Introduction
Cervical cancer ranks as the third most commonly
diagnosed cancer and is the second leading cause of
cancer-associated mortality in females worldwide (1). Despite improvements in screening and
diagnostic techniques, regional or local relapse, and lymph node
metastasis remain the most prevalent causes of cervical
cancer-related mortality (2).
Similarly, the motility and invasiveness of cancer cells serve a
critical role in the mortality of patients with cervical cancer.
Therefore, advances in understanding the mechanism of metastasis in
cervical cancer may provide insight for developing effective
approaches to prevent and treat cervical cancer.
S100A9, known as Calgranulin B or MRP-14, is a
member of the low-molecular calcium binding S100 protein family. In
our previous study, we demonstrated that exogenous S100A9 promoted
the proliferation and invasion of HepG2 cells, which was partly
mediated by activation of the MAPK signaling pathway (3) and that the underlying mechanism may
be dependent on receptor of advanced glycation end products
(RAGE)/MAPK signaling cascades (4). Interestingly, increasing evidence
indicates that S100A9 is overexpressed in cervical cancer (5,6) and
could be a candidate marker for the early diagnosis of cervical
cancer and a potential therapeutic target for therapy(5). few studies have investigated the
precise role of S100A9 in the tumorigenesis and development of
cervical cancer, and the underlying potential molecular
mechanism.
Epithelial-mesenchymal transition (EMT) is a vital
process associated with the pathogenesis of numerous cancers,
including cervical cancer. This transition is characterized by a
process in which epithelial cells lose their polarity enabling them
to assume a mesenchymal cell phenotype, which imparts cancer cells
several properties, such as tumor invasion, carcinoma metastasis
and chemotherapy resistance (7). A
previous study demonstrated that cellular RAGE binding to S100A8/A9
can cause a notable increase and accumulation of endogenous Snail
in the nucleus of breast cancer cells; thus, EMT and cancer
metastasis are enhanced via the nuclear factor (NF)-κB signaling
module (8). Additionally, the
effects of transforming growth factor β1 on EMT were antagonized by
S100A9 via the formation of heterodimers between them in pancreatic
cancer; however, the effects of S100A9 on EMT in cervical cancer
require further investigation.
Recent studies reported that cytoplasmic/nuclear
expression of β-catenin was associated with the malignant phenotype
and the pathogenesis of cervical cancer (9,10).
In addition, it was indicated that the Wnt/β-catenin pathway and
EMT may act synergistically in the progression of cervical cancer
(11). Tumor cells exhibiting
β-catenin accumulation in the nucleus appear to promote EMT, which
involves the loss of epithelial markers and the expression of
mesenchymal markers (12). Members
of the S100 protein family, including S100A6, S100B, and S100P were
reported to negatively regulate β-catenin degradation. Our
consensus finding is that recombinant S100A8 and S100A9 proteins
could promote the accumulation of β-catenin, and upregulate mRNA
expression of the target genes of the Wnt/β-catenin signaling
pathway in colorectal carcinoma (13). However, whether S100A9 regulates
the Wnt/β-catenin pathway, thereby exerting its effects on the
progression of cervical cancer remains unknown.
In the present study, the functional role of S100A9
in the Wnt/β-catenin signaling pathway and EMT was investigated to
elucidate how S100A9 modulates the progression of cervical cancer.
We reported that S100A9 promoted the proliferation, migration, and
EMT of cervical cancer cells, and the activation of the
Wnt/β-catenin signaling pathway may be involved in the modulation
of these processes. These findings provide novel insight to the
importance of S100A9 in the progression of cervical cancer and a
basis for the development of targeted therapeutic strategies for
treating cervical cancer.
Materials and methods
Cell culture
The human embryonic kidney cell line 293 and the
human cervical cancer cell lines SiHa, CaSki, and HeLa were
obtained from the American Type Culture Collection. All cells were
maintained in Dulbecco's modified Eagle's medium (DMEM)
supplemented with fetal bovine serum (FBS; 10%; both obtained from
Gibco; Thermo Fisher Scientific, Inc.), 100 U/ml penicillin and 100
μg/ml streptomycin (both obtained from HyClone; GE
Healthcare) at 37°C in a humidified atmosphere of 5%
CO2. 293 cells were used for adenovirus amplifica
tion.
Reagents and antibodies
Adenovirus expressing S100A9 and green fluorescent
protein (AdS100A9), adenovirus expressing small interfering (si)RNA
targeting β-catenin and red fluorescent protein (Adsiβ-catenin),
control adenovirus expressing green fluorescent protein (AdGFP) and
red fluorescent protein (AdRFP), and pGST-moluc and
pGST-moluc-hS100A9 plasmids were kindly donated by Professor T.C.
He (The University of Chicago Medical Center, Chicago, IL, USA).
Competent bacteria E. coli (BL21) were saved in our
laboratory. Adenoviruses expressing siRNA targeting S100A9 and red
fluorescent protein (AdsiS100A9), and control adenoviruses
expressing red fluorescent protein (AdsiControl) were constructed
in house. The kit used for semi-quantitative PCR was purchased from
Takara Bio, Inc. Antibodies, including mouse anti-β-actin,
anti-vimentin and anti-β-catenin were purchased from Santa Cruz
Biotechnology, Inc. (cat. nos. sc-47778, sc-66001 and sc-59737).
Rabbit anti-S100A9 antibody was purchased from Abcam (cat. no.
ab92507). Rabbit anti-E-cadherin antibody was purchased from
ImmunoWay (cat. no. YM3353, Plano). Rabbit anti-histone H3 antibody
was purchased from Abmart (cat. no. P30266). Secondary antibody
reagents, such as goat anti-mouse IgG serum and goat anti-rabbit
IgG serum were obtained from Beijing Zhongshan Golden Bridge
Biotechnology (cat. no. 2305 and no. 2301). Western blot reagents
and radioimmunoprecipitation assay (RIPA) buffer were purchased
from Beyotime Institute of Biotechnology. Phosphatase and protease
inhibitors were purchased from Roche Diagnostics GmbH.
Polyvinylidene difluoride (PVDF) membranes and an enhanced
chemiluminescence (ECL) kit were purchased from EMD Millipore.
Adenovirus infection
HeLa cells were infected with AdS100A9 and AdGFP,
whereas SiHa cells were infected with AdsiS100A9 and AdsiControl.
After 8-12 h of incubation, the medium was replaced with complete
medium containing FBS followed by continued cell culture for
subsequent experiments. The cells were maintained at 37°C in a
humidified atmosphere of 5% CO2.
Recombinant protein preparation
The pGST-moluc and pGST-moluc-hS100A9 plasmids used
in the present study has been described previously (4). In brief, pGST-moluc and
pGST-moluc-hS100A9 was transfected into E. coli (BL21) by
calcium chloride-mediated transformation.
Isopropylthio-β-D-galactoside was used to induce the expression of
GST and GST-hS100A9 proteins. The bacteria were then collected and
sonicated on ice at 4°C. The supernatants were incubated with
glutathione-sepharose 4B beads, GST and GST-hS100A9 proteins on the
beads were eluted by elution buffer with reduced glutathione on
ice. Finally the GST and GST-hS100A9 proteins were filtered and
stored at −80°C. Cells were treated with 20 μg/ml of
recombinant proteins for 48 h at 37°C.
RNA extraction and semi-quantitative
polymerase chain reaction (PCR) analysis
Total RNA was extracted from cells using
TRIzol® reagent (Invitrogen; Thermo Fisher Scientific,
Inc.), and cDNA samples were synthesized using random primers and a
Reverse Transcriptase PCR kit (Takara Bio, Inc.) according to the
manufacturer's protocols. cDNA was used as a template for
semi-quantitative PCR. The thermocycling conditions were as
follows: 94°C for 5 min, 94°C for 30 sec, 68°C for 30 sec and 72°C
for 12 cycles with a decrease in 1°C/cycle; then, 94°C for 30 sec,
55°C for 30 sec, and 72°C for 30 sec for 18-27 cycles depending on
the abundance of the target genes. The PCR products were identified
by electrophoresis using 1.5% agarose gels. GAPDH was used as an
internal reference control. The results were recorded using Gel Doc
1000 imaging system and Quantity One version 4.5.0 software
(Bio-Rad Laboratories, Inc.). The primers used in this study are
shown in Table I.
 | Table IPrimers employed in the present
study. |
Table I
Primers employed in the present
study.
| Gene | Primer
sequences | Product size
(bp) |
|---|
| S100A9 | Forward:
5′-ACCCAGACACCCTGAACC-3′ | |
| Reverse:
5′-AGCATGATGAACTCCTCGA-3′ | 163 |
| c-Myc | Forward:
5′-TACCCTCTCAACGACAGCAG-3′ | |
| Reverse:
5′-TCTTGACATTCTCCTCGGTG-3′ | 478 |
| Snail | Forward:
5′-ACCCCACATCCTTCTCACTG-3′ | |
| Reverse:
5′-TACAAAAACCCACGCAGACA-3′ | 217 |
| Twist | Forward:
5′-TCTTACGAGGAGCTGCAGAC-3′ | |
| Reverse:
5′-TATCCAGCTCCAGAGTCTCT-3′ | 406 |
| GAPDH | Forward:
5′-CAGCGACACCCACTCCTC-3′ | |
| Reverse:
5′-TGAGGTCCACCACCCTGT-3′ | 120 |
Western blot analysis
Total cellular protein of cervical cancer cells were
prepared using RIPA buffer containing phosphatase and protease
inhibitors. Nuclear and cytoplasmic proteins were extracted using
Nuclear-Cytosol Extraction Kit (cat. no. KGP1100, Nanjing KeyGen
Biotech Co., Ltd.) according to the manufacturer's instructions.
Protein extracts were separated by 6-15% SDS-PAGE and transferred
to PVDF membranes. The membranes were blocked with 5% bovine serum
albumin at 37°C for 2 h, washed with Tris-buffered saline with
Tween-20 (TBST) and incubated with primary antibodies against
S100A9, vimentin, E-cadherin, β-catenin, histone H3, and β-actin
separately (1:1,000 dilution) at 4°C overnight. After washing with
TBST, the membranes were incubated with the a secondary antibody
conjugated with horseradish peroxidase (1:5,000 dilution) for 1 h
at 37°C. Finally, the blots were washed with TBST, and then
visualized using the Bio-Rad Gel Doc 1000 imaging system and
analyzed by Quantity One Version 4.5.0.
Cell proliferation assay
Cell proliferation was analyzed via an MTT assay
(Promega Corporation). A total of 3.5×103 HeLa cells or
4.5×103 SiHa cells were seeded in 96-well flat-bottomed
microplates. After infection with or without adenoviruses in DMEM
containing 1% FBS for 24, 48 and 72 h, 10 μl of MTT reagent
was added to each well and incubated at 37°C for 4 h. Dimethyl
sulfoxide (100 μl) was added to dissolve the formazan
crystals for 10 min at room temperature. Finally, the absorbance
was measured at 490 nm using an automatic enzyme immunoassay
analyzer (Bio-Rad Laboratories, Inc.). The experiments were
performed five times in three independent experimental trials.
Wound healing assay
Cervical cancer cells were seeded in 6-well plates
and incubated at 37°C until the cells attained 90% confluence. The
cells were then infected with adenoviruses. Cell monolayers were
carefully wounded by scratching with a sterile 10-μl pipette
tip. The cells were then washed twice with cold PBS and incubated
in serum-free DMEM at 37°C. Wound areas were observed and imaged at
×100 magnification at 0, 24 and 36 h with an inverted fluorescent
microscope (Nikon eclipse 80i; Nikon Corporation). Overall, the
experiments were repeated three times.
Transwell migration assay
A chamber of non-type I-collagen- coated 24-well
Transwell cell culture inserts (EMD Millipore) was used for the
Transwell migration assay. Briefly, after infection with or without
adenoviruses and recombination proteins for various durations, the
cells were trypsinized, washed, and suspended in 400 μl of
serum-free DMEM and finally seeded in the upper chamber (HeLa,
4×104 cells per well; SiHa, 6×104 cells per
well). The lower chamber was coated with 700 μl of DMEM
containing 20% FBS as a chemoattractant. After 24 h, cells were
fixed with methanol for 20 min and stained with 0.05% crystal
violet for 30 min at room temperature. Cells on the upper surface
of the insert membrane were removed with cotton swabs. Finally, the
cells were counted using an inverted microscope (magnification,
×100) in five randomly selected fields for each well. The
experiment was repeated three times.
Immunofluorescence
The cells were seeded on sterile glass coverslips in
24-well culture plates. After treatment with or without AdS100A9,
cells were washed with PBS, fixed with 4% paraformaldehyde for 30
min and permeabilized with 0.3% Triton X-100 at 37°C for 30 min.
After blocking with goat serum at 37°C for 30 min, the cells were
incu- bated with antibodies against E-cadherin, N-cadherin (1:100
dilution in PBS) at 4°C overnight. Subsequently, slides were washed
with PBS and incubated with fluorescein isothiocyanate-conjugated
secondary antibodies (Beyotime Institute of Biotechnology) for 1 h
at room temperature in the dark. Then, the cells were washed with
PBS, counter stained with DAPI (Beyotime Institute of
Biotechnology) for 5 min at room temperature, washed with PBS again
and then the coverslips were mounted using antifade mounting
medium. The fluorescence of the various groups was visualized
imaged with a Nikon eclipse 80i inverted fluorescent microscope
(Nikon Corporation).
Statistical analysis
All data are presented as the mean ± standard
deviation. Differences were analyzed using one-way ANOVA followed
by a Tukey's multiple comparison test. All statistical analyses
were performed using SPSS statistical package 17.0 (SPSS, Inc.,
Chicago, IL, USA) and GraphPad Prism 5 (GraphPad Software, Inc.).
P≤0.05 was considered to indicate a statistically significant
difference.
Results
Expression of endogenous S100A9 in
cervical cancer cells
We first evaluated the endogenous expression of
S100A9 in SiHa, CaSki, and HeLa cell lines using semi-quantitative
PCR and western blot analysis. As shown in Fig. 1A, the mRNA expression levels of
S100A9 in HeLa cells were significantly reduced than in SiHa and
CaSki cells (P≤0.001 and P≤0.01). These results were confirmed by
western blot analysis (P≤0.01; Fig.
1B). Subsequently, semi-quantitative PCR and western blot
analysis (Fig. 1C and D) were
performed to detect the expression of S100A9 in HeLa and SiHa cells
infected with AdS100A9 and AdsiS100A9 for 48 h, respectively. The
expression of S100A9 observed in the AdS100A9 group was
significantly increased compared with the AdGFP and Blank groups in
HeLa cells (P≤0.001 and P≤0.05), whereas a significant decrease in
S100A9 expression was detected in the AdsiS100A9 group compared
with AdsiControl and Blank groups in SiHa cells (P≤0.05 and
P≤0.01). These results indicated that S100A9 was successfully
overexpressed in HeLa cells and downregulated in SiHa cells via
infection with AdS100A9 and AdsiS100A9, respectively.
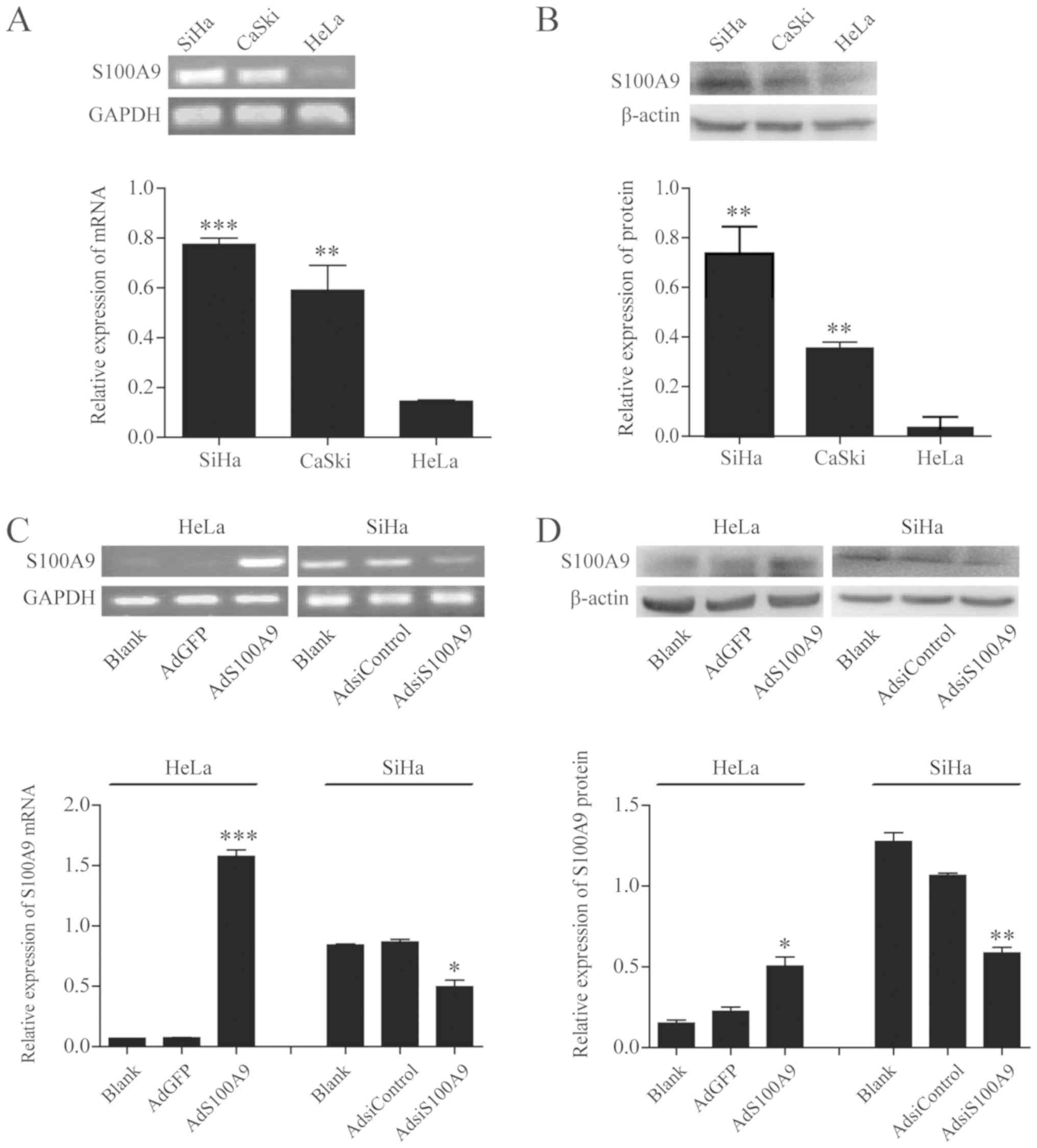 | Figure 1S100A9 is successfully overexpressed
in HeLa cells and downregulated in SiHa cells via adenoviral
infection. (A) mRNA expression of endogenous S100A9 in SiHa, CaSki
and HeLa cell lines was detected by semi-quantitative PCR analysis.
***P≤0.001, HeLa vs. SiHa and **P≤0.01, HeLa
vs. CaSki. (B) Protein expression of endogenous S100A9 in SiHa,
CaSki and HeLa cell lines was detected by western blot analysis.
**P≤0.01, HeLa vs. SiHa and CaSki. HeLa cells were
infected with AdGFP and AdS100A9 for 48 h, whereas SiHa cells were
infected with AdsiControl and AdsiS100A9 for 48 h. The mRNA and
protein expression of S100A9 in cervical carcinoma cell lines was
detected by (C) semi-quantitative PCR and (D) western blot
analysis. Data are presented as the mean ± standard deviation of
three independent experiments. ***P≤0.001,
**P≤0.01 and *P≤0.05, AdS100A9 vs. AdGFP and
AdsiControl. No significant difference between the
AdGFP/AdsiControl and Blank groups of HeLa and SiHa cell lines were
observed (P>0.05). Ad, adenovirus; Adsi, adenovirus containing
small interfering RNA; GFP, green fluorescent protein; PCR,
polymerase chain reaction. |
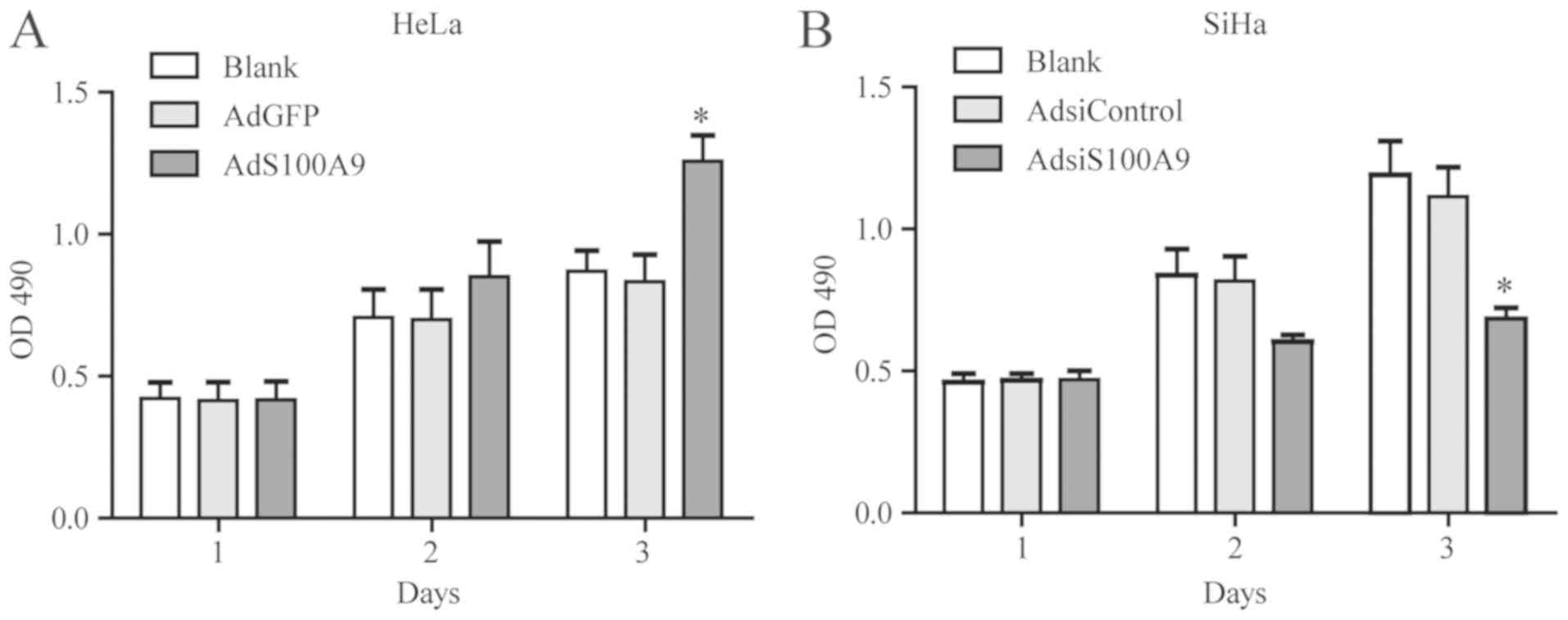
S100A9 promotes the proliferation of
cervical cancer cells
To investigate the effects of S100A9 on the
proliferation of cervical cancer cells, we treated the cells with
AdS100A9 and AdsiS100A9 for 3 days and examined by an MTT assay
(Fig. 2). Compared with the
control groups, the proliferation of HeLa cells was significantly
enhanced in the S100A9-overexpressing group on day 3 (P≤0.05;
Fig. 2A). On the contrary, the
proliferation of SiHa cells was significantly inhibited in the
S100A9-knockdown group compared with the control (P≤0.05; Fig. 2B). These results demonstrated that
overexpressed S100A9 could promote the proliferative potential of
cervical cancer cells.
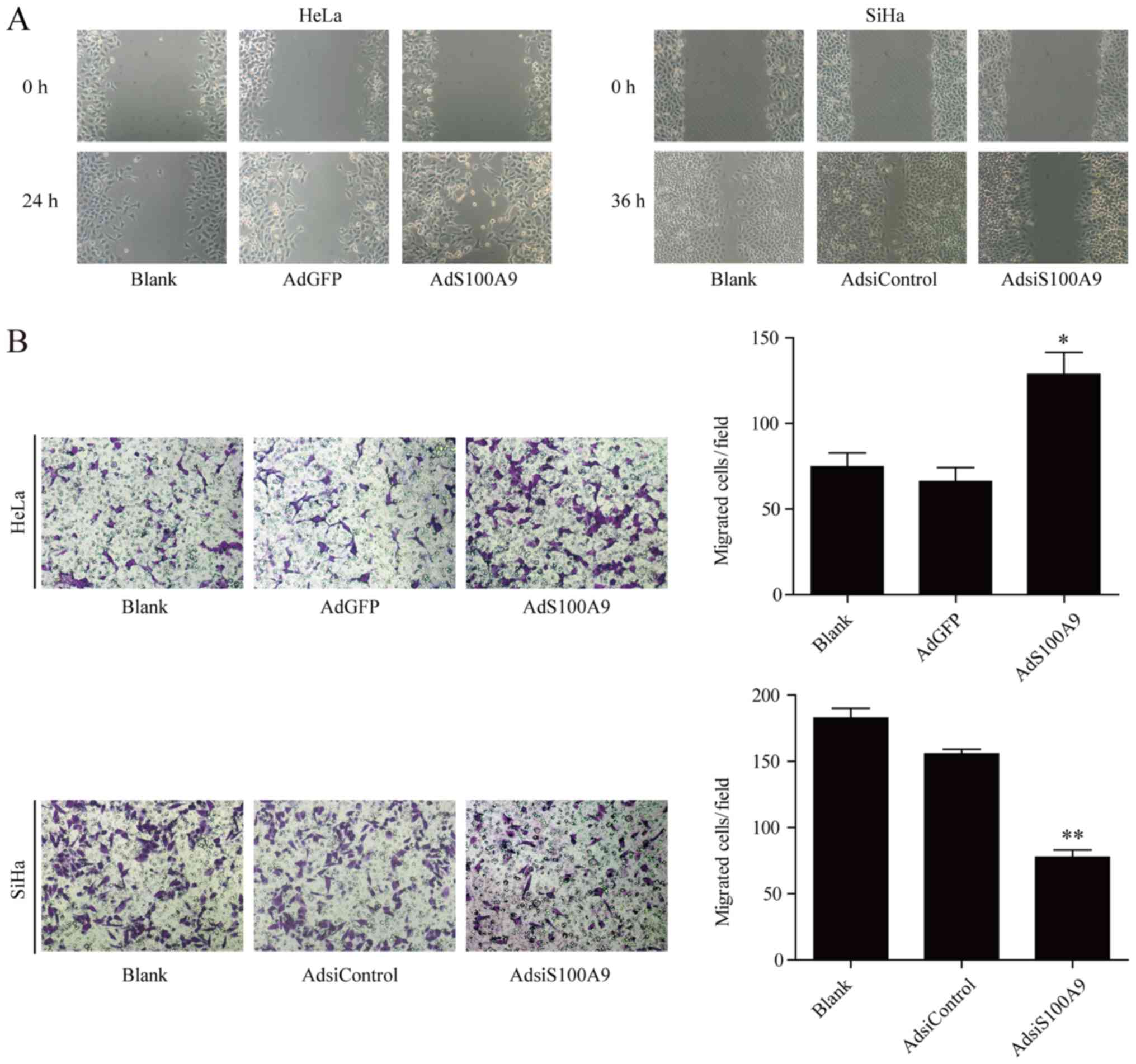
S100A9 promotes the migration of cervical
cancer cells
We further studied the role of S100A9 in the
migration of cervical cancer cells by wound healing and Transwell
migration assays. The results revealed that the wound closure rate
of S100A9-overexpressing HeLa cells was notably increased than the
control group, whereas that of SiHa cells was impaired following
knockdown of S100A9 (Fig. 3A).
Similar results were observed in the Transwell migration assay. The
number of HeLa cells migrated across the membrane in AdS100A9 group
was significantly higher than that in the AdGFP group (P≤0.05;
Fig. 3B). These results are
consistent with the wound healing ability of SiHa cells. The number
of S100A9-depleted SiHa cells that migrated across the membrane was
significantly lower compared with the control (P≤0.01).
Collectively, these results suggested that upregulated S100A9
expression may facilitate the migration of cervical cancer
cells.
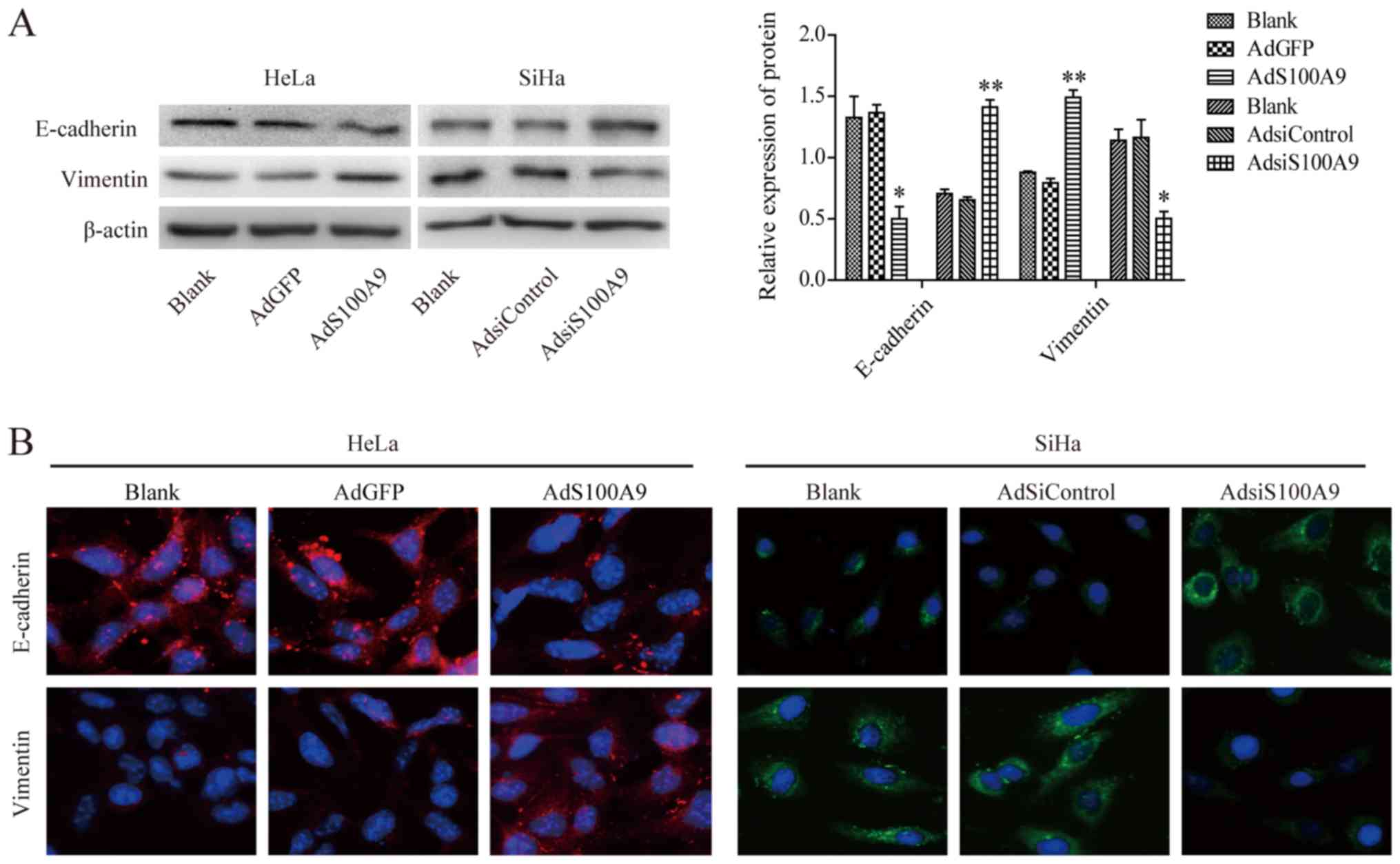
S100A9 induces EMT in cervical cancer
cells
Emerging evidences support that EMT has a crucial
role in the metastasis of primary tumors. To evaluate whether
S100A9 promoted EMT in cervical cancer cells, western blot and
immunofluorescence analyses were conducted to detect the levels of
E-cadherin and vimentin. As presented in Fig. 4A, overexpression of S100A9 in HeLa
cells significantly decreased the expression of E-cadherin
(P≤0.05), and increased that of of vimentin (P≤0.01). On the
contrary, knockdown of S100A9 in SiHa cells significantly decreased
the expression of vimentin (P≤0.01), and increased that of
E-cadherin (P≤0.05). These results were further confirmed by
immunofluorescence assays (Fig.
4B). These findings suggested that S100A9 is involved in the
regulation of EMT-related protein expression, and may act as a
predictive factor and therapeutic target for preventing metastasis
in cervical cancer.
S100A9 promotes the activation of the
Wnt/β-catenin signaling pathway in cervical cancer cells
It is well known that aberrant activation of the
Wnt/β-catenin pathway serves a critical role in the development of
cervical cancer. To investigate whether S100A9 was involved in the
regulation of Wnt/β-catenin pathway in cervical cancer cells,
western blot and semi-quantitative PCR analyses were performed to
study the effects of S100A9 on the expression of β-catenin protein,
and the transcriptional levels of c-Myc, Snail, and twist-related
protein-1 (Twist), which are the classical target genes of
Wnt/β-catenin pathway. As shown in Fig. 5A, after infecting HeLa cells with
AdS100A9, overexpression of S100A9 significantly enhanced the
accumulation of total β-catenin and nuclear β-catenin (P≤0.01), but
knockdown of S100A9 via the infection of SiHa cells with AdsiS100A9
decreased the accumulation of total β-catenin and nuclear β-catenin
compared with the control groups (P≤0.001 and P≤0.05).
Additionally, overexpression of S100A9 caused by infecting HeLa
cells with AdS100A9 significantly promoted the transcription of
c-Myc (P≤0.05), Snail (P≤0.01), and Twist (P≤0.05) compared with
the control. Conversely, the mRNA levels of the aforementioned
target genes of the Wnt/β-catenin signaling pathway were
significantly reduced when SiHa cells were infected with AdsiS100A9
compared with the control (P≤0.01, P≤0.01 and P≤0.001,
respectively; Fig. 5B). These
results demonstrated that S100A9-induced upregulation of c-Myc,
Snail, and Twist expression in cervical cancer cell lines may be
associated with the accumulation of total β-catenin and nuclear
β-catenin.
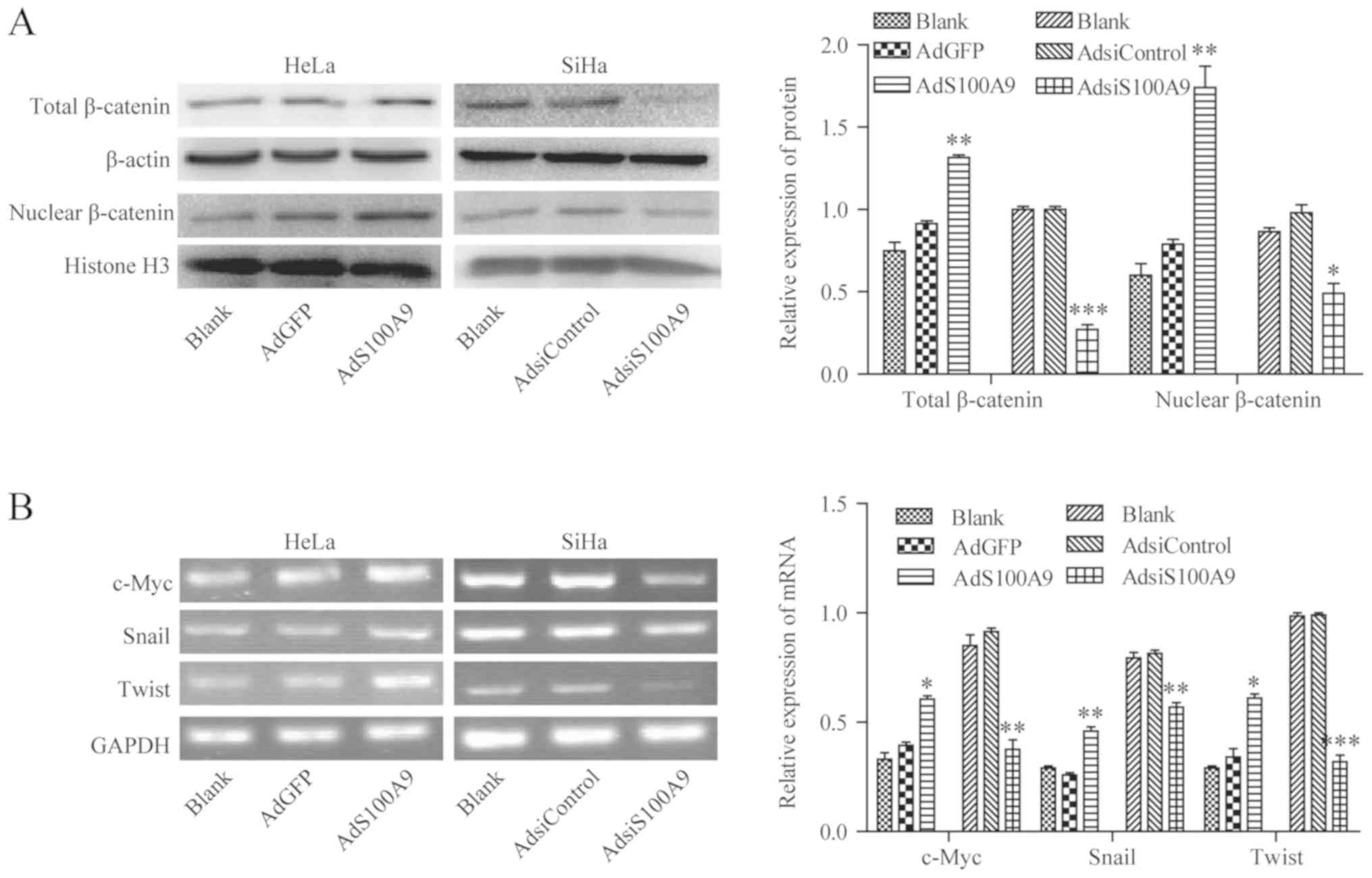 | Figure 5S100A9 activates the Wnt/β-catenin
signaling pathway in cervical cancer cells. (A) After HeLa and SiHa
cells were infected with AdS100A9 or AdsiS100A9, the accumulation
of total and nuclear β-catenin was measured by western blot
analysis. β-actin and histone were used as internal reference
controls. **P≤0.01, AdS100A9 vs. AdGFP;
***P≤0.001, *P≤0.05, AdsiS100A9 vs.
AdsiControl. (B) After infecting HeLa and SiHa cells with AdS100A9
or AdsiS100A9, the mRNA expression of c-Myc, Snail, and Twist was
detected using semi-quantitative polymerase chain reaction. GAPDH
was used as an internal reference control. *P≤0.05,
**P≤0.01, ***P≤0.001, AdS100A9 vs. AdGFP and
AdsiControl. Data are presented as mean ± standard deviation of
three individual measurements. Ad, adenovirus; Adsi, adenovirus
containing small interfering RNA; Twist, twist-related
protein-1. |
S100A9-induced promotion of
proliferation, migration and EMT in cervical cancer cells via
activation of the Wnt/β-catenin pathway
To determine whether Wnt/β-catenin pathway is
involved in S100A9-induced promotion of proliferation, migration
and EMT in cervical cancer cells, adenovirus Adsiβ-catenin was used
as the antagonist to impair the activation of the Wnt/β-catenin
pathway. As presented in Fig. 6A,
the effects of S100A9 on the proliferation of HeLa cells was
investigated by an MTT assay. After the treatment of cells with
GST-hS100A9 at 10 and 20 μg/ml for 72 h, the proliferation
of HeLa cells was significantly increased than that of the control
groups (P≤0.05). Additionally, the results further showed a more
notable effect in response to high concentration recombinant
protein; thus, we selected 20 μg/ml of recombinant protein
for the following experiments. As shown in Fig. 6B, β-catenin expression was
significantly downregulated by adenovirus Adsiβ-catenin in HeLa
cells compared with the control (P≤0.05). After treatment with
GST-hS100A9 and adenovirus Adsiβ-catenin, the mRNA levels of Snail,
and Twist were significantly reduced in HeLa cells compared with
the control groups (P≤0.05 and P≤0.01; Fig. 6C). In addition, treatment with
Adsiβ-catenin significantly inhibited the proliferation and
migration of HeLa cells induced by GST-hS100A9 (P≤0.05 and P≤0.01;
Fig. 6D and E). Furthermore, the
expression of EMT markers was examined in HeLa cells after
treatment with and without AdRFP, Adsiβ-catenin, GST and
GST-hS100A9. We observed that downregulation of β-catenin
significantly suppressed the effects of S100A9 on the expression of
E-cadherin and vimentin compared with the control groups (P≤0.05
and P≤0.01; Fig. 6F). These
findings demonstrated that S100A9 induced promotion of
proliferation, migration and EMT through activation of
Wnt/β-catenin pathway in cervical cancer cells.
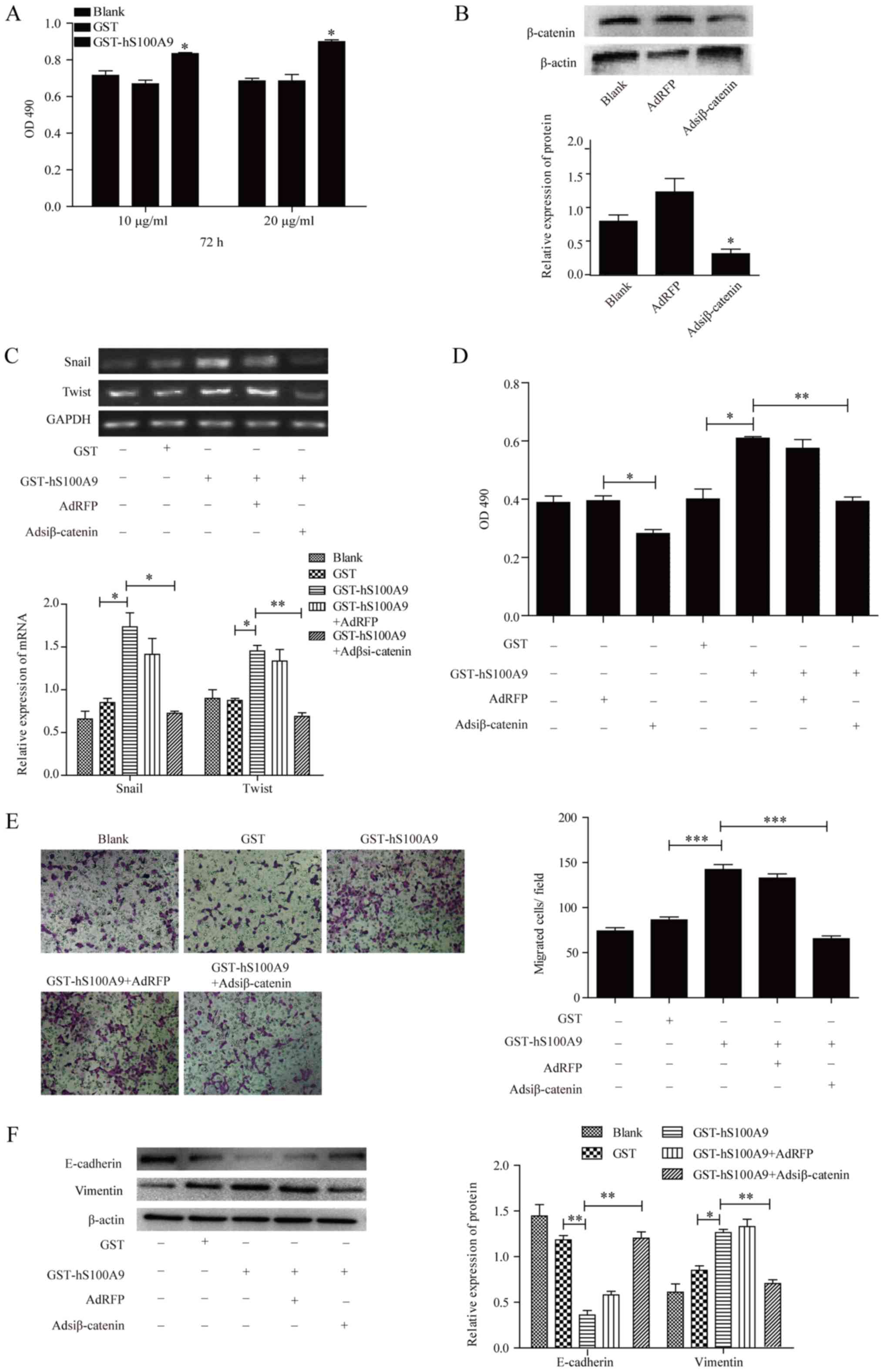 | Figure 6Underlying molecular mechanisms of
S100A9-induced promotion of proliferation, migration and EMT in
cervical cancer cells. (A) After treatment with or without
GST-hS100A9 at different concentrations for 72 h, the proliferation
of HeLa cells was measured by an MTT assay. *P≤0.05,
GST-hS100A9 vs. GST control. (B) Adsiβ-catenin-mediated β-catenin
knockdown in HeLa cells. After infection with Adsiβ-catenin, the
expression of β-catenin was analyzed by western blot analysis.
β-actin was used as internal reference control. *P≤0.05,
Adsiβ-catenin vs. AdRFP. After infection with Adsiβ-catenin for 24
h, the infected cells were treated with or without GST-hS100A9 at
20 μg/ml for 48 h. The mRNA levels of Snail, and Twist were
measured by (C) semi-quantitative polymerase chain reaction and the
proliferation of HeLa cells was measured by (D) an MTT assay. GAPDH
was used as an internal reference control. *P≤0.05, GST
vs. GST-hS100A9; **P≤0.01, GST-hS100A9 vs.GST-hS100A9 +
Adsiβ-catenin. (E) Transwell migration assay for analyzing the
migration of HeLa cells that were infected with Adsiβ-catenin for
24 h and treated with or without GST-hS100A9 at 20 μg/ml for
48 h. Representative images of transmembrane cells are shown. The
mean of transmembrane cells ± standard deviation per microscopic
field of three independent experiments was calculated.
Magnification, ×100. ***P≤0.001, GST vs. GST-hS100A9,
GST-hS100A9 + AdRFP vs. GST-hS100A9 + Adsiβ-catenin. (F) Western
blot for analyzing the expression of EMT markers of HeLa cells that
were infected with Adsiβ-catenin for 24 h and treated with and
without GST-hS100A9 at 20 μg/ml for 48 h. β-actin was used
as internal reference control. Data are presented as mean ±
standard deviation of three individual measurements.
*P≤0.05, **P≤0.01, GST vs. GST-hS100A9,
GST-hS100A9 + AdRFP vs. GST-hS100A9 + Adsiβ-catenin. Ad,
adenovirus; Adsi, adenovirus containing small interfering RNA; EMT,
epithelial-mesenchymal transition; GST, glutathione S-transferase;
OD, optical density; RFP, red fluorescent protein. |
Discussion
Increasing evidence has indicated that the enhanced
motility and invasiveness of cancer cells are associated with the
progression of cervical cancer (14-16).
In the present study, we observed that overexpression of S100A9
promoted the proliferation and migration of cervical cancer cells,
eventually facilitating EMT, which was regulated by the
Wnt/β-catenin signaling pathway. These findings suggested that
S100A9 may be an important carcinogenic factor in the occurrence
and progression of cervical cancer and may serve as an attractive
therapeutic target for cervical cancer driven by various signaling
pathways.
S100 proteins participate in a broad range of
intracellular and extracellular functions by regulating calcium
balance, cell apoptosis, migration, proliferation, differentiation,
energy metabolism and inflammation (17). Overexpressed S100 proteins,
including S100A9, were detected in a variety of cancers, including
pancreatic, liver, and colorectal carcinomas (4,13,18).
Compared with adjacent normal cervical tissues, increased levels of
S100A9 were detected in squamous cervical cancer by two-dimensional
gel electrophoresis followed by mass spectrometry, and the
expression of S100A9 and RAGE in squamous cervical cancer was
closely associated with histological differentiation and the tumor
progression (6). Furthermore,
various studies have shown that S100A9 could promote the
proliferation and migration of different cell types, such as
fibroblasts (19), MCF-7,
MDA-MB-231, SH-EP, Kelly (20),
HepG2 (4) and PNT1A cells
(21). In accordance with these
studies, our results suggested that overexpression of S100A9
enhanced the proliferation and migration properties of HeLa and
SiHa cells.
EMT is required for several complex differentiation
processes, including embryonic development, wound healing and tumor
progression (22,23). Interestingly, Li et al
(24) reported that S100A6 could
facilitate the metastatic ability and EMT of cervical cancer cells,
which was mediated by activating the PI3K/Akt signaling pathway.
Additionally, S100A14 was determined to be a mediator of EMT that
regulated the proliferation, migration and invasion of human
cervical cancer cells (25). Based
on these findings, we propose that overexpression of S100A9
resulted in a decrease in E-cadherin and an increase in vimentin
expression in cervical cancer cells. Conversely, knockdown of
S100A9 exhibited an antagonistic effect on the regulation of
E-Cadherin and vimentin. These results suggested that S100A9 could
enhance the mesenchymal properties of cervical cancer cells, which
may be attributed to the induction of EMT.
The pivotal role of Wnt/β-catenin signaling pathway
in tumor progression has been generally accepted, and cervical
cancer has been linked with the aberrant activation of the
Wnt/β-catenin pathway (22,26).
In the present study, we reported that S100A9 enhanced the
accumulation of β-catenin, and upregulated the mRNA expression of
the target genes c-Myc, Snail, and Twist in cervical cancer cells.
The results suggested that S100A9 may exert significant effects on
the induction of Wnt/β-catenin pathway in cervical cancer cells. In
addition, previous studies have shown that β-catenin upregulation
was involved in the proliferation and migration of cancer cells
(27,28). Our results revealed that
Wnt/β-catenin pathway was responsible for S100A9-induced promotion
cervical cancer cell proliferation and migration, which could be
partially suppressed by downregulation of β-catenin. Furthermore,
we observed that activation of the Wnt/β-catenin pathway may be
responsible for S100A9-induced EMT of cervical cancer cells, which
could be partially suppressed by downregulating β-catenin. These
findings suggested that S100A9 could induce EMT in cervical cancer
cells via the Wnt/β-catenin signaling pathway.
Collectively, the present study revealed that
S100A9-induced increases in cervical cancer cell proliferation and
migration may be attributed to the promotion of EMT and aberrant
activation of the Wnt/β-catenin signaling pathway. Additionally,
S100A9 may be considered as a novel molecular target for the
treatment of cervical cancer in future.
Funding
This work was supported by National Natural Science
Foundation grants of China (grant no. 81760475).
Availability of data and materials
The datasets used and/or analyzed during the present
study are available from the corresponding author on request.
Authors' contributions
HZ and XL performed most experiments and were major
contributors in writing the manuscript. SY, HL, AL, YG, JZ and JX
prepared experimental materials and reviewed the article. HS and LD
made substantial contributions to the design of the study, and
drafted the manuscript and revising it critically for important
intellectual content. LZ gave final approval of the version to be
published. All authors read and approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interest.
Acknowledgments
We would like to thank T.C He (Medical Center, The
University of Chicago) for the kind provision of the AdGFP, AdRFP,
AdS100A9, Adsiβ-catenin recombinant adenoviruses, pGST-moluc and
pGST-moluc-hS100A9 plasmids.
References
|
1
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2018. CA Cancer J Clin. 68:7–30. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Zhang X, Wang Y, Cao Y, Zhang X and Zhao
H: Increased CCL19 expression is associated with progression in
cervical cancer. Oncotarget. 8:73817–73825. 2017.PubMed/NCBI
|
|
3
|
Wu R, Duan L, Ye L, Wang H, Yang X, Zhang
Y, Chen X, Zhang Y, Weng Y, Luo J, et al: S100A9 promotes the
proliferation and invasion of HepG2 hepatocellular carcinoma cells
via the activation of the MAPK signaling pathway. Int J Oncol.
42:1001–1010. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Wu R, Duan L, Cui F, Cao J, Xiang Y, Tang
Y and Zhou L: S100A9 promotes human hepatocellular carcinoma cell
growth and invasion through RAGE-mediated ERK1/2 and p38 MAPK
pathways. Exp Cell Res. 334:228–238. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Zhao Q, He Y, Wang XL, Zhang YX and Wu YM:
Differentially expressed proteins among normal cervix, cervical
intraepithelial neoplasia and cervical squamous cell carcinoma.
Clin Transl Oncol. 17:620–631. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Zhu X, Jin L, Zou S, Shen Q, Jiang W, Lin
W and Zhu X: Immunohistochemical expression of RAGE and its ligand
(S100A9) in cervical lesions. Cell Biochem Biophys. 66:843–850.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Qureshi R, Arora H and Rizvi MA: EMT in
cervical cancer: Its role in tumour progression and response to
therapy. Cancer Lett. 356B:B321–B331. 2015. View Article : Google Scholar
|
|
8
|
Yin C, Li H, Zhang B, Liu Y, Lu G, Lu S,
Sun L, Qi Y, Li X and Chen W: RAGE-binding S100A8/A9 promotes the
migration and invasion of human breast cancer cells through actin
polymerization and epithelial-mesenchymal transition. Breast Cancer
Res Treat. 142:297–309. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Yang M, Wang M, Li X, Xie Y, Xia X, Tian
J, Zhang K and Tang A: Wnt signaling in cervical cancer? J Cancer.
9:1277–1286. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Sun X and Liu Y: Activation of the
Wnt/β-catenin signaling pathway may contribute to cervical cancer
pathogenesis via upregulation of Twist. Oncol Lett. 14:4841–4844.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Shin S, Im HJ, Kwon YJ, Ye DJ, Baek HS,
Kim D, Choi HK and Chun YJ: Human steroid sulfatase induces
Wnt/β-catenin signaling and epithelial-mesenchymal transition by
upregulating Twist1 and HIF-1α in human prostate and cervical
cancer cells. Oncotarget. 8:61604–61617. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Busch EL, McGraw KA and Sandler RS: The
potential for markers of epithelial-mesenchymal transition to
improve colorectal cancer outcomes: A systematic review. Cancer
Epidemiol Biomarkers Prev. 23:1164–1175. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Duan L, Wu R, Ye L, Wang H, Yang X, Zhang
Y, Chen X, Zuo G, Zhang Y, Weng Y, et al: S100A8 and S100A9 are
associated with colorectal carcinoma progression and contribute to
colorectal carcinoma cell survival and migration via Wnt/β-catenin
pathway. PLoS One. 8:e620922013. View Article : Google Scholar
|
|
14
|
Li X, Zhou Q, Tao L and Yu C:
MicroRNA-106a promotes cell migration and invasion by targeting
tissue inhibitor of matrix metalloproteinase 2 in cervical cancer.
Oncol Rep. 38:1774–1782. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Ye H, Zhang Y, Geng L and Li Z: Cdc42
expression in cervical cancer and its effects on cervical tumor
invasion and migration. Int J Oncol. 46:757–763. 2015. View Article : Google Scholar
|
|
16
|
Wang C, Gu W, Zhang Y, Ji Y, Wen Y and Xu
X: Nicotine promotes cervical carcinoma cell line HeLa migration
and invasion by activating PI3k/Akt/NF-κB pathway in vitro. Exp
Toxicol Pathol. 69:402–407. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Xia C, Braunstein Z, Toomey AC, Zhong J
and Rao X: S100 proteins as animportant regulator of macrophage
inflammation. Front Immunol. 8:19082018. View Article : Google Scholar
|
|
18
|
Chen KT, Kim PD, Jones KA, Devarajan K,
Patel BB, Hoffman JP, Ehya H, Huang M, Watson JC, Tokar JL, et al:
Potential prognostic biomarkers of pancreatic cancer. Pancreas.
43:22–27. 2014. View Article : Google Scholar
|
|
19
|
Shibata F, Ito A, Ohkuma Y and Mitsui K:
Mitogenic activity of S100A9 (MRP-14). Biol Pharm Bull.
28:2312–2314. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Ghavami S, Rashedi I, Dattilo BM, Eshraghi
M, Chazin WJ, Hashemi M, Wesselborg S, Kerkhoff C and Los M:
S100A8/A9 at low concentration promotes tumor cell growth via RAGE
ligation and MAP kinase-dependent pathway. J Leukoc Biol.
83:1484–1492. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Hermani A, De Servi B, Medunjanin S,
Tessier PA and Mayer D: S100A8 and S100A9 activate MAP kinase and
NF-kappaB signaling pathways and trigger translocation of RAGE in
human prostate cancer cells. Exp Cell Res. 312:184–197. 2006.
View Article : Google Scholar
|
|
22
|
Jolly MK, Ware KE, Gilja S, Somarelli JA
and Levine H: EMT and MET: Necessary or permissive for metastasis?
Mol Oncol. 11:755–769. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Klymkowsky MW and Savagner P:
Epithelial-mesenchymal transition: A cancer researcher's conceptual
friend and foe. Am J Pathol. 174:1588–1593. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Li A, Gu Y, Li X, Sun H, Zha H, Xie J,
Zhao J, Huang M, Chen L, Peng Q, et al: S100A6 promotes the
proliferation and migration of cervical cancer cells via the
PI3K/Akt signaling pathway. Oncol Lett. 15:5685–5693.
2018.PubMed/NCBI
|
|
25
|
Wang X, Yang J, Qian J, Liu Z, Chen H and
Cui Z: S100A14, a mediator of epithelial-mesenchymal transition,
regulates proliferation, migration and invasion of human cervical
cancer cells. Am J Cancer Res. 5:1484–1495. 2015.PubMed/NCBI
|
|
26
|
Cain JM, Ngan H and Garland S; FIGO
Working Group on Combating Cervical Cancer: Control of cervical
cancer: Women's options and rights. Int J Gynaecol Obstet.
106:141–143. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Lustig B and Behrens J: The Wnt signaling
pathway and its role in tumor development. J Cancer Res Clin Oncol.
129:199–221. 2003.PubMed/NCBI
|
|
28
|
Kahn M: Can we safely target the WNT
pathway? Nat Rev Drug Discov. 13:513–532. 2014. View Article : Google Scholar : PubMed/NCBI
|