Introduction
Lung cancer is the leading cause of cancer-related
mortality, and the major histological type of lung cancer is
classified as non-small cell lung cancer (NSCLC) (occurring in 75%
of cases), of which adenocarcinoma (40%) and squamous cell
carcinoma (35%) are the two most common subtypes (1,2). At
present, although surgery is the most effective treatment method
for NSCLC, approximately 80% of patients with NSCLC require
chemotherapy, as they are diagnosed at an advanced stage and are
unable to undergo surgery. However, in clinical practice, >90%
of patients with NSCLC receiving chemotherapy will eventually
become resistant, which leads to treatment failure; thus, the
5-year survival rate is <20% (3). Therefore, the identification of the
molecular mechanisms responsible for resistance to chemotherapy and
a method with which to overcome drug resistance in NSCLC are
urgently required.
In recent years, cancer stem cells have been
considered to be the source of tumor heterogeneity and tumor
expansion, and are the source of resistance to tumor chemotherapy
in numerous types of cancer, including NSCLC (4,5).
Cancer stem cells refer to a very small part of the tumor tissue
and are also known as side population cells (6). Similar to normal stem cells, cancer
stem cells have stem cell-like properties, such as differentiation
potential, self-renewal capability, dormancy capability, DNA repair
capability and drug efflux capability during tumor development and
play important roles in tumor progression (7,8).
Moreover, cancer stem cells cannot be eliminated completely by drug
treatment and result in resistance to tumor chemotherapy, although
the underlying basis of cancer stem cells in resistance to
chemotherapy remains elusive (9,10).
Therefore, cancer stem cells are considered to drive
chemotherapeutic resistance in NSCLC, and the investigation into
the regulatory mechanisms of cancer stem cell properties is
essential for NSCLC therapy.
In the present study, DNA methylation microarray was
used to detect the DNA methylation profiles of cisplatin-resistant
A549/DDP cells and cisplatin-sensitive A549 cells, and it was found
that the DNA methylation levels of forkhead box F1 (FOXF1)
decreased significantly in the A549/DDP cells compared with the
A549 cells. FOXF1 is a member of the forkhead box (FOX)
transcription factor family, which is involved in various complex
cellular processes and significantly implicated in cancer (11,12).
Herein, it was found that the hypomethylation of FOXF1 increased
the expression of FOXF1 and that the overexpression of FOXF1
enhanced the resistance of NSCLC cells to cisplatin by promoting
cancer stem cell properties, such as self-renewal capability. In
addition, clinical analysis found that high expression levels of
FOXF1 in NSCLC tissues were associated with platinum-based
chemotherapeutic resistance. These results not only shed light onto
the mechanisms of cancer stem cells in chemotherapy-resistant
NSCLC, but may also lead to the discovery of a biomarker that may
be used for the identification of patients with NSCLC who are
resistant to chemotherapy.
Materials and methods
Cell culture and transfections
A549/DDP (cisplatin-resistant A549) cells were
established using increasing concentrations of cisplatin. Briefly,
A549 cells (ATCC) in logarithmic growth were treated with 0.5
µmol/l of cisplatin. After 48 h, cisplatin was withdrawn and the
cells were cultured without cisplatin until they recovered. The
same treatment was performed again, and when the cells were
considered to be resistant to cisplatin (cells can grow normally at
the current cisplatin concentration), the concentration was
gradually increased up to a final concentration of 3 µmol/l. When
the induced cells had survived in 3 µmol/l of cisplatin for
approximately 2 months with normal activity, the cells were
confirmed to be cisplatin-resistant and named A549/DDP (the
IC50 increased from 0.49 to 4.12 µmol/l) (data not
shown). The A549/DDP cells were cultured with 2 µmol/l
cisplatin.
NSCLC epithelial cell lines (A549, A549/DDP and
H1299) and the normal lung epithelial cell line (16HBE) (ATCC) were
cultured in RPMI-1640 medium supplemented with 10% fetal bovine
serum at 37˚C in a humidified atmosphere with 5% CO2.
The expression plasmid of FOXF1 was constructed by inserting cDNA
into the pCDNA3.1 plasmid (Invitrogen; Thermo Fisher Scientific),
and the shRNAs of FOXF1 was designed for lentivirus production
(Shanghai GeneChem Co.). The plasmids were transfected into the
cells using Lipofectamine 2000 (Invitrogen; Thermo Fisher
Scientific), and shRNA lentiviral vectors were transfected into the
cells by the lentivirus. After 72 h, stable transfected cell lines
were established for analysis.
Tissue sample collection
Primary NSCLC tissues were collected from the Cancer
Center of Guangzhou Medical University (Guangzhou, China) with
informed consent and Institutional Review Board (IRB) permission. A
total of 70 patients with NSCLC were recruited into this study. All
of the following criteria were met: Patients with primary NSCLC; a
histological diagnosis of NSCLC with at least one measurable
lesion; a TNM clinical stage of III to IV; first-line chemotherapy
with platinum-based chemotherapy every 3 weeks for a maximum of 4
cycles. According to the Response Evaluation Criteria in Solid
Tumors (RECIST), tissue samples were divided into 2 groups
according to the patient's response assessed by medical image
analysis and the detection of serum tumor markers after 4 cycles of
platinum-based chemotherapy: Response or partial response, at least
a 30% decrease in the sum of diameters of target lesions from
pre-chemotherapy levels, taking as a reference the baseline sum
diameters; stable or progressive disease, a decrease of <30% or
an increase from pre-chemotherapy levels, taking as a reference the
baseline sum diameters. Patients with chemotherapy response or
partial response were considered to be chemotherapy-sensitive (R,
responder), whereas patients with stable or progressive disease
were grouped together and considered to be chemotherapy-resistant
(NR, non-responder). The patient characteristics are presented in
Table SI. Fresh NSCLC tissues were obtained by aspiration biopsy
and immediately snap-frozen in liquid nitrogen and stored at -80˚C
until use. All clinical and biological data on these samples were
available.
All patients provided written informed consent, and
the collection of NSCLC tissues for research purposes was approved
by the relevant human research ethics committees of the Affiliated
Cancer Hospital and Institute of Guangzhou Medical University.
DNA methylation assays
According to the manufacturer's instructions, total
DNA was extracted from cells and tissue samples using the Genomic
DNA Purification kit (Promega) and were then bisulfite-modified
using the EpiTect Bisulfite kit (Qiagen).
Genome-wide methylation analysis was performed using
the validated Illumina Infinium HumanMethylation 450K BeadChip. The
methylation score of each CpG is represented as a β-value.
The detection of FOXF1 methylation was achieved by
the quantitative measurement of methylated (C)/unmethyl-ated (T)
allele peak ratios by pyrosequencing analysis. The assay was
designed to detect FOXF1 CpG site methylation at chromosome 16 at
position 86542659-86542696 (position 1, 6 CpG sites) and
86542770-86542808 (position 2, 6 CpG sites). Sequence accessions
matched the UCSC Genome Browser Human assembly (GRCh37/hg19). The
sequence reaction and detection were performed by pyrosequencing
following the manufacturer's protocol (PyroMark Q96 Plate, Qiagen).
The results are reported as a percentage of the methylated (C)
allele over the background of un-methylated (T) allele (Methylation
level = mC/(mC + umT) ×100%). The primers for FOXF1 pyrosequencing
are listed in Table SII.
The DNA methyltransferase inhibitor
5-aza-2′-deoxycyti-dine (5-Aza-dC) (Sigma) was used to block DNA
methylation. The cells were treated with 5-Aza-dC at 10 µM for 3
days. Drugs and culture medium were refreshed every day during
treatment.
Reverse transcription-quantitative PCR
(RT-qPCR)
Total RNA in cells was extracted using TRIzol
reagent (Invitrogen; Thermo Fisher Scientific). Total RNA (1 µg)
was used for cDNA synthesis using a Reverse Transcription kit
(Takara), and the cDNA was then used for quantitative polymerase
chain reaction (qPCR) using SYBR-Green Real-time PCR Master Mix
(Toyobo). RT-qPCR was performed using the ABI ViiA™7Dx Real-Time
PCR System (Life Technologies; Thermo Fisher Scientific)
(thermocycling conditions: 50˚C, 2 min; 95˚C, 10 min; (95˚C, 15
sec; 60˚C, 1 min) 40 cycles). The expression levels of mRNA were
normalized to GAPDH (method of quantification, 2-ΔΔCq)
(13). The primers used for
RT-qPCR are listed in Table SII.
Western blot analysis
The cells were harvested and lysed using RIPA buffer
for 30 min at 4˚C. A total of 50 µg heat-denatured proteins were
loaded onto a 15% SDS-polyacrylamide gel electrophoresis (SDS-PAGE)
system, and then transferred to a polyvinylidene difluoride
membrane for western blot analysis. After blocking non-specific
binding sites with 5% (wt/vol) non-fat milk, 0.1% (vol/vol)
Tween-20 diluted in Tris (pH 7.8)-buffered saline, rabbit
polyclonal anti-FOXF1 (ab23194, 1:400 dilution), anti- aldehyde
dehydrogenase 1 (ALDH1; ab52492, 1:500 dilution),
anti-octamer-binding transcription factor 4 (OCT4; ab18976, 1:500
dilution) and anti-GAPDH (ab37168, 1:1,000 dilution) (all from
Abcam) primary antibodies were added followed by incubation for 2 h
at 37˚C. Subsequently, horseradish peroxidase (HRP)-conjugated goat
anti-rabbit secondary antibody (1:2,000 dilution, A0208, Beyotime)
was added and incubated for 2 h at 25˚C. The bound antibodies were
detected using the ECL Plus Western Blotting Detection system (GE
Healthcare). Actin was used as an internal control.
Cell immunofluorescence staining
NSCLC cells were stained using the standard
immunofluorescence (IF) protocol. Briefly, NSCLC cells were
pre-fixed with 4% paraformaldehyde and permeabilized with PBS
containing 0.2% Triton X-100 and 5% BSA, and the cells were stained
with direct anti-FOXF1 (ab23194, 1:200 dilution), anti-ALDH1
(ab52492, 1:200 dilution) and anti-OCT4 (ab18976, 1:200 dilution)
antibodies (all from Abcam) overnight at 4˚C. The cells were then
washed with PBS 3 times, and an appropriate secondary antibody
(1:400 dilution, goat anti-rabbit-Texas Red, ab6719, Abcam) was
added followed by incubation for 1 h at 25˚C. Finally, the cells
were stained with DAPI and images were visualized using a
fluorescence microscope (Leica).
Xenograft tumors in nude mice
For the xenograft model assay, 1x107 of
A549/FOXF1-plasmid and A549/control-plasmid cells were
subcutaneously injected into 4-week-old BALB/c athymic nude mice,
respectively [8 mice (weight, 19.0, 18.0, 18.2, 18.5, 17.0, 16.3,
19.3 and 20.3 g), temperature, 18-22˚C; humidity, 50-60%, free
access to food and water]. Each cell line was injected
subcutaneously into the armpit of the right forelimb, for a total
of 8 injections (4 A549/FOXF1-plasmid cells and 4
A549/control-plasmid cells). The longest diameter 'L' and the
shortest diameter 'W' of tumors were measured 5 times in 30 days.
The tumor volume was calculated using the following formula: Tumor
volume (mm3) = π/6 xLxWxW. After 30 days, all
experimental mice were sacrificed simultaneously and tumor weights
were measured.
Animal experiments were approved by the Animal
Ethics Committee of Guangzhou Medical University (2018-114). The
experimental mice were anesthetized with an intraperitoneal
injection of 0.7% pentobarbital sodium (70 mg/kg). The experimental
mice were euthanized with carbon dioxide (flow rate, 20% of the
chamber volume per minute) and subsequently also subjected to
cervical dislocation.
Cell cytotoxicity assays
NSCLC cells were incubated with various
concentrations of cisplatin (for the A549 and H1299 cells: 0, 0.5,
1, 1.5, 2, 2.5, 3, 3.5 and 4 µmol/l; for the A549/DDP cells: 0, 1,
2, 3, 4, 5, 6, 7 and 8 µmol/l). After 48 h, cisplatin-induced
cytotoxicity was determined using the CCK8 kit (Beyotime) and
represented as IC50 (µmol/l).
Colony-formation assay
NSCLC cells were treated with cisplatin (2 µmol/l)
for 24 h, and the cells were then plated in 6-well plates (1,000
cells per well) and allowed to form colonies over 7 days. Cells
were stained with Giemsa (Sigma-Aldrich, cells were pre-fixed with
4% paraformaldehyde and incubated for 15 min at 25˚C) and counted
using ImageJ software (NIH, 1.52r).
Cell apoptosis assay
NSCLC cells were treated with cisplatin (2 µmol/l)
for 24 h. Cell apoptosis was then determined using the Annexin
V-FITC Apoptosis Detection kit (Beyotime) and by flow cytometry
(Guava easyCyte HT, Millipore).
Sphere formation assay
NSCLC cells were plated in DMEM F12 serum-free
medium reconstituted with 20 ng/ml of epidermal growth factor
(EGF), 20 ng/ml of basic fibroblast growth factor (bFGF), 2% B27
and 1% methylcellulose (all from Sigma-Aldrich) (5,000 cells per
well in a 6-well plate). After 4-7 days, microsphere-like
structures were visible, and images of the microspheres were
captured using a microscope (Leica).
Statistical analysis
All values are expressed as the means ± standard
deviation (SD) from at least 3 separate experiments. The Student's
unpaired t-test, ANOVA and receiver operating characteristic (ROC)
curves were performed using SPSS 21.0 statistical software (IBM).
Multiple comparisons between the groups was performed using the
Bonferroni method. A two-tailed P-value was used in all analyses,
and differences were considered statistically significant if the
P-value was <0.05 (P<0.05).
Results
FOXF1 is epigenetically activated in
cisplatin-resistant A549/DDP cells
Firstly, a DNA methylation microarray was used
(Illumina Infinium 450K BeadChip) to detect the DNA methylation
profiles (β-values) of the cisplatin-resistant A549/DDP cells and
cisplatin-sensitive A549 cells. The results revealed that the 32
probes of FOXF1 were defined as 1,500 bp upstream region 1 of the
transcription start site (TSS1500 region 1, 10 probes), 1,500 bp
upstream region 2 of the transcription start site (TSS1500 region
2, 6 probes), First Exon (1stExon, 5 probes), gene body (3 probes)
and 3′ untranslated region (3′UTR, 8 probes), and the differential
region (fold change <0.5, P<0.01) were mapped to the TSS1500
region 1 of the FOXF1 gene and the locations of these 10 probes
concentrated on a CpG island (Fig. 1A
and B, and Table SIII).
Moreover, the methylation status of the FOXF1 CpG island was
determined in NSCLC epithelial cell lines (A549, A549/DDP and
H1299) and a normal lung epithelial cell line (16HBE) using
pyrosequencing analysis. It was confirmed that the DNA methylation
levels of the FOXF1 CpG island were decreased in the A549/DDP cells
compared with the A549, H1299 and 16HBE cells (Figs. 1A and C and S1, and Table SIV). Using western blot analysis
and RT-qPCR, the mRNA and protein expression of levels FOXF1 were
found to be higher in the A549/DDP compared with the A549, H1299
and 16HBE cells (Fig. 1D).
According to the observation that the hypomethylation of the CpG
island of FOXF1 facilitated the expression of FOXF1, it was also
found that demethylation using the DNA methyltransferase inhibitor,
5-Aza-dC, promoted FOXF1 expression in the A549, 16HBE and H1299
cells (Fig. 1E). Therefore, these
results indicate that cisplatin-induced DNA hypomethylation of
FOXF1 can epigenetically activate the expression of FOXF1.
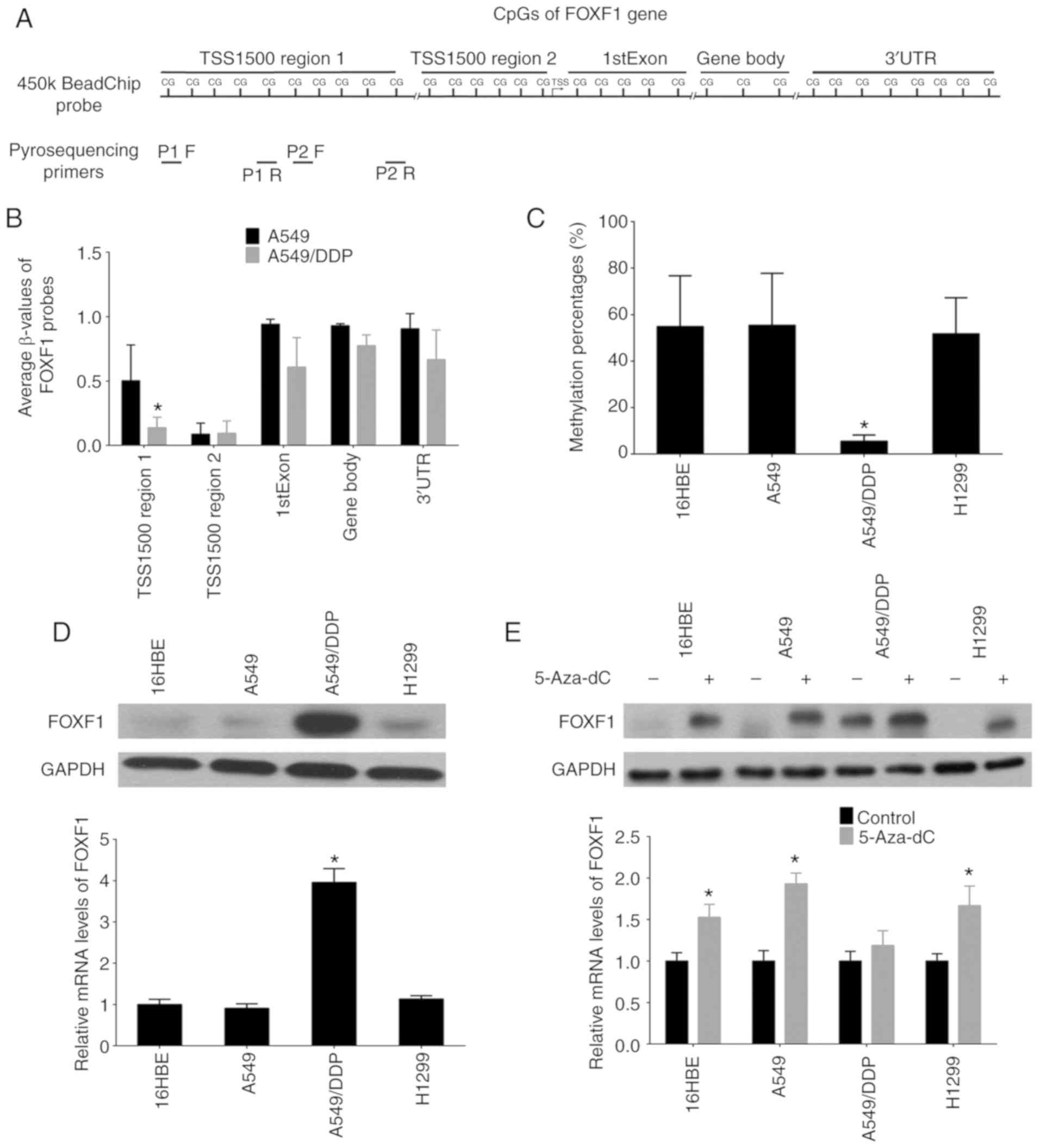 | Figure 1Discovery of differential methylation
region around FOXF1 gene upstream regulatory region in
cisplatin-resistant A549/DDP cells compared with
cisplatin-sensitive A549 cells. (A) The location of Illumina
Infinium HumanMethylation 450k BeadChip FOXF1 probes and
pyrosequencing primers. (B) Average β-values of probes mapping to
different regions of the FOXF1 gene in A549/DDP cells and A549
cells. (C) Statistical analysis of the percentage of methylated
CpGs in the FOXF1 gene. (D) FOXF1 protein levels detected by
western blot analysis (upper panel) and FOXF1 mRNA levels detected
by RT-qPCR (lower panel) in 1 normal lung cell lines (16HBE cells)
and 3 NSCLC cell lines (A549, A549/DDP and H1299 cells). (E) FOXF1
protein levels detected by western blot analysis (upper panel) and
FOXF1 mRNA levels detected by RT-qPCR (lower panel) in 16HBE, A549,
A549/DDP and H1299 cells following treatment with the demethylating
agent, 5-Aza-dC. n=3, *P<0.05 [(B) A549 compared to
A549/DDP; (C) A549/DDP compared to 16HBE, A549 and H1299; (D)
A549/DDP compared to 16HBE, A549 and H1299; (E) 5-Aza-dC compared
to Control]. 5-Aza-dC, 5-aza-2′-deoxycytidine; FOXF1, forkhead box
F1; NSCLC, non-small cell lung cancer. |
FOXF1 regulates cisplatin resistance and
is associated with platinum-based chemotherapeutic resistance in
NSCLC
In order to investigate the regulatory functions of
FOXF1 in cisplatin resistance, A549 and H1299 cells were
established with the stable overexpression of FOXF1 (FOXF1
plasmids) and A549/DDP cells were established with the stable
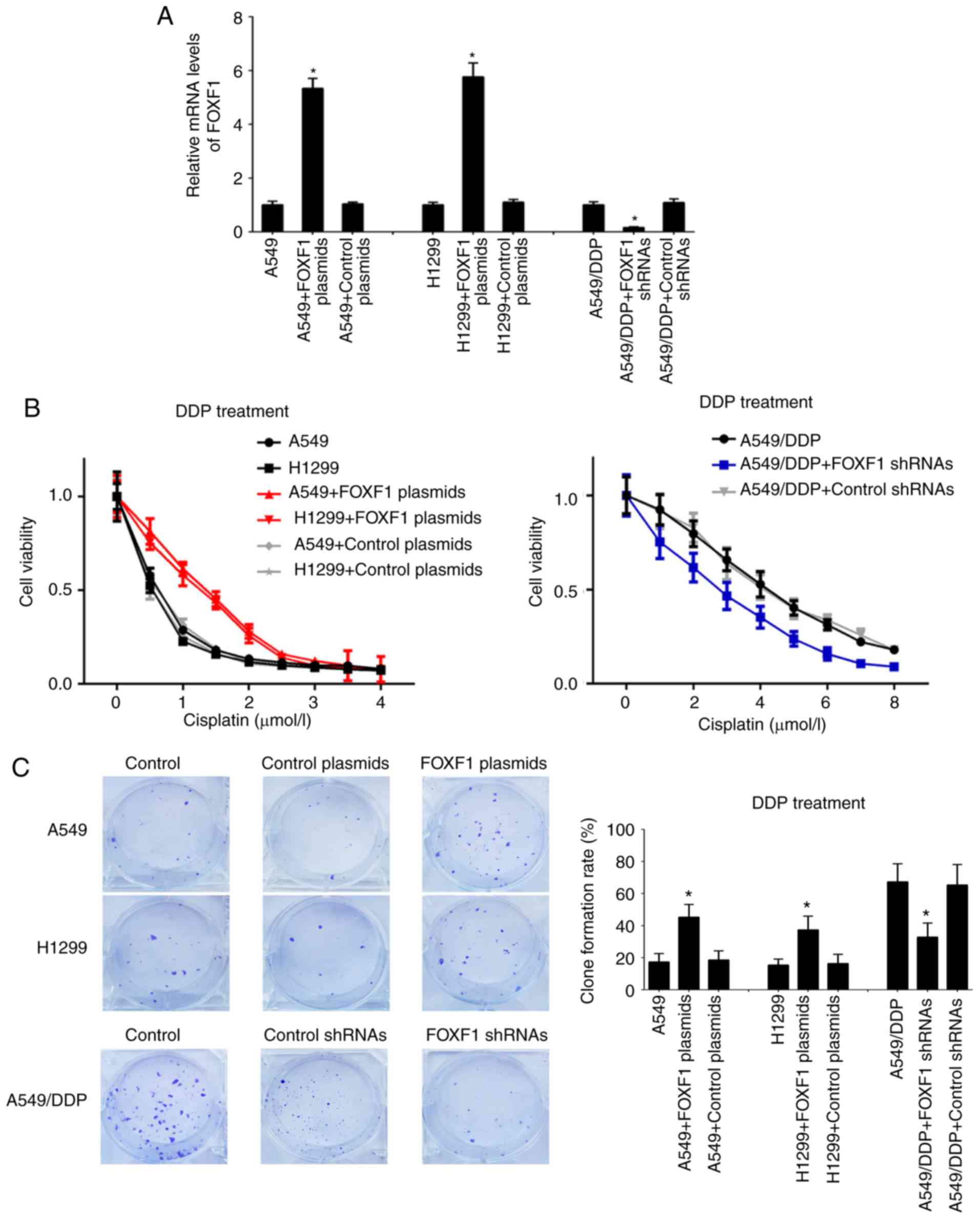
knockdown of FOXF1 (FOXF1 shRNAs) (Fig. 2A). In A549 and H1299 cells, the
overexpression of FOXF1 protected the A549 and H1299 cells from
cisplatin (IC50 increased from 0.51 to 1.33 µmol/l for
the A549 cells and from 0.43 to 1.15 µmol/l for the H1299 cells);
in the A549/DDP cells, the knockdown of FOXF1 restored sensitivity
to cisplatin (IC50 decreased from 4.23 to 2.77 µmol/l)
(Fig. 2B). The results of the
colony-formation assay demonstrated that the overexpression of
FOXF1 increased the proliferation of the A549 and H1299 cells
treated with cisplatin, and the knockdown of FOXF1 decreased the
proliferation of the A549/DDP cells treated with cisplatin
(Fig. 2C). Moreover, the
overexpression of FOXF1 decreased the cisplatin-induced apoptosis
of A549 and H1299 cells, and the knockdown of FOXF1 increased the
cisplatin-induced apoptosis of A549/DDP cells (Fig. 2D). In addition, the overexpression
of FOXF1 decreased the cisplatin-induced apoptosis of 16HBE cells
(Fig. S2). Thus, these findings
suggest that FOXF1 plays important roles in the regulation of
cisplatin resistance in NSCLC by promoting cell proliferation and
inhibiting cell apoptosis.
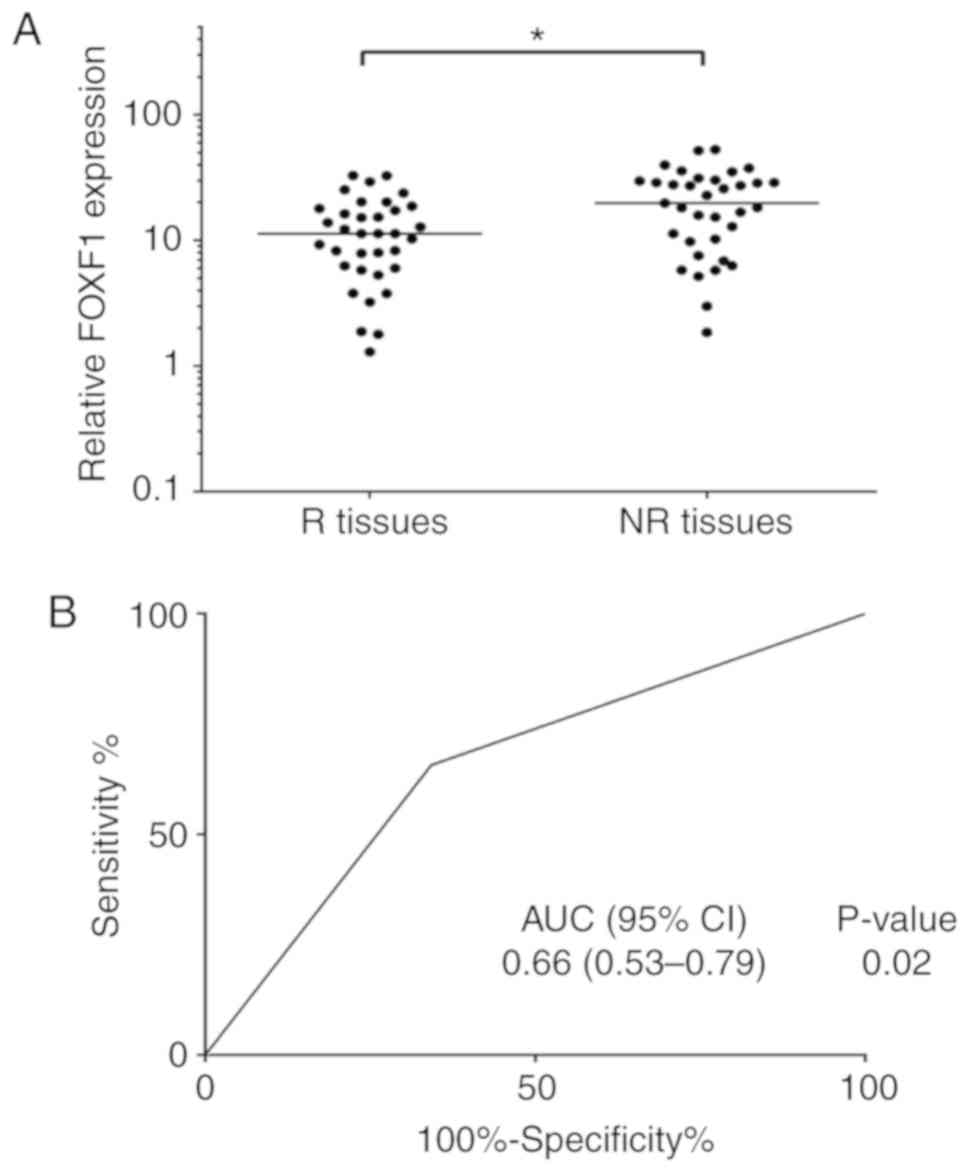
FOXF1 has been found to be associated with cisplatin
resistance in NSCLC cells; therefore, the authors wished to
determine whether the expression levels of FOXF1 may be associated
with the outcome of chemotherapy in patients with NSCLC. A total of
35 platinum-based chemotherapy-resistant NSCLC tissues (NR tissues)
and 35 platinum-based chemotherapy-sensitive NSCLC tissues (R
tissues) were collected, and the expression levels of FOXF1 were
measured in these samples using RT-qPCR. The results revealed that
the expression levels of FOXF1 were significantly higher in the NR
tissues compared with the R tissues (Fig. 3A). To determine the potential of
FOXF1 in predicting the clinical outcome of platinum-based
chemotherapy, ROC analysis of the expression levels of FOXF1 was
performed and the area under the curve (AUC) of the ROC was
calculated to assess the sensitivity and specificity of the
prediction. The AUC value of the FOXF1 expression level was 0.66
(P<0.05) (Fig. 3B). On the
whole, these findings suggested that FOXF1 was significantly
associated with platinum-based chemotherapeutic resistance in
patients with NSCLC and may thus be used as a potential biomarker
for predicting the clinical outcome of platinum-based chemotherapy
in NSCLC.
FOXF1 induces the development of cancer
stem cell properties in NSCLC cells
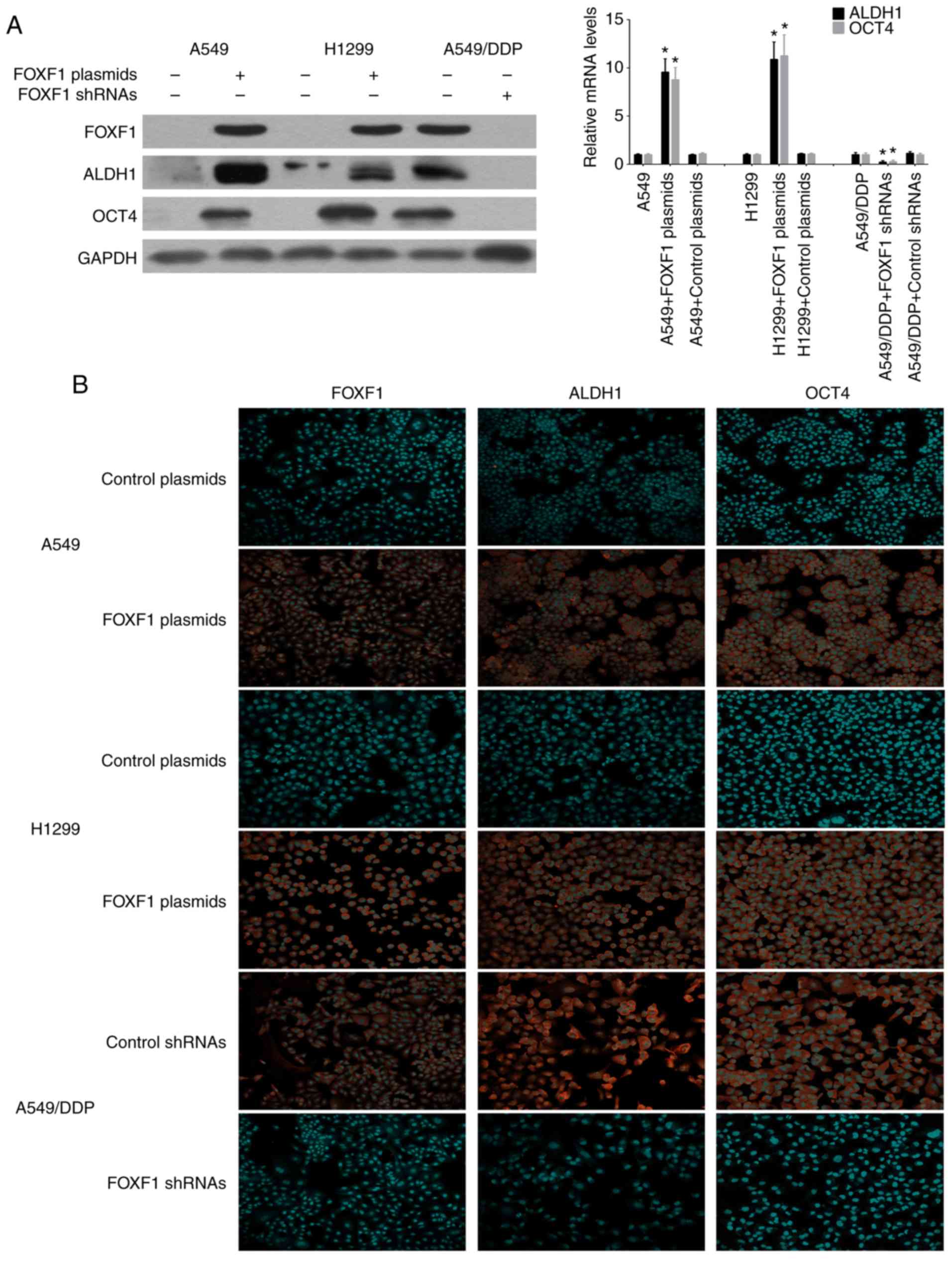
The stem cell markers, ALDH1 and OCT4, have been
successfully used to isolate cancer stem cells from NSCLC cell
lines and tissues, and it is widely accepted that NSCLC cancer stem
cells are contained exclusively in the high ALDH1 and OCT4
expression cell compartment (14,15).
Therefore, this study investigated the influence of the
overex-pression and knockdown of FOXF1 on the expression of the
stem cell markers, ALDH1 and OCT4. Using western blot analysis and
RT-qPCR, ALDH1 and OCT4 were found to be highly expressed in the
A549/DDP cells compared with the A549 and H1299 cells. The
overexpression of FOXF1 resulted in an increase in ALDH1 and OCT4
expression in the A549 and H1299 cells, and the knockdown of FOXF1
resulted in a decrease in ALDH1 and OCT4 expression in the A549/DDP
cells (Fig. 4A). Moreover, the
regulatory effects of FOXF1 on ALDH1 and OCT4 expression were
confirmed by immunofluorescence staining (Fig. 4B), and it was also found that the
overexpression of FOXF1 increased the expression of ALDH1 and OCT4
in the 16HBE cells (Fig. S3).
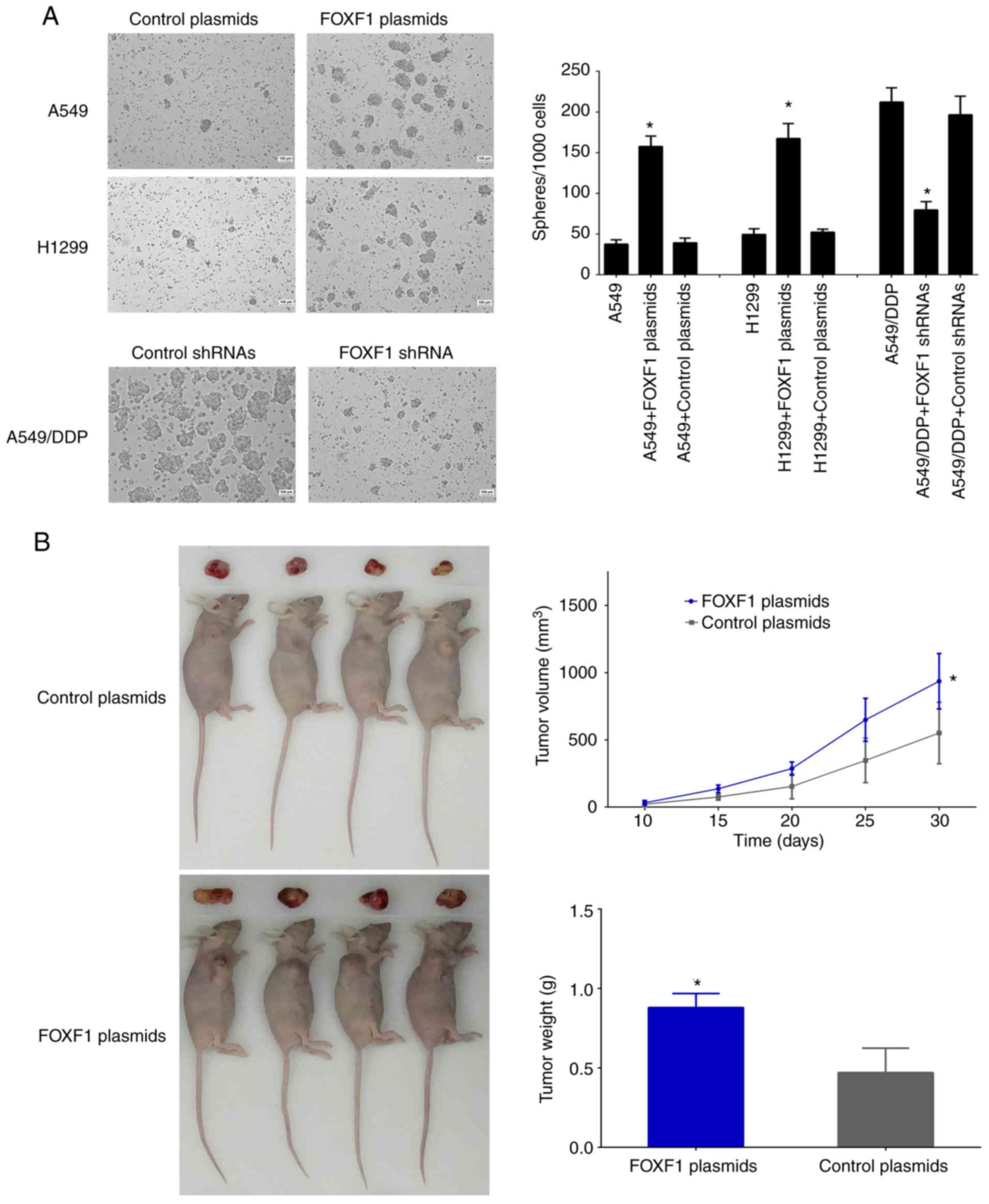
The effect of FOXF1 on the self-renewal capacity of
the cells was then examined using sphere formation assay. The
results revealed that the number and size of spheres formed by the
A549/DDP cells were greater than those formed by the A549 and H1299
cells. The overexpression of FOXF1 resulted in an increase in the
number and size of spheres formed by the A549 and H1299 cells, and
the knockdown of FOXF1 resulted in a decrease in the number and
size of spheres formed by the A549/DDP cells (Fig. 5A). These data suggested that FOXF1
promoted the self-renewal ability of the NSCLC cells. Moreover,
according to the results of the xenograft model assay, the
overexpression of FOXF1 resulted in a significant increase in tumor
volumes and weights compared with control-plasmid, indicating that
FOXF1 promoted tumorigenesis (Fig.
5B). Taken together, these data demonstrate that the
cisplatin-induced upregulation of FOXF1 expression is generally a
critical determinant of stem cell-like properties, which results in
cisplatin resistance in NSCLC cells.
Discussion
Resistance to chemotherapy is the main cause of
chemotherapy failure and disease recurrence, and is a major
obstacle in the treatment of a number of types of cancer, including
NSCLC (16,17). The mechanisms responsible for
resistance to chemotherapy are complex, and are related to several
factors, such as drug uptake, cell cycle, DNA repair and others;
the molecular mechanisms responsible for chemotherapeutic
resistance have not yet been fully elucidated (18,19).
Recently, it was found that the abnormal DNA methylation of the
genome plays an important role in the chemotherapy resistance of
cancers, including NSCLC (20,21).
It was found that platinum-based chemotherapy can influence DNA
methylation modification in cancer (22). In the present study, it was found
that cisplatin influenced the genomic DNA methylation levels in
A549/DDP cells, and it was confirmed that the DNA methylation
levels of the FOXF1 gene upstream regulatory region were
significantly decreased. FOXF1 is located in 16q24.1, and the
upstream regulatory region of FOXF1 has a CpG island which can
inhibit the transcription of FOXF1 by DNA methylation modification
(23,24). It was also found that the
hypomethylation of FOXF1 can increase the mRNA and protein
expression levels of FOXF1 in NSCLC cells. FOXF1 has been found to
be abnormally expressed in cancers, which is closely related to
tumor occurrence and progression (25-27).
Gialmanidis et al found that FOXF1 expression was elevated
in NSCLC tissue samples and was associated with lymph node
metastasis (28). Saito et
al found that FOXF1 was involved in the regulation of the
tumor-promoting properties of lung cancer-associated fibroblasts
(29). In this study, it was found
that the overexpression of FOXF1 promoted cisplatin resistance in
NSCLC by increasing cell proliferation and inhibiting cell
apoptosis. In addition, the expression levels of FOXF1 in were
determined in NSCLC samples, and it was found that the expression
levels of FOXF1 were higher in platinum-based
chemotherapy-resistant NSCLC tissues compared with platinum-based
chemotherapy-sensitive NSCLC tissues; the expression levels of
FOXF1 were associated with the platinum-based chemotherapeutic
response in patients with NSCLC. Therefore, these results indicate
that cisplatin can promote the expression of FOXF1, which results
in cisplatin resistance, by the hypomethylation of the FOXF1 gene
upstream regulatory region.
There is strong evidence to indicate that
abnormalities in DNA methylation can influence the
dedifferentiation of cancer cells, which promotes stem cell
properties in cancer cells, which then form cancer stem cells
(30,31). In the course of cancer treatment,
cancer stem cells have the characteristics of dormancy, strong DNA
repair and self-renewal. As a result, they are 'difficult to kill'
and become the source of chemotherapy resistance and cancer
recurrence (32-35). Cancer stem cells have also been
found in NSCLC and are closely related to chemotherapeutic
resistance in NSCLC (36).
Lopez-Ayllon et al found that cancer stem cells isolated
from NSCLC cells were resistant to cisplatin (37). Bora-Singhal et al found that
Gli1 promoted the self-renewal of NSCLC cancer stem cells by
regulating Sox2, and promoted drug resistance (38). FOXF1 is one of the important
transcription factors involved in the regulation of lung tissue
differentiation and development, and is mainly involved in the
regulation of the correct development of airway smooth muscle and
cartilage (39). Wei et al
found that abnormalities in FOXF1 can influence the
dedifferentiation of cancer cells which promotes stem cell
properties in cancer cells, which then form cancer stem cells
(40). In this study, it was found
that the overexpression of FOXF1 increased the expression of the
stem cell markers, ALDH1 and OCT4, and promoted the self-renewal
capacity and tumorigenesis capacity of NSCLC cells, suggesting that
the abnormal high expression of FOXF1 induced by cisplatin can
promote the stem cell-like properties of NSCLC cells. Thus, the
ability of FOXF1 to initiate cisplatin resistance is dependent in
turn on their ability to promote the cancer stem cell properties of
NSCLC cells. Moreover, it was also found that the overexpression of
FOXF1 increased the expression of stem cell markers and promoted
cisplatin resistance in 16HBE normal human bronchial epithelial
cells. As FOXF1 is involved in the regulation of normal lung tissue
differentiation, the abnormal expression of FOXF1 can also promote
the stem cell-like properties of normal lung epithelial cells and
may be related to the occurrence of NSCLC.
According to the results of this study, we suggest
that cisplatin can promote the transcription of FOXF1 by
hypo-methylation of the upstream regulatory region of FOXF1 gene,
and FOXF1 further promotes cancer stem cell properties which
ultimately result in cisplatin resistance in NSCLC. Furthermore,
the understanding of the molecular mechanisms of cisplatin
resistance regulated by FOXF1 may also provide biomarkers and
therapeutic targets for NSCLC chemotherapy.
Supplementary Data
Funding
This study was supported by grants from the National
Natural Science Foundation of China (81501969, 81572258), Guangdong
Basic and Applied Basic Research Foundation (2019A1515011092,
2019A1515010026) and Guangzhou key medical discipline construction
project fund.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
JZ and JY conceived and designed the experiments.
JZ, JY, and SZ performed the experiments and assembled the data.
XX, WF, LD and ZJ obtained the tumors and tissues with clinical
information where available. JY and BD performed the statistical
analysis. JZ and JY analyzed the data and drafted the manuscript.
All authors have read and approved the final manuscript.
Ethics approval and consent to
participate
NSCLC tissues were collected from the Cancer Center
of Guangzhou Medical University (Guangzhou, China) with written
informed consent and permission from the Institutional Review
Board. All patients provided written informed consent. The study
protocol was approved by the ethics committee of Cancer Center of
Guangzhou Medical University (approval no (2014) 100). Animal
experiments were approved by the animal ethics committees of the
Guangzhou Medical University (2018-114).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Acknowledgements
Not applicable
References
|
1
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2018. CA Cancer J Clin. 68:7–30. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Chen W, Zheng R, Baade PD, Zhang S, Zeng
H, Bray F, Jemal A, Yu XQ and He J: Cancer statistics in China. CA
Cancer J Clin. 66:115–132. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Lemjabbar-Alaoui H, Hassan OU, Yang YW and
Buchanan P: Lung cancer: Biology and treatment options. Biochim
Biophys Acta. 1856:189–210. 2015.PubMed/NCBI
|
|
4
|
Batlle E and Clevers H: Cancer stem cells
revisited. Nat Med. 23:1124–1134. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Adorno-Cruz V, Kibria G, Liu X, Doherty M,
Junk DJ, Guan D, Hubert C, Venere M, Mulkearns-Hubert E, Sinyuk M,
et al: Cancer stem cells: Targeting the roots of cancer, seeds of
metastasis, and sources of therapy resistance. Cancer Res.
75:924–929. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Beck B and Blanpain C: Unravelling cancer
stem cell potential. Nat Rev Cancer. 13:727–38. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Pisco AO, Brock A, Zhou J, Moor A,
Mojtahedi M, Jackson D and Huang S: Non-Darwinian dynamics in
therapy-induced cancer drug resistance. Nat Commun. 4:24672013.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Hu Y, Guo R, Wei J, Zhou Y, Ji W, Liu J,
Zhi X and Zhang J: Effects of PI3K inhibitor NVP-BKM120 on
overcoming drug resistance and eliminating cancer stem cells in
human breast cancer cells. Cell Death Dis. 6:e20202015. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Pattabiraman DR and Weinberg RA: Tackling
the cancer stem cells-what challenges do they pose? Nat Rev Drug
Discov. 13:497–512. 2014. View
Article : Google Scholar : PubMed/NCBI
|
|
10
|
MacDonagh L, Gray SG, Breen E, Cuffe S,
Finn SP, O'Byrne KJ and Barr MP: Lung cancer stem cells: The root
of resistance. Cancer Lett. 372:147–156. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Bach DH, Long NP, Luu TT, Anh NH, Kwon SW
and Lee SK: The dominant role of forkhead box proteins in cancer.
Int J Mol Sci. 19:E32792018. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Laissue P: The forkhead-box family of
transcription factors: Key molecular players in colorectal cancer
pathogenesis. Mol Cancer. 18:52019. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
14
|
Sterlacci W, Savic S, Fiegl M, Obermann E
and Tzankov A: Putative stem cell markers in non-small-cell lung
cancer: A clini-copathologic characterization. J Thorac Oncol.
9:41–49. 2014. View Article : Google Scholar
|
|
15
|
Curtarelli RB, Gonçalves JM, Dos Santos
LGP, Savi MG, Nör JE, Mezzomo LAM and Rodríguez Cordeiro MM:
Expression of cancer stem cell biomarkers in human head and neck
carcinomas: A systematic review. Stem Cell Rev. 14:769–784. 2018.
View Article : Google Scholar
|
|
16
|
Kartal-Yandim M, Adan-Gokbulut A and Baran
Y: Molecular mechanisms of drug resistance and its reversal in
cancer. Crit Rev Biotechnol. 36:716–726. 2016.
|
|
17
|
Kunjachan S, Rychlik B, Storm G, Kiessling
F and Lammers T: Multidrug resistance: Physiological principles and
nanomedical solutions. Adv Drug Deliv Rev. 65:1852–1865. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Szakács G, Paterson JK, Ludwig JA,
Booth-Genthe C and Gottesman MM: Targeting multidrug resistance in
cancer. Nat Rev Drug Discov. 5:219–234. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Fletcher JI, Williams RT, Henderson MJ,
Norris MD and Haber M: ABC transporters as mediators of drug
resistance and contributors to cancer cell biology. Drug Resist
Updat. 26:1–9. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Grasse S, Lienhard M, Frese S, Kerick M,
Steinbach A, Grimm C, Hussong M, Rolff J, Becker M, Dreher F, et
al: Epigenomic profiling of non-small cell lung cancer xenografts
uncover LRP12 DNA methylation as predictive biomarker for
carboplatin resistance. Genome Med. 10:552018. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Stone A, Zotenko E, Locke WJ, Korbie D,
Millar EK, Pidsley R, Stirzaker C, Graham P, Trau M, Musgrove EA,
et al: DNA methylation of oestrogen-regulated enhancers defines
endocrine sensitivity in breast cancer. Nat Commun. 6:77582015.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Flanagan JM, Wilson A, Koo C, Masrour N,
Gallon J, Loomis E, Flower K, Wilhelm-Benartzi C, Hergovich A,
Cunnea P, et al: Platinum-based chemotherapy induces methylation
changes in blood DNA associated with overall survival in patients
with ovarian cancer. Clin Cancer Res. 23:2213–2222. 2017.
View Article : Google Scholar
|
|
23
|
Dharmadhikari AV, Szafranski P,
Kalinichenko VV and Stankiewicz P: Genomic and epigenetic
complexity of the FOXF1 locus in 16q24.1: Implications for
development and disease. Curr Genomics. 16:107–116. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Lo PK, Lee JS, Liang X, Han L, Mori T,
Fackler MJ, Sadik H, Argani P, Pandita TK and Sukumar S: Epigenetic
inactivation of the potential tumor suppressor gene FOXF1 in breast
cancer. Cancer Res. 70:6047–6058. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Fulford L, Milewski D, Ustiyan V,
Ravishankar N, Cai Y, Le T, Masineni S, Kasper S, Aronow B,
Kalinichenko VV and Kalin TV: The transcription factor FOXF1
promotes prostate cancer by stimulating the mitogen-activated
protein kinase ERK5. Sci Signal. 9:ra482016. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Milewski D, Pradhan A, Wang X, Cai Y, Le
T, Turpin B, Kalinichenko VV and Kalin TV: FoxF1 and FoxF2
transcription factors synergistically promote rhabdomyosarcoma
carcinogenesis by repressing transcription of p21Cip1 CDK
inhibitor. Oncogene. 36:850–862. 2017. View Article : Google Scholar :
|
|
27
|
Ran L, Chen Y, Sher J, Wong EWP, Murphy D,
Zhang JQ, Li D, Deniz K, Sirota I, Cao Z, et al: FOXF1 defines the
core-regulatory circuitry in gastrointestinal stromal tumor. Cancer
Discov. 8:234–251. 2018. View Article : Google Scholar
|
|
28
|
Gialmanidis IP, Bravou V, Petrou I, Kourea
H, Mathioudakis A, Lilis I and Papadaki H: Expression of Bmi1,
FoxF1, Nanog, and γ-catenin in relation to hedgehog signaling
pathway in human non-small-cell lung cancer. Lung. 191:511–521.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Saito RA, Micke P, Paulsson J, Augsten M,
Peña C, Jönsson P, Botling J, Edlund K, Johansson L, Carlsson P, et
al: Forkhead box F1 regulates tumor-promoting properties of
cancer-associated fibroblasts in lung cancer. Cancer Res.
70:2644–2654. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Toh TB, Lim JJ and Chow EK: Epigenetics in
cancer stem cells. Mol Cancer. 16:292017. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Easwaran H, Tsai HC and Baylin SB: Cancer
epigenetics: Tumor heterogeneity, plasticity of stem-like states,
and drug resistance. Mol Cell. 54:716–727. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Wickström M, Dyberg C, Milosevic J, Einvik
C, Calero R, Sveinbjörnsson B, Sandén E, Darabi A, Siesjö P, Kool
M, et al: Wnt/β-catenin pathway regulates MGMT gene expression in
cancer and inhibition of Wnt signalling prevents chemore-sistance.
Nat Commun. 6:89042015. View Article : Google Scholar
|
|
33
|
Carnero A, Garcia-Mayea Y, Mir C, Lorente
J, Rubio IT and LLeonart ME: The cancer stem-cell signaling network
and resistance to therapy. Cancer Treat Rev. 49:25–36. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Shibue T and Weinberg RA: EMT, CSCs, and
drug resistance: The mechanistic link and clinical implications.
Nat Rev Clin Oncol. 14:611–629. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Holohan C, Van Schaeybroeck S, Longley DB
and Johnston PG: Cancer drug resistance: An evolving paradigm. Nat
Rev Cancer. 13:714–726. 2013. View
Article : Google Scholar : PubMed/NCBI
|
|
36
|
Suresh R, Ali S, Ahmad A, Philip PA and
Sarkar FH: The role of cancer stem cells in recurrent and
drug-resistant lung cancer. Adv Exp Med Biol. 890:57–74. 2016.
View Article : Google Scholar
|
|
37
|
Lopez-Ayllon BD, Moncho-Amor V, Abarrategi
A, Ibañez de Cáceres I, Castro-Carpeño J, Belda-Iniesta C, Perona R
and Sastre L: Cancer stem cells and cisplatin-resistant cells
isolated from non-small-lung cancer cell lines constitute related
cell populations. Cancer Med. 3:1099–1111. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Bora-Singhal N, Perumal D, Nguyen J and
Chellappan S: Gli1-mediated regulation of Sox2 facilitates
self-renewal of stem-like cells and confers resistance to EGFR
inhibitors in non-small cell lung cancer. Neoplasia. 17:538–551.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Mahlapuu M, Enerbäck S and Carlsson P:
Haploinsufficiency of the forkhead gene Foxf1, a target for sonic
hedgehog signaling, causes lung and foregut malformations.
Development. 128:2397–2406. 2001.PubMed/NCBI
|
|
40
|
Wei HJ, Nickoloff JA, Chen WH, Liu HY, Lo
WC, Chang YT, Yang PC, Wu CW, Williams DF, Gelovani JG and Deng WP:
FOXF1 mediates mesenchymal stem cell fusion-induced reprogramming
of lung cancer cells. Oncotarget. 5:9514–9529. 2014. View Article : Google Scholar : PubMed/NCBI
|