to reduce
miRNA-mediated mRNA inhibition (54-56).
Based on the interaction between these three types of ncRNAs and
the current research status in pancreatic cancer, it was considered
that an investigation of the effect of lncRNAs and circRNAs as
competitive endogenous RNAs (ceRNAs) on the targeting of miRNAs in
the development of pancreatic cancer is beneficial.
Recently, the number of autophagy-associated lncRNAs
has been determined to be limited (57-60),
and investigations on circRNAs and their associations with
autophagy have not been reported to date. However, due to autophagy
being involved in the occurrence and development of a variety of
diseases, including breast (61,62),
gastric (63) and lung cancer
(64), in the present study, the
human pancreatic cancer cell line PANC-1 and gene chip technology
was used to detect differential expression of mRNAs, miRNAs,
lncRNAs and circRNAs under different autophagy levels, and to
investigate the genes associated with autophagy in pancreatic
cancer and their underlying molecular mechanisms.
The human pancreatic cancer cell line PANC-1 was
obtained from the Cell Laboratory of Chinese Academy of Sciences
(Shanghai, China) and cultured in complete Dulbecco’s modified
Eagle’s medium (DMEM; Gibco; Thermo Fisher Scientific, Inc.,
Waltham, MA, USA) at 37°C. Due to chloroquine diphosphate being
frequently used as a classic autophagic inhibitor (65,66),
the following concentration gradient of chloroquine diphosphate
(Sigma-Aldrich; Merck KGaA, Darmstadt, Germany) was designed as
recommended by the manufacturer’s protocols: 0, 25, 50, 75 and 100
µM. When the number of cells in the culture bottle grew to
~5×106, the original medium was replaced by a complete
DMEM containing chloroquine diphosphate. Following treating the
cells for 12 h at 37°C with the drug, the cellular proteins were
immediately extracted using Radioimmunoprecipitation Assay Lysis
Buffer (Beyotime Institute of Biotechnology, Haimen, China).
The lysis solution (radioimmunoprecipitation assay
buffer:phenylmethylsulfonyl fluoride; 100:1, Beyotime Institute of
Biotechnology) was prepared. The protein concentration in the
lysate was determined with a bicinchoninic acid kit (Beyotime
Institute of Biotechnology), according to the manufacturer’s
instructions. Western blotting was performed according to routine
procedures, and conducted with a 12% separating gel and a 5%
stacking gel. A total of 20 µg protein was added to the glue
holes. The electrophoresis condition was set to 120 Ma for 1 h.
Following the electrophoresis, the protein was transferred to the
polyvinylidene (PVDF) membrane (Beyotime Institute of
Biotechnology). Subsequently, the PVDF membrane was placed into the
Bull Serum Albumin blocking buffer (Beyotime Institute of
Biotechnology) for shaking at 37°C for 2 h. Primary antibodies were
incubated at 4°C overnight and secondary antibodies were incubated
at room temperature for 2 h. The primary antibodies were LC3
β-specific rabbit polyclonal (1:1,000; cat. no. 18725-1-AP) and P62
mouse monoclonal antibodies (1:2,000; cat. no. 66184-1-Ig), which
represented the state of autophagy, and the GAPDH horseradish
peroxidase-labeled internal reference antibody (1:10,000; cat. no.
HRP-60004), andhe secondary antibodies used were goat anti-mouse
IgG (H+L), allopregnanolone conjugate (1:1,000; cat. no. SA00002-1)
(all from ProteinTech Group, Inc., Chicago, IL, USA). The gray
scale values of the protein bands from the raw image were
determined using the ImageJ 1.48 (National Institutes of Health,
Bethesda, MD, USA) and the formula Final gray scale value (G) = [G
× (target band) − G × (background)] / [G × (internal reference band
of the group) − G × (background)]. The data were analyzed using
SPSS software 22 (SPSS, Inc., Chicago, IL, USA), and the results
were plotted using the GraphPad Prism 5 software (GraphPad
Software, Inc., La Jolla, CA, USA).
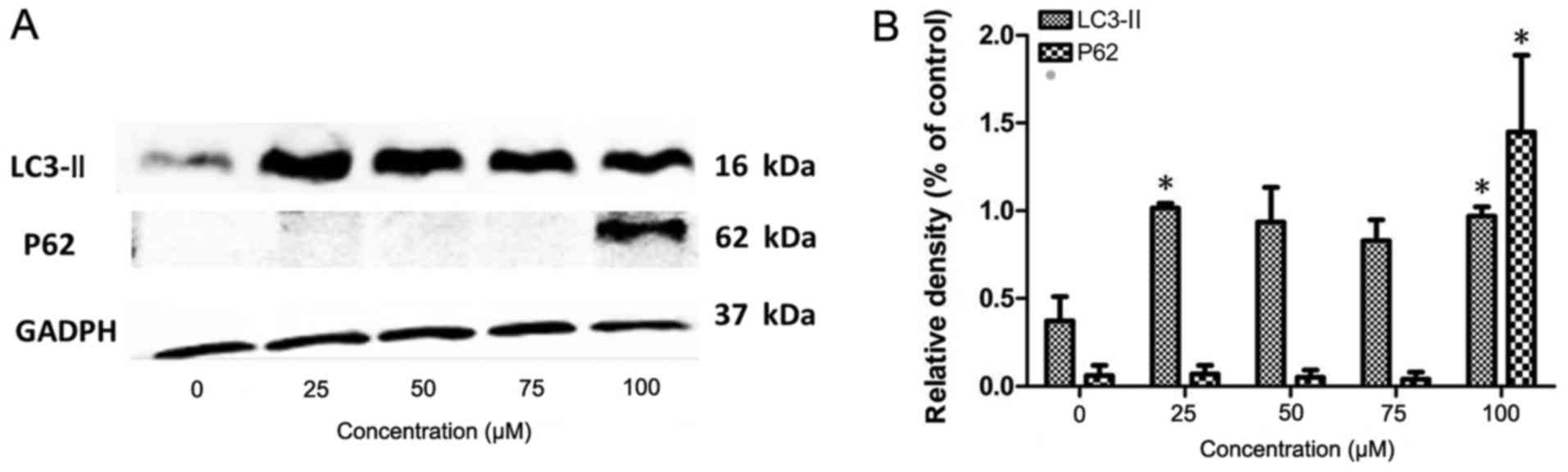
Based on the results of the western blotting
experiment, the human pancreatic cancer PANC-1 cells treated with
100 µM chloroquine diphosphate were selected as the
autophagic inhibition group, and the cells cultured in normal DMEM
medium were selected as the control group. Subsequent experiments
were compared between the autophagic inhibition group and the
control group. Following chloroquine diphosphate treatment, total
RNA was prepared using the TRIzol® reagent (Beyotime
Institute of Biotechnology) with three replicates per group.
The quality control for the gene analyses in the
present study was performed by Shanghai Biotechnology Corporation
(Shanghai, China). The starting sample of the microarray test was
total RNA, which was analyzed with the NanoDrop ND-2000
spectrophotometer and the Agilent Bioanalyzer 2100 (Agilent
Technologies, Inc., Santa Clara, CA, USA) to conduct the quality
control assay. Only RNA samples that passed the quality control
proceeded to the subsequent microarray experiments. The qualifying
sample standards were that the RNA integrity number of each sample
was ≥7.0 and that the 28S/18S was ≥0.7.
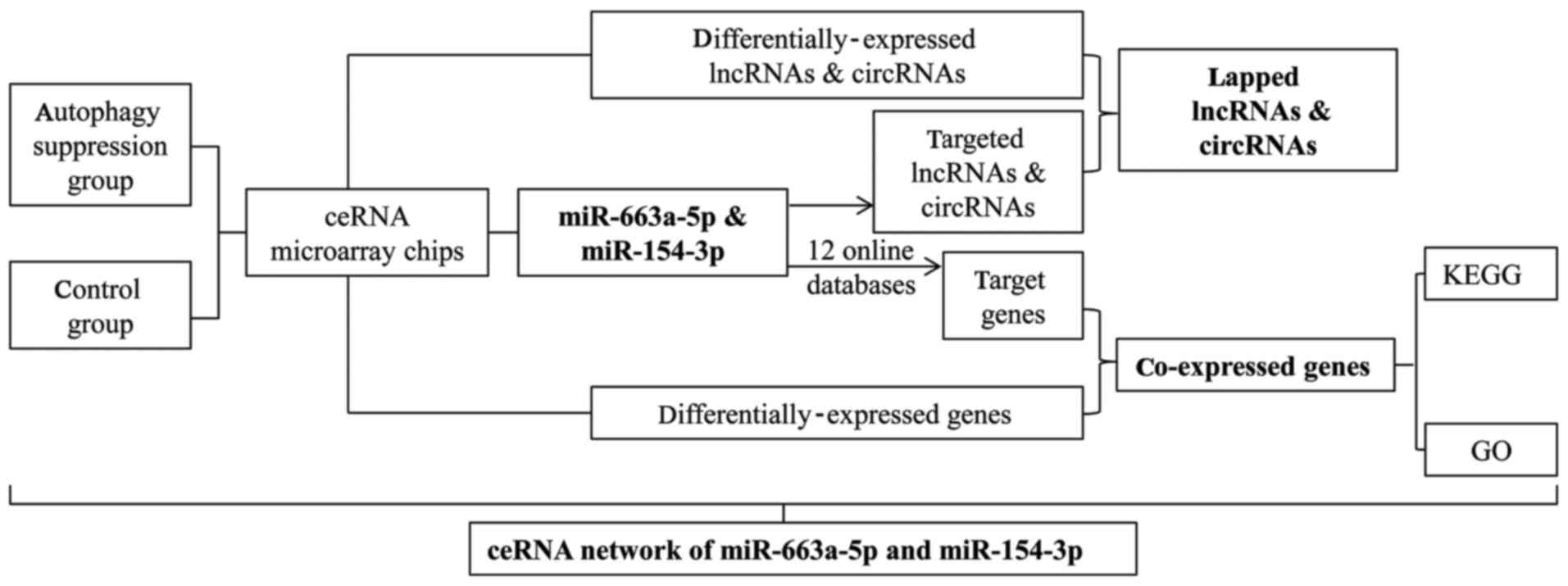
The ceRNA microarray detection assay included the
detection of three types of RNA (lncRNA, circRNA and mRNA). The
cutoff values of the differentials were all set to a fold change
(FC)>2 or FC<0.5 and P<0.05. The assays and data analyses
are described below. The analyses primarily included the
normalization of raw data, sample association analysis, screening
of genes with differential expression, Gene Ontology (GO)
enrichment analysis of the differentially-expressed mRNAs, Kyoto
Encyclopedia of Genes and Genomes (KEGG) pathway enrichment
analysis, PANTHER pathway analysis of the differentially-expressed
mRNAs, prediction of lncRNA and circRNA target genes, and
prediction of lncRNA and circRNA adsorption of miRNAs. GO and KEGG
pathway were generated by DAVID (https://david.ncifcrf.gov/) while PANTHER was analyzed
using the WebGestalt 2017 tool (http://www.webgestalt.org/option.php). The clinical
roles of genes were analyzed through the online database of Gene
Expression Profiling Interactive Analysis (GEPIA; http://gepia.cancer-pku.cn/) Which is based on the
data obtained from The Cancer Genome Atlas and GTEx. The miRNA
microarray analyses primarily included the normalization of raw
data, sample association analysis, screening of
differentially-expressed miRNAs and prediction of miRNA target
genes, which was predicted by 12 online miRNA target prediction
databases [DIANA-microTv4.0 (67),
DIANA-microT-CDS (68),
miRanda-rel2010 (69), mirbridge
(70), miRDB4.0 (71), miRMap (72), miRNAMap (73), PicTar2 (74), PITA (75), RNA22v2 (76), RNAhybrid2.1 (77) and Targetscan6.2 (78)] (http://zmf.umm.uni-heidelberg.de/apps/zmf/mirwalk2/).
The crossed genes from target prediction and
differentially-expressed mRNAs were depicted in Compendia
expression profiles (Novartis; http://software.broadinstitute.org/gsea/msigdb/annotate.jsp).
Network visualization was performed in Cytoscape 3.5.1 (The
Cytoscape Consoritum, New York, NY, USA).
The raw data of ceRNA microarray has been deposited
in the Gene Expression Omnibus (GEO) database (https://www.ncbi.nlm.nih.gov/geo/; accession
number GSE115517).
A total of 31 formalin-fixed paraffin-embedded
(FFPE) tissues, including 18 cases of Pancreatic ductal
adenocarcinoma and 13 paracancerous tissues, were collected from
the Department of Pathology of the First Affiliated Hospital of
Guangxi Medical University (Nanning, China) between June 2015 and
June 2018. The present study was approved by the Ethics Committee
of the First Affiliated Hospital of Guangxi Medical University. All
cases were from patients who have not been treated with
chemotherapy or radiation prior to resection and have signed
informed consent. There were 11 male and 7 female aged 29-77 years
(mean age, 56 years) in 18 patients with adenocarcinoma.
According to the user guides provided by the
manufacturer, the total RNA of tissues was extracted using an
E.Z.N.A.® FFPE RNA kit (Omega Bio-Tek, Inc., Norcross,
GA, USA). The complementary DNA (cDNA) was reverse transcribed
through the kit of miRNA First Strand cDNA Synthesis [Tailing
Reaction (79,80)] (Sangon Biotech Co., Ltd., Shanghai,
China), and RT-qPCR was conducted by applying a MicroRNAs qPCR kit
[SYBR® Green method (81,82)]
(Sangon Biotech Co., Ltd.), according to the manufacturer’s
protocols, on an ABI Prism 7500 (Applied Biosystems; Thermo Fisher
Scientific, Inc.). The temperature of pre-denaturation and
denaturation were both set at 95°C, and the temperature of
annealing/extension was set at 60°C. Pre-denaturation was conducted
for 10min, denaturation for 15 sec and annealing/extension for 60
sec. The number of cycles was set to 40. Subsequently, the
expression of miRNA relative to U6 was calculated via the
2−ΔΔCq method on identical samples (83). The PCR primers included: i)
miR-663a-5p for ward, 5′-ATAGGCGGGGCGCCGCGGGAC-3′; ii) miR-154-3p
forward, 5′-CCGGGAATCATACACGGTTGA CCTATT-3′; and iii) U6 forward
and universal PCR reverse primer were attached to the kits. Their
primer sequences are 5′-CTCGCTTCGGCAGCACA-3′ and 5′-AACGCTTCACGA
ATTTGCGT-3′, respectively.
Within GEO, ‘pancreas’ or ‘pancreatic’, and
‘adenocarcinoma’, ‘carcinoma’, ‘cancer’, ‘neoplasm’, ‘tumor’,
‘tumour’, ‘neoplas*’, ‘malignan*’, ‘PDAC’,
‘OR’, ‘PAAD’ or ‘PC’ were employed as a search strategy in GEO to
determine the expression of miR-663a-5p and miR-154-3p. STATA 12.0
(StataCorp LLC, College Station, TX, USA) was applied to estimate
the pooled effects, heterogeneity and publication bias.
The data of GEO and RT-qPCR were calculated using
SPSS 22 software for mean and standard deviation (SD). Continuous
data are presented as mean ± SD. The forest plots were produced by
Stata 12.0. The heterogeneity test was used to analyze the
existence of heterogeneity and the source of the occurrence of
heterogeneity (I2>50% or P<0.05 is the existence
of heterogeneity). Funnel plots were used for the analysis of
publication bias. The comparison between the two groups was
performed using two-sample Student’s t-test. The method for
differential gene expression analysis in GEPIA was one-way analysis
of variance, using pathological stage as variable for calculating
differential expression. GEPIA performs overall survival analysis
based on gene expression. GO analysis was performed in DAVID tool
which used Fisher’s exact test and the techniques of Kappa
statistics (84). PANTHER analysis
was performed in WebGestalt, which used Hypergeometric and Fisher’s
exact tests. P<0.05 was considered to indicate a statistically
significant difference.
Electrophoresis of the RNA samples indicated that
the RNA integrity number of each sample was ≥7.0 and that the
28S/18S was ≥0.7, which reached the qualifying sample standards.
Thus, the samples were qualified for use in the subsequent
experiments.
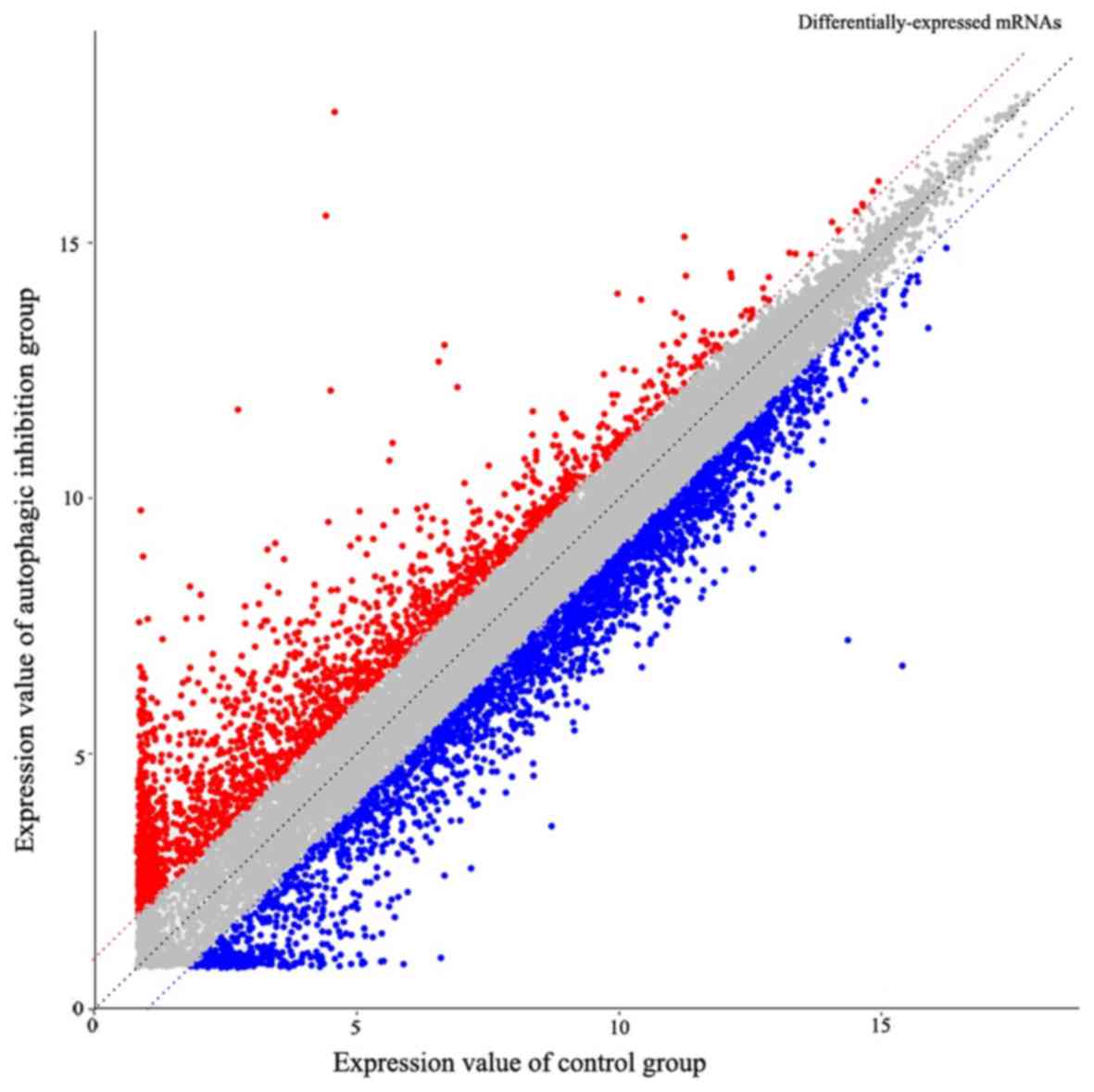
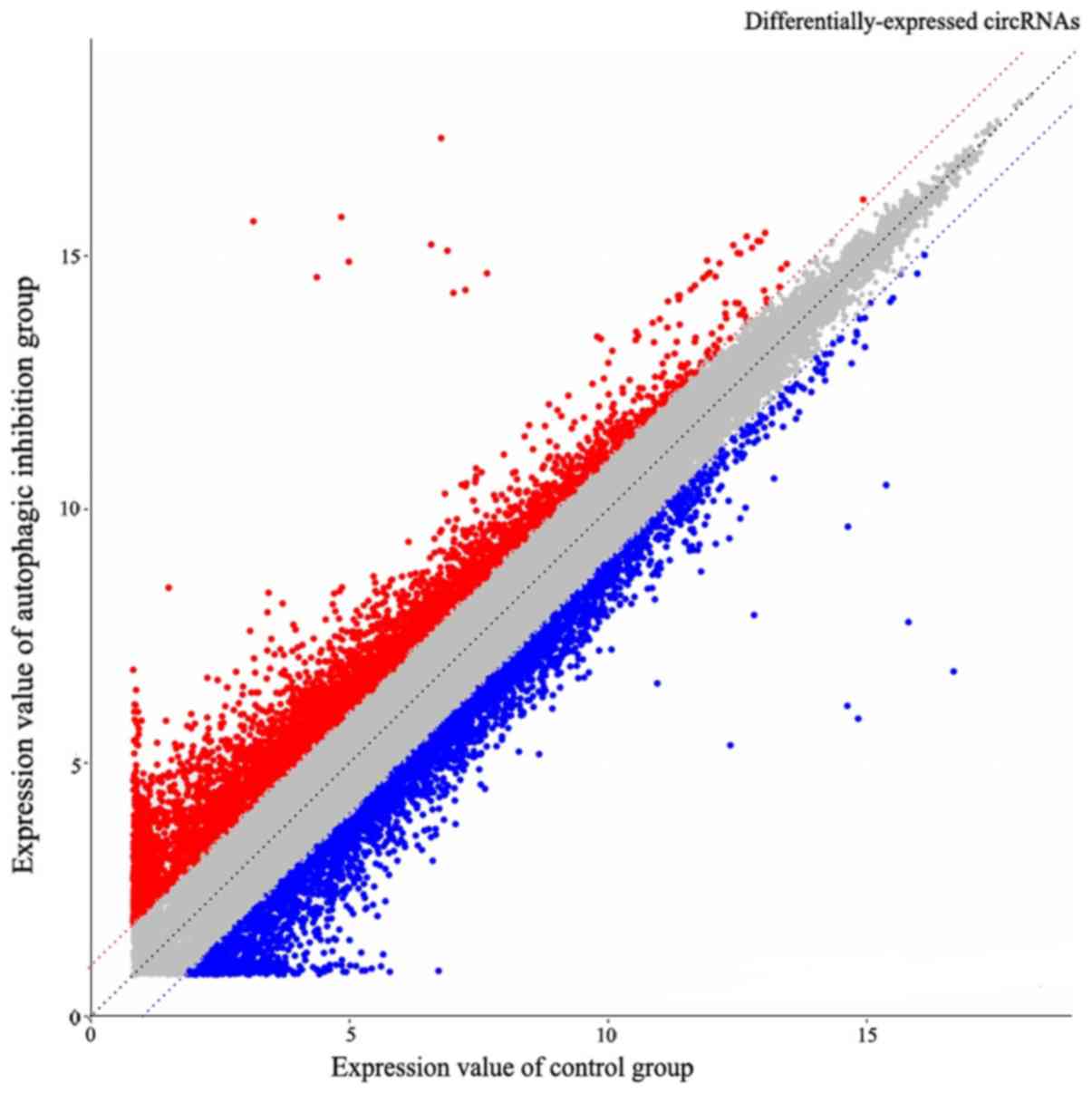
The ceRNA microarray results determined 3,966
differentially-expressed mRNAs in total. The expression levels of
2,445 mRNAs in the PANC-1 cells from the chloroquine diphosphate
treatment group (autophagy suppression group) were downregulated
and 1,521 mRNAs were upregulated, compared with the control group
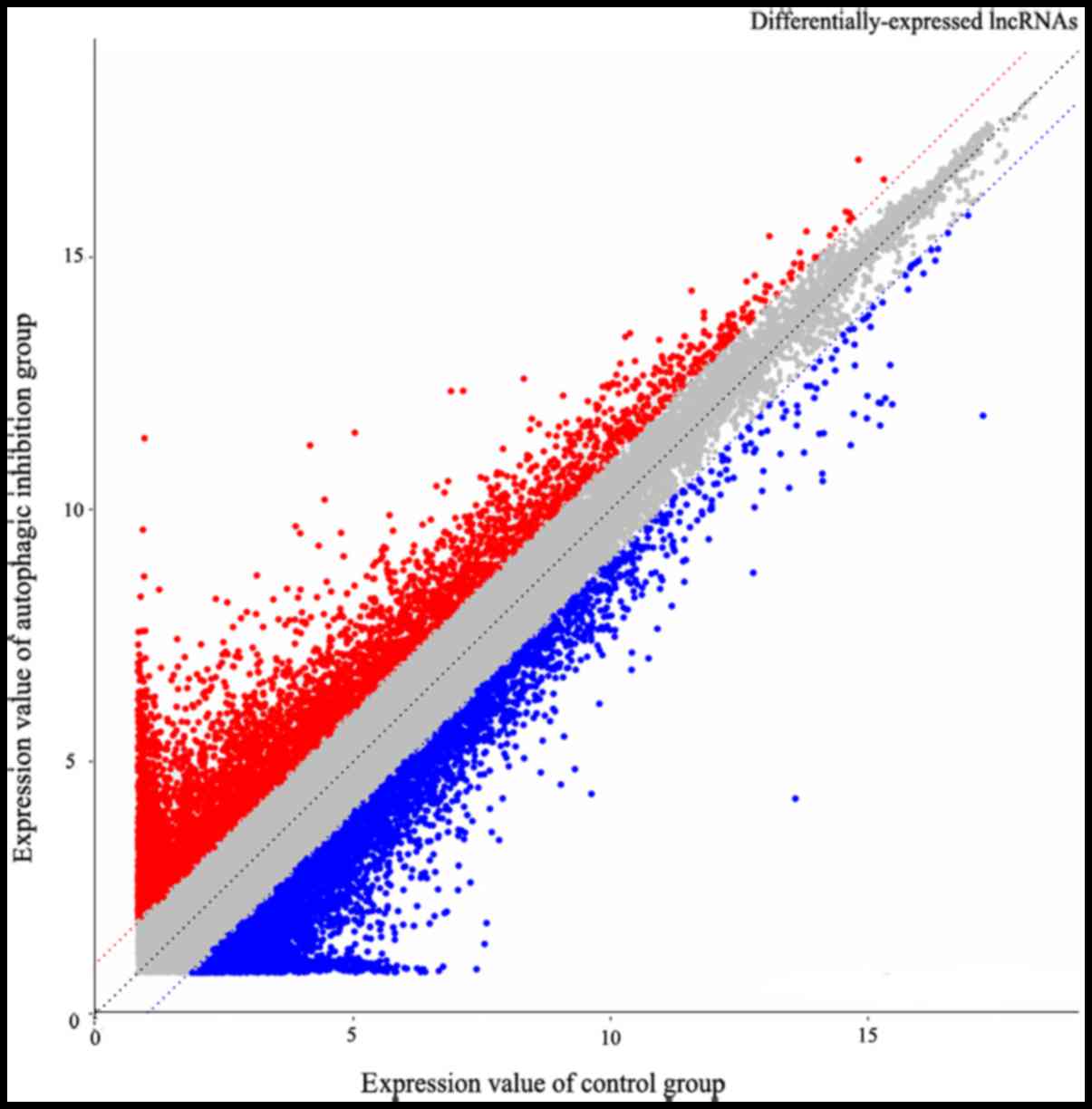
(Fig. 3). Additionally, 3,184
differentially-expressed lncRNAs were observed, including 1,637
downregulated and 1,547 upregulated (Fig. 4), whereas 9,420 circRNAs were
determined to be differentially expressed, including 4,223
downregulated and 5,197 upregulated (Fig. 5). The miRNA microarray results
demonstrated that the expression levels of two miRNAs
(hsa-miR-663a-5p and hsa-miR-154-3p) were underexpressed in the
PANC-1 cells in the autophagy-suppression group, compared with the
control group. No upregulated miRNA was determined. The cutoff
values of the aforementioned differentials were all set to a fold
change>2 or FC<0.5 and P<0.05.
GO and KEGG pathway analyses were performed using
the DAVID database. The 3,966 differentially-expressed mRNAs in the
chloroquine diphosphate treatment group (autophagy suppression
group), compared with the control group, were subjected to
bioinformatics analysis of their functions. The results
demonstrated that these genes were concentrated in the biological
processes, molecular functions and cellular components categories
in the GO analysis (Table I), and
were involved in the regulation of multiple KEGG signaling pathways
including pathways in cancer and the mitogen-activated protein
kinase signaling pathway (Table
II). Of these functions, the involvement of the
autophagy-associated pathway (Autophagy) was notable. Additionally,
the differentially-expressed genes in this pathway included a
number of recognized autophagy-associated genes, including
autophagy-related 12 (ATG12), GABA type A receptor associated
protein like 1 (GABARAPL1) and ULK2, which were differentially
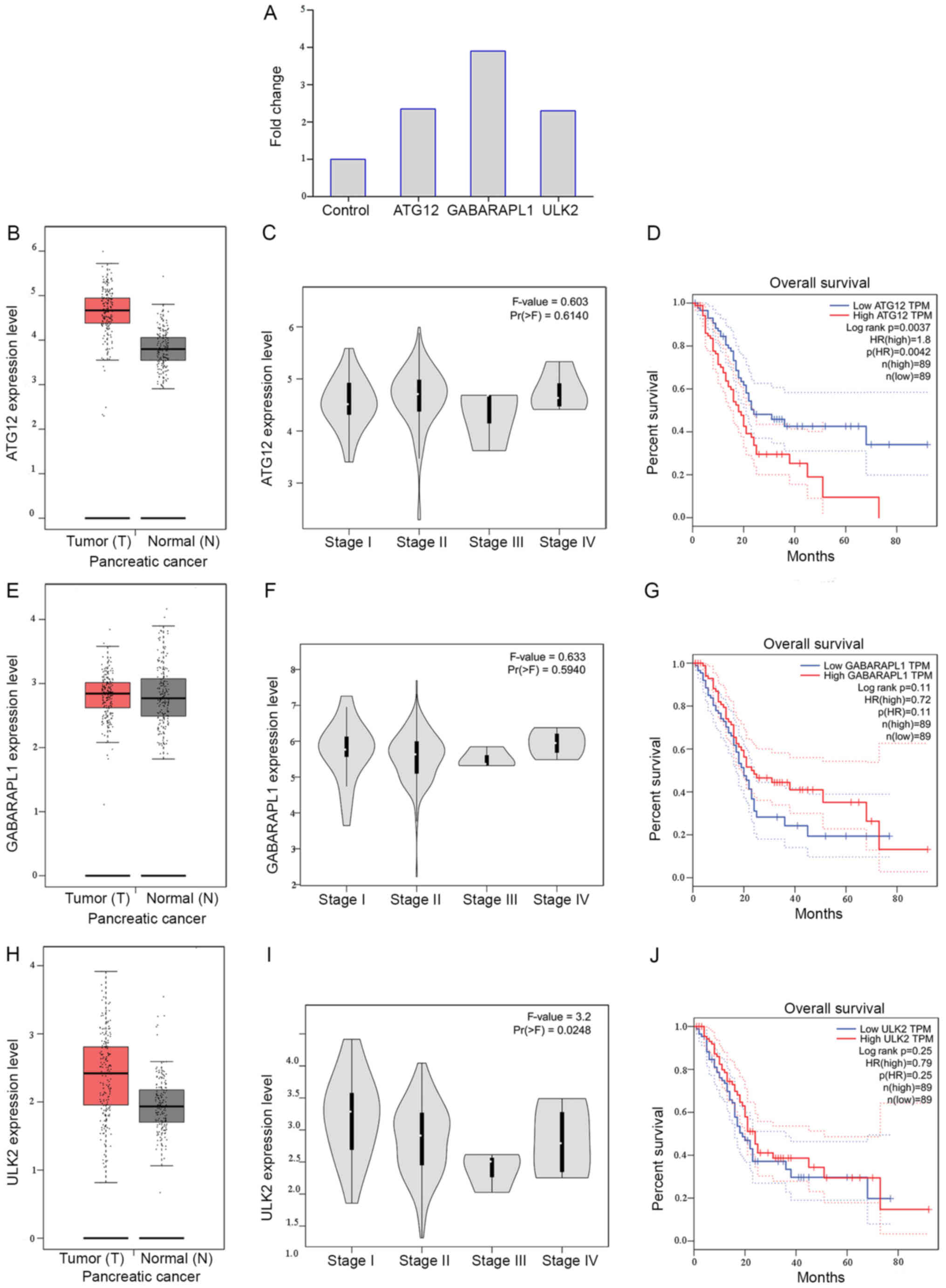
expressed in the present ceRNA microarray results also (Fig. 6). Following analyzing the clinical
roles of the three genes via the Gene Expression Profiling
Interactive Analysis online database, which is based on the data
obtained from The Cancer Genome Atlas and GTEx, it was determined
that the expression of ATG12 was upregulated in pancreatic cancer
tissue, compared with non-tumor tissue. Furthermore, high
expression of ATG12 was associated with significantly reduced
survival time (P<0.01). Notably, the expression of ATG12,
GABARAPL1 and ULK2 were increased in pancreatic cancer, compared
with the control group (Fig. 6).
These results validated that these autophagy-associated genes were
involved in the onset and progression of pancreatic cancer.
These results were integrated with the
differentially-expressed mRNAs that were actually measured. The
co-expressed genes were subjected to GO and KEGG pathway analyses
using the DAVID database.
A total of 1,726 target genes of miR-663a-5p were
detected by at least 6 platforms using the aforementioned 12 online
miRNA target gene prediction databases. Following cross-checking
with the differentially-expressed genes in the autophagy
suppression group derived from chloroquine diphosphate treatment,
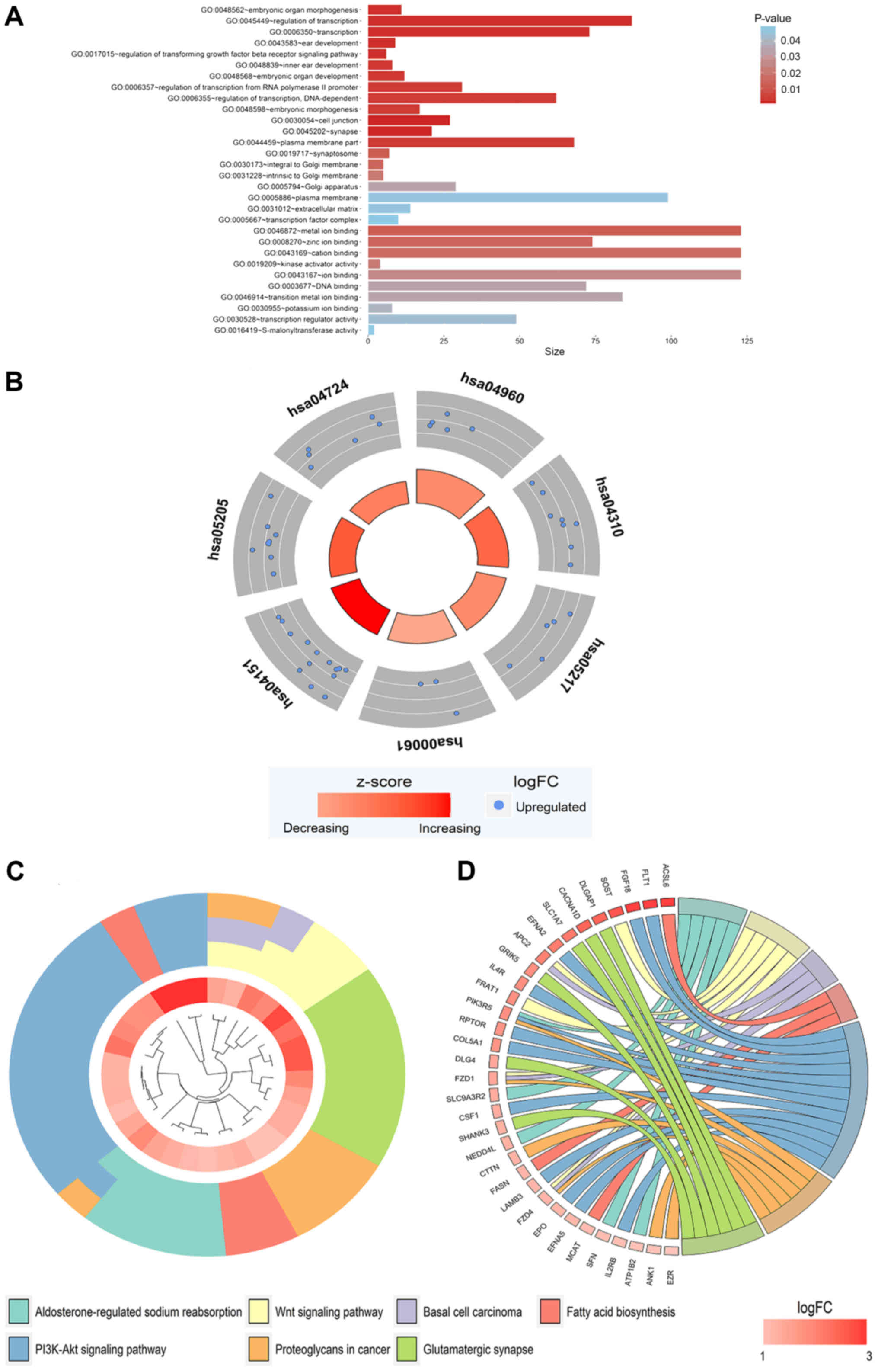
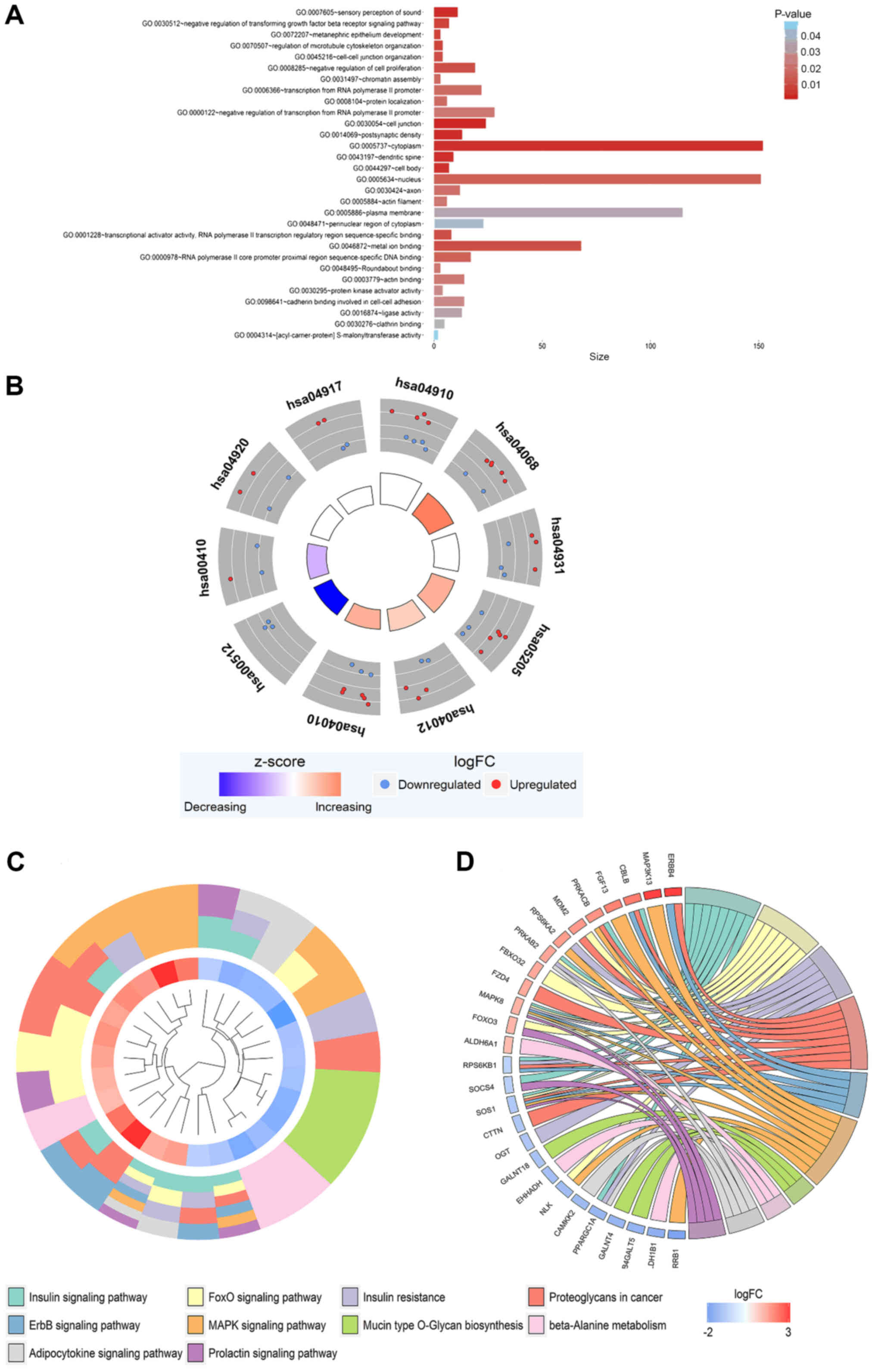
462 co-expressed genes were obtained (Fig. 7). These genes were subjected to GO
and KEGG pathway analyses. The results demonstrated that these
genes were concentrated in the biological processes, molecular
functions and cellular components categories in the GO analysis
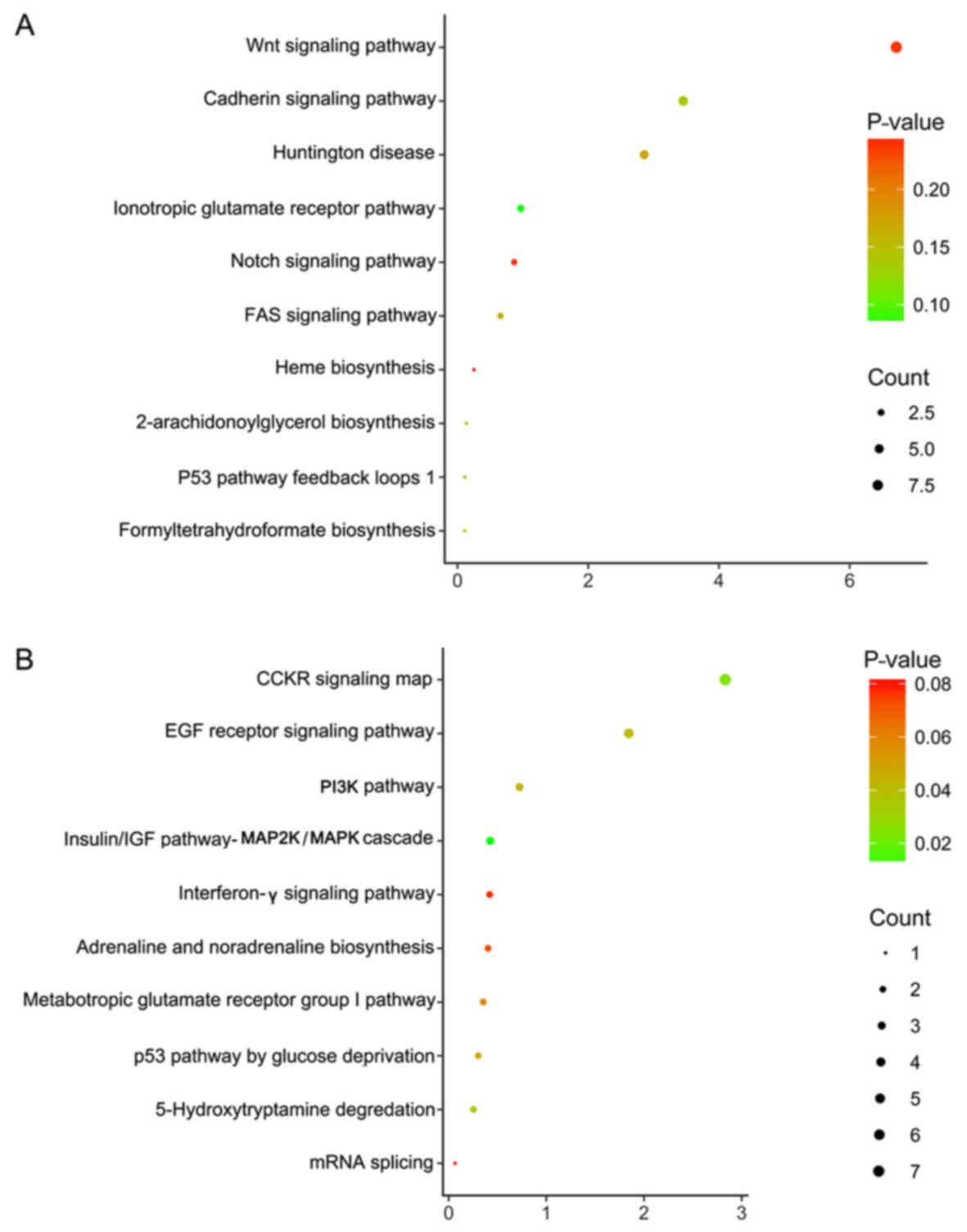
(Fig. 8A, Table III) and were involved in the
regulation of multiple KEGG signaling pathways (Table III), including the
aldosterone-regulated sodium reabsorption and Wnt signaling
pathways (Fig. 8B-D). However,
there were no significant pathways determined by PANTHER analysis
(P>0.05).
A total of 294 target genes of miR-154-3p were
detected by at least 6 programs using the aforementioned 12 online
miRNA target gene prediction databases. Following cross-checking
with the differentially-expressed genes in the autophagy
suppression group derived from chloroquine diphosphate treatment,
294 co-expressed genes were obtained (Fig. 9). These genes were subjected to GO
and KEGG pathway analyses. The results demonstrated that these
genes were concentrated in the biological processes, molecular
functions and cellular components categories in the GO analysis
(Fig. 10A, Table IV), and were involved in the
insulin signaling, Forkhead Box O (FoxO) signaling and
proteoglycans in cancer pathways (Table IV, Fig. 10B-D). Additionally, the results of
PANTHER analysis revealed these genes were concentrated in a
numbered of pathways, including the p53 pathway by glucose
deprivation and the classical gastrin cholecystokinin receptor
(CCKR) signaling map (Table V,
Fig. 11).
Different online platforms were used to predict the
target genes of the differentially-expressed miRNAs. The target
circRNAs and lncRNAs of the differentially-expressed miRNAs were
searched based on the prediction of the target miRNAs of the
differentially-expressed circRNAs and lncRNAs revealed by
microarray detection, and the differentially-expressed miRNAs that
were detected. The ceRNAs that were negatively associated with the
expression of the miRNAs were selected.
A total of 21 differentially-expressed circRNAs that
targeted miR-663a-5p were determined, of which nine
(hsa_circ_0003176, hsa_circ_0048579, hsa_circ_0063706,
hsa_circ_0071922, hsa_circ_0078989, hsa_circ_0079319,
hsa_circ_0083080, hsa_circ_0089643 and hsa_circ_0090372) indicated
negative associations with miR-663a-5p expression. hsa_circ_0071922
had the highest number of binding sites for miR-663a-5p, with six.
Simultaneously, 45 differentially-expressed lncRNAs that targeted
miR-663a-5p were determined, of which 8 lncRNAs (RP11-59C5.3,
RP13-516M14.8, RP11-196G18.24, AJ006995.3, AC024560.2, PPP1R1C,
LINC00595 and HAGLROS) were negatively associated with miR-663a-5p
expression. All of these lncRNAs had one miR-663a-5p binding site
(Table VI). Subsequently, 144
common predicted targets from at least 8 among 12 programs were
selected to overlap with the 3,966 differentially-expressed genes
following autophagy inhibition. Collectively, 46 potential targets
were determined. Thus, a ceRNA hypothesis figure with miR-663a-5p,
9 circRNAs, 8 lncRNAs and 46 genes was depicted in Fig. 12.
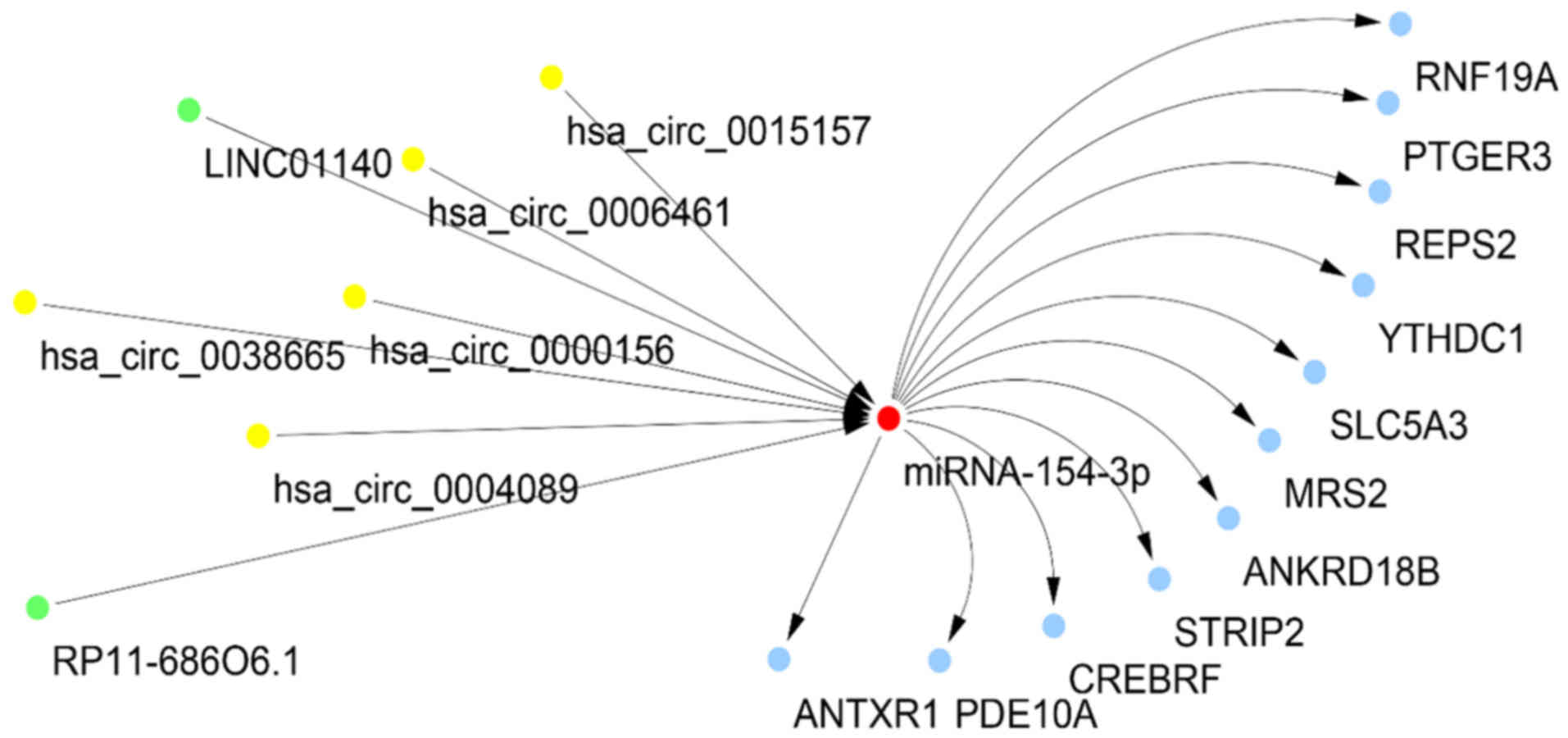
A total of 6 differentially-expressed circRNAs that
targeted miR-154-3p were determined, of which five
(hsa_circ_0000156, hsa_circ_0004089, hsa_circ_0006461,
hsa_circ_0015157 and hsa_circ_0038665) indicated a negative
association with miR-154-3p expression. All of these circRNAs had
two miR-154-3p binding sites. Simultaneously, 16
differentially-expressed lncRNAs that targeted miR-154-3p were
determined, of which two (RP11-686O6.1 and LINC01140) were
negatively associated with miR-154-3p expression. These lncRNAs had
one miR-154-3p binding site (Table
VI). Similarly to miR-663a-5p, 67 common predicted targets of
miR-154-3p from at least 8 among 12 platforms were collected, and
then they were intersected into the 3,966 differentially-expressed
genes following autophagy suppression. Eventually, 11 potential
targets were collected. Hence, a ceRNA hypothesis network with
miR-154-3p, 5 circRNAs, 2 lncRNAs and 11 genes was presented
(Fig. 13).
A comprehensive analysis for the expression of
miR-663a-5p and miR-154-3p in pancreatic cancer tissues was
performed based on RT-qPCR and GEO data. The combined standardized
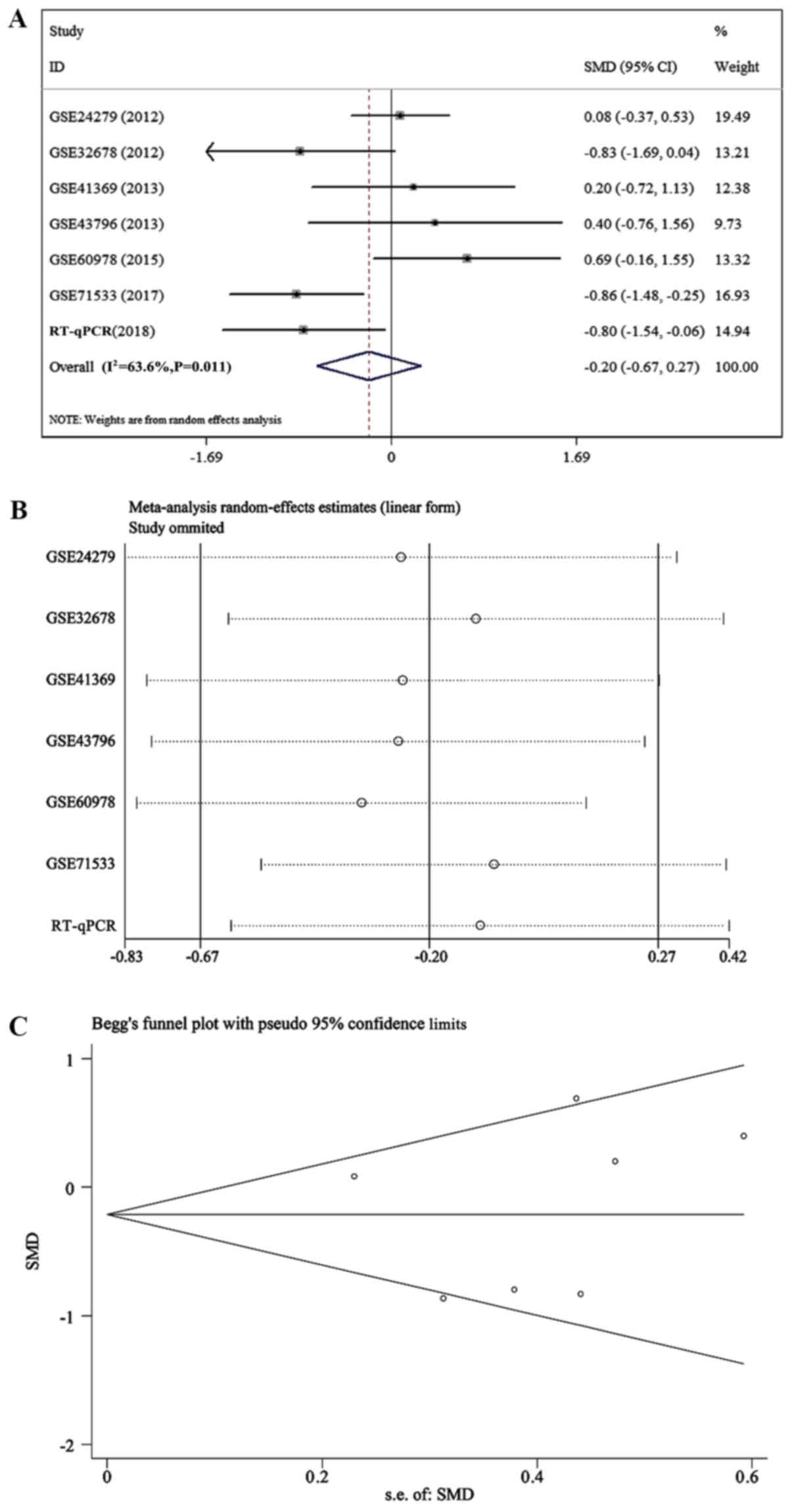
mean difference (SMD) values of miR-663a-5p was −0.203 [95%
confidence interval (CI), −0.675-0.269; Fig. 14A)], which indicated that miR-663a
had low expression in pancreatic cancer tissues; however,
heterogeneity existed (I2=63.6%; P=0.011; Fig. 14B). Sensitivity analysis and
publication bias of miR-663a-5p were depicted in Fig. 14C. The combined SMD values of
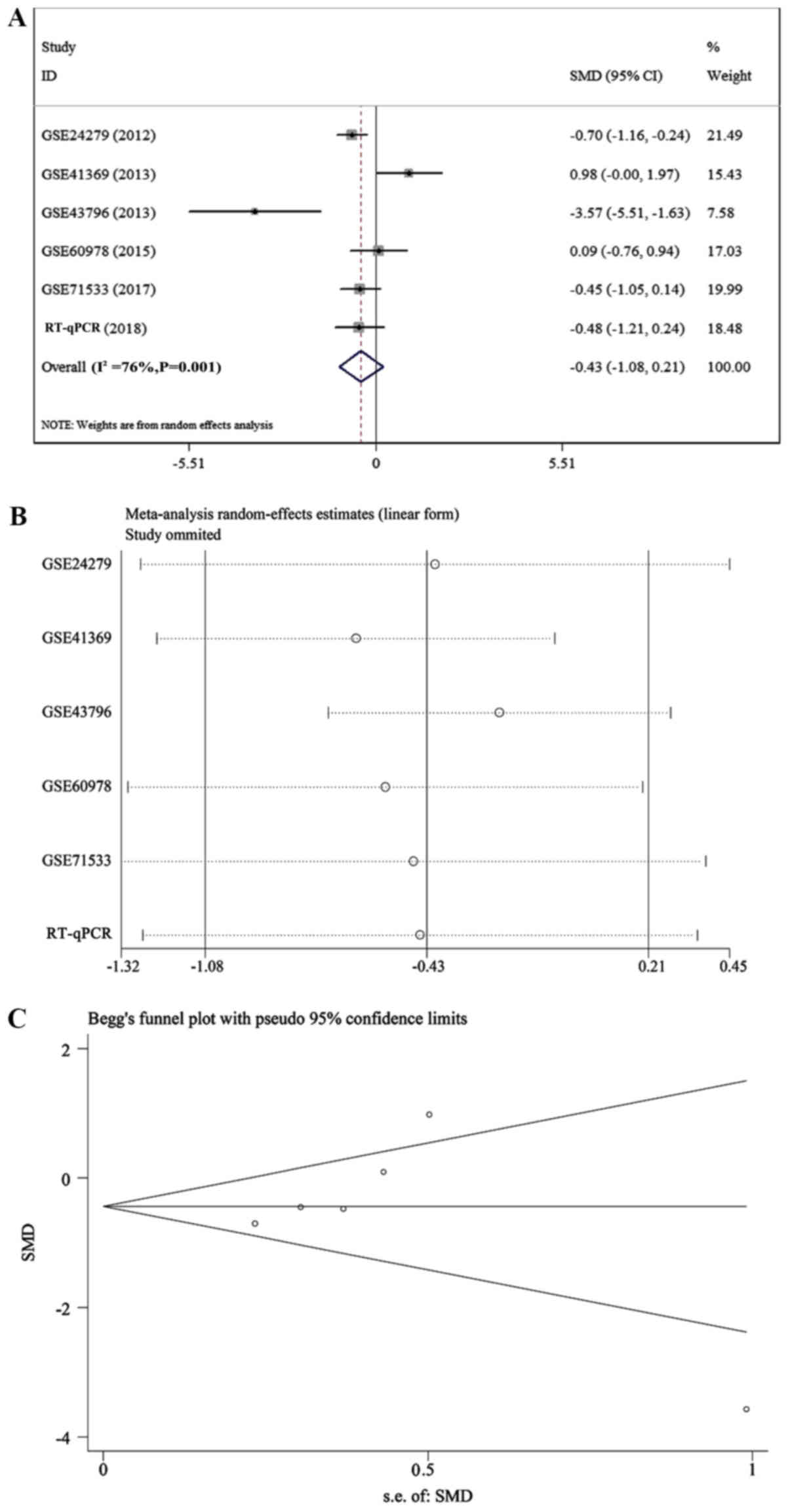
miR-154-3p was −0.434 (95% CI, −1.079-0.212; Fig. 15A)], which indicated that
miR-154-3p had low expression in pancreatic cancer tissues;
however, heterogeneity existed (I2=76.0%; P=0.001;
Fig. 15B). Sensitivity analysis
and publication bias of miR-154-3p were depicted in Fig. 15C.
Pancreatic cancer is one of the common malignancy
types of the digestive system. Due to the lack of an effective
early diagnosis, numerous patients are already in the advanced
stage of the cancer when diagnosed (85). Currently, pancreatic cancer
treatment remains dominated by surgical resection, which has a low
five-year survival rate (86-88).
Therefore, there is an urgent requirement for a series of effective
markers for pancreatic cancer to change the current status of the
poor efficacy of individualized treatment. Recent studies
demonstrated that although ncRNAs do not encode proteins, they
serve a pivotal role in the regulation of a variety of malignant
tumor types, including gastric (89), pancreatic (90), prostate (91) and breast cancer (92). Over the past two years, the
extensively investigated ncRNAs, including lncRNAs and miRNAs, and
circRNAs, which were newly revealed to have similar regulatory
functions, particularly to miRNAs, were indicated to also serve
crucial roles (44,93-97).
Abnormal expression of a large number of miRNAs has been determined
in malignant tumor tissues (98-100). For example, it has been reported
that miR-221/222 and miR-15b serve a role in causing malignant
tumors (101). Additionally, it
was determined that miR-451 and miR-126 have an abnormal expression
in lung cancer (102), and the
targeted delivery of miRNA therapeutics was a promising strategy
for cancer (103). This abnormal
expression pattern affects the occurrence, development and
prognosis of the tumor by directly regulating the biological
functions of the targeted mRNAs (102,104-112). Studies have demonstrated that a
variety of miRNAs exhibit abnormal expression patterns in
pancreatic cancer cells that affect the occurrence, development and
prognosis of pancreatic cancer; for example, the upregulated
expression of miR-10b, miR-21, miR23a and miR-27a in pancreatic
cancer affects cell growth, proliferation and apoptotic metastasis
by targeting programmed cell death 4, BTG anti-proliferation factor
2, neural precursor cell expressed, developmentally downregulated
4-like, phosphatase and tensin homolog, HIV-1 Tat interactive
protein 2 and p16 (113-117).
With the increased understanding of the regulatory
mechanism of ncRNAs, ceRNAs have attracted increasing attention as
important regulators of miRNA activity. ceRNA refers to lncRNAs and
circRNAs that suppress miRNA expression through targeted binding to
the miRNA, and thus regulate its activity, which affects the
development, progression and prognosis of tumors (118-120). Studies have indicated that
lncRNA-metastasis associated lung adenocarcinoma transcript 1 can
target miR-124 to reduce its expression and activate
cyclin-dependent kinase 4, thereby accelerating the progression of
breast cancer (121-123). A recent study on pancreatic
cancer demonstrated that lncRNA-urothelial cancer associated 1 was
overexpressed in pancreatic cancer tissues and reduced miR-135a
expression by adsorbing it, thereby inhibiting genes associated
with tumor growth and metastasis (124). These examples highlight the
significance of the interactions among lncRNAs, miRNAs and mRNAs
for cancer diagnosis and treatment.
Therefore, our aim was to mine pancreatic cancer
autophagy-associated ceRNAs and miRNAs through assays with ceRNA
and miRNA microarrays, respectively, to reveal their targets using
bioinformatics methods, construct ceRNA, miRNA and mRNA pathways,
predict the potential molecular mechanisms of the ceRNA, miRNA and
mRNA pathways in the autophagy of pancreatic cancer, and provide
novel ideas and directions for further studies of pancreatic
cancer.
Firstly, the differentially-expressed mRNAs were
analyzed. The results demonstrated that PANC-1 cells treated with
chloroquine diphosphate had multiple differentially-expressed
mRNAs, compared with the control group, using the FC value FC>2
or FC<0.5 as the threshold. The bioinformatics analysis of these
genes indicated that as the autophagic level changed, the
differentially-expressed genes were primarily concentrated in
tumor-associated and pancreatic cancer-associated pathways.
Notably, the autophagy-associated pathway, in which three genes
(ATG12, GABARAPL1 and ULK2) were abnormally expressed when the
autophagic level, was inhibited. ATG12 is an autophagy-related
protein that has been demonstrated to function as a key
autophagy-associated target gene regulated by miR-23b, and
suppression of ATG12 significantly increased the radiosensitivity
of pancreatic cancer cells, whereas the miR-23b-induced
radiosensitivity was eliminated by ATG12 overexpression (125). Studies indicated that ULK served
an important role in the autophagy process (126-128). ULK2 and ULK1 are highly
homologous, functionally complementary and indispensable for
affecting autophagy (129-131).
GABARAPL1 is considered as a target of miR-195 and regulates the
proliferation, migration, angiogenesis and autophagy of endothelial
progenitor cells (132). These
three genes (ATG12, GABARAPL1 and ULK2) may serve pivotal roles in
the autophagy suppressed by chloroquine diphosphate in pancreatic
cancer cells.
To determine relevant miRNAs and elucidate their
potential regulatory mechanisms in the autophagy of pancreatic
cancer, differentially-expressed miRNAs were analyzed and their
target genes were predicted. Subsequently, the results were
compared with the target genes detected in the microarray analysis.
The co-expressed genes were considered the putative target genes
regulated by the abnormally-expressed miRNAs during the autophagy
process in pancreatic cancer cells. Bioinformatics analyses were
performed on these target genes to identify the signaling pathways
and biological processes that were regulated. The results
demonstrated that the expression of miR-663a-5p and miR-154-3p was
downregulated in the PANC-1 cells treated with chloroquine
diphosphate, compared with the control group. miR-663a-5p
expression in pancreatic cancer tissues is significantly
downregulated and negatively associated with eukaryotic translation
elongation factor 1α 2 (eEF1A2) expression due to miR-663 reducing
the proliferation and invasion of pancreatic cells by directly
targeting eEF1A2 in vitro and in vivo (133). Additionally, miR-663a-5p has been
reported to have a low expression in hepatocellular carcinoma
(134), non-small cell lung
cancer (135) and colorectal
cancer (136). Similarly, the
present comprehensive analysis indicated that the expression of
miR-663a-5p was downregulated in pancreatic ductal adenocarcinoma
(PDAC). In the present study, the bioinformatics analyses indicated
that the putative autophagy pathways in the pancreatic cancer cells
regulated by miR-663a-5p included the aldosterone-regulated sodium
reabsorption, Wnt signaling, basal cell carcinoma, fatty acid
biosynthesis and PI3K-Akt signaling pathways, of which the Wnt
signaling and PI3K-Akt signaling pathways were considered to have a
function in the regulation of autophagy (137-143). Currently, whether miR-154-3p is
involved in the biological processes of pancreatic cancer cells,
including occurrence, development, prognosis and autophagy, remains
unknown; however, miR-154-3p was reported to be expressed at a low
level in colorectal cancer and was associated with the degree of
malignancy of colorectal cancer (144). Similarly, miR-154-3p exhibited
low expression in breast cancer and affected the treatment outcome
of breast cancer by regulating E2F transcription factor 5 (145). Additionally, the present
comprehensive analysis indicated that the expression of miR-154-3p
was downregulated in PDAC. Furthermore, the bioinformatics analyses
indicated that the putative autophagy pathways in the pancreatic
cancer cells regulated by miR-154-3p included the insulin signaling
pathway, FoxO signaling, mitogen-activated protein kinase
signaling, ErbB signaling, p53 pathway by glucose deprivation and
CCKR signaling map pathways. Activation of insulin like growth
factor 1 receptor (IGF-1R) signaling antagonizes the decrease in
cell viability of human disc cells through the suppression of
apoptosis and enhancement of autophagy (146). IGF-1R increases cell viability
during hypoxia, which may be dependent on promoting autophagy by
suppressing the PI3K/Akt/mTOR signaling pathway (147). The present study demonstrated
that a number of genes, including suppressor of cytokine signaling
1 (SOCS1), plasma membrane intrinsic protein 3 (PIP3) and
serine/threonine protein phosphatase type 1 α (PP1), which were
involved in the insulin signaling pathway, were abnormally
expressed when the autophagic level was decreased. Overexpression
of SOCS1 suppressed the PI3K/PIP3/Akt signaling pathway with the
subsequent PP1 activation, which has been demonstrated to induce
autophagy (148,149). Autophagy is inhibited under the
condition of oxygen-glucose deprivation (150). Therefore, we hypothesized that
miR-154-3p influences the key genes in the insulin signaling or p53
pathways by glucose deprivation, thereby affecting autophagy.
Additionally, due to the involvement of CCKR signaling in
pancreatic enzyme secretion (151,152), miR-154-3p may serve a key role in
the secretion process. Therefore, the data in the present study
will provide novel directions for the investigation of pancreatic
cancer and its autophagy.
To elucidate which genes inhibited the miRNAs
regulating the autophagy of pancreatic cancer cells through
competitive binding, potential target associations between circRNA
and miRNA, and between lncRNA-miRNA were predicted based on the
competitive expression of ceRNA and miRNA. When these results were
combined with the differentially-expressed circRNAs and lncRNAs
under different autophagic levels detected by the microarrays,
numerous ceRNAs that exhibited target associations with miR-663a-5p
and miR-154-3p, and negative associations with the expression of
the targeted miRNAs under the same changes in the autophagic level
were determined. AC024560.2 is an effective lncRNA residing on
chromosome 3 that can be used to predict the metastasis of early
cervical cancer to lymph nodes (153). In the present study, AC024560.2
was determined to competitively bind to miR-663a-5p and thus
regulate the autophagic level of pancreatic cancer cells by
inhibiting the expression of this miRNA. For the other ceRNAs,
their associations with tumors have not been reported, and further
investigations are required.
Since the molecular mechanisms of tumor occurrence
and development are complex and the numerous different
physiological processes, including differentiation (154) and aging (155), are frequently accompanied by an
aberrant autophagic level (156,157), it is not surprising that the
interference of autophagy is determined to be one of the
therapeutic methods for tumors. The growth of PDAC could be
repressed by hydroxychloroquine in vivo and in vitro
(158). PDAC was dependent on
autophagy and the use of an autophagy inhibitor may become the
breakthrough point of treatment (35). However, in the treatment of PDAC
with gemcitabine, a high concentration of gemcitabine is required
following adding autophagy inhibitors (37). Therefore, autophagy may serve a
dual role in the development of PDAC. Investigating the ceRNA
network following inhibiting autophagy, of which these genes may
serve role in inhibiting autophagy or promoting autophagy, will
provide novel ideas for determining new ways to treat pancreatic
cancer. On this basis, the biological function and action mechanism
of these two miRNAs will be investigated in further studies.
In summary, the miRNAs that were significantly
associated with the autophagy of pancreatic cancer included
miR-663a-5p and miR-154-3p. These miRNAs were used as the
intermediate regulatory sites, upstream to which multiple putative
ceRNAs, including AC024560.2, were present and downstream to
multiple targeted regulatory genes, including ATG12 and ULK1. The
expression of these miRNAs was regulated through ceRNA, miRNA and
mRNA interactions, which could serve important regulatory roles in
pancreatic cancer autophagy.
In conclusion, autophagy serves an important role in
the development and progression of various malignant tumor types,
including lung (159), gastric
(160) and renal cancer (161). The investigation of
differentially-expressed genes in pancreatic cancer cells under
different autophagic levels not only clarifies the regulatory
mechanism of the autophagic process in pancreatic cancer cells, but
also provides novel ideas for the effective diagnosis and treatment
of pancreatic cancer. In the present study, microarray technology
was employed to confirm that the change in the autophagic level of
pancreatic cancer cells was accompanied by various
differentially-expressed genes. Following establishing multiple
ceRNA, miRNA and mRNA associations, there is more reason to
consider that these genes will serve important roles in the
diagnosis and treatment of pancreatic cancer, and thus lay the
theoretical foundation for subsequent investigations on pancreatic
cancer while indicating new research directions. Subsequently, the
regulatory functions of these genes at the cellular and tissue
levels will be investigated and their specificity and accuracy will
be further validated through a series of in vivo and in
vitro experiments.
The study was supported by the funds of the National
Natural Science Foundation of China (grant no. NSFC81560448), the
Natural Science Foundation of Guangxi, China (grant no.
2016GXNSFBA380039), Medical Excellence Award Funded by the Creative
Research Development Grant from the First Affiliated Hospital of
Guangxi Medical University, Guangxi Medical University Training
Program for Distinguished Young Scholars, and the Promoting Project
of Basic Capacity for University Young and Middle-aged Teachers in
Guangxi (grant no. KY2016LX031). The funders had no role in the
study design, the data collection and analysis, the decision to
publish, or the preparation of the manuscript.
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
DMW and MTJ wrote the paper, performed the
experiments and conducted bioinformatics analysis. PL, HY and YWD
wrote the paper and conducted bioinformatics analysis. QY and DYL
conducted bioinformatics analysis and statistical data analysis.
DZL and GC designed the project, supervised the experiments and
corrected the draft.
The present study was approved by the Ethics
Committee of the First Affiliated Hospital of Guangxi Medical
University, and all patients provided signed informed consent.
Consent for the publication of the pathological data
was obtained from all patients who were involved in the present
study.
The authors declare that they have no competing
interests.
The authors would like to thank all publicly
available data used in the present study.
|
1
|
Ryan DP, Hong TS and Bardeesy N:
Pancreatic adenocarcinoma. N Engl J Med. 371:2140–2141. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Swords DS, Firpo MA, Scaife CL and
Mulvihill SJ: Biomarkers in pancreatic adenocarcinoma: Current
perspectives. OncoTargets Ther. 9:7459–7467. 2016. View Article : Google Scholar
|
|
3
|
Siegel RL, Miller KD and Jemal A: Cancer
Statistics, 2017. CA Cancer J Clin. 67:7–30. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2018. CA Cancer J Clin. 68:7–30. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Song HY, Wang Y, Lan H and Zhang YX:
Expression of Notch receptors and their ligands in pancreatic
ductal adenocarcinoma. Exp Ther Med. 16:53–60. 2018.PubMed/NCBI
|
|
6
|
Xu Q, Gao J and Li Z: Identification of a
novel alternative splicing transcript variant of the suppressor of
fused: Relationship with lymph node metastasis in pancreatic ductal
adenocarcinoma. Int J Oncol. 49:2611–2619. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Li Q, Zhang L, Li X, Yan H, Yang L, Li Y,
Li T, Wang J and Cao B: The prognostic significance of human
epidermal growth factor receptor family protein expression in
operable pancreatic cancer: HER14 protein expression and prognosis
in pancreatic cancer. BMC Cancer. 16:9102016. View Article : Google Scholar
|
|
8
|
Li H, Hao X, Wang H, Liu Z, He Y, Pu M,
Zhang H, Yu H, Duan J and Qu S: Circular RNA Expression Profile of
Pancreatic Ductal Adenocarcinoma Revealed by Microarray. Cell
Physiol Biochem. 40:1334–1344. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Zhu L, Staley C, Kooby D, El-Rays B, Mao H
and Yang L: Current status of biomarker and targeted nanoparticle
development: The precision oncology approach for pancreatic cancer
therapy. Cancer Lett. 388:139–148. 2017. View Article : Google Scholar :
|
|
10
|
Kocaturk NM and Gozuacik D: Crosstalk
Between Mammalian Autophagy and the Ubiquitin-Proteasome System.
Front Cell Dev Biol. 6:1282018. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Hong JC, Czito BG, Willett CG and Palta M:
A current perspective on stereotactic body radiation therapy for
pancreatic cancer. OncoTargets Ther. 9:6733–6739. 2016. View Article : Google Scholar
|
|
12
|
Marsoner K, Haybaeck J, Csengeri D, Waha
JE, Schagerl J, Langeder R, Mischinger HJ and Kornprat P:
Pancreatic resection for intraductal papillary mucinous neoplasm -
a thirteen-year single center experience. BMC Cancer. 16:8442016.
View Article : Google Scholar
|
|
13
|
Kang MJ, Jang JY and Kim SW: Surgical
resection of pancreatic head cancer: What is the optimal extent of
surgery. Cancer Lett. 382:259–265. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Ellerhoff TP, Berchtold S, Venturelli S,
Burkard M, Smirnow I, Wulff T and Lauer UM: Novel
epi-virotherapeutic treatment of pancreatic cancer combining the
oral histone deacetylase inhibitor resminostat with oncolytic
measles vaccine virus. Int J Oncol. 49:1931–1944. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Zuo C, Sheng X, Ma M, Xia M and Ouyang L:
ISG15 in the tumorigenesis and treatment of cancer: An emerging
role in malignancies of the digestive system. Oncotarget.
7:74393–74409. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Chang JH, Jiang Y and Pillarisetty VG:
Role of immune cells in pancreatic cancer from bench to clinical
application: An updated review. Medicine (Baltimore). 95:e55412016.
View Article : Google Scholar
|
|
17
|
Kim KH and Lee MS: Autophagy--a key player
in cellular and body metabolism. Nat Rev Endocrinol. 10:322–337.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Tao Z, Li T, Ma H, Yang Y, Zhang C, Hai L,
Liu P, Yuan F, Li J, Yi L, et al: Autophagy suppresses self-renewal
ability and tumorigenicity of glioma-initiating cells and promotes
Notch1 degradation. Cell Death Dis. 9:10632018. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Mowers EE, Sharifi MN and Macleod KF:
Autophagy in cancer metastasis. Oncogene. 36:1619–1630. 2017.
View Article : Google Scholar :
|
|
20
|
Tan YQ, Zhang J and Zhou G: Autophagy and
its implication in human oral diseases. Autophagy. 13:225–236.
2017. View Article : Google Scholar :
|
|
21
|
Green DR, Oguin TH and Martinez J: The
clearance of dying cells: Table for two. Cell Death Differ.
23:915–926. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Sionov RV, Vlahopoulos SA and Granot Z:
Regulation of Bim in Health and Disease. Oncotarget. 6:23058–23134.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Gafar AA, Draz HM, Goldberg AA, Bashandy
MA, Bakry S, Khalifa MA, AbuShair W, Titorenko VI and Sanderson JT:
Lithocholic acid induces endoplasmic reticulum stress, autophagy
and mitochondrial dysfunction in human prostate cancer cells.
PeerJ. 4:e24452016. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Jawhari S, Ratinaud MH and Verdier M:
Glioblastoma, hypoxia and autophagy: A survival-prone
‘ménage-à-trois’. Cell Death Dis. 7:e24342016. View Article : Google Scholar
|
|
25
|
Mao S and Zhang J: Role of autophagy in
chronic kidney diseases. Int J Clin Exp Med. 8:22022–22029.
2015.
|
|
26
|
Mo He Y, Luo Q, Qiao B, Xu Y, Zuo R, Deng
Z, Nong J, Peng X, He GW, et al: Induction of apoptosis and
autophagy via mitochondria- and PI3K/Akt/mTOR-mediated pathways by
E. adenophorum in hepatocytes of saanen goat. Oncotarget.
7:54537–54548. 2016.PubMed/NCBI
|
|
27
|
Goulielmaki M, Koustas E, Moysidou E,
Vlassi M, Sasazuki T, Shirasawa S, Zografos G, Oikonomou E and
Pintzas A: BRAF associated autophagy exploitation: BRAF and
autophagy inhibitors synergise to efficiently overcome resistance
of BRAF mutant colorectal cancer cells. Oncotarget. 7:9188–9221.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Vera-Ramirez L, Vodnala SK, Nini R, Hunter
KW and Green JE: Autophagy promotes the survival of dormant breast
cancer cells and metastatic tumour recurrence. Nat Commun.
9:19442018. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Li SJ, Sun SJ, Gao J and Sun FB: Wogonin
induces Beclin-1/PI3K and reactive oxygen species-mediated
autophagy in human pancreatic cancer cells. Oncol Lett.
12:5059–5067. 2016. View Article : Google Scholar
|
|
30
|
Klein K, Werner K, Teske C, Schenk M,
Giese T, Weitz J and Welsch T: Role of TFEB-driven autophagy
regulation in pancreatic cancer treatment. Int J Oncol. 49:164–172.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Kwon JJ, Willy JA, Quirin KA, Wek RC, Korc
M, Yin XM and Kota J: Novel role of miR-29a in pancreatic cancer
autophagy and its therapeutic potential. Oncotarget. 7:71635–71650.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Ranjan A and Srivastava SK: Penfluridol
suppresses pancreatic tumor growth by autophagy-mediated apoptosis.
Sci Rep. 6:261652016. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Seo JW, Choi J, Lee SY, Sung S, Yoo HJ,
Kang MJ, Cheong H and Son J: Autophagy is required for PDAC
glutamine metabolism. Sci Rep. 6:375942016. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Yan Y, Jiang K, Liu P, Zhang X, Dong X,
Gao J, Liu Q, Barr MP, Zhang Q, Hou X, et al: Bafilomycin A1
induces caspase-independent cell death in hepatocellular carcinoma
cells via targeting of autophagy and MAPK pathways. Sci Rep.
6:370522016. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Yang S, Wang X, Contino G, Liesa M, Sahin
E, Ying H, Bause A, Li Y, Stommel JM, Dell’antonio G, et al:
Pancreatic cancers require autophagy for tumor growth. Genes Dev.
25:717–729. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Yang A, Herter-Sprie G, Zhang H, Lin EY,
Biancur D, Wang X, Deng J, Hai J, Yang S, Wong KK, et al: Autophagy
sustains pancreatic cancer growth through both cell autonomous and
non-autonomous mechanisms. Cancer Discov. 8:276–287. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Mukubou H, Tsujimura T, Sasaki R and Ku Y:
The role of autophagy in the treatment of pancreatic cancer with
gemcitabine and ionizing radiation. Int J Oncol. 37:821–828.
2010.PubMed/NCBI
|
|
38
|
Marinković M, Šprung M, Buljubašić M and
Novak I: Autophagy Modulation in Cancer: Current Knowledge on
Action and Therapy. Oxid Med Cell Longev. 2018.8023821:2018.
|
|
39
|
Islam MA, Reesor EK, Xu Y, Zope HR, Zetter
BR and Shi J: Biomaterials for mRNA delivery. Biomater Sci.
3:1519–1533. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Liu F, Gao S, Yang Y, Zhao X, Fan Y, Ma W,
Yang D, Yang A and Yu Y: Antitumor activity of curcumin by
modulation of apoptosis and autophagy in human lung cancer A549
cells through inhibiting PI3K/Akt/mTOR pathway. Oncol Rep.
39:1523–1531. 2018.PubMed/NCBI
|
|
41
|
Chen JF, Wu P, Xia R, Yang J, Huo XY, Gu
DY, Tang CJ, De W and Yang F: STAT3-induced lncRNA HAGLROS
over-expression contributes to the malignant progression of gastric
cancer cells via mTOR signal-mediated inhibition of autophagy. Mol
Cancer. 17:62018. View Article : Google Scholar
|
|
42
|
Gong J, Muñoz AR, Chan D, Ghosh R and
Kumar AP: STAT3 down regulates LC3 to inhibit autophagy and
pancreatic cancer cell growth. Oncotarget. 5:2529–2541. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Raimondi M, Cesselli D, Di Loreto C, La
Marra F, Schneider C and Demarchi F: USP1 (ubiquitin specific
peptidase 1) targets ULK1 and regulates its cellular
compartmentalization and autophagy. Autophagy. Oct 18–2018.Epub
ahead of print. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Pan Y, Li C, Chen J, Zhang K, Chu X, Wang
R and Chen L: The Emerging Roles of Long Noncoding RNA ROR
(lincRNA-ROR) and its Possible Mechanisms in Human Cancers. Cell
Physiol Biochem. 40:219–229. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Bamodu OA, Huang WC, Lee WH, Wu A, Wang
LS, Hsiao M, Yeh CT and Chao TY: Aberrant KDM5B expression promotes
aggressive breast cancer through MALAT1 overexpression and
downregulation of hsa-miR-448. BM. Cancer. 16:1602016.
|
|
46
|
Zhang Y, He RQ, Dang YW, Zhang XL, Wang X,
Huang SN, Huang WT, Jiang MT, Gan XN, Xie Y, et al: Comprehensive
analysis of the long noncoding RNA HOXA11-AS gene interaction
regulatory network in NSCLC cells. Cancer Cell Int. 16:892016.
View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Peng L, Yuan XQ and Li GC: The emerging
landscape of circular RNA ciRS-7 in cancer (Review). Oncol Rep.
33:2669–2674. 2015.Review. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Deng T, Yuan Y, Zhang C, Zhang C, Yao W,
Wang C, Liu R and Ba Y: Identification of Circulating MiR-25 as a
Potential Biomarker for Pancreatic Cancer Diagnosis. Cell Physiol
Biochem. 39:1716–1722. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Zhang YH, Fu J, Zhang ZJ, Ge CC and Yi Y:
LncRNA-LINC00152 down-regulated by miR-376c-3p restricts viability
and promotes apoptosis of colorectal cancer cells. Am J Transl Res.
8:5286–5297. 2016.
|
|
50
|
Qu S, Yang X, Li X, Wang J, Gao Y, Shang
R, Sun W, Dou K and Li H: Circular RNA: A new star of noncoding
RNAs. Cancer Lett. 365:141–148. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Li X, Yang L and Chen LL: The Biogenesis,
Functions, and Challenges of Circular RNAs. Mol Cell. 71:428–442.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Xuan L, Qu L, Zhou H, Wang P, Yu H, Wu T,
Wang X, Li Q, Tian L, Liu M, et al: Circular RNA: A novel biomarker
for progressive laryngeal cancer. Am J Transl Res. 8:932–939.
2016.PubMed/NCBI
|
|
53
|
Jin X, Feng CY, Xiang Z, Chen YP and Li
YM: CircRNA expression pattern and circRNA-miRNA-mRNA network in
the pathogenesis of nonalcoholic steatohepatitis. Oncotarget.
7:66455–66467. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Liu Q, Zhang X, Hu X, Dai L, Fu X, Zhang J
and Ao Y: Circular RNA Related to the Chondrocyte ECM Regulates
MMP13 Expression by Functioning as a MiR-136 ‘Sponge’ in Human
Cartilage Degradation. Sci Rep. 6:225722016. View Article : Google Scholar
|
|
55
|
Xie H, Ren X, Xin S, Lan X, Lu G, Lin Y,
Yang S, Zeng Z, Liao W, Ding YQ, et al: Emerging roles of
circRNA_001569 targeting miR-145 in the proliferation and invasion
of colorectal cancer. Oncotarget. 7:26680–26691. 2016.PubMed/NCBI
|
|
56
|
Zhong Z, Lv M and Chen J: Screening
differential circular RNA expression profiles reveals the
regulatory role of circTCF25-miR-103a-3p/miR-107-CDK6 pathway in
bladder carcinoma. Sci Rep. 6:309192016. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Deng X, Feng N, Zheng M, Ye X, Lin H, Yu
X, Gan Z, Fang Z, Zhang H, Gao M, et al: PM2.5 exposure-induced
autophagy is mediated by lncRNA loc146880 which also promotes the
migration and invasion of lung cancer cells. Biochim Biophys Acta,
Gen Subj. 1861.112–125. 2017.
|
|
58
|
Li C, Zhao Z, Zhou Z and Liu R: Linc-ROR
confers gemcitabine resistance to pancreatic cancer cells via
inducing autophagy and modulating the miR-124/PTBP1/PKM2 axis.
Cancer Chemother Pharmacol. 78:1199–1207. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Chen ZH, Wang WT, Huang W, Fang K, Sun YM,
Liu SR, Luo XQ and Chen YQ: The lncRNA HOTAIRM1 regulates the
degradation of PML-RARA oncoprotein and myeloid cell
differentiation by enhancing the autophagy pathway. Cell Death
Differ. 24:212–224. 2017. View Article : Google Scholar :
|
|
60
|
Pawar K, Hanisch C, Palma Vera SE,
Einspanier R and Sharbati S: Down regulated lncRNA MEG3 eliminates
mycobacteria in macrophages via autophagy. Sci Rep. 6:194162016.
View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Vega-Rubín-de-Celis S, Zou Z, Fernández
AF, Ci B, Kim M, Xiao G, Xie Y and Levine B: Increased autophagy
blocks HER2-mediated breast tumorigenesis. Proc Natl Acad Sci USA.
115:4176–4181. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Zarzynska JM: The importance of autophagy
regulation in breast cancer development and treatment. BioMed Res
Int. 2014.710345:2014.
|
|
63
|
Qin W, Li C, Zheng W, Guo Q, Zhang Y, Kang
M, Zhang B, Yang B, Li B, Yang H, et al: Inhibition of autophagy
promotes metastasis and glycolysis by inducing ROS in gastric
cancer cells. Oncotarget. 6:39839–39854. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Cai J, Li R, Xu X, Zhang L, Lian R, Fang
L, Huang Y, Feng X, Liu X, Li X, et al: CK1α suppresses lung tumour
growth by stabilizing PTEN and inducing autophagy. Nat Cell Biol.
20:465–478. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Chen PM, Gombart ZJ and Chen JW:
Chloroquine treatment of ARPE-19 cells leads to lysosome dilation
and intracellular lipid accumulation: possible implications of
lysosomal dysfunction in macular degeneration. Cell Biosci.
1:102011. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Li ML, Xu YZ, Lu WJ, Li YH, Tan SS, Lin
HJ, Wu TM, Li Y, Wang SY and Zhao YL: Chloroquine potentiates the
anticancer effect of sunitinib on renal cell carcinoma by
inhibiting autophagy and inducing apoptosis. Oncol Lett.
15:2839–2846. 2018.PubMed/NCBI
|
|
67
|
Maragkakis M, Vergoulis T, Alexiou P,
Reczko M, Plomaritou K, Gousis M, Kourtis K, Koziris N, Dalamagas T
and Hatzigeorgiou AG: DIANA-microT Web server upgrade supports Fly
and Worm miRNA target prediction and bibliographic miRNA to disease
association. Nucleic Acids Res 39 (Web Server issue):. W145–W148.
2011. View Article : Google Scholar
|
|
68
|
Paraskevopoulou MD, Georgakilas G,
Kostoulas N, Vlachos IS, Vergoulis T, Reczko M, Filippidis C,
Dalamagas T and Hatzigeorgiou AG: DIANA-microT web server v5.0:
Service integration into miRNA functional analysis workflows.
Nucleic Acids Res. 41(Web Server issue): W169–W173. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
69
|
John B, Enright AJ, Aravin A, Tuschl T,
Sander C and Marks DS: Human MicroRNA targets. PLoS Biol.
2:e3632004. View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Tsang JS, Ebert MS and van Oudenaarden A:
Genome-wide dissection of microRNA functions and cotargeting
networks using gene set signatures. Mol Cell. 38:140–153. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Wang X: Improving microRNA target
prediction by modeling with unambiguously identified
microRNA-target pairs from CLIP-ligation studies. Bioinformatics.
32:1316–1322. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Vejnar CE, Blum M and Zdobnov EM: miRmap
web: Comprehensive microRNA target prediction online. Nucleic Acids
Res. 41(Web Server issue): W165–W168. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Hsu SD, Chu CH, Tsou AP, Chen SJ, Chen HC,
Hsu PW, Wong YH, Chen YH, Chen GH and Huang HD: miRNAMap 2.0:
Genomic maps of microRNAs in metazoan genomes. Nucleic Acids Res.
36(Database): D165–D169. 2008. View Article : Google Scholar :
|
|
74
|
Krek A, Grün D, Poy MN, Wolf R, Rosenberg
L, Epstein EJ, MacMenamin P, da Piedade I, Gunsalus KC, Stoffel M,
et al: Combinatorial microRNA target predictions. Nat Genet.
37:495–500. 2005. View
Article : Google Scholar : PubMed/NCBI
|
|
75
|
Kertesz M, Iovino N, Unnerstall U, Gaul U
and Segal E: The role of site accessibility in microRNA target
recognition. Nat Genet. 39:1278–1284. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
76
|
Miranda KC, Huynh T, Tay Y, Ang YS, Tam
WL, Thomson AM, Lim B and Rigoutsos I: A pattern-based method for
the identification of MicroRNA binding sites and their
corresponding heteroduplexes. Cell. 126:1203–1217. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
77
|
Rehmsmeier M, Steffen P, Hochsmann M and
Giegerich R: Fast and effective prediction of microRNA/target
duplexes. RNA. 10:1507–1517. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
78
|
Lewis BP, Burge CB and Bartel DP:
Conserved seed pairing, often flanked by adenosines, indicates that
thousands of human genes are microRNA targets. Cell. 120:15–20.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
79
|
Michelson AM and Orkin SH:
Characterization of the homo-polymer tailing reaction catalyzed by
terminal deoxynucleotidyl transferase. Implications for the cloning
of cDNA. J Biol Chem. 257:14773–14782. 1982.PubMed/NCBI
|
|
80
|
Hoshino T and Inagaki F: A comparative
study of microbial diversity and community structure in marine
sediments using poly(A) tailing and reverse transcription-PCR.
Front Microbiol. 4:1602013. View Article : Google Scholar : PubMed/NCBI
|
|
81
|
Tajadini M, Panjehpour M and Javanmard SH:
Comparison of SYBR Green and TaqMan methods in quantitative
real-time polymerase chain reaction analysis of four adenosine
receptor subtypes. Adv Biomed Res. 3:852014. View Article : Google Scholar : PubMed/NCBI
|
|
82
|
Mardis E and McCombie WR: Library
Quantification Using SYBR Green-Quantitative Polymerase Chain
Reaction (qPCR). Cold Spring Harb Protoc. 2017:pdb prot0947142017.
View Article : Google Scholar
|
|
83
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
84
|
Huang W, Sherman BT and Lempicki RA:
Bioinformatics enrichment tools: Paths toward the comprehensive
functional analysis of large gene lists. Nucleic Acids Res.
37:1–13. 2009. View Article : Google Scholar :
|
|
85
|
Kang MJ, Jang JY, Chang YR, Kwon W, Jung W
and Kim SW: Revisiting the concept of lymph node metastases of
pancreatic head cancer: Number of metastatic lymph nodes and lymph
node ratio according to N stage. Ann Surg Oncol. 21:1545–1551.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
86
|
Georgiadou D, Sergenta nis T N,
Sakellariou S, Vlachodimitropoulos D, Psaltopoulou T, Lazaris AC,
Gounaris A and Zografos GC: Prognostic role of sex steroid
receptors in pancreatic adenocarcinoma. Pathol Res Pract.
212:38–43. 2016. View Article : Google Scholar
|
|
87
|
Lovecek M, Skalicky P, Klos D, Bebarova L,
Neoral C, Ehrmann J, Zapletalova J, Svebisova H, Vrba R, Stasek M,
et al: Long-term survival after resections for pancreatic ductal
adeno-carcinoma. Single centre study. Biomed Pap Med Fac Univ
Palacky Olomouc Czech Repub. 160:280–286. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
88
|
Ramacciato G, Nigri G, Petrucciani N,
Pinna AD, Ravaioli M, Jovine E, Minni F, Grazi GL, Chirletti P,
Tisone G, et al: Pancreatectomy with Mesenteric and Portal Vein
Resection for Borderline Resectable Pancreatic Cancer: Multicenter
Study of 406 Patients. Ann Surg Oncol. 23:2028–2037. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
89
|
Li PF, Chen SC, Xia T, Jiang XM, Shao YF,
Xiao BX and Guo JM: Non-coding RNAs and gastric cancer. World J
Gastroenterol. 20:5411–5419. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
90
|
Kishikawa T, Otsuka M, Ohno M, Yoshikawa
T, Takata A and Koike K: Circulating RNAs as new biomarkers for
detecting pancreatic cancer. World J Gastroenterol. 21:8527–8540.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
91
|
Bolton EM, Tuzova AV, Walsh AL, Lynch T
and Perry AS: Noncoding RNAs in prostate cancer: the long and the
short of it. Clin Cancer Res. 20:35–43. 2014. View Article : Google Scholar
|
|
92
|
Zhao Z, Li S, Song E and Liu S: The roles
of ncRNAs and histone-modifiers in regulating breast cancer stem
cells. Protein Cell. 7:89–99. 2016. View Article : Google Scholar :
|
|
93
|
Lan PH, Liu ZH, Pei YJ, Wu ZG, Yu Y, Yang
YF, Liu X, Che L, Ma CJ, Xie YK, et al: Landscape of RNAs in human
lumbar disc degeneration. Oncotarget. 7:63166–63176. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
94
|
Xu CZ, Jiang C, Wu Q, Liu L, Yan X and Shi
R: A Feed-Forward Regulatory Loop between HuR and the Long
Noncoding RNA HOTAIR Promotes Head and Neck Squamous Cell Carcinoma
Progression and Metastasis. Cell Physiol Biochem. 40:1039–1051.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
95
|
Dou C, Cao Z, Yang B, Ding N, Hou T, Luo
F, Kang F, Li J, Yang X, Jiang H, et al: Changing expression
profiles of lncRNAs, mRNAs, circRNAs and miRNAs during
osteoclastogenesis. Sci Rep. 6:214992016. View Article : Google Scholar : PubMed/NCBI
|
|
96
|
Huang M, Zhong Z, Lv M, Shu J, Tian Q and
Chen J: Comprehensive analysis of differentially expressed profiles
of lncRNAs and circRNAs with associated co-expression and ceRNA
networks in bladder carcinoma. Oncotarget. 7:47186–47200.
2016.PubMed/NCBI
|
|
97
|
Zhou X, Ye F, Yin C, Zhuang Y, Yue G and
Zhang G: The Interaction Between MiR-141 and lncRNA-H19 in
Regulating Cell Proliferation and Migration in Gastric Cancer. Cell
Physiol Biochem. 36:1440–1452. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
98
|
Zeng JH, Xiong DD, Pang YY, Zhang Y, Tang
RX, Luo DZ and Chen G: Identification of molecular targets for
esophageal carcinoma diagnosis using miRNA-seq and RNA-seq data
from The Cancer Genome Atlas: A study of 187 cases. Oncotarget.
8:35681–35699. 2017.PubMed/NCBI
|
|
99
|
Massillo C, Dalton GN, Farré PL, De Luca P
and De Siervi A: Implications of microRNA dysregulation in the
development of prostate cancer. Reproduction. 154:R81–R97. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
100
|
Murray MJ, Bell E, Raby KL, Rijlaarsdam
MA, Gillis AJ, Looijenga LH, Brown H, Destenaves B, Nicholson JC
and Coleman N: A pipeline to quantify serum and cerebrospinal fluid
microRNAs for diagnosis and detection of relapse in paediatric
malignant germ-cell tumours. Br J Cancer. 114:151–162. 2016.
View Article : Google Scholar :
|
|
101
|
Ouyang Q, Xu L, Cui H, Xu M and Yi L:
MicroRNAs and cell cycle of malignant glioma. Int J Neurosci.
126:1–9. 2016. View Article : Google Scholar
|
|
102
|
Del Vescovo V and Denti MA: microRNA and
Lung Cancer. Adv Exp Med Biol. 889:153–177. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
103
|
Rupaimoole R and Slack FJ: MicroRNA
therapeutics: Towards a new era for the management of cancer and
other diseases. Nat Rev Drug Discov. 16:203–222. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
104
|
Zhang X, Tang W, Chen G, Ren F, Liang H,
Dang Y and Rong M: An Encapsulation of Gene Signatures for
Hepatocellular Carcinoma, MicroRNA-132 Predicted Target Genes and
the Corresponding Overlaps. PLoS One. 11:e01594982016. View Article : Google Scholar : PubMed/NCBI
|
|
105
|
Lan D, Zhang X, He R, Tang R, Li P, He Q
and Chen G: MiR-133a is downregulated in non-small cell lung
cancer: A study of clinical significance. Eur J Med Res. 20:502015.
View Article : Google Scholar : PubMed/NCBI
|
|
106
|
Gan TQ, Tang RX, He RQ, Dang YW, Xie Y and
Chen G: Upregulated MiR-1269 in hepatocellular carcinoma and its
clinical significance. Int J Clin Exp Med. 8:714–721.
2015.PubMed/NCBI
|
|
107
|
Zhang X, Li P, Rong M, He R, Hou X, Xie Y
and Chen G: MicroRNA-141 is a biomarker for progression of squamous
cell carcinoma and adenocarcinoma of the lung: Clinical analysis of
125 patients. Tohoku J Exp Med. 235:161–169. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
108
|
Ren F, Ding H, Huang S, Wang H, Wu M, Luo
D, Dang Y, Yang L and Chen G: Expression and clinicopathological
significance of miR-193a-3p and its potential target astrocyte
elevated gene-1 in non-small lung cancer tissues. Cancer Cell Int.
15:802015. View Article : Google Scholar : PubMed/NCBI
|
|
109
|
Liu Y, Ren F, Luo Y, Rong M, Chen G and
Dang Y: Down-Regulation of MiR-193a-3p Dictates Deterioration of
HCC: A Clinical Real-Time qRT-PCR Study. Med Sci Monit.
21:2352–2360. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
110
|
Yang He R, Lin L, Chen X, Lin X, Wei X,
Liang F, Luo X, Wu Y, Gan YT, et al: MiR-30a-5p suppresses cell
growth and enhances apoptosis of hepatocellular carcinoma cells via
targeting AEG-1. Int J Clin Exp Pathol. 8:15632–15641. 2015.
|
|
111
|
Huang WT, Wang HL, Yang H, Ren FH, Luo YH,
Huang CQ, Liang YY, Liang HW, Chen G and Dang YW: Lower expressed
miR-198 and its potential targets in hepatocellular carcinoma: A
clinicopathological and in silico study. OncoTargets Ther.
9:5163–5180. 2016. View Article : Google Scholar
|
|
112
|
Zhang X, Tang W, Li R, He R, Gan T, Luo Y,
Chen G and Rong M: Downregulation of microRNA-132 indicates
progression in hepatocellular carcinoma. Exp Ther Med.
12:2095–2101. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
113
|
Frampton AE, Castellano L, Colombo T,
Giovannetti E, Krell J, Jacob J, Pellegrino L, Roca-Alonso L, Funel
N, Gall TM, et al: MicroRNAs cooperatively inhibit a network of
tumor suppressor genes to promote pancreatic tumor growth and
progression. Gastroenterology. 146:268-277e182014. View Article : Google Scholar
|
|
114
|
Frampton AE, Krell J, Jamieson NB, Gall
TM, Giovannetti E, Funel N, Mato Prado M, Krell D, Habib NA,
Castellano L, et al: microRNAs with prognostic significance in
pancreatic ductal adenocarcinoma: A meta-analysis. Eur J Cancer.
51:1389–1404. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
115
|
Giovannetti E, Funel N, Peters GJ, Del
Chiaro M, Erozenci LA, Vasile E, Leon LG, Pollina LE, Groen A,
Falcone A, et al: MicroRNA-21 in pancreatic cancer: Correlation
with clinical outcome and pharmacologic aspects underlying its role
in the modulation of gemcitabine activity. Cancer Res.
70:4528–4538. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
116
|
Ouyang H, Gore J, Deitz S and Korc M:
microRNA-10b enhances pancreatic cancer cell invasion by
suppressing TIP30 expression and promoting EGF and TGF-β actions.
Oncogene. 33:4664–4674. 2014. View Article : Google Scholar
|
|
117
|
Nakata K, Ohuchida K, Mizumoto K,
Kayashima T, Ikenaga N, Sakai H, Lin C, Fujita H, Otsuka T, Aishima
S, et al: MicroRNA-10b is overexpressed in pancreatic cancer,
promotes its invasiveness, and correlates with a poor prognosis.
Surgery. 150:916–922. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
118
|
Qi X, Zhang DH, Wu N, Xiao JH, Wang X and
Ma W: ceRNA in cancer: Possible functions and clinical
implications. J Med Genet. 52:710–718. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
119
|
Shao T, Wu A, Chen J, Chen H, Lu J, Bai J,
Li Y, Xu J and Li X: Identification of module biomarkers from the
dysregulated ceRNA-ceRNA interaction network in lung
adenocarcinoma. Mol Biosyst. 11:3048–3058. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
120
|
Xu J, Li Y, Lu J, Pan T, Ding N, Wang Z,
Shao T, Zhang J, Wang L and Li X: The mRNA related ceRNA-ceRNA
landscape and significance across 20 major cancer types. Nucleic
Acids Res. 43:8169–8182. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
121
|
Feng T, Shao F, Wu Q, Zhang X, Xu D, Qian
K, Xie Y, Wang S, Xu N, Wang Y, et al: miR-124 downregulation leads
to breast cancer progression via LncRNA-MALAT1 regulation and
CDK4/E2F1 signal activation. Oncotarget. 7:16205–16216.
2016.PubMed/NCBI
|
|
122
|
Jin H, Li Q, Cao F, Wang SN, Wang RT, Wang
Y, Tan QY, Li CR, Zou H, Wang D, et al: miR-124 Inhibits Lung
Tumorigenesis Induced by K-ras Mutation and NNK. Mol Ther Nucleic
Acids. 9:145–154. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
123
|
Wang M, Meng B and Liu Y, Yu J, Chen Q and
Liu Y: MiR-124 Inhibits Growth and Enhances Radiation-Induced
Apoptosis in Non-Small Cell Lung Cancer by Inhibiting STAT3. Cell
Physiol Biochem. 44:2017–2028. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
124
|
Zhang X, Gao F, Zhou L, Wang H, Shi G and
Tan X: UCA1 Regulates the Growth and Metastasis of Pancreatic
Cancer by Sponging miR-135a. Oncol Res. 25:1529–1541. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
125
|
Wang P, Zhang L, Chen Z and Meng Z:
MicroRNA targets autophagy in pancreatic cancer cells during cancer
therapy. Autophagy. 9:2171–2172. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
126
|
Ganley IG, Lam H, Wang J, Ding X, Chen S
and Jiang X: ULK1.ATG13.FIP200 complex mediates mTOR signaling and
is essential for autophagy. J Biol Chem. 284:12297–12305. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
127
|
Hosokawa N, Hara T, Kaizuka T, Kishi C,
Takamura A, Miura Y, Iemura S, Natsume T, Takehana K, Yamada N, et
al: Nutrient-dependent mTORC1 association with the
ULK1-Atg13-FIP200 complex required for autophagy. Mol Biol Cell.
20:1981–1991. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
128
|
Jung CH, Jun CB, Ro SH, Kim YM, Otto NM,
Cao J, Kundu M and Kim DH: ULK-Atg13-FIP200 complexes mediate mTOR
signaling to the autophagy machinery. Mol Biol Cell. 20:1992–2003.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
129
|
Yan J, Kuroyanagi H, Tomemori T, Okazaki
N, Asato K, Matsuda Y, Suzuki Y, Ohshima Y, Mitani S, Masuho Y, et
al: Mouse ULK2, a novel member of the UNC-51-like protein kinases:
Unique features of functional domains. Oncogene. 18:5850–5859.
1999. View Article : Google Scholar : PubMed/NCBI
|
|
130
|
Chan EY, Longatti A, McKnight NC and Tooze
SA: Kinase-inactivated ULK proteins inhibit autophagy via their
conserved C-terminal domains using an Atg13-independent mechanism.
Mol Cell Biol. 29:157–171. 2009. View Article : Google Scholar :
|
|
131
|
Kundu M, Lindsten T, Yang CY, Wu J, Zhao
F, Zhang J, Selak MA, Ney PA and Thompson CB: Ulk1 plays a critical
role in the autophagic clearance of mitochondria and ribosomes
during reticulocyte maturation. Blood. 112:1493–1502. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
132
|
Mo J, Zhang D and Yang R: MicroRNA-195
regulates proliferation, migration, angiogenesis and autophagy of
endothelial progenitor cells by targeting GABARAPL1. Biosci Rep.
36:362016. View Article : Google Scholar
|
|
133
|
Zang W, Wang Y, Wang T, Du Y, Chen X, Li M
and Zhao G: miR-663 attenuates tumor growth and invasiveness by
targeting eEF1A2 in pancreatic cancer. Mol Cancer. 14:372015.
View Article : Google Scholar : PubMed/NCBI
|
|
134
|
Huang W, Li J, Guo X, Zhao Y and Yuan X:
miR-663a inhibits hepatocellular carcinoma cell proliferation and
invasion by targeting HMGA2. Biomed Pharmacother. 81:431–438. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
135
|
Zhang Y, Xu X, Zhang M, Wang X, Bai X, Li
H, Kan L, Zhou Y, Niu H and He P: MicroRNA-663a is downregulated in
non-small cell lung cancer and inhibits proliferation and invasion
by targeting JunD. BMC Cancer. 16:3152016. View Article : Google Scholar : PubMed/NCBI
|
|
136
|
Kuroda K, Fukuda T, Krstic-Demonacos M,
Demonacos C, Okumura K, Isogai H, Hayashi M, Saito K and Isogai E:
miR-663a regulates growth of colon cancer cells, after
administration of antimicrobial peptides, by targeting CXCR4-p21
pathway. BMC Cancer. 17:332017. View Article : Google Scholar : PubMed/NCBI
|
|
137
|
Lin Y, Kuang W, Wu B, Xie C, Liu C and Tu
Z: IL-12 induces autophagy in human breast cancer cells through
AMPK and the PI3K/Akt pathway. Mol Med Rep. 16:4113–4118. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
138
|
Wu L, Zhang Q, Mo W, Feng J, Li S, Li J,
Liu T, Xu S, Wang W, Lu X, et al: Quercetin prevents hepatic
fibrosis by inhibiting hepatic stellate cell activation and
reducing autophagy via the TGF-β1/Smads and PI3K/Akt pathways. Sci
Rep. 7:92892017. View Article : Google Scholar
|
|
139
|
Ren L, Han W, Yang H, Sun F, Xu S, Hu S,
Zhang M, He X, Hua J and Peng S: Autophagy stimulated proliferation
of porcine PSCs might be regulated by the canonical Wnt signaling
pathway. Biochem Biophys Res Commun. 479:537–543. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
140
|
Lin R, Feng J, Dong S, Pan R, Zhuang H and
Ding Z: Regulation of autophagy of prostate cancer cells by
β-catenin signaling. Cell Physiol Biochem. 35:926–932. 2015.
View Article : Google Scholar
|
|
141
|
Zhao S, Li L, Wang S, Yu C, Xiao B, Lin L,
Cong W, Cheng J, Yang W, Sun W, et al: H2O2 treatment or serum
deprivation induces autophagy and apoptosis in naked mole-rat skin
fibroblasts by inhibiting the PI3K/Akt signaling pathway.
Oncotarget. 7:84839–84850. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
142
|
Lu X, Lv S, Mi Y, Wang L and Wang G:
Neuroprotective effect of miR-665 against sevoflurane
anesthesia-induced cognitive dysfunction in rats through PI3K/Akt
signaling pathway by targeting insulin-like growth factor 2. Am J
Transl Res. 9:1344–1356. 2017.PubMed/NCBI
|
|
143
|
Su N, Wang P and Li Y: Role of
Wnt/β-catenin pathway in inducing autophagy and apoptosis in
multiple myeloma cells. Oncol Lett. 12:4623–4629. 2016. View Article : Google Scholar
|
|
144
|
Kai Y, Qiang C, Xinxin P, Miaomiao Z and
Kuailu L: Decreased miR-154 expression and its clinical
significance in human colorectal cancer. World J Surg Oncol.
13:1952015. View Article : Google Scholar : PubMed/NCBI
|
|
145
|
Xu H, Fei D, Zong S and Fan Z:
MicroRNA-154 inhibits growth and invasion of breast cancer cells
through targeting E2F5. Am J Transl Res. 8:2620–2630.
2016.PubMed/NCBI
|
|
146
|
Liu ZQ, Zhao S and Fu WQ: Insulin-like
growth factor 1 antagonizes lumbar disc degeneration through
enhanced autophagy. Am J Transl Res. 8:4346–4353. 2016.PubMed/NCBI
|
|
147
|
Liu Q, Guan JZ, Sun Y, Le Z, Zhang P, Yu D
and Liu Y: Insulin-like growth factor 1 receptor-mediated cell
survival in hypoxia depends on the promotion of autophagy via
suppression of the PI3K/Akt/mTOR signaling pathway. Mol Med Rep.
15:2136–2142. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
148
|
Ceperuelo-Mallafré V, Ejarque M, Serena C,
Duran X, Montori-Grau M, Rodríguez MA, Yanes O, Núñez-Roa C, Roche
K, Puthanveetil P, et al: Adipose tissue glycogen accumulation is
associated with obesity-linked inflammation in humans. Mol Metab.
5:5–18. 2015. View Article : Google Scholar
|
|
149
|
Ciarcia R, Damiano S, Montagnaro S,
Pagnini U, Ruocco A, Caparrotti G, d’ Angelo D, Boffo S, Morales F,
Rizzolio F, et al: Combined effects of PI3K and SRC kinase
inhibitors with imatinib on intracellular calcium levels,
autophagy, and apoptosis in CML-PBL cells. Cell Cycle.
12:2839–2848. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
150
|
Hua R, Han S, Zhang N, Dai Q, Liu T and Li
J: cPKCγ-Modulated Sequential Reactivation of mTOR Inhibited
Autophagic Flux in Neurons Exposed to Oxygen Glucose
Deprivation/Reperfusion. Int J Mol Sci. 19:192018. View Article : Google Scholar
|
|
151
|
Shivaram S, Crist KA, Chaudhuri B, Mucci
SJ and Chaudhuri PK: Effect of CCK receptor antagonist on growth of
pancreatic adenocarcinoma. J Surg Res. 53:234–237. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
152
|
Clerc P, Saillan-Barreau C, Desbois C,
Pradayrol L, Fourmy D and Dufresne M: Transgenic mice expressing
cholecystokinin 2 receptors in the pancreas. Pharmacol Toxicol.
91:321–326. 2002. View Article : Google Scholar
|
|
153
|
Shang C, Zhu W, Liu T, Wang W, Huang G,
Huang J, Zhao P, Zhao Y and Yao S: Characterization of long
non-coding RNA expression profiles in lymph node metastasis of
early-stage cervical cancer. Oncol Rep. 35:3185–3197. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
154
|
Bronietzki AW, Schuster M and Schmitz I:
Autophagy in T-cell development, activation and differentiation.
Immunol Cell Biol. 93:25–34. 2015. View Article : Google Scholar
|
|
155
|
Martinez-Lopez N, Athonvarangkul D and
Singh R: Autophagy and aging. Adv Exp Med Biol. 847:73–87. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
156
|
Tan Y, Gong Y, Dong M, Pei Z and Ren J:
Role of autophagy in inherited metabolic and endocrine myopathies.
Biochim Biophys Acta Mol Basis Dis. 1865.48–55. 2018.
|
|
157
|
Chu CT: Autophagy in Neurological
Diseases: An update. Neurobiol Dis. S0969–9961(18): 30728–9.
2018.
|
|
158
|
Yang A, Rajeshkumar NV, Wang X, Yabuuchi
S, Alexander BM, Chu GC, Von Hoff DD, Maitra A and Kimmelman AC:
Autophagy is critical for pancreatic tumor growth and progression
in tumors with p53 alterations. Cancer Discov. 4:905–913. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
159
|
Zhan Y, Wang K, Li Q, Zou Y, Chen B, Gong
Q, Ho HI, Yin T, Zhang F, Lu Y, et al: The Novel Autophagy
Inhibitor Alpha-Hederin Promoted Paclitaxel Cytotoxicity by
Increasing Reactive Oxygen Species Accumulation in Non-Small Cell
Lung Cancer Cells. Int J Mol Sci. 19:192018. View Article : Google Scholar
|
|
160
|
Chen S, Wu J, Jiao K, Wu Q, Ma J, Chen D,
Kang J, Zhao G, Shi Y, Fan D, et al: MicroRNA-495-3p inhibits
multidrug resistance by modulating autophagy through GRP78/mTOR
axis in gastric cancer. Cell Death Dis. 9:10702018. View Article : Google Scholar : PubMed/NCBI
|
|
161
|
Verma SP and Das P: Monensin induces cell
death by autophagy and inhibits matrix metalloproteinase 7 (MMP7)
in UOK146 renal cell carcinoma cell line. In Vitro Cell Dev Biol
Anim. 54:736–742. 2018. View Article : Google Scholar : PubMed/NCBI
|