Introduction
Esophageal squamous cell carcinoma (ESCC) is a
common histological subtype of esophageal cancer that occurs
primarily in China (1). Standard
therapeutic interventions are surgery, radiotherapy and
chemotherapy, which result in a poor 5-year survival rate of
<20% (2,3). Although recent advances in clinical
diagnosis and therapeutics have improved the survival rate, the
local recurrence and refractory radioresistance remain high,
leading to a limited chance of cure (4). ESCC cells treated by fractionated
irradiation (FIR) may survive due to high adaptive evolution of the
cancer cells (5). Therefore, the
development of promising therapeutic strategies to increase
radiosensitivity and reverse irradiation resistance of ESCC cells
is crucial. Understanding the dynamic changes in the gene
expression regulation network and heterogeneity of ESCC cells
before and after radiotherapy is urgently needed for researchers
and clinicians.
Numerous studies have identified differentially
expressed genes (DEGs) associated with tumor radioresistance by
comparing gene expression in parental cell lines with that in
radioresistant sub-clones exhibiting different levels of radiation
sensitivity (6,7). In radiosensitive ESCC cells, inactive
protein tyrosine kinase 7 (PTK7) overexpression has been
demonstrated to promote cell survival and inhibit radiation-induced
apoptosis, whereas in radioresistant ESCC cells, knockdown of PTK7
decreased the survival of irradiated cells and increased
radiation-induced apoptosis; PTK7 regulates radioresistance through
nuclear factor-κB in ESCC (8). In
addition, microRNA-205 was upregulated in radioresistant ESCC cells
compared with their parental cells by enhancing DNA repair,
inhibiting apoptosis and activating epithelial-mesenchymal
transition (9). Next-generation
sequencing (NGS) has been widely used as a tool for cancer
transcriptome analysis (10,11),
which can provide detailed transcriptome variations between cancer
samples. In addition, using single-cell transcriptome sequencing
(scRNA-seq) (12), the
heterogeneity of cancer cells with various gene signatures and
crucial cancer-associated signaling pathways has been revealed
(13). In addition, bioinformatics
approaches that identify the regulation networks between genes and
phenotypes have revealed functional pathways, as well as indicated
the key molecules in complex diseases. The Weighted Gene
Co-expression Network Analysis (WGCNA) has been extensively
utilized to analyze whole-genome gene expression profiles of
microarray and RNA-Seq data (14-17).
To characterize the acquired radioresistance in a
population of cancer cells, KYSE-180 ESCC cells were continuously
exposed to different cumulative irradiation doses (2, 6, 12, 18, 24
and 30 Gy) to induce radioresistance. This FIR exposure procedure
mimics the process of radiotherapy in patients with ESCC. KYSE-180
cells with different FIR dosages were collected and their gene
expression profiles were analyzed using whole transcriptome
sequencing with non-radiated KYSE-180 cells as controls. It was
hypothesized that cancer cells with acquired radioresistance
accumulated and that the genes associated with radioresistance
dynamically changed during FIR treatment. To identify the subgroups
of radioresistant cancer cells and to understand the gene
expression dynamics in these subgroups, single-cell transcriptome
sequencing was performed on the 0, 12 and 30 Gy FIR-treated
KYSE-180 cells to characterize the heterogeneity of irradiated
cancer cells. By examining the whole genome transcriptome profiles
of in vitro cultured ESCC cells and patient tumor samples
before and after acquisition of radioresistance at the population
and single cell level, the present study aimed to reveal
transcriptomic features that corresponded to FIR treatment and to
further clarify the potential mechanisms and significant genes
responsible for acquired tumor radioresistance.
Materials and methods
Cell culture and irradiation
treatment
The human ESCC cell line KYSE-180 was purchased from
the Cell Bank of Type Culture Collection of the Chinese Academy of
Sciences and passaged for <4 generations. The cells were
cultured in RPMI-1640 medium (Gibco; Thermo Fisher Scientific,
Inc.) with 500 ng/ml penicillin-streptomycin and 10% fetal bovine
serum at 37°C with 5% CO2.
The irradiation treatment was performed as
previously described (18).
KYSE-180 cells (3×106 cells/flask) were seeded into 75
cm2 culture flasks. When the cells reached ~70%
confluence, the medium was replaced and the cells were irradiated
with 2 Gy X-rays using a linear accelerator (Elekta Instrument AB)
at an average dose rate of 100 cGy/min, a 20×20 cm field and a
source-skin distance of 100 cm. Following irradiation, cells were
immediately returned to the incubator. The irradiation (2 Gy) was
repeated on days 2 and 3, and the cells were cultured for 4 days to
recover. When 90% confluence was reached, cells were trypsinized
and sub-cultured into new flasks. These procedures were repeated
five times to achieve a total dose of 30 Gy (KYSE-180-30 Gy, RA30)
(18). Parental cells used as the
irradiation control were trypsinized, counted and passaged under
the same conditions without irradiation. When the repeated
procedures were completed, cells were trypsinized and washed, and
single cells were captured by a micromanipulator for scRNA-seq. The
remaining cells were collected for bulk cell RNA-seq.
Colony formation assay
KYSE-180 cells (400 cells/well) were seeded in a
6-well plate and cultured for 24 h. The plates were irradiated with
8 Gy FIR; the control group was not irradiated. Following
treatment, all plates were returned to the incubator and cultured
for one week. The cells were washed with PBS, fixed with methanol
for 10 min at room temperature and stained with 0.1% crystal violet
(cat. no. C0121; Beyotime Institute of Biotechnology). Following
incubation for 30 min at 37°C, the crystal violet staining was
removed, and the plates were washed with deionized water and dried
at room temperature. The colonies in each plate were counted to
calculate the colony formation rate with light microscope (SZX16,
OLYMPUS), magnification 4×.
Immunofluorescence assay of
phosphorylated H2A histone family member X (γ-H2AX) expression
Primary KYSE180 cells (2,000 cells/well) in 96-well
plates at 1daypost-seeding were exposed to 4 Gy radiation for 2 h,
fixed with acetone/methanol (1:1) 15 min, and permeabilized with
Triton-X 100 (0.1%) in PBS 30 min. Non-specific binding was blocked
by 3% BSA (Sigma-Aldrich; Merck KGaA) in PBS at room temperature
for 1 h. Cells were incubated with an anti-γ-H2AX antibody
(dilution, 1:100; cat. no. 2595; Cell Signaling Technology, Inc.)
at room temperature for 2 h in PBS with 0.1% BSA followed by Alexa
Fluor 488-conjugated secondary antibody at room temperature for 1 h
(dilution, 1:1,000; cat. no. R32723; Invitrogen; Thermo Fisher
Scientific, Inc.) to complete the indirect immunofluorescence
procedure. Immunofluorescence images were obtained by confocal
laser scanning microscopy (magnification, ×40; 4 fields of
view).
Bulk RNA-seq for irradiated ESCC cells
and tissues
Bulk KYSE-180 (RA0), KYSE-180-2 Gy (RA2), KYSE-180-6
Gy (RA6), KYSE-180-12 Gy (RA12), KYSE-180-18 Gy (RA18), KYSE-180-24
Gy (RA24) and KYSE-180-30 Gy (RA30) cells (4×106 cells)
were collected, when the cells reached the respective doses of
irradiation during continuous daily irradiation (two replicates per
sample).
Samples of post-radiotherapy and recurrent tumors
were obtained from a patient with ESCC. The recruitment period was
from May 2016 to May 2018. A male ESCC patient (48 years) was
recruited and the following inclusion criteria were applied: (i)
FIR treatment; and (ii) cancer tissues collectable via biopsy. Two
samples were collected; in May 2016 cancer tissue after
radiotherapy and in March 2017 cancer tissue after recurrence.
Patient sample collection, patient consent and recruitment followed
protocols approved by the Institutional Review Board of The First
Affiliated Hospital of Kunming Medical University (Kunming,
China).
Total RNA from cancer tissues (homogenized prior to
use) and cells were extracted using the RNeasy Mini kit (Qiagen,
Inc.) and the quality of the RNA was evaluated using Agilent
Bioanalyzer 2100 (Agilent Technologies, Inc.). Sequence libraries
were prepared using a TruSeq Stranded mRNA Library Prep kit
(Illumina, Inc.) according to the manufacturer's instructions and
sequenced.
Single-cell (sc)RNA-seq for irradiated
KYSE-180 cells
FIR-treated KYSE-180 cells (RA0, RA12 and RA30) were
trypsinized and resuspended in PBS. Cell numbers were determined
using a cell counter and adjusted to 1×103 cells/ml.
Single cells were placed in 0.2 ml PCR tubes with a
micromanipulation system (cat. no. MPC-285; Sutter Instrument
Company) (19). A total of 48
cells were prepared for RA0, 96 cells for RA12 and 96 cells for
RA30. Single-cell transcriptomes were amplified with SMART-seq2
(20) and cDNAs were purified
using Agencourt Ampure XP beads (cat. no. A63881; Beckman Coulter,
Inc.). The yield and quality of cDNAs were determined by
fluorometric quantitation. DNA >0.5 ng/µl and fragment
distributions of 500-7,000 bp were used to evaluate the quality of
the single-cell transcriptome amplification. The cDNA (1 ng/cell)
was used for library preparation with the Nextera XT DNA Sample
Preparation kit (Illumina, Inc.). Sequencing libraries were
prepared using the Illumina HiSeq 2500 platform (Illumina, Inc.). A
total of 229 FIR-treated single cells (scRA0, scRA12 and scRA30)
were sequenced following 3 days of recovery, yielding ~2 million
high quality reads for each cell. ScRNA-seq data were uploaded to
the National Center for Biotechnology Information reference
sequence database with the accession number GSE81812 (https://www.ncbi.nlm.nih.gov/gds/?term=GSE81812).
ScRNA-seq data selection, normalization
and DEG identification
Raw RNA-seq reads were trimmed using Trimmomatic
0.35 (21) and mapped to the
GENCODE GRCh38 human reference genome (GENCODE v22) using Tophat
2.1.0 software (22). Gene counts
were obtained using the Rsubread 1.34.7 and cleaned by scde 2.12.0
(23). Expression levels were
calculated by normalized gene counts and variance-stabilizing
transformation for clustering by the DESeq2 1.24.0 in Bioconductor
3.9 (24). Relative standard
deviations (RSDs) were calculated as standard deviations of each
gene divided by average normalized gene count (25). Standard methods were used to
evaluate statistical significance. Normalized counts of each gene
were used for the analysis of DEGs by Monocle 2.12.0 (26). The sincell 1.16.0 package (27) was used to draw principal component
analysis (PCA) plots and the cluster-Profiler 3.12.0 package
(28) was used for gene enrichment
analysis. To ensure the fidelity of the data, single cells with
<1,800 detected genes were excluded. Genes with <10 counts in
each cell were excluded and genes that appeared in ≥5 cells were
included. All data from scRNA-seq were analyzed and DEGs were
identified by comparing scRA0, scRA12 and scRA30.
WGCNA
WGCNA was used for scale-free network topology
analysis of RNA-seq data. The WGCNA 1.51 R package (https://horvath.genetics.ucla.edu/html/CoexpressionNetwork/Rpackages/WGCNA/)
was used to cluster highly correlated genes and determine clusters
in which gene expression was associated with the examined
characteristics. An adjacency matrix based on expression
correlation was created using a soft threshold procedure to allow
scale-free topology. The clusters created by WGCNA are referred to
as modules and the minimum number of genes in a module was set to
30. The functional annotation tool DAVID Bioinformatics Resources
6.7 (https://david.ncifcrf.gov/summary.jsp) was used to
determine Gene Ontology (GO) terms enriched by the identified
genes. DAVID analyses were performed using genes corresponding to
significant WGCNA modules.
DAVID analysis
DAVID (http://david.abcc.ncifcrf.gov) functional annotation
bioinformatics microarray analysis was used to identify
significantly enriched GO and Kyoto Encyclopedia of Genes and
Genomes (KEGG) terms among the genes that were differentially
expressed in control samples. Statistically overrepresented GO and
KEGG categories with P≤0.05 were considered significant.
Statistical analysis
Data are presented as the mean ± SEM. All of the
experiments were independently performed in triplicate. All graphs
were plotted and analyzed using GraphPad Prism 5 (GraphPad
Software, Inc.) with one-way ANOVA followed by Dunnett's multiple
comparisons tests. Correlation coefficients were calculated by
Pearson's correlation analysis using R software 3.6.1 (https://www.r-project.org/). P<0.05 was considered
to indicate a statistically significant difference.
Results
Induction of radioresistance in human
ESCC cells
To characterize the transcriptome dynamics
associated with the FIR response, the radiosensitive human ESCC
cell line KYSE-180 was exposed to FIR following the procedure
described by Jing et al (18). KYSE-180 cells were isolated from
well-differentiated human ESCC (JCRB1083), and carried a cyclin D1
amplification and a p53 mutation that commonly occurs in ESCC in
the Asian population (29,30). A preliminary study demonstrated
that compared with KYSE150 and KYSE30, KYSE180 was more sensitive
to FIR (data no shown). The in vitro radioresistance
acquisition model was established to simulate a daily FIR treatment
process in patients with ESCC. The results demonstrated that daily
low-dose irradiation (2 Gy/day) for 3 days did not influence
overall cell viability, and a cumulative dose of 12 Gy for 2 weeks
caused instability and apoptosis in KYSE-180 cells (Fig. 1A). Following 4 days of recovery,
surviving RA12 cells exhibited dramatic cell morphology changes
compared with untreated RA0; ~24.4% of RA12 were abnormal and
exhibited fusiform morphology (Fig.
1B). When continuous FIRs reaching a 30 Gy cumulative
irradiation dose in a period of 5 weeks was applied to these cells,
the morphology of the majority of RA30 cancer cells was aberrantly
changed (fusiform cells, 60.4%; Fig.
1B). The levels of radioresistance of RA0, RA12 and RA30 cells
were determined using colony formation assays combined with DNA
damage analysis using γ-H2AX immunostaining. In the colony
formation assay, cancer cells were treated with 4 and 8 Gy FIR, the
number of colonies was quantified on day 7. RA12 and RA30 displayed
increased resistance to FIR by exhibiting significantly more
colonies compared with untreated RA0 controls (Fig. 1C). In addition, γ-H2AX
immunostaining demonstrated that RA12 and RA30 cells exhibited
significantly fewer γ-H2AX-positive cells compared with RA0,
indicating that RA12 and RA30 cancer cells rapidly repaired DNA
damage following FIR treatment (Fig.
1D). In summary, these results suggested that an in
vitro radioresistance acquisition model for ESCC was
successfully established.
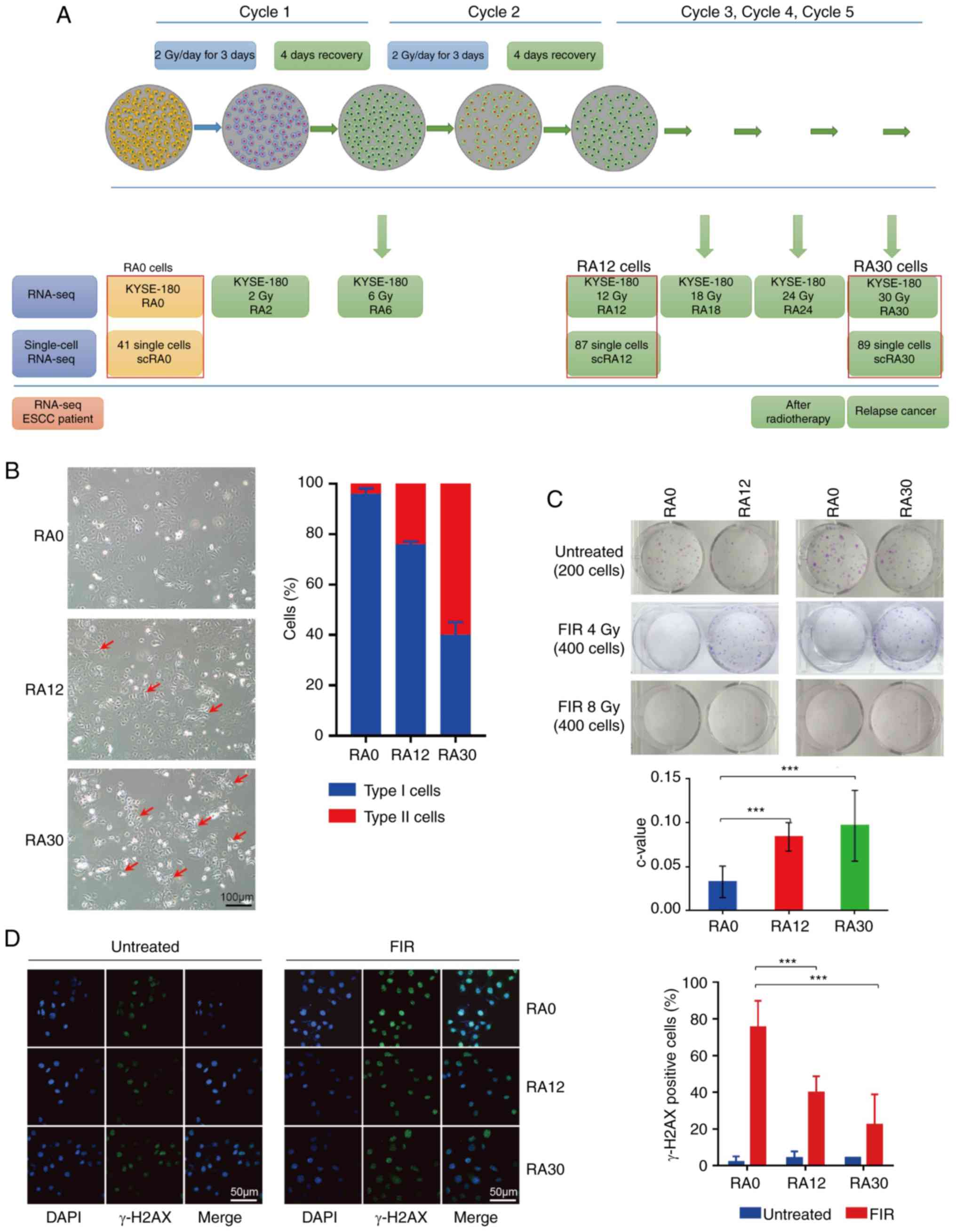 | Figure 1Cancer radioresistance in response to
FIR in vitro. (A) Regimen for induction of radioresistance
in ESCC cells. KYSE-180 cells (yellow) were treated with FIR as
indicated: For one cycle, cells were treated FIR 2 Gy/day on days
1, 2 and 3. From days 4-7, cells were cultured. Most stressed cells
(red) recovered and few cells died. Five cycles of FIR treatment
were performed, and populations in each cycle (total radiation 0,
2, 6, 12, 18, 24 and 30 Gy) were collected and sequenced with bulk
RNA-seq. Single cells of RA0, RA12 and RA30 (red box) were picked
and sequenced with SMART-seq2. Patient tissues were collected and
directly sequenced with bulk RNA-seq. (B) Cell morphology of RA0,
RA12 and RA30 cells; images were collected with an inverted
microscope (magnification, ×20; scale bar, 100 µm). Type I,
original KYSE-180 cells; type II, spindle cells (morphology
changed; red arrows). (C) Colony formation assay of RA0, RA12 and
RA30 cells; relative colony formation was measured with c-value
(colony number with FIR/colony number without treatment). (D)
Immunofluorescence analysis regarding the ability of DNA damage
repair in RA0, RA12 and RA30 cells; cells were stained with
anti-γ-H2AX. Images were recorded using a fluorescence microscope
(magnification, ×40; scale bar, 50 µm). γ-H2AX positive
cells (damaged cells) were defined with ≥5 foci per nuclei. Data
are presented as the mean ± SEM; representative of triplicates.
***P<0.01. FIR, fractionated irradiation; ESCC,
esophageal squamous cell carcinoma; seq, sequencing; RA, samples
with final radiation dose in Gy; γ-H2AX, phosphorylated H2A histone
family member X. |
Genes involved in the radioresistance of
irradiated ESCC cells
To analyze the response of ESCC cells to FIR, whole
transcriptome sequencing of irradiated ESCC cells at cumulative
irradiation doses of 0, 2, 6, 12, 18, 24 and 30 Gy were conducted
(Table SI). The transcriptional
profiles of all irradiated ESCC cells were distinguished from that
of the non-irradiated RA0 cancer cells. Unsupervised hierarchical
clustering of gene expression in the irradiated ESCC cells
demonstrated that transcriptional profiles of RA2+RA6, RA12+RA18,
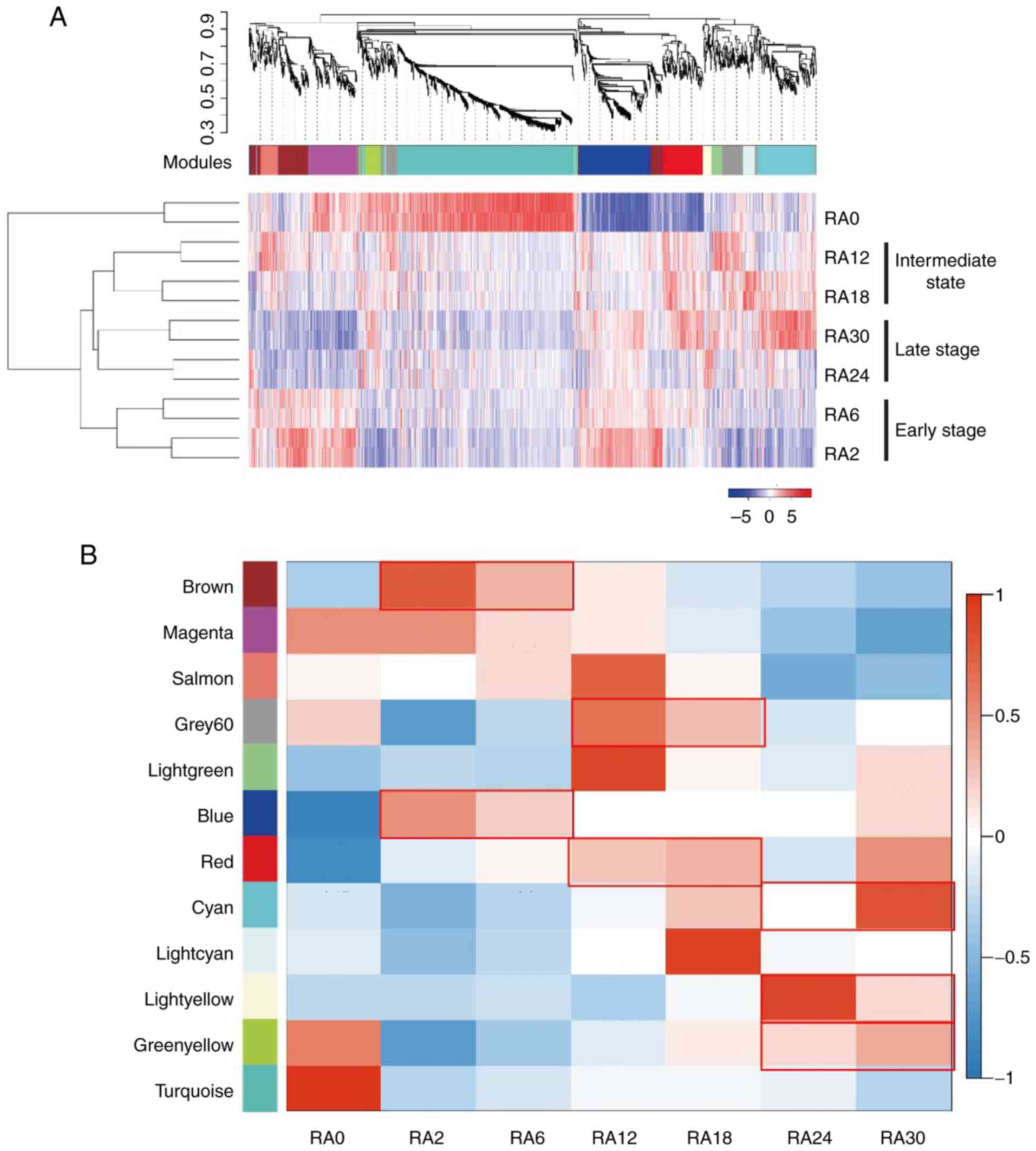
and RA24+RA30 were clustered together (Fig. 2A). These results suggested that
RA2/RA6, RA12/RA18 and RA24/RA30 cells were at an early,
intermediate and late stage of acquired radioresistance,
respectively. To assess the transcriptome dynamics of ESCC cells at
different stages of irradiation, WGCNA was performed and multiple
gene-network modules associated with early, intermediate and late
stages of irradiation were obtained (Fig. 2A; Table SII). At the early stage, genes in
the 'brown', 'magenta' and 'blue' modules were highly expressed
(Fig. 2B). GO term biological
process analysis of these genes revealed significant enrichment in
the pathways of 'RNA metabolic signaling' (P<0.001), 'cell
cycle' (P<0.001) and 'protein translation' (P<0.001)
(Fig. 2C and D; Table SIII). These results indicated that
during the early stage of irradiation, ESCC cells undergo a rapid
cellular response to environmental stress. During the intermediate
stage, genes in the 'grey' and 'red' modules were highly expressed
(Fig. 2B). Genes in these modules
were significantly enriched in the pathways of 'cell adhesion'
(P<0.001), 'cell migration' (P<0.001) and 'cell
proliferation' (P<0.001) (Fig. 2E
and F; Table SIV), which was
consistent with the observed morphological changes in the RA12
cells (Fig. 1B). During the late
stage of irradiation, genes in the 'cyan', 'light yellow' and
'green-yellow' modules were significantly upregulated (Fig. 2B). Genes in these modules were
enriched in cancer radioresistance-associated signaling pathways,
including 'response to hypoxia' (P=0.0015) (31), 'regulation of autophagy' (P=0.0038)
(32) and 'DNA damage response'
(P=0.016) (33) (Fig. 2G; Table SIII). In addition, KEGG enrichment
analysis demonstrated that 'mTOR signaling pathway' (P=0.057) was
also activated in the RA24 and RA30 cells (Fig. 2H; Table SIV) (34,35).
In summary, transcriptome dynamics revealed that ESCC cells respond
to FIR rapidly with increased RNA metabolism at the beginning of
radiotherapy, followed by upregulated cell adhesion and finally
acquired radioresistance with the activation of multiple known
radioresistance-associated signaling pathways during continuous
irradiation.
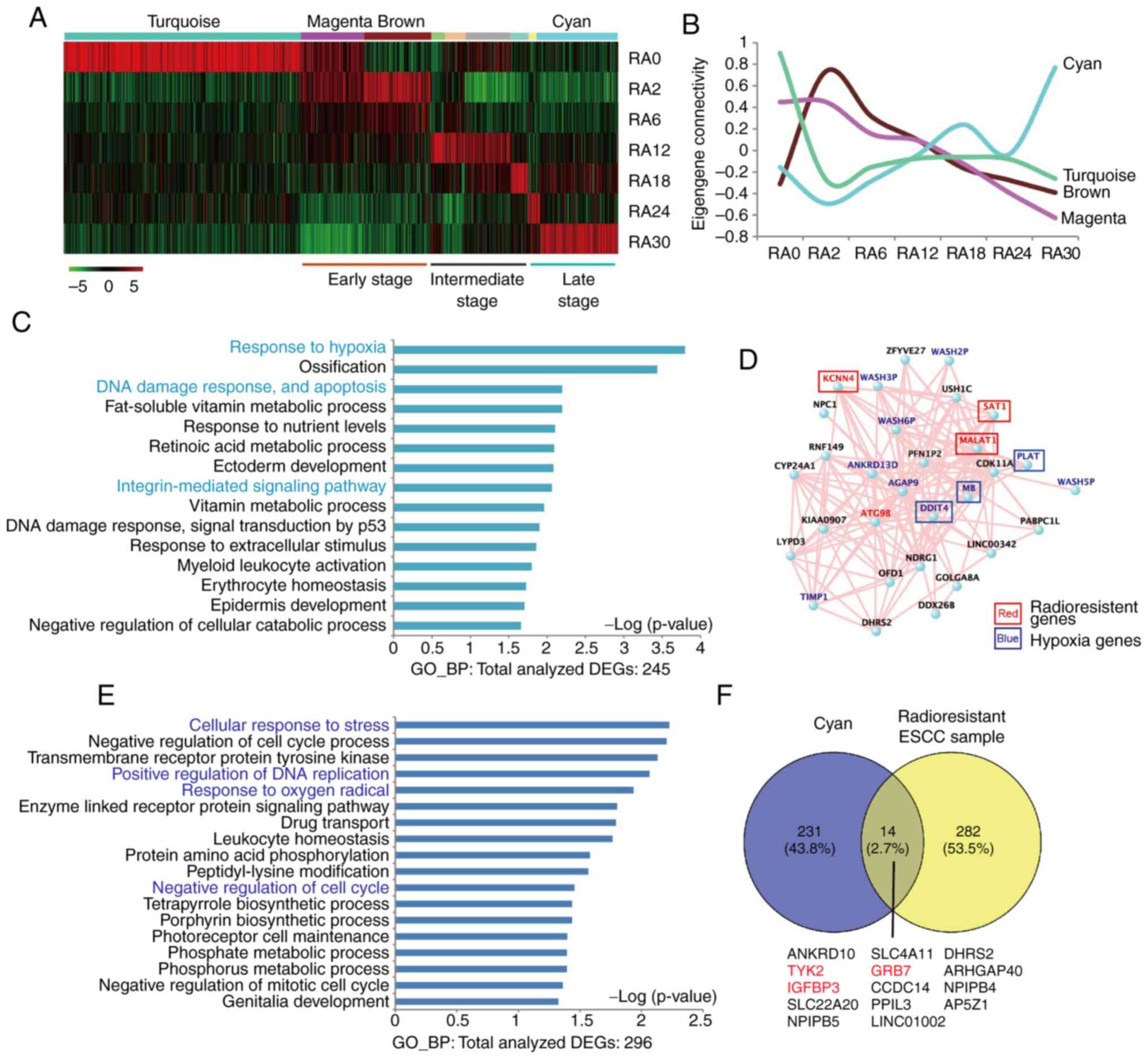
The modules that reflected unique gene expression
dynamics across irradiated ESCC cells were further analyzed. A
total of four modules of interest were identified: 'Turquoise',
'brown', 'magenta' and 'cyan' (Fig. 3A
and B). In addition, hub-gene-network analysis of these modules
revealed highly connected genes in each module, through which key
controlling genes in the modular network were identified (Figs. 3C and D and S1). The 'turquoise' module contained a
significant number of known cancer-related genes, such as
WNT5A (36,37), transmembrane protease serine 2
(TMPRSS2) (38), ATPase
copper-transporting α (ATP7A) (39) and others. The 'turquoise' module
gene expression significantly decreased in irradiated ESCC cells
(Fig. S1), suggesting that these
genes were downregulated by radiotherapy. The 'brown' module, which
contained genes that were upregulated in the early stage of
irradiation and declined during further irradiation treatment, was
significantly enriched in genes involved in RNA metabolism
(Fig. S1). The expression of
genes in the 'magenta' module, which were abundantly expressed in
untreated RA0 and early-stage RA2 and RA6 cells, continuously
decreased following cumulative irradiation treatment. Thus, the
expression of the genes in the 'magenta' module was negatively
associated with the acquisition of radioresistance in ESCC and may
serve functional roles in the development of cancer cell
radioresistance. GO term analysis demonstrated that the genes in
the 'magenta' module were significantly enriched in biological
processes involved in 'WNT signaling' (P=0.043) and 'chromosome
organization' (P=0.0032) (Table
SIII). The network of the top hub-genes in the 'magenta' module
is presented in Fig. S1. By
contrast, genes in the 'cyan' module, which were aberrantly
overexpressed in RA30 cells compared with untreated and irradiated
ESCC cells at the early and intermediate stages, may be associated
with the acquisition of radioresistance in ESCC. GO term analysis
demonstrated that the genes in the 'cyan' module were significantly
enriched in the biological processes involved in 'response to
hypoxia' (P<0.0001) and 'DNA damage' (P=0.006) (Fig. 3C; Table SIII). Previously reported
radioresistance-related genes in the 'cyan' module were potassium
calcium-activated channel subfamily N member 4 (KCNN4)
(40), spermidine/spermine
N1-acetyltransferase 1 (SAT1) (41) and metastasis-associated lung
adenocarcinoma transcript 1 (MALAT1) (42,43)
(Fig. 3D). The hub-gene
autophagy-related 9B (ATG9B) in the 'cyan' module may serve
important roles in ESCC radioresistance through autophagy. DNA
damage-inducible transcript 4 (DDIT4), myoglobin (MB)
and plasminogen activator tissue type (PLAT) are associated
with hypoxia (44), which may also
be associated with radioresistance. Pseudogenes of the WASH protein
family that recruit and activate the Arp2/3 complex to induce actin
polymerization, which have a key role in the fission of tubules
that serve as transport intermediates during endosome sorting
(45), WASH2P,
WASH3P, WASH5P and WASH6P may be related to
radioresistance. Overall, these results demonstrated that
transcriptome dynamics were associated with cell adhesion,
autophagy, hypoxia and DNA damage during irradiation treatment and
revealed a number of genes that may serve crucial roles in the
development of radioresistance.
Radioresistance genes in tumor samples
from FIR-treated patients with ESCC
To validate the radioresistant genes identified in
the in vitro experiments in the present study, paired tumor
tissue samples were collected from one radiotherapy-treated and
relapsed patient with ESCC. Comparison between the gene expression
profiles of the primary and relapsed tumor samples revealed that
296 genes were upregulated in the relapsed tumor samples (Table SV). GO term biological process
analysis of these genes demonstrated significant enrichment in the
pathways associated with 'cell response to stress' (P=0.006), 'cell
cycle' (P=0.006), 'DNA repair' (P=0.009) and 'hypoxia' (P=0.012).
The result obtained using patient samples were consistent with the
in vitro results (Fig. 3E;
Table SVI). To examine whether
the relapsed tumor was derived from residual radioresistant cancer
cells, the 296 upregulated genes were compared with the previously
identified radioresistance-associated genes in the 'cyan' module. A
total of 14 overlapping genes were identified, including certain
known resistance-related genes tyrosine kinase 2 (TYK2)
(46), growth factor
receptor-bound protein 7 (GRB7) (47) and insulin-like growth
factor-binding protein 3 (IGFBP3) (48) (Fig.
3F). These results demonstrated a conserved common molecular
mechanism in radioresistance between the in vitro ESCC model
and the patient samples, indicating that radioresistance genes
('cyan' module) identified in the irradiated cell line model serve
crucial roles in acquired radioresistance.
Radioresistance dynamics at the
single-cell level
To determine the mechanism of the acquisition of
radioresistance in ESCC, whole transcriptome analysis was performed
at the single-cell level in RA0, RA12 and RA30 cancer cells.
ScRNA-seq reads were mapped to the reference human genome with an
average of 65.4% reads mapped within the genome. To test the
reproducibility of data, the sequencing was repeated using the same
templates; a strong correlation (r=0.96) was obtained, which
confirmed the reliability of the sequencing data (Fig. S2A). To obtain accurate estimates
of the gene expression levels, the sequencing depth was normalized
to ensure that all cells exhibited approximately the same median
read depth (Fig. S2B). A total of
217 cells passed this filter, which resulted in 41 qualified RA0
single cells (scRA0), 87 qualified RA12 single cells (scRA12) and
89 qualified RA30 single cells (scRA30). To examine the gene
expression levels in individual cells, the RSD of normalized gene
counts was used to estimate the dispersion. Genes with higher
expression levels exhibited lower degrees of dispersion (Fig. S2C). Genes with <4 normalized
read counts (≤2 log2 value) were removed as they
provided an unreliable measure of gene expression levels (25). The reproducibility of the Illumina
sequencing platform was guaranteed by a high correlation
coefficient (r=0.98; Fig. S2D).
Overall, the results demonstrated that the quality of the scRNA-seq
data was satisfactory for further analysis.
One-way ANOVA was used to analyze the differences
between the RNA-seq data of each group. A total of 3,174 DEGs were
identified for further analysis. PCA demonstrated thatscRA0 was
distinct from scRA12 and scRA30, whereas scRA12 and scRA30 were
partially separated, and a number of single cells from scRA12 and
scRA30 were clustered together (Figs.
4A, S3A and B). The scRNA-seq
data were subjected to WGCNA and multiple gene-network modules were
obtained (Table SVII).
Unsupervised hierarchical clustering of 217 single cell
transcriptome results revealed three major groups (I-III) and three
subgroups in group II: (i) Group I, scR0; (ii) group II, scR1,
scR2, and scR3; and (iii) Group III, scR4 (Fig. 4B). The distribution of single cells
in each group was further examined; the results demonstrated that
scR1, scR2, scR3 and scR4 contained different percentages of scRA12
and scRA30. All cells in group I were untreated RA0 cells. From
scR1 to scR4, the number of scRA12 gradually decreased, whereas the
number of scRA30 gradually increased (Fig. 4C). These results suggested that the
cells in different groups obtained various levels of
radioresistance, and the ESCC cells in the scR4 subgroup may have
developed the highest expression levels of
radioresistance-associated genes compared with the cells in the
other subgroups (Fig. 4C; Table SVIII).
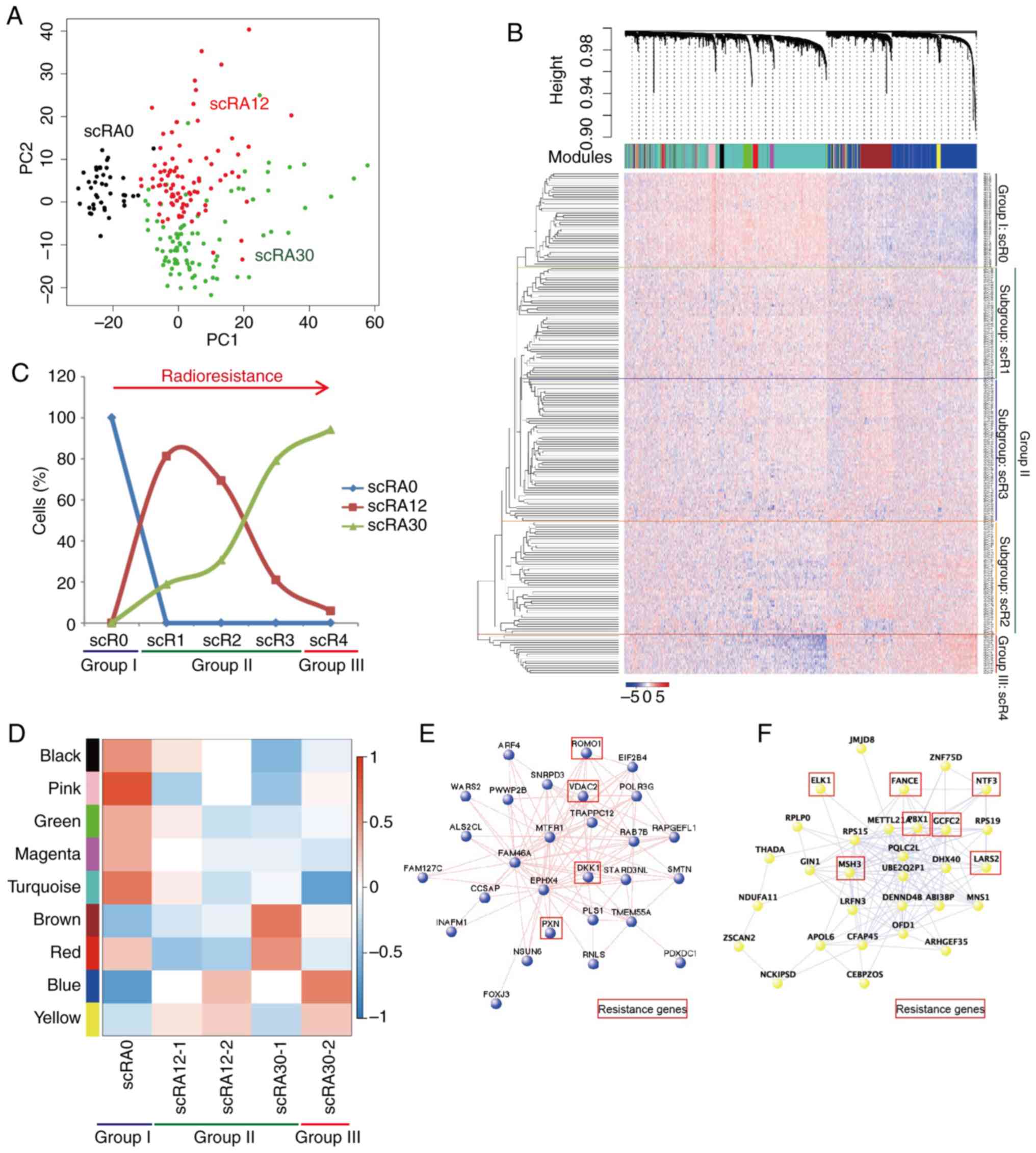 | Figure 4Single-cell transcriptome sequencing
reveals radioresistance. (A) PCA of 217 single cells with
differentially expressed genes. (B) WGCNA dendrogram indicating
expression of gene modules in single cells; based on the
unsupervised hierarchical clustering, with group I including scR0,
group II including scR1, scR2, and scR3, and group III including
scR4. (C) Distribution of scRA0, scR12 and scR30 in clustered
groups. (D) Modules-trait relationships in single-cell groups;
color code of modules is preserved. (E) Hub-gene network of scBlue;
cancer resistance associated genes highlighted with red box. (F)
Hub-gene network of scYellow; cancer resistance associated genes
highlighted with red box. WGCNA, Weighted Gene Co-expression
Network Analysis; PCA, principal component analysis; RA, samples
with final radiation dose in Gy; scR, single radioresistant
cell. |
The scRA12 and scRA30 were divided into two
subgroups (scRA12, scRA12-1 and scRA12-2; and scRA30, scRA30-1 and
scRA30-2), and the module-trait relationships were determined
(Fig. 4D). The results
demonstrated that the genes in the 'yellow', 'blue' and 'brown'
modules were highly expressed in single cells from groups II and
III, whereas genes in the 'black', 'pink', 'green', 'magenta' and
'turquoise' modules were highly expressed in single cells from
group I. Hub-gene network analysis in each module revealed the key
controlling genes in the modular network (Figs. 4E, F and S4). The results demonstrated that the
hub-genes in the 'blue' module of single-cell data (scBlue)
contained multiple known cancer resistant-related genes, including
reactive oxygen species modulator 1 (ROMO1) (49), voltage-dependent anion channel 2
(VDAC2) (50), Dickkopf WNT
signaling pathway inhibitor (DKK1) (49,51)
and paxillin (PXN) (52-54),
which have been reported in previous studies. The hub-genes in the
'yellow' module (scYellow) contained neurotrophin 3 (NTF3)
(55), FA complementation group E
(FANCE) (56) and mutS
homolog 3 (MSH3) (57),
which have been previously demonstrated to be associated with
apoptosis or DNA damage repair processes. Other genes in this
module are also reported to associate with cancer resistance, such
as ETS transcription factor (ELK1) (58), PBX homeobox 1 (PBX1)
(59), GC-rich sequence
DNA-binding factor 2 (GCFC2) (60) and mitochondrial leucyl-tRNA
synthetase 2 (LARS2) (61).
These results revealed the heterogeneity and dynamic gene
expression changes of ESCC cells during the acquisition of
radioresistance.
Discussion
The present study successfully established an in
vitro radioresis-tance acquisition model using radiosensitive
ESCC KYSE-180 cells through consistent low dose FIR treatment. The
results demonstrated that this in vitro model recapitulated
the acquisition process of cancer cell radioresistance following
radiotherapy in patients with ESCC. Throughout the irradiation
process, multiple ESCC cells receiving different doses of
irradiation were collected and, for the first time, the
transcriptome profiles of an in vitro ESCC model at the
population and single-cell level were analyzed. The results
revealed that irradiation-induced dynamics of gene expression and
modules were associated with the stage of radioresistance. The
genes in the 'cyan' module from the population RNA-seq were
specifically enriched in ESCC cells at the late stage of
irradiation. These genes were involved in signaling pathways
associated with autophagy, hypoxia and DNA damage repair.
Considering the crucial roles of autophagy (62) and hypoxia (63) in radioresistance development, the
transcriptome analysis results in the present study suggested that
MALAT1-ATG9B (autophagy) and
DDIT4-MB-PLAT (hypoxia) may induce
radioresistance in cancer cells.
The dynamic changes in gene expression profiles were
compared between the FIR-induced in vitro radioresistance
model and tumor samples from one patient with ESCC undergoing
radiotherapy before and after relapse. Although only 5% (14/296) of
the genes were consistent between radioresistant ESCC cells and
patient-derived samples, two signaling pathways, DNA damage and
hypoxia, were conserved in the model and the patient. More patient
samples are needed to validate these results; other mechanisms may
contribute to radioresistance in other patients in addition to
these two signaling pathways. Among the overlapping genes,
TYK2 and IGFBP3, which were previously reported as
chemoresistance-associated genes (46), were signifi-cantly upregulated in
RA30 ESCC cells and in the relapsed tumor sample. GRB7,
which has been reported to be associated with Erb-B2 receptor
tyrosine kinase 2(HER2) amplification (64), HER2 signaling inhibition (65) and an increased risk of recurrence
in triple negative breast cancer (47), may functionally regulate
radioresistance in ESCC. These results suggested that the
FIR-induced in vitro radioresistance model shared a similar
molecular regulation network involving radioresistance; thus, the
in vitro model established in the present study may bean
efficient tool to study clinical cancer radioresistance.
Single-cell transcriptome analysis of the
FIR-induced in vitro radioresistance model in the present
study revealed cellular heterogeneity in irradiated ESCC cells.
During the process of irradiation, radioresistance is achieved by a
continuous dynamic selection process. Different single ESCC cells
may activate diverse radioresistance-related signaling pathways and
acquire various levels of radioresistance. Among all irradiated
single cancer cells (scRA12 and scRA30), four subgroups were
identified. Single cells in the scR1 subgroup, in which the
expression profile was similar to that in the original single ESCC
cells (scRA0 or scR0), may still be sensitive to irradiation. The
cells in subgroups scR2, scR3 and scR4, which exhibited significant
differences in gene expression profiles compared with the cells in
the scR0 subgroup, may acquire varying levels of radioresistance.
Analysis of the percentage of scRA12 and scRA30 cells in each
subgroup revealed that the cells in scR2, scR3 and scR4 may obtain
increased sensitivity to radioresistance, and the cells in scR4
acquired the highest level of radioresistance. The scRA12 cells
were distributed among the scR1, scR2, scR3 subgroups, and one cell
in the scR4 subgroup, suggesting that cancer radioresistance may
occur in the stage of RA12. By contrast, 10% (9/89) of scRA30 cells
were in the scR1 subgroup and 18% (16/89) of scRA30 cells were in
the scR4 subgroup, which indicated a heterogeneous cell population
of scRA30 cells.
In the present study, WGCNA revealed that the
scBlue, scYellow (upregulated) and scTurquoise (downregulated)
modules significantly represented the molecular features of each
subgroup. Gene network analyses of these modules identified the
hub-genes involved in cancer radioresistance. The scBlue and
scYellow modules exhibited low consistence with the bulk 'cyan'
module and samples from the patient with ESCC (Fig. S5), which may be due to a mixed
population of radioresistant cancer cells in RA30. However, a
number of overlapping genes were identified among these samples,
including GRB7 (64,65).
Other overlapping genes in subgroup scR4 and relapsed tumor samples
may be of importance in acquired radioresistance. Further
validation and investigation, including extensive single-cell
transcriptome analysis in radioresistant ESCC tissues or ESCC
derived PDX models, will be required to determine the precise
molecular pathways of acquired radioresistance. The results of the
present study supported the hypothesis that single-cell
transcriptome analysis of cancer cell lines and patient tumor
samples may bring invaluable insight into the development of
acquired radioresistance from a specific population of cancer
cells. Overall, these results are of potential clinical relevance,
as they revealed genes and signaling pathways involved in
radioresistance and identified opportunities for developing novel
therapeutic options for ESCC.
Supplementary Data
Acknowledgments
The authors would like to thank Dr Xinghua Pan
(Southern Medical University) for single-cell RNA-seq performed at
Yale University. The abstract of the current study was previously
presented at ASTRO2017 (https://www.redjournal.org/article/S0360-3016(17)33168-1/fulltext).
Funding
This work was supported by the National Natural
Science Foundation of China (grant nos. 81903046 and 31860306), the
Health Department of Yunnan Province (grant no. 2018NS0083), Yunnan
Engineering Technology Center of Digestive Disease (grant no.
2018DH006) and Yunling Scholar (grant no. YLXL20170002).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request. The datasets generated and/or analyzed during the current
study are available in the NCBI GEO database with the accession
number GSE81812 (https://www.ncbi.nlm.nih.gov/gds/?term=GSE81812) and
the NCBI SRA database with the accession number SRP110339
(https://www.ncbi.nlm.nih.gov/sra/?term=SRP110339).
All data generated or analyzed during this study are included in
this published article.
Authors' contributions
HW and KW designed the study. DK, YX and ZZ
performed the experiments. HW, JY, and HL analyzed the data and
performed statistical analyses. HW, JY, HL, JS and ZL interpreted
and discussed the data, and wrote the manuscript. All authors read
and approved the final version of the manuscript.
Ethics approval and consent to
participate
All patient sample collections, patient consent and
recruitment followed protocols approved by the institutional review
board of The First Affiliated Hospital, Kunming Medical University.
The study was conducted according to the principles of the
Declaration of Helsinki.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Song Y, Li L, Ou Y, Gao Z, Li E, Li X,
Zhang W, Wang J, Xu L, Zhou Y, et al: Identification of genomic
alterations in oesophageal squamous cell cancer. Nature. 509:91–95.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Song QK, Li J, Jiang HD, He YM, Zhou XQ
and Huang CY: Esophageal cancer mortality during 2004-2009 in
yanting county, China. Asian Pac J Cancer Prev. 13:5003–5006. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Wei WQ, Yang J, Zhang SW, Chen WQ and Qiao
YL: Esophageal cancer mortality trends during the last 30 years in
high risk areas in China: Comparison of results from national death
surveys conducted in the 1970's, 1990's and 2004-2005. Asian Pac J
Cancer Prev. 12:1821–1826. 2011.PubMed/NCBI
|
|
4
|
Brooks-Brunn JA: Esophageal cancer: An
overview. Medsurg Nurs. 9:248–254. 2000.
|
|
5
|
Raman NV and Small W Jr: The role of
radiation therapy in the management of esophageal cancer. Cancer
Control. 6:53–62. 1999. View Article : Google Scholar
|
|
6
|
Michna A, Schotz U, Selmansberger M,
Zitzelsberger H, Lauber K, Unger K and Hess J: Transcriptomic
analyses of the radiation response in head and neck squamous cell
carcinoma subclones with different radiation sensitivity:
Time-course gene expression profiles and gene association networks.
Radiat Oncol. 11:942016. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
McDermott N, Meunier A, Mooney B, Nortey
G, Hernandez C, Hurley S, Lynam-Lennon N, Barsoom SH, Bowman KJ,
Marples B, et al: Fractionated radiation exposure amplifies the
radioresistant nature of prostate cancer cells. Sci Rep.
6:347962016. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Park M, Yoon HJ, Kang MC, Kwon J and Lee
HW: PTK7 regulates radioresistance through nuclear factor-kappa B
in esophageal squamous cell carcinoma. Tumour Biol. 37:14217–14224.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Pan F, Mao H, Bu F, Tong X, Li J, Zhang S,
Liu X, Wang L, Wu L, Chen R, et al: Sp1-mediated transcriptional
activation of miR-205 promotes radioresistance in esophageal
squamous cell carcinoma. Oncotarget. 8:5735–5752. 2017.
|
|
10
|
Roychowdhury S and Chinnaiyan AM:
Translating cancer genomes and transcriptomes for precision
oncology. CA Cancer J Clin. 66:75–88. 2016. View Article : Google Scholar :
|
|
11
|
Shyr D and Liu Q: Next generation
sequencing in cancer research and clinical application. Biol Proced
Online. 15:42013. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Kolodziejczyk AA, Kim JK, Svensson V,
Marioni JC and Teichmann SA: The technology and biology of
single-cell RNA sequencing. Mol Cell. 58:610–620. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Wu H, Yu J, Li Y, Hou Q, Zhou R, Zhang N,
Jing Z, Jiang M, Li Z, Hua Y, et al: Single-cell RNA sequencing
reveals diverse intratumoral heterogeneities and gene signatures of
two types of esophageal cancers. Cancer Lett. 438:133–143. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Fuller TF, Ghazalpour A, Aten JE, Drake
TA, Lusis AJ and Horvath S: Weighted gene coexpression network
analysis strategies applied to mouse weight. Mamm Genome.
18:463–472. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Langfelder P and Horvath S: WGCNA: An R
package for weighted correlation network analysis. BMC
Bioinformatics. 9:5592008. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Presson AP, Sobel EM, Papp JC, Suarez CJ,
Whistler T, Rajeevan MS, Vernon SD and Horvath S: Integrated
weighted gene co-expression network analysis with an application to
chronic fatigue syndrome. BMC Syst Biol. 2:952008. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Zhang B and Horvath S: A general framework
for weighted gene co-expression network analysis. Stat Appl Genet
Mol Biol. 4:172005. View Article : Google Scholar
|
|
18
|
Jing Z, Gong L, Xie CY, Zhang L, Su HF,
Deng X and Wu SX: Reverse resistance to radiation in KYSE-150R
esophageal carcinoma cell after epidermal growth factor receptor
signal pathway inhibition by cetuximab. Radiother Oncol.
93:468–473. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Wu H, Zhang XY, Hu Z, Hou Q, Zhang H, Li
Y, Li S, Yue J, Jiang Z, Weissman SM, et al: Evolution and
heterogeneity of non-hereditary colorectal cancer revealed by
single-cell exome sequencing. Oncogene. 36:2857–2867. 2017.
View Article : Google Scholar
|
|
20
|
Picelli S, Björklund ÅK, Faridani OR,
Sagasser S, Winberg G and Sandberg R: Smart-seq2 for sensitive
full-length transcrip-tome profiling in single cells. Nat Methods.
10:1096–1098. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Bolger AM, Lohse M and Usadel B:
Trimmomatic: A flexible trimmer for Illumina sequence data.
Bioinformatics. 30:2114–2120. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Trapnell C, Roberts A, Goff L, Pertea G,
Kim D, Kelley DR, Pimentel H, Salzberg SL, Rinn JL and Pachter L:
Differential gene and transcript expression analysis of RNA-seq
experiments with TopHat and Cufflinks. Nat Protoc. 7:562–578. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Kharchenko PV, Silberstein L and Scadden
DT: Bayesian approach to single-cell differential expression
analysis. Nat Methods. 11:740–742. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Love MI, Huber W and Anders S: Moderated
estimation of fold change and dispersion for RNA-seq data with
DESeq2. Genome Biol. 15:5502014. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Suzuki A, Matsushima K, Makinoshima H,
Sugano S, Kohno T, Tsuchihara K and Suzuki Y: Single-cell analysis
of lung adenocarcinoma cell lines reveals diverse expression
patterns of individual cells invoked by a molecular target drug
treatment. Genome Biol. 16:662015. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Trapnell C, Cacchiarelli D, Grimsby J,
Pokharel P, Li S, Morse M, Lennon NJ, Livak KJ, Mikkelsen TS and
Rinn JL: The dynamics and regulators of cell fate decisions are
revealed by pseudotem-poral ordering of single cells. Nat
Biotechnol. 32:381–386. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Julia M, Telenti A and Rausell A: Sincell:
An R/Bioconductor package for statistical assessment of cell-state
hierarchies from single-cell RNA-seq. Bioinformatics. 31:3380–3382.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Yu G, Wang LG, Han Y and He QY:
clusterProfiler: An R package for comparing biological themes among
gene clusters. OMICS. 16:284–287. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Kanda Y, Nishiyama Y, Shimada Y, Imamura
M, Nomura H, Hiai H and Fukumoto M: Analysis of gene amplification
and overexpression in human esophageal-carcinoma cell lines. Int J
Cancer. 58:291–297. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Tanaka H, Shibagaki I, Shimada Y, Wagata
T, Imamura M and Ishizaki K: Characterization of p53 gene mutations
in esophageal squamous cell carcinoma cell lines: Increased
frequency and different spectrum of mutations from primary tumors.
Int J Cancer. 65:372–376. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Wilson WR and Hay MP: Targeting hypoxia in
cancer therapy. Nat Rev Cancer. 11:393–410. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Yang Y, Yang Y, Yang X, Zhu H, Guo Q, Chen
X, Zhang H, Cheng H and Sun X: Autophagy and its function in
radiosensi-tivity. Tumour Biol. 36:4079–4087. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Centurione L and Aiello FB: DNA repair and
cytokines: TGF-β, IL-6, and thrombopoietin as different biomarkers
of radioresis-tance. Front Oncol. 6:1752016. View Article : Google Scholar
|
|
34
|
Chang L, Graham PH, Ni J, Hao J, Bucci J,
Cozzi PJ and Li Y: Targeting PI3K/Akt/mTOR signaling pathway in the
treatment of prostate cancer radioresistance. Crit Rev Oncol
Hematol. 96:507–517. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Zheng H, Wang M, Wu J, Wang ZM, Nan HJ and
Sun H: Inhibition of mTOR enhances radiosensitivity of lung cancer
cells and protects normal lung cells against radiation. Biochem
Cell Biol. 94:213–220. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Jiang G, Lin J, Wang W, Sun M, Chen K and
Wang F: WNT5A promoter methylation is associated with better
responses and longer progression-free survival in colorectal cancer
patients treated with 5-fluorouracil-based chemotherapy. Genet Test
Mol Biomarkers. 21:74–79. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Hu B, Wang Q, Wang YA, Hua S, Sauve CG,
Ong D, Lan ZD, Chang Q, Ho YW, Monasterio MM, et al: Epigenetic
activation of WNT5A drives glioblastoma stem cell differentiation
and invasive growth. Cell. 167:1281–1295.e18. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Deplus R, Delliaux C, Marchand N, Flourens
A, Vanpouille N, Leroy X, de Launoit Y and Duterque-Coquillaud M:
TMPRSS2-ERG fusion promotes prostate cancer metastases in bone.
Oncotarget. 8:11827–11840. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Li ZH, Zheng R, Chen JT, Jia J and Qiu M:
The role of copper transporter ATP7A in platinum-resistance of
esophageal squamous cell cancer (ESCC). J Cancer. 7:2085–2092.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Bonito B, Sauter DR, Schwab A, Djamgoz MB
and Novak I: KCa3.1 (IK) modulates pancreatic cancer cell
migration, invasion and proliferation: Anomalous effects on
TRAM-34. Pflugers Arch. 468:1865–1875. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Brett-Morris A, Wright BM, Seo Y,
Pasupuleti V, Zhang J, Lu J, Spina R, Bar EE, Gujrati M, Schur R,
et al: The polyamine catabolic enzyme SAT1 modulates tumorigenesis
and radiation response in GBM. Cancer Res. 74:6925–6934. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Li Z, Zhou Y, Tu B, Bu Y, Liu A and Xie C:
Long noncoding RNA MALAT1 affects the efficacy of radiotherapy for
esophageal squamous cell carcinoma by regulating Cks1 expression. J
Oral Pathol Med. 46:583–590. 2017. View Article : Google Scholar
|
|
43
|
Jin C, Yan B, Lu Q, Lin Y and Ma L: The
role of MALAT1/miR-1/slug axis on radioresistance in nasopharyngeal
carcinoma. Tumour Biol. 37:4025–4033. 2016. View Article : Google Scholar
|
|
44
|
Minchenko OH, Kharkova AP, Kubaichuk KI,
Minchenko DO, Hlushchak NA and Kovalevska OV: Effect of hypoxia on
the expression of CCN2, PLAU, PLAUR, SLURP1, PLAT and ITGB1 genes
in ERN1 knockdown U87 glioma cells. Ukr Biochem J. 86:79–89. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Wang F, Zhang L, Zhang GL, Wang ZB, Cui
XS, Kim NH and Sun SC: WASH complex regulates Arp2/3 complex for
actin-based polar body extrusion in mouse oocytes. Sci Rep.
4:55962014. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Shah K and Bradbury NA: Lemur tyrosine
kinase 2, a novel target in prostate cancer therapy. Oncotarget.
6:14233–14246. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Sparano JA, Goldstein LJ, Childs BH, Shak
S, Brassard D, Badve S, Baehner FL, Bugarini R, Rowley S, Perez EA,
et al: Relationship between quantitative GRB7 RNA expression and
recurrence after adjuvant anthracycline chemotherapy in
triple-negative breast cancer. Clin Cancer Res. 17:7194–7203. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Xu QY, Gao Y, Liu Y, Yang WZ and Xu XY:
Identification of differential gene expression profiles of
radioresistant lung cancer cell line established by fractionated
ionizing radiation in vitro. Chin Med J (Engl). 121:1830–1837.
2008. View Article : Google Scholar
|
|
49
|
Kim IG, Kim SY, Kim HA, Kim JY, Lee JH,
Choi SI, Han JR, Kim KC and Cho EW: Disturbance of DKK1 level is
partly involved in survival of lung cancer cells via regulation of
ROMO1 and γ-radiation sensitivity. Biochem Biophys Res Commun.
443:49–55. 2014. View Article : Google Scholar
|
|
50
|
Roy SS, Ehrlich AM, Craigen WJ and
Hajnoczky G: VDAC2 is required for truncated BID-induced
mitochondrial apoptosis by recruiting BAK to the mitochondria. EMBO
Rep. 10:1341–1347. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Salim H, Zong D, Hååg P, Novak M, Mörk B,
Lewensohn R, Lundholm L and Viktorsson K: DKK1 is a potential novel
mediator of cisplatin-refractoriness in non-small cell lung cancer
cell lines. BMC Cancer. 15:6282015. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Du C, Wang X, Zhang J, Liu X, Zhu J and
Liu Y: Paxillin is positively correlated with the
clinicopathological factors of colorectal cancer, and knockdown of
Paxillin improves sensitivity to cetux-imab in colorectal cancer
cells. Oncol Rep. 35:409–417. 2016. View Article : Google Scholar
|
|
53
|
Wu DW, Chen CY, Chu CL and Lee H: Paxillin
confers resistance to tyrosine kinase inhibitors in EGFR-mutant
lung cancers via modulating BIM and Mcl-1 protein stability.
Oncogene. 35:621–630. 2016. View Article : Google Scholar
|
|
54
|
Wu DW, Huang CC, Chang SW, Chen TH and Lee
H: Bcl-2 stabilization by paxillin confers 5-fluorouracil
resistance in colorectal cancer. Cell Death Differ. 22:779–789.
2015. View Article : Google Scholar :
|
|
55
|
Howe EN, Cochrane DR, Cittelly DM and
Richer JK: miR-200c targets a NF-κB up-regulated TrkB/NTF3
autocrine signaling loop to enhance anoikis sensitivity in triple
negative breast cancer. PLoS One. 7:e499872012. View Article : Google Scholar
|
|
56
|
Taniguchi T, Tischkowitz M, Ameziane N,
Hodgson SV, Mathew CG, Joenje H, Mok SC and D'Andrea AD: Disruption
of the Fanconi anemia-BRCA pathway in cisplatin-sensitive ovarian
tumors. Nat Med. 9:568–574. 2003. View
Article : Google Scholar : PubMed/NCBI
|
|
57
|
Helleman J, van Staveren IL, Dinjens WN,
van Kuijk PF, Ritstier K, Ewing PC, van der Burg ME, Stoter G and
Berns EM: Mismatch repair and treatment resistance in ovarian
cancer. BMC Cancer. 6:2012006. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Kawahara T, Ide H, Kashiwagi E, Patterson
JD, Inoue S, Shareef HK, Aljarah AK, Zheng Y, Baras AS and Miyamoto
H: Silodosin inhibits the growth of bladder cancer cells and
enhances the cytotoxic activity of cisplatin via ELK1 inactivation.
Am J Cancer Res. 5:2959–2968. 2015.PubMed/NCBI
|
|
59
|
Jung JG, Shih IM, Park JT, Gerry E, Kim
TH, Ayhan A, Handschuh K, Davidson B, Fader AN, Selleri L and Wang
TL: Ovarian cancer chemoresistance relies on the stem cell
reprogramming factor PBX1. Cancer Res. 76:6351–6361. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Yang C, Yin L, Zhou P, Liu X, Yang M, Yang
F, Jiang H and Ding K: Transcriptional regulation of IER5 in
response to radiation in HepG2. Cancer Gene Ther. 23:61–65. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Yu Y, Guo M, Wei Y, Yu S, Li H, Wang Y, Xu
X, Cui Y, Tian J, Liang L, et al: FoxO3a confers cetuximab
resistance in RAS wild-type metastatic colorectal cancer through
c-Myc. Oncotarget. 7:80888–80900. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Sannigrahi MK, Singh V, Sharma R, Panda NK
and Khullar M: Role of autophagy in head and neck cancer and
therapeutic resistance. Oral Dis. 21:283–291. 2015. View Article : Google Scholar
|
|
63
|
Peitzsch C, Perrin R, Hill RP, Dubrovska A
and Kurth I: Hypoxia as a biomarker for radioresistant cancer stem
cells. Int J Radiat Biol. 90:636–652. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Bivin WW, Yergiyev O, Bunker ML, Silverman
JF and Krishnamurti U: GRB7 expression and correlation with HER2
amplification in invasive breast carcinoma. Appl Immunohistochem
Mol Morphol. 25:553–558. 2017. View Article : Google Scholar
|
|
65
|
Nencioni A, Cea M, Garuti A, Passalacqua
M, Raffaghello L, Soncini D, Moran E, Zoppoli G, Pistoia V, Patrone
F and Ballestrero A: Grb7 upregulation is a molecular adaptation to
HER2 signaling inhibition due to removal of Akt-mediated gene
repression. PLoS One. 5:e90242010. View Article : Google Scholar : PubMed/NCBI
|