Introduction
Axon guidance molecules modulate the growth and
migration of neurons during development of the neural system. These
molecules are classified according to their genetic and biochemical
properties into four highly conserved families; netrins, slits,
semaphorins and ephrins (1). Unc-5
Netrin Receptor (UNC5)C is one of seven cognate receptors,
including neogenin, deleted in colorectal cancer (DCC), UNC5A,
UNC5B, UNC5C, UNC5D and adenosine A2b receptor, for netrin-1
(NTN1), which mediates the directed extension and migration of
axons during neural development (2). UNC5C mediates the repellent response
to NTN1. UNC5C-deletion in mice disrupts the long-range dorsal
guidance of inferior olivary and pontine axons after crossing the
midline (3). UNC5C is also
involved in tumor progression (4).
Thiebault et al (5) used a
full-length complementary DNA (cDNA) probe that recognizes UNC5A,
UNC5B and UNC5C mRNA, and found that UNC5A, UNC5B and UNC5C were
significantly downregulated in 93, 88, 49, 48, 68 and 74% of
colorectal, ovarian, breast, uterine, gastric and lung cancer
samples, respectively, indicating that UNC5A, UNC5B and UNC5C are
potential tumor suppressor genes. UNC5 receptors are members of the
dependence receptor family, which elicit an apoptotic signal in the
absence of their ligand, NTN1, instead of being inactive (5). The loss of function of UNC5C
typically occurs in early stages of colorectal cancer (6), and inherited UNC5C mutations can
inhibit cell apoptosis and increase the risk of colorectal cancer
(7). UNC5A, UNC5B and UNC5C are
downregulated in colorectal cancer by 48, 27 and 74–77%,
respectively (5), suggesting that
UNC5C plays an important inhibitory role in colorectal cancer. In
addition to the loss of heterozygosity, several studies have
attributed UNC5C downregulation to the abnormal methylation of its
promoter (5,8).
UNC5 receptors perform functions through
interactions with other axon guidance molecule receptors. For
example, UNC5B was shown to interact with the netrin-4 receptor
neogenin (9) or roundabout
guidance receptor 4 (10) to
inhibit angiogenesis. UNC5B also interacts with DCC to convert
NTN1-induced growth cone attraction to repulsion (11). However, the impact of these
interactions on tumor progression remains unknown.
Breast cancer is the most malignant type of cancer
in females, and it is difficult to treat due to its high rates of
recurrence and mortality. Breast cancer alone accounted for 11.6%
of all cancer cases and 6.6% of all cancer-associated mortalities
among females in 2018 (12).
Metastasis is the leading cause of mortality in breast cancer
patients (13). Most studies on
the role of UNC5C in tumorigenesis have focused on colorectal
cancer (7,8,14).
To the best of our knowledge, the function and mechanism of UNC5C
in breast cancer have not been widely reported. Fitamant et
al (15) found that metastatic
breast cancer expresses NTN1 as a mechanism by which breast cancer
cells escape apoptosis.
The present study investigated the effects of UNC5C
on cell growth and metastasis both in vitro and in
vivo by knocking down and overexpressing UNC5C in breast cancer
cell lines. It was identified that UNC5C interacted with the
integrin α6 subunit and UNC5C-knockdown enhanced the growth and
metastasis of breast cancer cells, which was likely partially
attributable to the upregulation of matrix metalloproteinase (MMP)9
expression via the NTN1/integrin α6/β4 signaling pathway. These
findings confirm the inhibitory effects of UNC5C on breast cancer
cell viability and metastasis.
Materials and methods
Cell lines and reagents
The SK-BR-3, ZR-75-30 and MDA-MB-231 breast cancer
cell lines were purchased from the American Type Culture
Collection. 293T, MCF-10a and MCF-7 cells were purchased from the
Cell Resource Center, Institute of Life Sciences, Chinese Academy
of Science. All cell lines were cultured in Dulbecco's modified
Eagle's medium (DMEM, HyClone; GE Healthcare) with 10% fetal bovine
serum (FBS; Gibco; Thermo Fisher Scientific, Inc.). All cells were
incubated in a 5% CO2 humidified atmosphere at 37°C. The
lentivirus vector pll3.7 for UNC5C-knockdown and packaging
plasmid pHR were obtained from Addgene, Inc. pVSVG was
obtained from Invitrogen; Thermo Fisher Scientific, Inc. The
pbobi plasmids for UNC5C (UNC5C full length gene sequence
ID: 8633) and integrin α6 subunit (ITGA6 full length gene sequence
ID: 3655) overexpression were provided by Professor Jiahuai Han
(Xiamen University, Xiamen, China). The mitogen activated protein
kinase (MAPK) inhibitors SB203580, U0126 and LY294002 were
purchased from Merck KGaA. Breast tissue chips that contained 30
pairs of tumor and adjacent tissues (cat. no. HBre-Duc060CS-02)
were purchased from Shanghai Outdo Biotech Co., Ltd.
Antibodies
Anti-UNC5C (cat. no. ab106949) and integrin α6 (cat.
no. ab20142 for immunoprecipitation and ab181551 for western
blotting) antibodies were purchased from Abcam. Anti-phosphorylated
(p)-Akt (cat. no. 4060; Ser473), Akt (cat. no. 4685), p-p38 MAPK
(clone D3F9; cat. no. 4511), p38 MAPK (clone D13E1; cat.no. 8690),
p-ERK1/2 (Thr202/Tyr204; cat. no. 9101), ERK1/2 (cat. no. 9102),
Src (cloneL4A1; cat. no. 2110), p-Src (Tyr416; cat. no. 2101), FAK
(cat. no. 3285) and p-FAK (clone D20B1; Tyr397; cat. no. 8556)
antibodies were purchased from Cell Signaling Technology, Inc.
Anti-α-tubulin (cat. no. T6199) and Flag (cat. no. F3040)
antibodies were purchased from Sigma-Aldrich; Merck KGaA. Anti-myc
antibody (cat. no. HT101) was purchased from Transgene SA.
Anti-integrin-linked kinase (ILK; clone E-2, cat. no. sc-137221)
and p-ILK antibodies (Thr173; cat. no. sc-130196) were purchased
from Santa Cruz Biotechnology, Inc. Goat anti-mouse horseradish
peroxidase (HRP)-conjugated immunoglobulin G (IgG; cat. no.
G-21040) and goat anti-rabbit HRP-conjugated IgG (cat. no. A16096)
were purchased from Pierce; Thermo Fisher Scientific, Inc.
Establishment of stable cell lines
The stable cell lines with UNC5C-knockdown or
overexpression were established as previously described (16). Briefly, virus stocks were prepared
by co-transfecting pll3.7 (for UNC5C mRNA knockdown) or
pbobi (cMyc tag; for UNC5C mRNA overexpression) with two
packaging plasmids (pHR and pVSVG) into 293T cells.
For UNC5C-knockdown, SK-BR-3 and ZR-75–30 cells were infected with
pll3.7 in the presence of polybrene (8 µg/ml;
Sigma-Aldrich; Merck KGaA) for 2 days. Infected cells were screened
by treatment with G418 (1,500 µg/ml; Sigma-Aldrich; Merck
KGaA) for 1 week. The targeted sequences for UNC5C-knockdown were
as follows: shRNA1, 5'-TCTTGGATTGCAAGACGAGG-3'; and shRNA2,
5'-CAAAGTCACGATGATTCTTC-3'. Empty plasmid was used as a control
(shCtrl). For UNC5C overexpression (UNC5C full length gene sequence
ID: 8633), MDA-MB-231 cells were infected with pbobi in the
presence of polybrene, and screened by treatment with puromycin
(2.5 µg/ml; Sigma-Aldrich; Merck KGaA) for 1 week. After the
screened cells were maintained in culture for 1 week
UNC5C-knockdown and overexpression efficiency were determined by
western blotting.
Western blotting and
immunoprecipitation
The MCF-10a, MCF-7, SK-BR-3, ZR-75–30 and MDA-MB-231
breast cancer cells were lysed with RIPA buffer (cat. no. P0013B;
Beyotime Institute of Biotechnology) that contained a protease
inhibitor (cat. no. 04906845001; Roche Diagnostics). Isolated
proteins were quantified using the Pierce™ BCA Protein assay kit
(cat. no. 23225; Thermo Fisher Scientific, Inc.). Protein samples
(30 µg) were fractionated by 7.5% SDS-PAGE and transferred to a
polyvinylidene difluoride membrane (EMD Millipore). After blocking
in 5% milk for 1 h at room temperature, membranes were incubated at
4°C overnight with primary antibodies at a dilution of 1:1,000 for
anti-phosphorylated (p)-Akt, Akt, p-p38 MAPK, p38 MAPK, p-ERK1/2,
ERK1/2, Src, p-Src, FAK, p-FAK, Unc5C and integrin α6, and at a
dilution of 1:500 for anti-integrin-linked kinase and p-ILK, The
membranes were then washed and incubated with the appropriate
HRP-conjugated secondary antibody (1:5,000) for 1 h at room
temperature. The bound antibody was developed using an enhanced
chemiluminescence system (Bio-Rad Laboratories, Inc.). The density
of the protein bands was analyzed by ImageJ software (version
1.52a; National Institutes of Health). For immunoprecipitation, the
cells were rinsed three times with ice-cold PBS, NP-40 buffer (cat.
no. P0013F; Beyotime Institute of Biotechnology) that contained
phenylmethylsulfonyl fluoride (cat. no. M145-5; Amresco, LLC) was
then added to the cells. The cell lysates were centrifuged at
12,000 × g for 20 min at 4°C. The supernatants were incubated with
the primary antibodies anti-myc, anti-Flag, anti-Unc5C and
anti-integrin α6 (all 1:500) at 4°C for 2 h, and then incubated
with protein G agarose beads (cat. no. 11719416001; Roche
Diagnostics) overnight at 4°C. After centrifugation and aspirating
the supernatants, the beads were washed 3–6 times, followed by
routine western blot analysis. Non-specific normal IgG (1:200; cat.
no. ab200699; Abcam) was used as a negative control.
Oncomine analysis
The raw data of Unc5C mRNA expression in ductal
breast carcinoma and normal breast tissues were extracted from
three datasets in the Oncomine database (https://www.oncomine.org/resource/login.html),
including the Perou et al (17), Sørlie et al (18) and Richardson et al (19) datasets. The threshold in the
analysis was set at P<1x10−4, fold change >2 and
gene rank in top 10%. The raw data were re-plotted using GraphPad
Prism software (version 6; GraphPad Software, Inc.).
Immunohistochemistry
The breast tissue chips with different
clinicopathological stages (grade II–III) were deparaffinized and
incubated in citrate buffer for antigen retrieval for ≥4 h. The
grades were determined according to the guidelines of the Chinese
Society of Clinical Oncology: Breast Cancer (20). After washing three times with PBS,
the chips were blocked with 3% H2O2 in PBS
for 10 min at room temperature, followed by three washes with PBS.
The chips were blocked by TNB blocking buffer at room temperature
for 1–2 h and then incubated with anti-UNC5C antibody (cat no.
ab106949; Abcam; 1:500) overnight at 4°C. After washing with TNT
buffer, the sections were incubated with the appropriate
HRP-conjugated secondary antibody for 1 h at room temperature.
After a 10–30 min wash with DAB solution and mounting, the slides
were visualized under a light microscope (magnification, ×40, ×200
or ×400) and images were captured. The analysis was performed as
described previously (21), with
minor modifications. The UNC5C-positive staining intensity was
scored as: 0, negative; 1, weak; 2, moderate; and 3, strong. The
scoring for the UNC5C stained area was: 0, <5%; 1, 5–25%; 2,
26–50%; 3, 51–75%; and 4, 76–100% of stained tumor cells. The final
staining score was calculated by multiplying the staining intensity
score by the staining area score, with a range between 0 and
12.
Quantitative PCR (qPCR) and
semi-qPCR
RNA isolation from SK-BR-3 and MDA-MB-231 cells was
performed using TRIzol reagent (Invitrogen; Thermo Fisher
Scientific, Inc.). cDNA was prepared using the ReverTra Ace RT-qPCR
kit (Toyobo Life Science) with 1 µg RNA per reaction. RT was
performed at 37°C for 15 min followed by 98°C for 5 min to
inactivate reverse transcriptase activity. qPCR was performed with
the Thunderbird SYBR qPCR mix (Toyobo Life Science) and the
thermocycling conditions were as previously described (22). The primers used are listed in
Table SI. Relative MMP gene
expression was calculated using the 2-ΔΔCq method
(23), normalized to GAPDH and
compared with the MMP3 level.
For semi-qPCR, the amplification conditions were the
following: Initial denaturation at 95°C for 1 min, followed by 38
cycles of 95°C for 30 sec, 58°C for 30 sec and 72°C for 1 min for
MMPs, and 23 cycles under the same conditions for GAPDH. Taq
polymerase (Takara Bio, Inc.) was used as the DNA polymerase. Each
condition was run in triplicate with the primers listed in Table SI. Amplified products were
resolved by 1% agarose gel electrophoresis and visualized by 4S
green plus (Sangon Biotech Co., Ltd.), and then photographed with a
Bio-Imaging system (DNR Bio Imaging Systems). The band intensity of
each lane was analyzed by ImageJ software (version 1.52a; National
Institutes of Health) as described previously (24).
Enzyme-linked immunosorbent assay
(ELISA)
A total of 1×106 cells/well were seeded
in 6-well microplates. The culture medium was replaced with
serum-free fresh DMEM (2 ml/well) 8 h later. After 24 h,
supernatants were collected and centrifuged at 200 × g for 5 min at
room temperature to remove cellular debris. Secreted NTN1 in the
supernatants was measured using an ELISA kit (cat. no. KTE62214;
Abbkine Scientific Co., Ltd.) according to the manufacturer's
protocol. The final optical density of each well was read at 450 nm
using a microplate reader (Multiscan MK3; Thermo Fisher Scientific,
Inc.).
Tumor xenograft and metastasis
Six-week old nude (BALB/c) mice (n=40, 18–22 g, 15
males and 25 females) were purchased from SLRC Laboratory Company
and maintained at the Laboratory Animal Center, Xiamen University
(Xiamen, China). The mice were housed in plastic cages and
maintained in a climate-controlled animal room (23±1°C; 55±5%
humidified atmosphere) with a 12/12 h light-dark cycle. Food and
water were freely available. The protocols for the xenograft and
metastasis experiments were approved by the Animal Ethics Committee
of Xiamen University (approval no. XMULAC20120030). For the
xenograft experiment, plasmid pll3.7 transfected-SK-BR-3
(Unc5C-shCtrl, Unc5C-shRNA1 and Unc5C-shRNA2) and pbobi
transfected-MDA-MB-231 (GFP and Unc5C) cells were detached and
resuspended in serum-free medium. Subsequently, 5.0×106
cells were subcutaneously injected in a volume of 200 µl
into the flank of 6-week-old female nude BALB/c mice (n≥4/group),
which were maintained in a specific-pathogen-free environment. The
mice were palpated every 3 days to monitor tumorigenesis. Tumor
growth was recorded using a Vernier caliper, and the volume of the
tumors was calculated according to the following formula: 0.52 ×
width2 × length. The humane endpoint was set when the
tumor diameters of any mouse exceeded 12 mm, at which point the
experiment was terminated. After 4 weeks, all of the mice were
euthanized by CO2 exposure at a flow rate of 1.2 l/min,
which displaces 20% of the cage volume per minute. Death of the
mice was verified by persistent unconsciousness and no breathing.
The resulting tumors were completely dissected and weighed.
For the analysis of lung metastasis, male nude
BALB/c mice were divided into three groups (n=5). Plasmid
pll3.7 transfected-SK-BR-3 cells (2.0×106) were
injected in a volume of 100 µl PBS into each mouse via the
tail vein. The body weight of the mice was monitored every 3 days.
A 20% reduction of body weight was defined as the humane endpoint.
After 8 weeks, all of the mice were euthanized by CO2
exposure at a flow rate of 1.2 l/min, which displaced 20% of the
cage volume per minute. Death of the mice was verified by
persistent unconsciousness and no breathing. Lung tissues were
harvested and washed twice with PBS. A cytomegalovirus-enhanced
green fluorescent protein reporter cassette was included in the
pll3.7 vector to monitor expression, thus allowing
evaluation of the metastasis of cancer cells by examining
GFP-labeled metastatic tumor nodules in all lung lobes. The lung
tissues were stored in 4% paraformaldehyde at room temperature for
24 h, embedded in paraffin and cut into 5-µm thick sections.
The sections were finally stained with hematoxylin for 3 min and
eosin for 2 min at room temperature, and visualized under a light
microscope (magnification, ×200).
MTT assay
SK-BR-3, ZR-75–30 and MDA-MB-231 cells were seeded
in 96-well plates at a density of 3.0×103/well. MTT
solution (10 µl; Beyotime Institute of Biotechnology) was
then added to each well every 24 h, and the cells were incubated at
37°C for 4 h with 5% CO2. The MTT solution was then
removed and replaced with 150 µl DMSO. The plate was further
incubated at 37°C for 15 min with 5% CO2 and agitated on
an orbital shaker for 10 min. The optical density of the wells was
read at a wavelength of 560 nm with a 630 nm reference filter.
Tumor cell invasion assay
The tumor cell invasion assay was performed as
described previously (16).
Briefly, the invasion chambers (8-µm; EMD Millipore) were
coated with 20 µl diluted Matrigel (BD Biosciences) at 37°C
for 30 min and inserted into a 24-well plate. Cells were added to
the top chambers at 1×104 cells/well. The upper chamber
was filled with 200 µl DMEM that contained 5% FBS, and the
lower chamber was filled with 500 µl DMEM that contained 20%
FBS. After incubation for 24 h, non-invading cells were removed
from the upper chamber with a cotton swab, and invading cells were
fixed with PAF for 10 min at room temperature and stained using 1%
crystal violet for 5 min at room temperature (cat. no. C0775;
Sigma-Aldrich; Merck KGaA). Subsequently, the cells were visualized
under an inverted microscope (magnification, ×20). The number of
cells was counted and calculated using ImagePro Plus 6.0 software
(Media Cybernetics, Inc.). Three identical replicates were
performed for each condition.
Statistical analysis
The statistical analysis was performed using SPSS
software (version 20; IBM Corp.). Unless otherwise indicated, data
are presented as the mean ± standard error of the mean. No samples
or animals were excluded from the analysis. Comparisons were
performed using Student's t-test for the comparison of two groups
or one-way analysis of variance followed by Tukey's post hoc test
for the comparison of multiple groups. P<0.05 was considered to
indicate a statistically significant difference.
Results
UNC5C is heterogeneously expressed in
breast cancer cell lines
UNC5C expression was detected by western blotting in
various commercially available breast cancer cell lines. The normal
breast cell line MCF-10a was used as a control. As presented in
Fig. 1A, UNC5C was detected in
MCF-10a, MCF-7, SK-BR-3 and ZR-75–30 cells, with the highest UNC5C
expression in ZR-75–30 cells and the lowest expression in
MDA-MB-231 cells. The MCF-7 and ZR-75-30 cells are derived from
ductal breast carcinoma. MDA-MB-231 and SK-BR-3 are derived from
breast adenocarcinoma. The present results indicate that the
heterogeneity of UNC5C expression in various cell lines may depend
on the specific type of cell line. To confirm the results that were
found in MCF-7 and ZR-75-30 cells, UNC5C expression in ductal
breast carcinoma was further examined by Oncomine data-mining
analysis and the immunostaining of breast tissue chips. The
Oncomine data indicated no significant differences between normal
tissues and ductal breast carcinoma (17–19)
(Fig. S1). Breast tissue chips
from normal tissue and ductal breast carcinoma were separately
stained with UNC5C antibodies. UNC5C protein was widely expressed
in most ductal breast carcinoma and adjacent normal tissues at
different clinical stages, and no significant difference was
observed between them (Fig. 1B).
UNC5 receptors are NTN1 dependence receptors (5). Therefore, ELISA was used to detect
the relative expression of NTN1 in culture media of the
aforementioned breast cancer cell lines. NTN1 expression in SK-BR-3
and ZR-75-30 cells, which express a relatively high level of UNC5C,
was significantly higher than in MDA-MB-231 cells and not detected
in MCF7 cells (Fig. 1C). NTN1
expression appears to be positively associated with the UNC5C
receptor.
 | Figure 1UNC5C is heterogeneously expressed in
clinical samples and breast cancer cell lines. (A) Detection of
relative UNC5C expression in MCF-10a, MCF-7, SK-BR-3, ZR-75–30 and
MDA-MB-231 breast cancer cell lines by western blotting. The
expression of UNC5C was first normalized to α-tubulin and then to
the value in the MCF-10a normal breast cell line.
*P<0.05, **P<0.01 vs. MCF-10a. (B)
Detection of UNC5C expression in breast tissue microarrays at
different clinical stages by immunostaining. Left panels, whole
tissue chip (magnification, ×40; scale bar, 200 µm). Middle
panels, part of the tissue chip (magnification, ×200; scale bar,
100 µm). Right panels, part of the tissue chip
(magnification, ×400; scale bar, 50 µm). Significant
differences were determined by paired Student's t-test. (C)
Detection of relative NTN1 expression in the culture medium of
breast cancer cell lines by ELISA. **P<0.01 vs.
MDA-MD-231. Data are expressed as the mean ± standard deviation.
NS, not significant; Adj, adjacent tissues; Ca, cancer; UNC5C,
Unc-5 Netrin Receptor C. |
UNC5C inhibits the viability and invasion
of breast cancer cells
To evaluate the function of UNC5C in breast cancer
cells, UNC5C-knockdown experiments were performed in SK-BR-3 and
ZR-75-30 cells, and UNC5C-overexpression experiments were performed
with MDA-MB-231 cells. Two RNA sequences were designed and inserted
into the pll3.7 vector to knockdown UNC5C expression in
SK-BR-3 and ZR-75-30 cells. An empty control vector was used as a
control. Plasmid pbobi that encoded the full-length human
UNC5C with a cMyc tag sequence was transfected into MDA-MB-231
cells. The plasmid with the GFP coding sequence was used as a
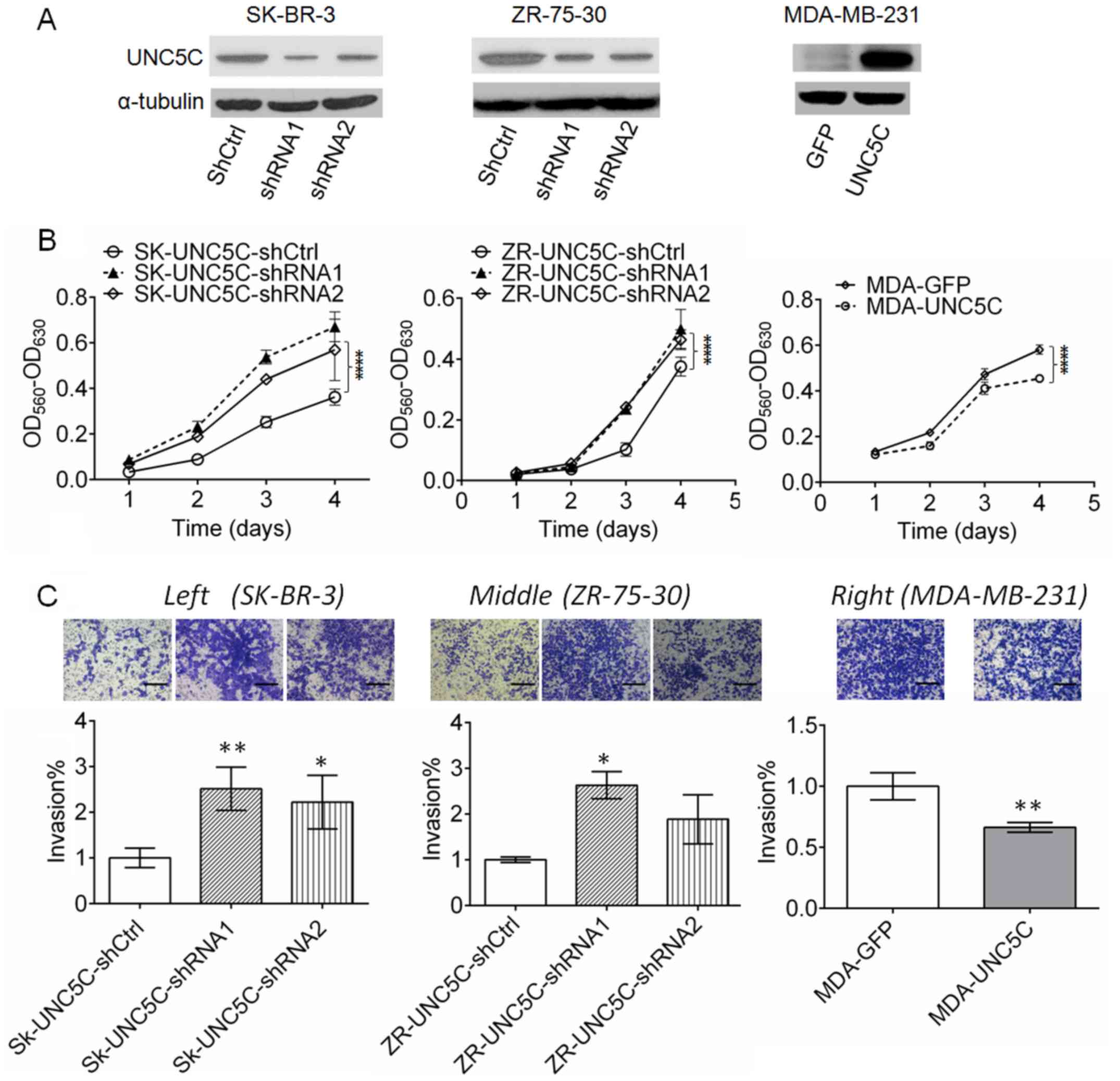
control. As shown in Fig. 2A,
UNC5C expression was effectively downregulated in SK-BR-3 and
ZR-75-30 cells, whereas UNC5C was markedly overexpressed in
MDA-MB-231 cells. To examine the effects of UNC5C expression on
NTN1, the concentration of NTN1 in culture media of these breast
cancer cells was detected by ELISA. The results demonstrated that
UNC5C-knockdown increased the concentration of NTN1, and
UNC5C-overexpression decreased NTN1 concentrations; however, these
changes were not statistically significant for cells transfected
with SK-UNC5C-shRNA1 or pbobi overexpression vector
(Fig. S2). Notably, NTN1
expression was not assessed by western blotting or RT-qPCR, which
may be a limitation of the present study.
The downregulation of UNC5C significantly enhanced
the viability of SK-BR-3 cells (P<0.001) and ZR-75-30 cells
(P<0.001). By contrast, the viability of MDA-MB-231 breast
cancer cells was significantly suppressed by UNC5C-overexpression
(P<0.001). These results indicate that UNC5C inhibits the
viability of breast cancer cells (Fig.
2B). To examine the effects of UNC5C on tumor invasion, the
Matrigel coated-Transwell assay was conducted. The number of cells
that passed through the Matrigel matrix was significantly increased
following UNC5C knockdown (P<0.01 for SK-BR-3-shRNA1 and
P<0.05 for ZR-75-30-shRNA1) and reduced by UNC5C overexpression
(P<0.01), suggesting that UNC5C suppresses tumor cell invasion
(Fig. 2C).
UNC5C suppresses the growth and
metastasis of breast cancer cells in vivo
To confirm the effects of UNC5C on tumor growth
in vivo, a subcutaneous tumor xenograft assay was performed
with SK-BR-3 and MDA-MB-231 cells. As presented in Fig. 3A, UNC5C-knockdown in SK-BR-3 cells
significantly enhanced tumor growth (P<0.001 for tumor volume
and P<0.01 for final tumor weight; n=5). By contrast,
UNC5C-overexpression (Fig. 3B)
resulted in the opposite effect in MDA-MB-231 cells (P<0.001 for
tumor volume and P<0.01 for final tumor weight; n=4).
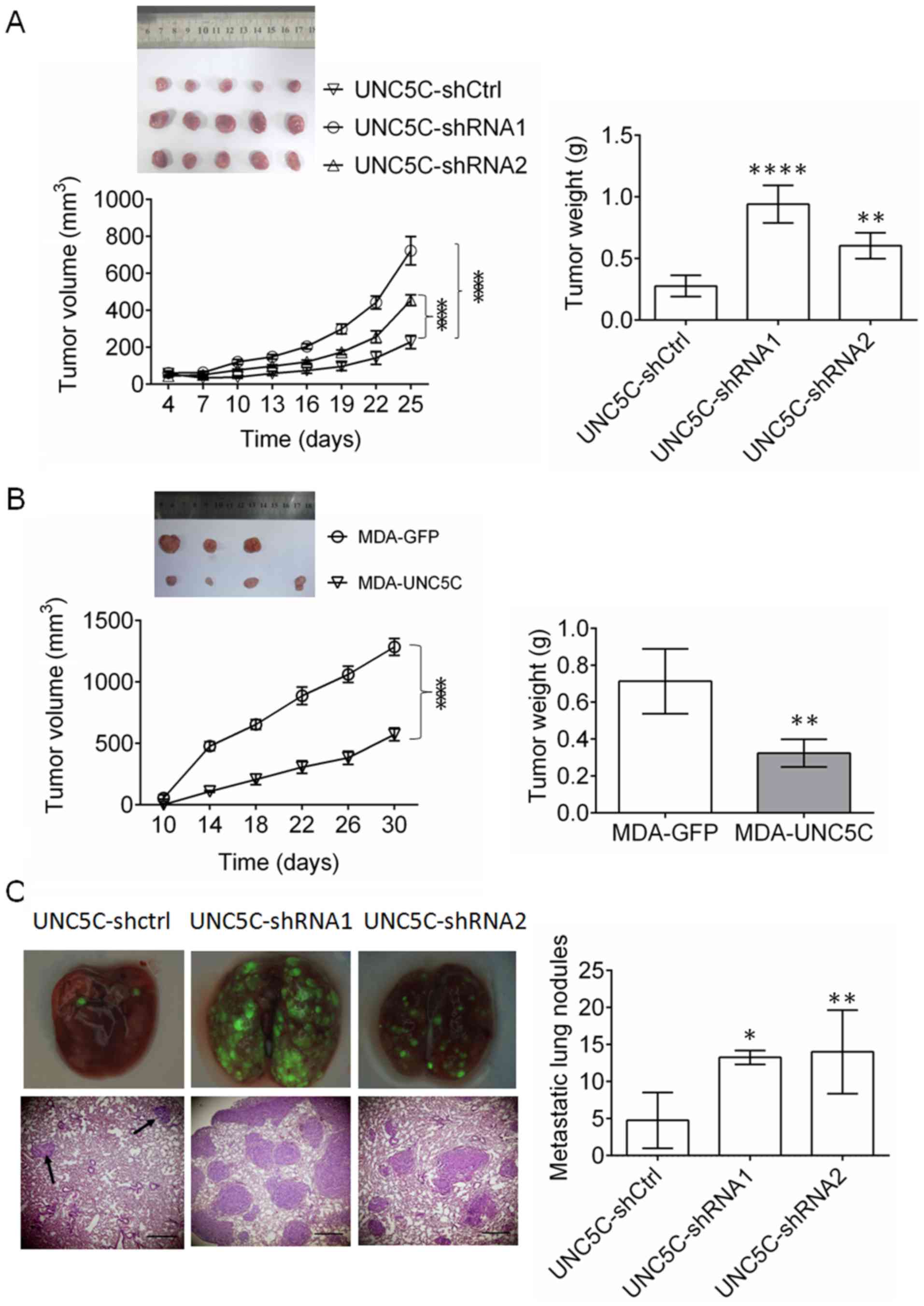 | Figure 3UNC5C suppresses the growth and
metastasis of breast cancer in vivo. For the tumor growth
assay, (A) SK-BR-3 cells (n=5) or (B) MDA-MB-231 cells (n=4) were
inoculated subcutaneously in the right flanks of 4-week-old nude
BALB/c mice. The volume of the resulting tumors was measured every
3 or 4 days according to the formula: Volume = width2 ×
length × 0.52. One month later, the resulting tumors were
completely dissected and weighed. The maximum diameter of the tumor
that formed at the end of the experiments was 9.2 mm in the
SK-UNC5C-Ctrl group, 12.4 mm in the SK-UNC5C-shRNA1 group, 12.3 mm
in the SK-UNC5C-shRNA2 group, 12.2 mm in the MDA-MB-231-GFP group
and 10.2 mm in the MDA-MB-231-Unc5C group. (C) For tumor
metastasis, SK-BR-3 cells were injected intravenously. Metastatic
tumor nodules in all lung lobes were photographed under a
fluorescence microscope and counted in the paraffin sections.
Magnification of upper panels, ×40. Magnification of lower panels,
×200. The data are expressed as the mean ± standard deviation.
*P<0.05, **P<0.01,
****P<0.0001 vs. UNC5C-shCtrl or MDA-GFP. UNC5C,
Unc-5 Netrin Receptor C; sh, short hairpin; Ctrl, control. |
To investigate the effects of UNC5C on tumor
metastasis in vivo, SB-BR-3 cells were injected in the tail
vein in mice. UNC5C-knockdown in SK-BR-3 cells significantly
promoted tumor metastasis (P<0.05 for shRNA1 and P<0.01 for
shRNA2; n=5; Fig. 3C). These
findings confirm that UNC5C also inhibits the growth and metastasis
of breast cancer cells in vivo.
UNC5C suppresses the expression of MMPs
in breast cancer cells
For tumor invasion and metastasis, tumor cells must
traverse the basement membrane that lines the basal face of tumor
cells, which contains collagen type IV as a major structural
component (25). Secreted MMPs
that can degrade collagen type IV, including MMP2, MMP3, MMP7, MMP9
and MMP10, of which MMP2 and MMP9 are the most efficient (26), play critical roles in tumor
invasion and metastasis. MMP3, MMP7, MMP9 and MMP10 expression was
upregulated by UNC5C-knockdown in SK-BR-3 cells and downregulated
by UNC5C-overexpression in MDA-MB-231 cells. However, no difference
in MMP2 expression was detected in either UNC5C-knockdown or
-overexpressing cells (Fig. 4A).
Western blotting further confirmed MMP2 and MMP9 expression at the
protein level (Fig. 4B). These
findings indicated that UNC5C inhibits the invasion and metastasis
of breast cancer cells, which is associated with suppression of the
expression and activity of MMP3, MMP7, MMP9 and MMP10.
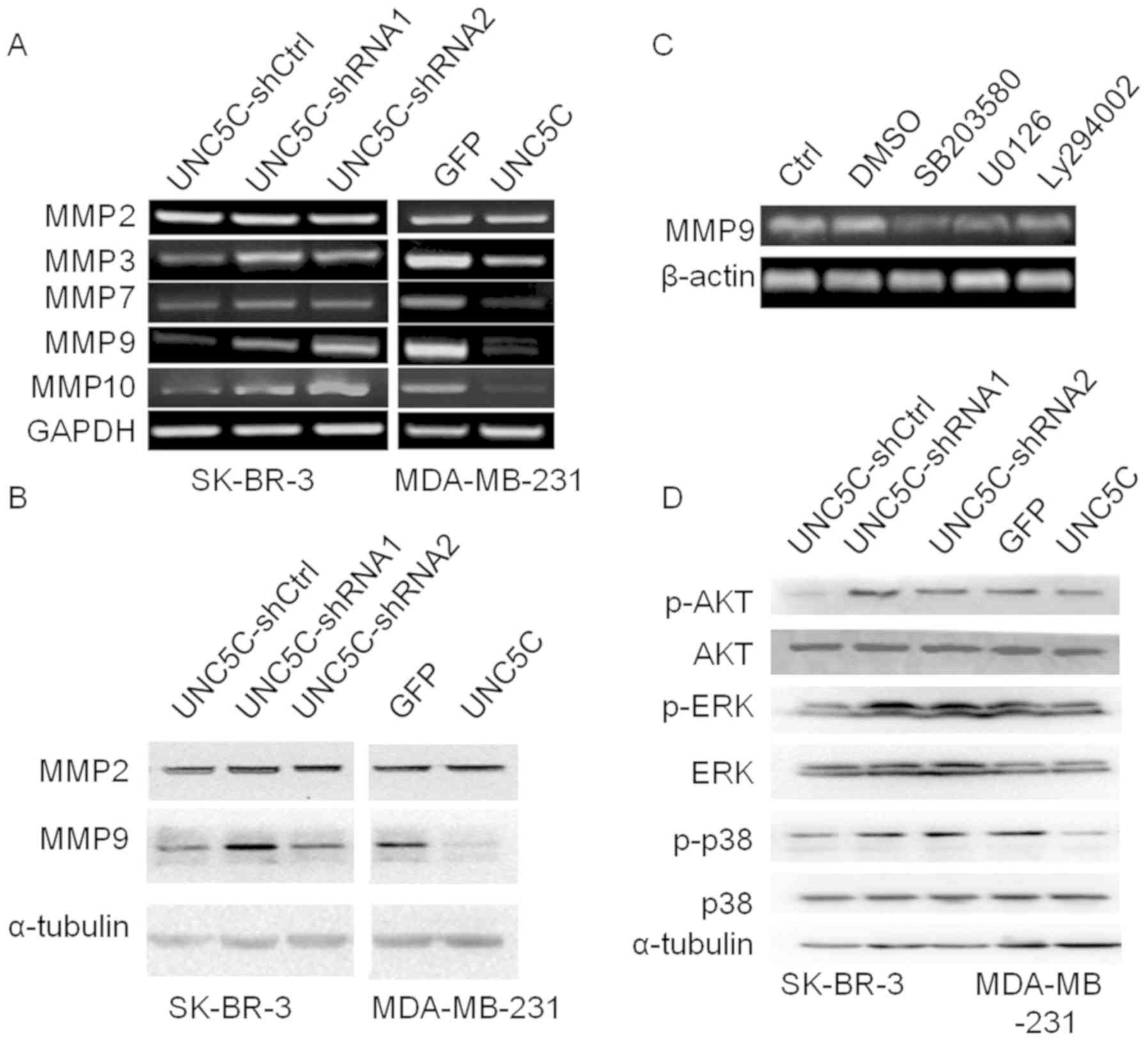 | Figure 4UNC5C suppresses MMP expression in
breast cancer cells and inhibits the phosphorylation of AKT,
MAPK/ERK and MAPK/p38. (A) Semi-quantitative PCR analysis.
UNC5C-knockdown in SK-BR-3 cells enhanced and overexpression in
MDA-MB-231 cells inhibited the expression of MMP3, MMP7, MMP9 and
MMP10, but not MMP2. (B) Western blotting. UNC5C-knockdown
upregulated MMP9 expression, but not MMP2. UNC5C-overexpression
inhibited MMP9 expression at the protein level. (C) MMP9 expression
in SK-BR-3 cells was markedly suppressed when the PI3K/AKT, ERK and
p38 signaling pathways were blocked. (D) UNC5C-knockdown enhanced
the phosphorylation of AKT, ERK1/2 and p38. UNC5C, Unc-5 Netrin
Receptor C; sh, short hairpin; Ctrl, control; MMP, matrix
metalloproteinase; p, phosphorylated. |
UNC5C suppresses the phosphorylation of
PI3K/AKT, ERK and p38 MAPK
Based on the findings that MMP9 expression is
maintained at a higher level than MMP3, MMP7 and MMP10 (Fig. S3) and that MMP9, not MMP2, is
affected by UNC5C (Fig. 4A and B),
we focused on MMP9 to further investigate the effects of UNC5C on
breast cancer cell invasion and metastasis. Several studies have
shown that the activation of PI3K/AKT (27), ERK (28) and p38 MAPK (29) is important for the regulation of
MMP expression in breast cancer cells. To examine the possibility
that MMP9 expression is regulated by the PI3K/AKT, ERK or p38 MAPK
signaling pathways, PI3K (LY294002), ERK (U0126) and p38 MAPK
(SB203580) inhibitors were used to investigate their influence on
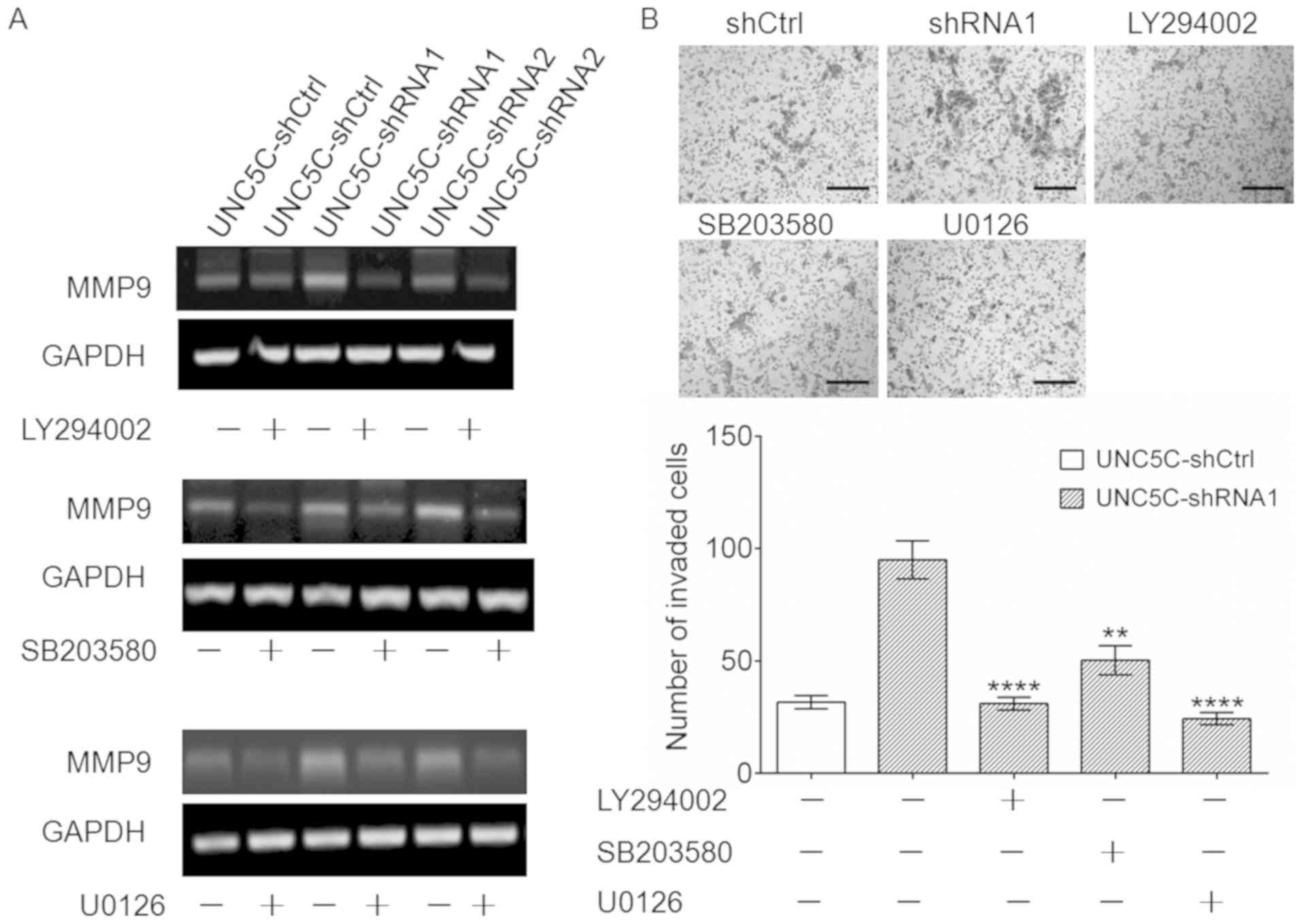
MMP9 expression. As shown in Fig.
4C, MMP9 expression was markedly suppressed when the PI3K/AKT,
ERK and p38 MAPK signaling pathways were blocked. Notably, these
inhibitors antagonized the upregulation of MMP9 (Figs. 5A and S4) and increased invasion ability
(Fig. 5B) that was induced by
UNC5C-knockdown. These results, combined with the findings that
phosphorylation levels of AKT, ERK and p38 MAPK increased after
UNC5C-knockdown in SK-BR-3 cells and decreased after
UNC5C-overexpression in MDA-MB-231 cells (Fig. 4D), suggest that UNC5C likely exerts
inhibitory effects on breast cancer metastasis at least partially
by inhibiting MMP9 expression through the PI3K/AKT, ERK and p38
MAPK signaling pathways.
UNC5C inhibits the growth and metastasis
of breast cancer cells through interactions with integrin α6
A notable characteristic of NTN1 receptors is their
mutual interactions (9–11). Integrins comprise a family of
receptors that usually mediate cell to extracellular matrix (ECM)
adhesion and migration. Integrin signaling pathways play crucial
roles in regulation of MMP expression in various cancer types
(30–32). NTN1 has been reported to promote
epithelial cell adhesion and migration during development of the
pancreas by binding to integrin α6/β4 and integrin α3β1, which have
been identified as NTN1 receptors (33). Integrin α6/β4 is normally expressed
in the breast epithelium and upregulated in invasive breast cancer
(34). One possibility is that
UNC5C may regulate metastasis in breast cancer through interactions
with integrin α6/β4 signaling pathways. Therefore, the present
study examined interactions between UNC5C and integrin α6.
Immunoprecipitation demonstrated that UNC5C interacted with
integrin α6 both exogenously (Fig.
6A) and endogenously (Fig.
6B). UNC5C also interacted with integrin α6 through
extracellular domains (Fig. 6C).
The effects of UNC5C on integrin α6 expression were then examined
and the results showed that UNC5C-knockdown did not affect integrin
α6 expression in SK-BR-3 cells. Unexpectedly, UNC5C-overexpression
in MDA-MB-231 cells enhanced integrin α6 expression (Figs. 6D and S5).
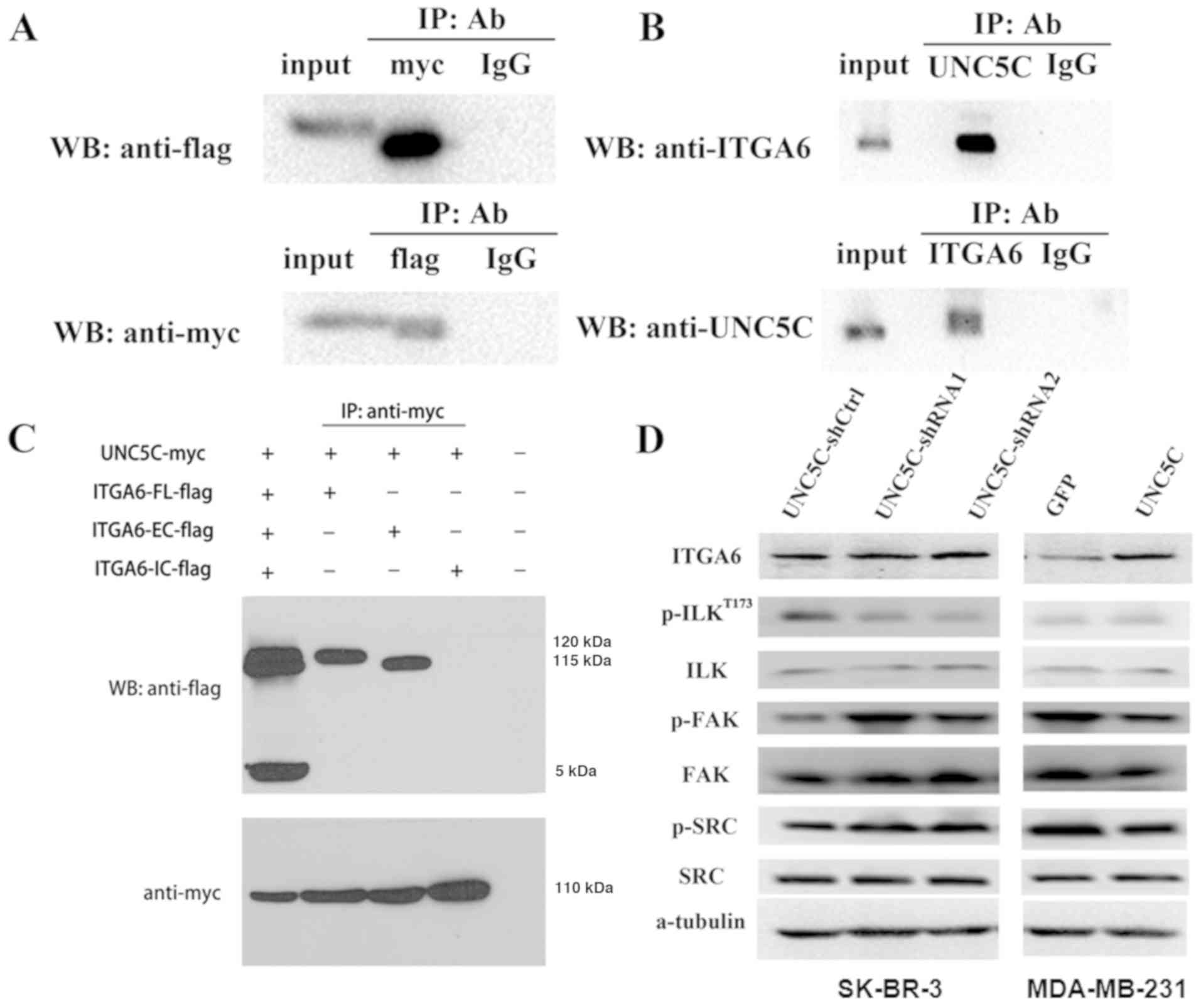 | Figure 6UNC5C activates integrin
α6-associated signaling through interactions with integrin α6. (A)
UNC5C interacts with integrin α6 exogenously. 293T cells were
co-transfected with a UNC5C-Myc plasmid and integrin α6-Flag
plasmid. Immunoprecipitation was performed with MYC and Flag
antibody separately. (B) Immunoprecipitation was performed with
antibodies against UNC5C and integrin α6 in SK-BR-3 cells for the
endogenous interaction analysis. (C) UNC5C interacts with integrin
α6 through an extracellular domain. 293T cells were transfected
with UNC5C-Myc, integrin α6-FL-Flag, integrin α6-EC-Flag and
integrin α6-IC-Flag in the indicated combinations.
Immunoprecipitation was performed to precipitate MYC. (D) Western
blotting. UNC5C-knockdown exerted no significant effects on
integrin α6 in SK-BR-3 cells, decreased ILK phosphorylation, and
increased FAK and SRC phosphorylation. Whereas UNC5C-overexpression
upregulated integrin α6 expression, increased ILK phosphorylation
and decreased FAK and SRC phos-phorylation. FL, full length; EC,
extracellular; IC, intracellular; WB, western blot; UNC5C, Unc-5
Netrin Receptor C; ITGA6, integrin α6; p, phosphorylated; ILK,
integrin-linked kinase; sh, short hairpin; Ctrl, control; IP,
immunoprecipitate; Ab, antibody. |
ILK, FAK and SRC kinases play critical roles in
integrin signaling (35,36). To investigate the effects of UNC5C
on integrin signaling, the present study examined the
phosphorylation of ILK, FAK and SRC kinases. The results showed
that ILK phosphorylation at Thr-173 decreased following
UNC5C-knockdown in SK-BR-3 cells. UNC5C-overexpression in
MDA-MB-231 cells increased the phosphorylation of ILK, although no
significant difference was found. By contrast, opposite results
were revealed for FAK and SRC phosphorylation (Fig. 6D and S5).
Discussion
A unique characteristic of dependence receptors is
their ability to perform dual opposing roles by activating opposite
signaling pathways. In the presence of their cognate ligand, they
trigger the activation of signaling pathways that are involved in
cell survival, migration and differentiation. In the absence of the
ligand, the receptors elicit apoptosis-inducing signals (37). The heterogeneous expression of
UNC5C was observed in various breast cancer cell lines in the
present study (Fig. 1A). The
results were consistent with Thiebault et al (5), who examined UNC5C expression in 53
breast cancer samples and corresponding normal samples. They found
a ≥2-fold lower expression in 49% of breast tumors, indicating the
heterogeneity of UNC5C expression in breast cancer samples. For
cancer cells without or with low levels of the cognate ligand NTN1,
such as the MCF-7 and MDA-MB-231 cell lines, to avoid the induction
of apoptosis, UNC5C expression was downregulated or silenced,
supposedly through promoter methylation or the loss of
heterogeneity. Conversely, for cells that expressed NTN1, such as
the SK-BR-3 and ZR-75-30 cell lines, the NTN1/Unc5C signaling
pathway transduced anti-apoptotic signals for survival or other
unknown aspects of cancer. Thus, dependence receptors, such as
UNC5C, are also termed 'conditional tumor suppressors'. Indeed, the
NTN1/Unc5 signaling pathway transmits signals for cancer survival,
which may be a self-rescuing reaction of cancer cells that are
affected by chemotherapy (38).
Several chemotherapeutic agents, such as doxorubicin,
5-fluorouracil, paclitaxel and cisplatin, trigger an increase in
the expression of NTN1 and its receptors, such as DCC and UNC5, in
various human cancer cell lines (38).
No significant difference was found between clinical
normal tissues and ductal breast carcinoma in the present study.
This would be expected because both normal and abnormal
physiological activities may need, depending on the tissue context,
the 'conditional tumor suppressor' function of UNC5C to eliminate
unnecessary cells in the absence of NTN1, and also need survival
signals that are elicited by the NTN1/UNC5C pathway (Fig. 1B) to control the number of cells in
tissues or to regulate directed migration, which usually occurs
during neural development.
Interference with NTN1 was reported to induce
apoptosis and inhibit metastasis in various cancer types (15,39).
Unexpectedly, UNC5C-knockdown in SK-BR-3 and ZR-75-30 cells that
expressed NTN1 enhanced tumor growth and metastasis both in
vitro (Fig. 2) and in
vivo (Fig. 3), indicating that
NTN1 and UNC5C exert opposite effects on tumor growth and
metastasis. One unresolved issue is why UNC5C-knockdown enhanced
tumor growth and metastasis in SK-BR-3 and ZR-75-30 cells even in
the presence of NTN1 (Figs. 1C and
2). It can be hypothesized that
NTN1 mediates a complex signaling process that integrates both
anti-apoptotic and survival signals, leading to the survival and
metastasis of cancer cells. NTN1 binding may limit the
pro-apoptotic role of UNC5C, and UNC5C-knockdown fully releases the
survival signal that is elicited by NTN1, thus enhancing tumor
growth and metastasis. Unknown are the receptors that mediate NTN1
survival signaling when UNC5C is disabled. UNC5B has been reported
to interact with another NTN1-dependence receptor, DCC, to regulate
growth cone orientation. Specifically, DCC attracted and UNC5B
repelled growth cones. However, the DCC-mediated attraction of
growth cones may be converted to repulsion when UNC5B is
overexpressed, and such a conversion may be initiated only by NTN1
(11). Thus, the NTN1 ligand
appears to serve as an integrator that ties the two receptors with
contrasting functions through a process whereby the amount of each
receptor determines the final effects of NTN1, such as either to
attract or repel growth cones. These findings prompted the present
study to explore whether UNC5C interacts with other NTN1 receptors
that are not NTN1-dependence receptors but instead mediate survival
signals to regulate tumor growth and metastasis. Integrin α6/β4,
whose cognate ligand is laminin-5, is one such NTN1 receptor,
although its NTN1-receptor identity was first recognized in the
process of pancreas development (33). In the present study, the
co-immunoprecipitation assay confirmed that UNC5C interacts with
integrin α6 via a mutual extracellular domain in breast cancer
cells (Fig. 6). Although integrin
α6 can bind to β1 or β4 subunits, integrin α6/β4 is normally
expressed in the breast epithelium and upregulated in invasive
breast cancer (34). Accordingly,
it was presumed that UNC5C predominantly interacts with integrin
α6/β4 in breast cancer. Survival signals that are transduced by
integrin α6/β4 for tumor progression have been reported in various
cancer types. For example, knockdown of integrin α6/β4 expression
leads to enhanced apoptosis in breast cancer cells (40) and negates parathyroid
hormone-related protein-promoted survival in prostate cancer cells
(41). MMP9 is crucially important
in tumor invasion and metastasis (42). Although, to the best of our
knowledge, no studies have associated integrin α6/β4 with MMP9
expression to date, the regulation of MMP9 expression by other
integrins has been frequently reported (43–45).
The present study found that UNC5C-knockdown promoted the growth
and metastasis of breast cancer cells, mechanistically at least
partially by upregulating MMP9 expression (Fig. 4A and B). Additionally, NTN1 has
been shown to promote the proliferation (46) and migration (47) of umbilical cord blood-derived
mesenchymal stem cells and notably protects cells from
hypoxia-induced apoptosis through the integrin α6/β4 signaling
pathway (35), indicating the
potential significance of the NTN1/integrin α6/β4 signaling pathway
in regulating tumor growth and metastasis. A reasonable postulation
is that loss of the UNC5C-receptor NTN1/integrin α6/β4 signaling
pathway in SK-BR-3 and ZR-75-30 cells provokes the enhancement of
tumor growth and metastasis. This is not necessarily ascribed to
the upregulation of integrin α6 expression, because UNC5C-knockdown
exerted no significant effects on integrin α6 expression. Notably,
UNC5C-overexpression upregulated integrin α6 expression in
MDA-MB-231 cells, demonstrating the complexity of cell
type-dependent interactions between UNC5C and integrin α6 (Fig. 6D).
The cytoplasmic tails of both UNC5C and integrins
are devoid of enzymatic features. Thus, UNC5C and integrins
transmit signals by associating with adapter and signaling proteins
(48,49). FAK and SRC are key kinases in the
integrin (50) and NTN1/Unc5C
(51) signaling cascades. Previous
studies have found that NTN1 activates both FAK and SRC
phosphorylation (52) and that FAK
and SRC directly phosphorylates UNC5C (51). For integrin signaling, upon
integrin clustering and interactions with the integrin β subunit,
FAK auto-phosphorylates itself, creating a binding site for SRC
whereby SRC kinase subsequently phosphorylates numerous downstream
components (50). The present
results demonstrated that UNC5C-knockdown enhanced FAK and SRC
phosphorylation (Fig. 6D). It can
therefore be proposed that the interaction between UNC5C and
integrin α6 may interfere with either integrin clustering or
binding between FAK and the integrin β4 subunit, thereby affecting
FAK and SRC activation. UNC5C-knockdown prevented such
interference, and led to enhancement of the activation of FAK and
SRC. Notably, crosstalk among FAK and SRC in the p38 MAPK (53), ERK (54) and PI3K/AKT signaling pathways
(55) has been well established.
Specifically, FAK can directly interact with PI3K (55). Furthermore, the p38 MAPK, ERK and
PI3K/AKT signaling pathways have been frequently reported to
regulate MMP9 (27–29). These finding support our hypothesis
that UNC5C-knockdown enhances MMP9 through the integrin α6/β4
signaling pathway by activating the FAK/SRC, p38 MAPK, ERK and
PI3K/AKT signaling pathways.
Although controversial, ILK has been proposed to be
both a scaffold protein and serine/threonine kinase in the integrin
signaling cascade. ILK activation is PI3K-dependent (36). Although numerous studies have
focused on the phosphorylation of ILK substrates, (AKT at Ser-473
and GSK at Ser-9) (36), to the
best of our knowledge, the regulation of ILK activity by direct
phosphorylation remains to be fully demonstrated. However,
phosphorylation at Thr-173, Ser-246 (56) and Ser-343 (57) has been reported to be important for
ILK function. Unexpectedly, the present study revealed that
UNC5C-knockdown did not affect ILK expression, but inhibited ILK
phosphorylation at Thr-173 (Fig.
6D). Based on the crucial role of ILK in stimulating MMP9
expression (58), it can be
hypothesized that, at least in SK-BR-3 cells, ILK phosphorylation
at Thr-173 may not exert an activating effect but rather an
inhibitory effect on ILK function. However, the precise mechanism
requires further research.
In summary, the present study found that UNC5C
exerts an inhibitory effect on the growth and metastasis of breast
cancer cells and that binding with NTN1 limits the effects of
UNC5C. Moreover, UNC5C interference potentiated the survival
signals that were delivered by the NTN1/integrin α6/β4 signaling
pathway and subsequently enhanced tumor growth and metastasis. The
NTN1/UNC5C pathway may be a therapeutic target for metastasized
cancers. Strategies may be developed to exploit the potential
tumor-suppressor role of UNC5C by enhancing its expression.
However, the present study was preliminary research on the function
of UNC5C in breast cancer, and the current study could not conduct
analyses of patient survival between a high-UNC5C group and
low-UNC5C group on a large scale. Given the heterogeneity of UNC5C
expression in both normal and tumor tissues and the critical roles
of integrin α6/β4 in cell-ECM interactions, further studies are
needed on the side effects of enhancing UNC5C expression in normal
tissues.
Supplementary Data
Funding
This work was supported by grants from the National
Natural Science Foundation of China (grant no. 31871375) and
Program of Fuzhou General Hospital for Distinguished Young
Scientists Development (grant no. 2016Q03).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request
Authors' contributions
JY and LY designed the experiments and wrote the
paper. MY and FX performed the RT-qPCR, western blotting,
immunoprecipitation and immunohistochemistry assays. XX and KZ
performed RNAi plasmid construction and functional assays, and
analyzed the data. LL and SZ performed the animal experiments. All
authors read and approved the final manuscript.
Ethics approval and consent to
participate
The animal protocols were approved by the Animal
Ethics Committee of Xiamen University (approval no.
XMULAC20120030).
Patient consent for publication
Competing interests
The authors declare that they have no competing
interests.
Acknowledgments
Not applicable.
References
|
1
|
Klagsbrun M and Eichmann A: A role for
axon guidance receptors and ligands in blood vessel development and
tumor angiogenesis. Cytokine Growth Factor Rev. 16:535–548. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Dillon AK, Jevince AR, Hinck L, Ackerman
SL, Lu X, Tessier-Lavigne M and Kaprielian Z: UNC5C is required for
spinal accessory motor neuron development. Mol Cell Neurosci.
35:482–489. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Kim D and Ackerman SL: The UNC5C netrin
receptor regulates dorsal guidance of mouse hindbrain axons. J
Neurosci. 31:2167–2179. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Chédotal A, Kerjan G and Moreau-Fauvarque
C: The brain within the tumor: New roles for axon guidance
molecules in cancers. Cell Death Differ. 12:1044–1056. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Thiebault K, Mazelin L, Pays L, Llambi F,
Joly MO, Scoazec JY, Saurin JC, Romeo G and Mehlen P: The netrin-1
receptors UNC5H are putative tumor suppressors controlling cell
death commitment. Proc Natl Acad Sci USA. 100:4173–4178. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Mehlen P and Tauszig-Delamasure S:
Dependence receptors and colorectal cancer. Gut. 63:1821–1829.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Coissieux MM, Tomsic J, Castets M, Hampel
H, Tuupanen S, Andrieu N, Comeras I, Drouet Y, Lasset C,
Liyanarachchi S, et al: Variants in the netrin-1 receptor UNC5C
prevent apoptosis and increase risk of familial colorectal cancer.
Gastroenterology. 141:2039–2046. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Hibi K, Mizukami H, Shirahata A, Goto T,
Sakata M and Sanada Y: Aberrant methylation of the netrin-1
receptor genes UNC5C and DCC detected in advanced colorectal
cancer. World J Surg. 33:1053–1057. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Lejmi E, Leconte L, Pédron-Mazoyer S,
Ropert S, Raoul W, Lavalette S, Bouras I, Feron JG, Maitre-Boube M,
Assayag F, et al: Netrin-4 inhibits angiogenesis via binding to
neogenin and recruitment of Unc5B. Proc Natl Acad Sci USA.
105:12491–12496. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Koch AW, Mathivet T, Larrivée B, Tong RK,
Kowalski J, Pibouin-Fragner L, Bouvrée K, Stawicki S, Nicholes K,
Rathore N, et al: Robo4 maintains vessel integrity and inhibits
angiogenesis by interacting with UNC5B. Dev Cell. 20:33–46. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Hong K, Hinck L, Nishiyama M, Poo MM,
Tessier-Lavigne M and Stein E: A ligand-gated association between
cytoplasmic domains of UNC5 and DCC family receptors converts
netrin-induced growth cone attraction to repulsion. Cell.
97:927–941. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Bray F, Ferlay J, Soerjomataram I, Siegel
RL, Torre LA and Jemal A: Global cancer statistics 2018: GLOBOCAN
estimates of incidence and mortality worldwide for 36 cancers in
185 countries. CA Cancer J Clin. 68:394–424. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Redig AJ and McAllister SS: Breast cancer
as a systemic disease: A view of metastasis. J Intern Med.
274:113–126. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Bernet A, Mazelin L, Coissieux MM, Gadot
N, Ackerman SL, Scoazec JY and Mehlen P: Inactivation of the UNC5C
Netrin-1 receptor is associated with tumor progression in
colorectal malignancies. Gastroenterology. 133:1840–1848. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Fitamant J, Guenebeaud C, Coissieux MM,
Guix C, Treilleux I, Scoazec JY, Bachelot T, Bernet A and Mehlen P:
Netrin-1 expression confers a selective advantage for tumor cell
survival in metastatic breast cancer. Proc Natl Acad Sci USA.
105:4850–4855. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Yuan M, Guo H, Li J, Sui C, Qin Y, Wang J,
Khan YH, Ye L, Xie F, Wang H, et al: Slit2 and Robo1 induce
opposing effects on metastasis of hepatocellular carcinoma Sk-hep-1
cells. Int J Oncol. 49:305–315. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Perou CM, Sørlie T, Eisen MB, van de Rijn
M, Jeffrey SS, Rees CA, Pollack JR, Ross DT, Johnsen H, Akslen LA,
et al: Molecular portraits of human breast tumours. Nature.
406:747–752. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Sørlie T, Perou CM, Tibshirani R, Aas T,
Geisler S, Johnsen H, Hastie T, Eisen MB, van de Rijn M, Jeffrey
SS, et al: Gene expression patterns of breast carcinomas
distinguish tumor subclasses with clinical implications. Proc Natl
Acad Sci USA. 98:10869–10874. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Richardson AL, Wang ZC, De Nicolo A, Lu X,
Brown M, Miron A, Liao X, Iglehart JD, Livingston DM and Ganesan S:
X chromosomal abnormalities in basal-like human breast cancer.
Cancer Cell. 9:121–132. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Diagnosis and Treatment Guidelines For
Colorectal Cancer Working Group C: Chinese Society of Clinical
Oncology (CSCO) diagnosis and treatment guidelines for breast
cancer. 2017.In Chinese.
|
|
21
|
Song S, Peng P, Tang Z, Zhao J, Wu W, Li
H, Shao M, Li L, Yang C, Duan F, et al: Decreased expression of
STING predicts poor prognosis in patients with gastric cancer. Sci
Rep. 7:398582017. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Zhang C, Guo H, Li B, Sui C, Zhang Y, Xia
X, Qin Y, Ye L, Xie F, Wang H, et al: Effects of Slit3 silencing on
the invasive ability of lung carcinoma A549 cells. Oncol Rep.
34:952–960. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
24
|
Li S, Quarto N, Senarath-Yapa K, Grey N,
Bai X and Longaker MT: Enhanced activation of canonical Wnt
signaling confers mesoderm-derived parietal bone with similar
osteogenic and skeletal healing capacity to neural crest-derived
frontal bone. PLoS One. 10:e01380592015. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Mao M, Alavi MV, Labelle-Dumais C, Gould
DB and Type IV: Type IV collagens and basement membrane diseases:
Cell biology and pathogenic mechanisms. Curr Top Membr. 76:61–116.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Tryggvason K, Höyhtyä M and Pyke C: Type
IV collagenases in invasive tumors. Breast Cancer Res Treat.
24:209–218. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Zhou R, Xu L, Ye M, Liao M, Du H and Chen
H: Formononetin inhibits migration and invasion of MDA-MB-231 and
4T1 breast cancer cells by suppressing MMP-2 and MMP-9 through
PI3K/AKT signaling pathways. Horm Metab Res. 46:753–760. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Kim S, Choi JH, Lim HI, Lee SK, Kim WW,
Cho S, Kim JS, Kim JH, Choe JH, Nam SJ, et al: EGF-induced MMP-9
expression is mediated by the JAK3/ERK pathway, but not by the
JAK3/STAT-3 pathway in a SKBR3 breast cancer cell line. Cell
Signal. 21:892–898. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Kim ES, Kim MS and Moon A:
TGF-beta-induced upregulation of MMP-2 and MMP-9 depends on p38
MAPK, but not ERK signaling in MCF10A human breast epithelial
cells. Int J Oncol. 25:1375–1382. 2004.PubMed/NCBI
|
|
30
|
Li W, Liu Z, Zhao C and Zhai L: Binding of
MMP-9-degraded fibronectin to β6 integrin promotes invasion via the
FAK-Src-related Erk1/2 and PI3K/Akt/Smad-1/5/8 pathways in breast
cancer. Oncol Rep. 34:1345–1352. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Rolli M, Fransvea E, Pilch J, Saven A and
Felding-Habermann B: Activated integrin alphavbeta3 cooperates with
metalloproteinase MMP-9 in regulating migration of metastatic
breast cancer cells. Proc Natl Acad Sci USA. 100:9482–9487. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Chung J, Bachelder RE, Lipscomb EA, Shaw
LM and Mercurio AM: Integrin (alpha 6 beta 4) regulation of eIF-4E
activity and VEGF translation: A survival mechanism for carcinoma
cells. J Cell Biol. 158:165–174. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Yebra M, Montgomery AM, Diaferia GR, Kaido
T, Silletti S, Perez B, Just ML, Hildbrand S, Hurford R,
Florkiewicz E, et al: Recognition of the neural chemoattractant
Netrin-1 by integrins alpha6beta4 and alpha3beta1 regulates
epithelial cell adhesion and migration. Dev Cell. 5:695–707. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Gilcrease MZ, Zhou X, Lu X, Woodward WA,
Hall BE and Morrissey PJ: Alpha6beta4 integrin crosslinking induces
EGFR clustering and promotes EGF-mediated Rho activation in breast
cancer. J Exp Clin Cancer Res. 28:672009. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Lee SS, Lee SJ, Lee SH, Ryu JM, Lim HS,
Kim JS, Song EJ, Jung YH, Lee HJ, Kim CH, et al: Netrin-1-induced
stem cell bioactivity contributes to the regeneration of injured
tissues via the lipid raft-dependent integrin α6β4 signaling
pathway. Sci Rep. 6:375262016. View Article : Google Scholar
|
|
36
|
Delcommenne M, Tan C, Gray V, Rue L,
Woodgett J and Dedhar S: Phosphoinositide-3-OH kinase-dependent
regulation of glycogen synthase kinase 3 and protein kinase B/AKT
by the integrin-linked kinase. Proc Natl Acad Sci USA.
95:11211–11216. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Bernet A and Mehlen P: Dependence
receptors: When apoptosis controls tumor progression. Bull Cancer.
94:E12–E17. 2007.PubMed/NCBI
|
|
38
|
Paradisi A, Creveaux M, Gibert B, Devailly
G, Redoulez E, Neves D, Cleyssac E, Treilleux I, Klein C,
Niederfellner G, et al: Combining chemotherapeutic agents and
netrin-1 interference potentiates cancer cell death. EMBO Mol Med.
5:1821–1834. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Grandin M, Mathot P, Devailly G, Bidet Y,
Ghantous A, Favrot C, Gibert B, Gadot N, Puisieux I, Herceg Z, et
al: Inhibition of DNA methylation promotes breast tumor sensitivity
to netrin-1 interference. EMBO Mol Med. 8:863–877. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Lipscomb EA, Simpson KJ, Lyle SR, Ring JE,
Dugan AS and Mercurio AM: The alpha6beta4 integrin maintains the
survival of human breast carcinoma cells in vivo. Cancer Res.
65:10970–10976. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Bhatia V, Mula RV, Weigel NL and Falzon M:
Parathyroid hormone-related protein regulates cell survival
pathways via integrin alpha6beta4-mediated activation of
phosphatidylinositol 3-kinase/Akt signaling. Mol Cancer Res.
7:1119–1131. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
van Kempen LCL and Coussens LM: MMP9
potentiates pulmonary metastasis formation. Cancer Cell. 2:251–252.
2002. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Missan DS, Mitchell K, Subbaram S and
DiPersio CM: Integrin α3β1 signaling through MEK/ERK determines
alternative polyadenylation of the MMP-9 mRNA transcript in
immortalized mouse keratinocytes. PLoS One. 10:e01195392015.
View Article : Google Scholar
|
|
44
|
Sil H, Sen T and Chatterjee A:
Fibronectin-integrin (alpha5beta1) modulates migration and invasion
of murine melanoma cell line B16F10 by involving MMP-9. Oncol Res.
19:335–348. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Lamar JM, Iyer V and DiPersio CM: Integrin
alpha3beta1 potentiates TGFbeta-mediated induction of MMP-9 in
immortalized keratinocytes. J Invest Dermatol. 128:575–586. 2008.
View Article : Google Scholar
|
|
46
|
Son TW, Yun SP, Yong MS, Seo BN, Ryu JM,
Youn HY, Oh YM and Han HJ: Netrin-1 protects hypoxia-induced
mitochondrial apoptosis through HSP27 expression via DCC- and
integrin α6β4-dependent Akt, GSK-3β, and HSF-1 in mesenchymal stem
cells. Cell Death Dis. 4:e5632013. View Article : Google Scholar
|
|
47
|
Lee SJ, Jung YH, Oh SY, Yong MS, Ryu JM
and Han HJ: Netrin-1 induces MMP-12-dependent E-cadherin
degradation via the distinct activation of PKCα and FAK/Fyn in
promoting mesenchymal stem cell motility. Stem Cells Dev.
23:1870–1882. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Taherian A, Li X, Liu Y and Haas TA:
Differences in integrin expression and signaling within human
breast cancer cells. BMC Cancer. 11:2932011. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Chisholm A and Tessier-Lavigne M:
Conservation and divergence of axon guidance mechanisms. Curr Opin
Neurobiol. 9:603–615. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Harburger DS and Calderwood DA: Integrin
signalling at a glance. J Cell Sci. 122:159–163. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Li W, Aurandt J, Jürgensen C, Rao Y and
Guan KL: FAK and Src kinases are required for netrin-induced
tyrosine phosphorylation of UNC5. J Cell Sci. 119:47–55. 2006.
View Article : Google Scholar
|
|
52
|
Li W, Lee J, Vikis HG, Lee SH, Liu G,
Aurandt J, Shen TL, Fearon ER, Guan JL, Han M, et al: Activation of
FAK and Src are receptor-proximal events required for netrin
signaling. Nat Neurosci. 7:1213–1221. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Aikawa R, Nagai T, Kudoh S, Zou Y, Tanaka
M, Tamura M, Akazawa H, Takano H, Nagai R and Komuro I: Integrins
play a critical role in mechanical stress-induced p38 MAPK
activation. Hypertension. 39:233–238. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Huang D, Khoe M, Befekadu M, Chung S,
Takata Y, Ilic D and Bryer-Ash M: Focal adhesion kinase mediates
cell survival via NF-kappaB and ERK signaling pathways. Am J
Physiol Cell Physiol. 292:C1339–C1352. 2007. View Article : Google Scholar
|
|
55
|
Chen HC, Appeddu PA, Isoda H and Guan JL:
Phosphorylation of tyrosine 397 in focal adhesion kinase is
required for binding phosphatidylinositol 3-kinase. J Biol Chem.
271:26329–26334. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Acconcia F, Barnes CJ, Singh RR, Talukder
AH and Kumar R: Phosphorylation-dependent regulation of nuclear
localization and functions of integrin-linked kinase. Proc Natl
Acad Sci USA. 104:6782–6787. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Persad S, Attwell S, Gray V, Mawji N, Deng
JT, Leung D, Yan J, Sanghera J, Walsh MP and Dedhar S: Regulation
of protein kinase B/Akt-serine 473 phosphorylation by
integrin-linked kinase: Critical roles for kinase activity and
amino acids arginine 211 and serine 343. J Biol Chem.
276:27462–27469. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Troussard AA, Costello P, Yoganathan TN,
Kumagai S, Roskelley CD and Dedhar S: The integrin linked kinase
(ILK) induces an invasive phenotype via AP-1 transcription
factor-dependent upregulation of matrix metalloproteinase-9
(MMP-9). Oncogene. 19:5444–5452. 2000. View Article : Google Scholar : PubMed/NCBI
|