Similarly to other malignant diseases, the incidence
of cutaneous malignant melanoma (CMM) is increasing worldwide
(1). This increased incidence
seems to be influenced by many factors, including ageing of the
population, behavioural habits, and climatic and environmental
changes. Formation of CMM is associated with the main genetic
drivers such as BRAF, NF1 and NRAS mutations, also usually
associated with chronic skin sun damage (2). Aberrant activation of the
RAS/BRAF/MEK/ERK signalling pathway causes uncontrolled
proliferation of malignant cells in the majority of CMM (3). Melanoma causes most of the skin
cancer-related deaths. The patient overall survival at five years
depends on the thickness of the primary melanoma. CMM is also known
for its remarkable ability to metastasise. Despite the new
therapeutic options, the curability of advanced-stage melanoma is
still limited. These recent therapeutic approaches modulate the
immune response of the organism (e.g., via application of
anti-CTLA-4 and anti-PD-1 antibodies) or target proliferation in
specifically mutated melanomas (e.g., by application of BRAF or MEK
inhibitors). In order to establish novel targeted melanoma
therapies, it is of fundamental importance to understand the
mechanisms activated in the permissive tumour microenvironment. In
particular, interactions between melanoma cells and the tissue
microenvironment play key roles in the disease progression. This
article summarises data on the multifaceted roles of CMM
microenvironment in tumour spreading. This concept may be extended
to the intravasation of bioactive molecules participating in the
melanoma cell crosstalk with non-malignant cells forming the CMM
microenvironment. These molecules also participate in premetastatic
niche formation. Finally, their role in patient wasting is also
widely discussed. The concept of CMM microenvironment as a complex
system suitable for therapeutic targeting is introduced in this
article.
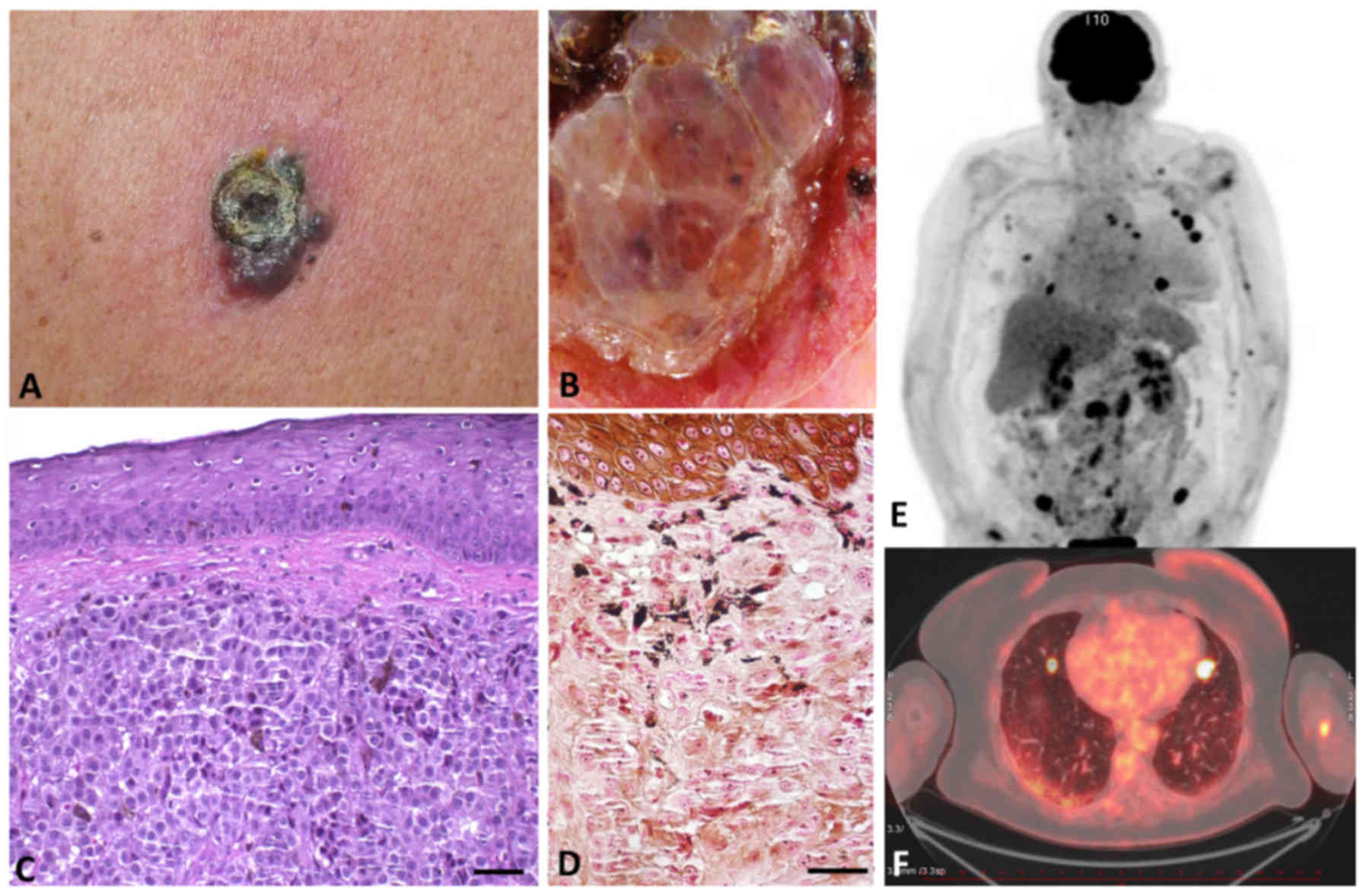
The critical feature associated with melanoma is its
enormous capability to spread and form lymph node or distant
visceral metastases (Fig. 1).
Almost any tissue in the patient's body can host metastatic cells,
and even a small and thin primary tumour can metastasise to the
entire body, leading to the death of the patient (1). Metastatic spread is a complex
multistep process, as was noted almost 200 years ago by surgeon
Stephen Paget, who coined the 'seed and soil' hypothesis (4). Surprisingly, cutaneous melanoma can
spread to different organs without any particular predilection, and
thus differs from, e.g., uveal melanoma of similar histogenesis.
However, the first predictive site of metastatic disease is a lymph
node. The presence of tumour cells in this lymph node is generally
investigated in melanoma patients with tumours thicker than 1 mm.
This procedure is routinely called sentinel lymph node biopsy. The
presence of melanoma cells in the lymph node is a powerful
predictor of melanoma recurrence, but not of survival, in the
melanoma patients (5). VEGF-C,
which is involved in lymphangiogenesis and promotes increased
lymphatic vessel density, can also play a role in lymph node
metastasis (6).
In the case of visceral melanoma metastasis, the
most predictive localisations are the lungs and pleura (7). Lung metastases are also the most
frequent metastases in mouse models of metastatic melanoma
(8). In these mouse models using
the B16 model of melanoma, chloride channel accessory 2 (CLCA2), an
extracellular protein expressed predominantly in the lung, was
identified as a factor mediating interactions with a6^4-integrin,
which is expressed by tumour cells (9). Brain metastases are associated with
poor prognosis. Historically, melanoma patients with brain
metastases have had dismal outcomes and very limited treatment
options. Systemic treatment with BRAF inhibitors and immunotherapy
offers therapeutic responses in up to 55-58% of patients (10). The actual mechanism of brain
metastases is not clear, but mouse models point to some factors
that play a role in this process. The original model suggested a
role of transferrin receptors and their interaction with their
ligand, transferrin, mediating metastases of melanoma cells to the
brain. Another study highlighted the importance of neurotrophins
and neurotrophin receptors in the process of brain-specific
melanoma metastases (11).
Other common sites for melanoma metastases are the
liver (up to 20% of patients), bones (11-17%), or skin and
subcutaneous tissue (12).
Skin metastasis represents haematogenous
dissemination of melanoma cells. Specific interactions between
chemokines C-C motif chemokine receptor 10 (CCR10) and C-C motif
chemokine ligand 27 (CCL27) have been determined as crucial factors
in melanoma metastasis to the skin (13). CCL27 is a chemokine expressed in
the epidermis by normal keratinocytes. In addition, high expression
in supratumoral epidermis is associated with more prolonged
melanoma-specific survival (14).
Presumably, CCL27 interacts with the chemokine receptor CCR10,
which is expressed in melanoma cells. Experiments with blocking
antibodies to CCL27 showed inhibition of development of skin
metastasis in a mouse model (15).
Despite that, no specific biomarker with predictive
potential to determine the metastatic site exists to date. In
melanoma, there is also an observed lack of association between the
site of visceral or lymph node metastasis and either the
clinicopathological variant or location of the primary tumour
(16). The dependence on the
presence of somatic mutations has been reported. One study
suggested that BRAF mutation is associated with lymph node
metastasis as the first metastasis and sentinel lymph node
positivity. BRAF and NRAS mutations were associated with different
metastatic patterns, with metastases more frequently affecting the
central nervous system and the liver. NRAS-mutated tumours formed
lung metastases (17). This
highlights an earlier-unexpected internal heterogeneity of the
group of tumours nowadays collectively called melanoma. Although
intense visceral organ-specific surveillance may be initiated in
patients with tumours harbouring these somatic mutations, this does
not necessarily lead to a decrease in mortality. It is not easy to
understand this metastatic potency of CMM, which represents the
main, frequently fatal, complication in the treatment of patients.
The complexity of these mechanisms is also shown by the concept of
pre-malignant melanocyte dissemination, suggesting that benign
melanocytes may exist at disseminated sites in the body and may be
capable of undergoing malignant progression. It is not uncommon to
find benign melanocytic nevi in the lymph node during sentinel
lymph node biopsy or in non-melanoma patients (up to 7% of
patients) (18). These findings
support the hypothesis mentioned above. It is also critically
important to identify the mechanisms driving the metastatic
behaviour.
CMM cells arise after malignant transformation from
pigment-producing cells called melanocytes. Melanocytes originate
from the embryonic neuroectoderm structure called the neural crest
(NC). NC cells are multipotent stem cells derived from the
neuroectoderm that delaminate from the neural tube in early
vertebrate development (in the 4th week) and migrate throughout the
developing embryo. Consequently, NC cells differentiate into
various cell lineages, including melanocytes (19).
NC cells are unique because of their remarkably
broad differentiation potential (Table
I) (20-22). Once they have reached the final
tissue niche in the skin, NC cells differentiate to melanocytes by
a cascade of events controlled by transcription factors such as
microphthalmia-associated transcription factor (MITF) and
sex-determining region Y-box 10 (Sox10). This process occurs during
the prenatal period of human development (23,24).
The signalling molecules and transcription factors that are
required for NC cell specification, migration and differentiation
form a highly orchestrated gene regulatory network. Every
individual signalling molecule has either individual or
combinatorial roles in transcriptional regulation (25). The precise understanding of this
mechanism seems to be critically important because similar pathways
are activated in malignancy, and they could control the biological
properties of malignant CMM cells (26). The signalling pathways regulating
epithelial to mesenchymal transition (EMT) can be triggered by
transcription factors that are active in both NC development and
cancer progression (27).
Interestingly, both melanoblasts and NC cells also
reside in the bulge region of the hair follicle in the outer root
sheath. In this highly specialised niche, NC cells retain their
multipotency during adult life. NC cells can be isolated and
expanded in vitro with the remarkable features of highly
multipotent stem cells (SCs). It is possible to differentiate NC
cells to various specialised cell types such as melanocytes,
adipocytes, osteoblasts, chondroblasts, smooth muscle cells,
neurons, and Schwann cells (28).
The NC cell phenotype is defined by expression of multiple markers,
and NC cell identification cannot be based on a single molecule. Of
note, there is a significant overlap with the marker profile of CMM
[Table II based on (29-37)]. This highlights the low
differentiation frequently observed in melanoma, where many cells
typically have properties of stem cells (37). These cancer-initiating cells of CMM
have an indispensable role in CMM resistance to therapy,
progression and generalisation (38).
The life-long postnatal presence of NC cells in hair
follicles raises important questions regarding the maintenance of
their multipotency and regulation of their normal behaviour within
this niche. There is strong evidence that the microenvironment is a
critical condition of this steady-state. The signalling cues within
the proper microenvironment, via both extrinsic and intrinsic
factors, orchestrate the interplay necessary for healthy tissue
dynamics. The importance of the normal tissue microenvironment was
highlighted in several studies using transplantation of malignant
cells to animal embryos. In experiments performed in the early
chicken embryo, labelled CMM cells were injected into the region of
the neural tube. It was demonstrated that melanoma cells migrate to
the same regions as the autologous embryonic NC cells (39). Similar experiments performed later
in zebrafish embryos supported these findings. Both the embryonic
NC cells and the cells of CMM in zebrafish express specific protein
crestin, which is absent in normal melanocytes (40).
Taking into account the low differentiation status
of NC cells and their natural migratory activity, the similarity of
CMM and NC cells can also explain the highly metastatic behaviour
observed in melanoma in the clinic.
Similarly to other types of malignant tumours, cells
of CMM can also be detected in the circulation. These circulating
melanoma cells harbour the functional properties of cells of the
primary tumour, including their SC-like properties (41,42).
These cells leave the primary tumour and penetrate the vessels and
use them as a highway for dissemination through the patient's body
to target the organ/tissue where they form metastases. Using an
identical vascular path, the normal adult tissue SCs can migrate in
order to facilitate body repair processes during wound healing
(43,44). From this point of view and based on
histological/molecular similarity, cancer again resembles wound
healing. With a certain hyperbole, cancer can be seen as a
distorted cascade of wound repair events (45). These data can also predict the
great invasive metastatic potential of CMM.
Malignant cells define the type of tumour. However,
there are other non-cancerous populations forming the tumour
stroma. It is the interaction of both components of this
microenvironment that finally defines the biological behaviour of
the tumour. It is truly applicable to solid tumours in general, and
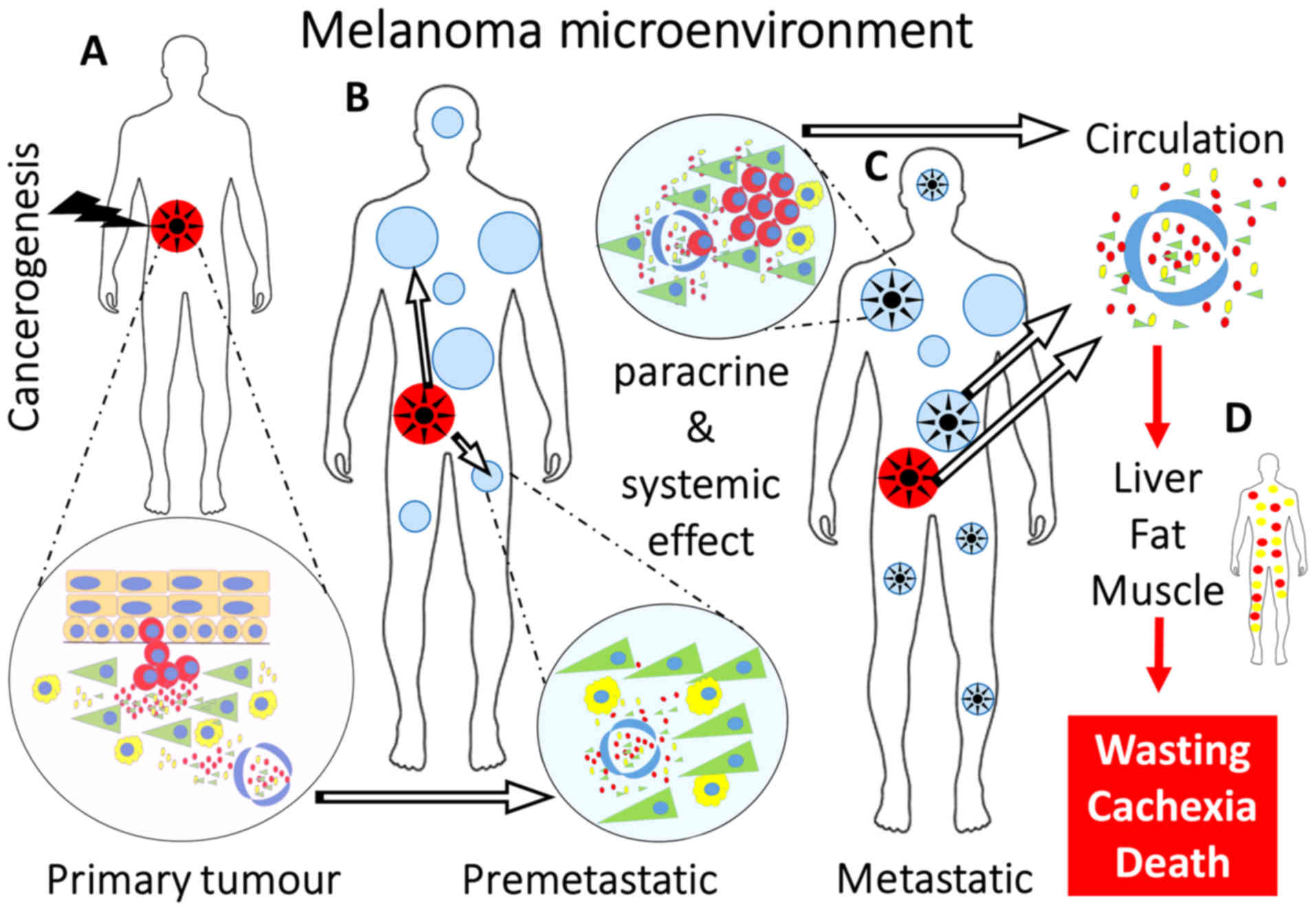
CMM is no exception (46-48). Concerning CMM, the cancer
microenvironment is formed by cancer-associated fibroblasts (CAFs)
and several types of leukocytes, as comprehensively reviewed by
Lacina et al (49)
(Fig. 2A).
The origin of CAFs is not fully understood. Local
normal dermal fibroblasts, attracted mesenchymal SCs and pericytes
are frequently mentioned as source cell populations from which CAFs
are recruited (50). However, the
transition of cancer cells to CAFs cannot be entirely excluded,
although its likeliness is not very high. This fact is difficult to
prove in the experimental model (51).
Treg lymphocytes, tumour-associated macrophages and
myeloid-derived immunosuppressive cells stimulating the CMM
progression, as well as NK cells, macrophages and CD8-positive T
lymphocytes, are attracted to the CMM site. Interestingly, CAFs
stimulate the activity of immune cells supporting melanoma cells
and inhibit the cancer-suppressing cells (52,53).
Unlike in other epidermal tumours, keratinocytes are
also an important component of the CMM microenvironment. Melanoma
cells can stimulate surrounding keratinocytes (54). On the other hand, keratinocytes
control growth and differentiation of melanocytes and potentiate
the invasiveness of melanoma cells during early progression as
observed in a reconstructed skin model (55).
Cell-cell adhesion molecules (cadherins) and
cell-extracellular matrix adhesion proteins (integrins) play a
critical role in the regulation of cancer invasion and metastasis.
Many members of the cadherin superfamily play an important role in
cancer biology. However, the most significant explanation is seen
in the E-/N-cadherin switch, and its role in epithelial to
mesenchymal transition (EMT), in cancer progression. N-cadherin
expression in CMM cells helps cancer cells to interact with
fibroblasts and extracellular matrix and stimulates the invasive
potential of melanoma cells and their proliferation, but also
activation of PI3/AKT, mTOR, and ERK kinase. Inhibition of
N-cadherin represents an interesting possibility, with potential
clinical use (56).
Intercellular contacts of normal melanocytes, or
malignant melanoma cells, respectively, and their non-cancerous
neighbours within the tissue environment influence their properties
mutually (62). Keratinocytes
reduce expression of N-cadherin not only via cell-cell contacts,
but also via cell-derived extracellular matrix and conditioned
medium with calcium regulators (57). These findings support the
importance of the balance in communications between melanoma cells
and non-cancerous cells in the melanoma microenvironment. Integrins
β1 and β3, as adhesion cell-extracellular matrix proteins, are
differentially expressed during the transformation of melanoma
radial growth to the vertical invasion (58,59).
The differentiation status of melanoma cells and the ability to
invade the surrounding tissue also highlights this impact. For
example, the expression of connexin-43 in CMM cells indicates the
ability of CMM cells to metastasise (52,60,61).
Expression of desmoglein-2, which participates in the contacts with
keratinocytes, has an inhibitory effect on CMM cell migration.
On the other hand, the expression of desmoglein-2
promotes the vasculogenic mimicry of CMM cells, which is associated
with a poor outcome of patients (62,63).
Furthermore, in melanoma cells that do not express β3 integrins, β1
integrins instead play a role in promoting their transendothelial
migration by binding to vascular cell adhesion molecule 1 (VCAM-1)
(64). Integrins also play an
important role in connecting the extracellular matrix with the
melanocyte and melanoma cell cytoskeleton. Cytoskeletal
rearrangements, such as the increase of the overall contractility,
impact cell mechanical properties and cell deformability. These
changes may then potentiate prometastatic phenotypes of melanoma
cells. Expression of avp3 integrin increases elasticity in human
melanoma cells in adherent and non-adherent conditions (65). Intercellular contacts and molecules
play an important role in the mechanisms of targeted therapy.
Targeting of the CMM cell surface receptor Notch-dependent pathway
improves the activity of Erk inhibitors in BRAF-V600E mutated
tumours. Further, it can be combined with inhibition of ERBB3 to
suppress melanoma cell growth (63). On the other hand, Notch expression
in CAFs reduces the growth and migration potential of melanoma
cells (66).
The paracrine mode of signalling between cancerous
and non-cancerous cells in CMM has been extensively studied
(Fig. 2B and C). For example,
CAFs, keratinocytes and infiltrating immune cells produce a variety
of growth factors/cytokines/chemokines that significantly influence
the biological properties of malignant CMM cells (67,68)
(Table III). Interestingly, many
of these factors are also associated with skin ageing.
Collectively, these bioactive molecules are called the
senescence-associated secretome (69). As expected, chemokines attract
inflammatory cells to the tumour site. However, they also have
multiple other functions (70).
CMM cells express receptors for CXCL1 and interleukin (IL)8, and
these factors enhance their invasiveness (71,72).
Antagonists of CXCR1/2 receptors have a well-documented inhibitory
effect on migration of CMM cells (73). The IL8 chemokine also stimulates
vascularisation of CMM (74,75),
and in general, its high expression indicates poor prognosis of CMM
patients (76).
Expression of chemokine CXCL16 seems to participate
in the malignant transformation of a melanocytic nevus to CMM
(77). Activation of CXCR6
recognising this chemokine induces SC-like properties in CMM cells
and initiates their migration (78).
IL1β is predominantly produced by macrophages. It
participates in the progression of CMM in collaboration with IL8
and caspase recruitment domain family member 8 (CARD8) (79).
IL6 is a crucial factor initiating the immune
response. IL6 has a multifaceted role in cancer progression
(80). While the initial stage of
CMM growth can be inhibited by IL6 (81), the more advanced stages are
associated with production of this interleukin (82,83).
IL6, frequently in cooperation with IL8, exhibits an additive
effect on WNT5A in the stimulation of CMM cell invasiveness
(67,84). IL6 induces Twist and N-cadherin
expression in CMM angiogenesis in a mechanism dependent on the p50
subunit of nuclear factor kb
(85).
Glycoprotein aggrecan is produced by cells of the
CMM, CAFs and keratinocytes. It is usually secreted during the
process of chondroblast differentiation, and it has an inhibitory
effect on CMM progression (90). A
similar anti-CMM effect is produced by insulin-like growth
factor-binding protein 7 (IGFBP-7), namely in BRAF-mutated
V600E-positive dysplastic nevi (91). On the other hand, heparin-binding
EGF-like growth factor has a stimulatory impact on CMM growth
(92). The same result was
described in the case of another growth factor, neurotrophin
(93,94). miRNA-328 controls production of
TGF-p2, and attenuation of its expression has a strong inhibitory
effect on CMM cell proliferation (95). VEGFA and VEGFC are generally
responsible for the activation of CMM neovascularisation by
blood/lymphatic vessels that support the CMM growth and
progression. Connective tissue growth factor has a synergistic
effect on VEGFA and stimulates the tumour site neovascularisation
(96).
As mentioned earlier, CAFs do not support tumour
growth and metastases exclusively. CAFs are also implicated in the
acquisition of resistance to the targeted therapy in BRAF-mutated
melanomas. Under the influence of BRAF inhibitors, CAFs secrete
factors that contribute to CMM cell survival and melanoma
resistance. CAFs release factors such as hepatocyte growth factor
(HGF) and neuregulin 1 (NRG1), which can trigger alternative
cascades in MAP kinase signalling (97,98).
Concluding this paragraph, paracrine signalling
represents a critical aspect in the control of biological
properties of CMM. In addition to the crosstalk between melanoma
cells, this process includes an exchange of information between CMM
cells and non-malignant cells of the microenvironment.
In addition to paracrine signalling via soluble
products, exosomes represent another tool of CMM cell communication
with non-cancerous partners within the microenvironment. All
affected cell populations of the cancer microenvironment produce
these bodies. Exosomes thus influence CMM cell biological
properties (99). Exosomes
stimulate CMM cell metastasis via support of the
epithelial-mesenchymal transition. Exosomes influence
vascularisation of the lymph node in order to prepare the vascular
bed for the metastasising (100,101). Exosomes significantly participate
in the regulation of local invasiveness and also in the entrance of
melanoma cells to the target organs (102). This effect is frequently
associated with the presence of miRNAs in CMM-derived exosomes. It
was confirmed both in vitro and in vivo in clinical
material (103). Exosomes exert a
robust immunosuppressive effect on the cancer microenvironment,
where they inhibit IL2-dependent proliferation of CD8-positive T
lymphocytes (104). Moreover, CMM
exosomal miRNA-125b-5p induces a tumour-promoting phenotype in
macrophages (105). These changes
can induce a mixed M1, and M2 tumour-promoting macrophage
activation included production of CCL22, IL-12B, IL-1β, IL-6,
i-NOS, and TNF-α (106). These
data highlight exosomes as a critically important component of the
CMM microenvironment significantly participating in its biological
properties, with the ability to stimulate the immune response of
the melanoma microenvironment.
Serological biomarkers represent a diverse group of
biomolecules with importance in diagnosis, staging, and monitoring
the therapeutic response. Serum lactate dehydrogenase (LDH) is the
only serum biomarker that has been accepted as a prognostic
biomarker for routine clinical use in melanoma patients with a
predictive therapeutic outcome and has been implemented in the
American Joint Committee on Cancer (8th edition) staging system
(107). Routinely used S100B
(S100 calcium binding protein B) protein is highly specific for
melanoma patients. Its increased levels can be detected in patients
with advanced melanoma during melanoma prognosis (108). Another serological protein is MIA
(melanoma inhibitory activity), which interacts with extracellular
matrix proteins. Its expression can also be detected in normal
tissue such as cartilage. In pathological processes, its
overexpression is observed in breast cancer or colorectal cancer,
in addition to melanoma (109).
Introduction of a new treatment strategy for
advanced melanoma leads to the search for new biomarkers to improve
both prognostic and predictive outcome. Likely, the high intensity
of molecular exchange between cancer cells and other members of the
microenvironment via cytokines, chemokines and growth factors can
lead to leakage out from the tumour microenvironment, and mediators
can be consequently detected in systemic circulations in the serum
(110,111) (Fig.
2A-D). Therapy by monoclonal antibodies targeting immune
checkpoint inhibitors is one of the most potent treatments of CMM
patients. Measurement of current concentrations and dynamics of
these mediators in the serum can have the potential of liquid
biopsy. Indeed, the serum protein signature even reflects the
efficiency of anti-PD-1 therapy of CMM patients and can be
substantial for therapeutic indications (112).
Similarly to other types of tumours, an elevated
serum level of IL6 in CMM has been observed (111), which has reached some prognostic
validity (113). Serum elevation
of IL6 sensibly reflected the tumour burden and indicated a relapse
of the disease or insensitivity to tumour therapy in several
studies (114-116).
Based on the selected examples, it is possible to
demonstrate the systemic effect of CMM. The serum/plasma of CMM
patients contains numerous bioactive proteins and exosomes that are
transported to the distant parts of the patient's body through the
vessels (Fig. 2B-D). These factors
participate in the preparation of a premetastatic niche suitable
for cancer cell homing and metastasis formation (124). Under the influence of CMM-derived
exosomes, the dermal fibroblasts reprogram their metabolism
significantly (125). The distant
dermal fibroblasts from CMM patients at the stage of metastatic
tumour dissemination differ from the normal dermal fibroblasts from
healthy donors. The phenotype of distant dermal fibroblasts, as
well as the expression profile and methylation profile of gene
promoters, is shifted closer to CAFs (126). Due to this activation, it is
possible to hypothesise that the tissue microenvironment in the
distant body parts is influenced by the released bioactive factors
from the melanoma microenvironment. It is, therefore, likely that
melanoma becomes a systemic disease very early. If so, it is the
signalling in the primary tumour that already prepares the rest of
the organism to host CMM cells and facilitate metastases (114) (Fig.
2B).
In recent years, the term 'migrastatics' has been
introduced for drugs interfering with all modes of cancer cell
invasion (115). Migrastatics
inhibit local invasion and consequent metastasis. This group of
drugs was recently established to define and distinguish them from
conventional cytostatic drugs that traditionally target cell
proliferation. Malignant melanoma, therefore, seems to be a
tempting disease for validation of this concept.
Endothelial cells of capillaries are also an
important structure of the cancer microenvironment. Migrating CMM
cells adhere to the capillary endothelium, intercalate between
endothelial cells, and migrate throughout the vessels in both
directions. From the endothelial cell perspective, this process is
not passive. Endothelial cells actively participate in
extravasation, where the role of N-cadherin has been broadly
investigated (117). It seems to
be also controlled by CD146, which cooperates with VEGFA. CD146 is
also elevated in the patients' serum/plasma (127).
On the other hand, VE-cadherin expressed on the
surface of endothelial cells prevents the migration of cancer cells
through the endothelium of capillaries. VE-cadherin must,
therefore, be eliminated from the site of malignant cell migration
(128). P-selectin has an
essential role in the recruitment of inflammatory cells to the site
of inflammation, so-called homing.
P-selectin expression on endothelial cells is under
the control of the local microenvironment. Expression of this
molecule on endothelial cells and blood platelets seems to be a
prerequisite for successful metastasising of CMM cells to the
target tissue (129,130). P-selectin expression on the
surface of endothelial cells is induced by STAT3 activation
(129). IL6 is available in the
serum of CMM patients, and it is known as a potent activator of
STAT3. The observation that capsular polysaccharides from E. coli
attenuate adhesion of CMM cells to the endothelium via P-selectin
demonstrates the specificity of this interaction (130). Endogenous lectin galectin-3
locally accumulates in inflamed tissues, including endothelium.
This lectin also enhances invasion of CMM cells, e.g., to the lungs
(131). These examples show that
an imbalance in serum proteins can participate in the process of
extravasation of CMM cells to the target tissue and
metastasising.
Therefore, the combination of migrastatics with
other groups of traditional oncologic drugs may be possible. Beyond
that, we suggest that directed therapy (biologics, small-molecule
receptor-associated kinase inhibitors) against the most prominent
inflammatory cytokines, namely IL6, could bring highly desirable
synergism. However, it seems evident that inhibition of the IL6
signalling axis is not sufficient and must, therefore, be
accompanied by simultaneous blockade of other proteins/receptors
such as IL8, VEGFA and MFGE8 (48,132). The therapeutic blockade of IL6,
in combination with checkpoint inhibitor anti-PD1, represents an
interesting possibility of overcoming some immunological mechanisms
of resistance. IL6 blockade upregulated expression of PD-L1 on
melanoma cells in a mouse model and may sensitise melanoma to this
treatment (133). These findings
underscore the importance of the IL6-PD1/PD-L1 crosstalk in the
tumour microenvironment of melanoma.
The local microenvironment and its control by
bioactive factors can be a highly relevant target in the prevention
of the deadly complication of malignant disease (Fig. 2D).
Advanced stages of cancer, including CMM, are
associated with metastasising, which in the case of CMM has a
character of extensive generalisation. The increasing burden of
tumour cells generates an imbalance in growth factors, cytokines
and chemokines, among which IL6 seems to have the leading position
(80). This stage of the disease
is usually terminated by cancer-associated cachexia (CAC), which
affects approximately 16 patients per 100,000 individuals (134) (Fig.
2D). CAC is a highly complex and multifaceted catabolic process
(135). IL6, in cooperation with
TNFa and IL1ß influences the metabolism of striated muscle fibres,
adipocytes and hepatocytes (136,137). The level of the mentioned factors
in the serum can even predict the onset of CAC and survival of
cancer patients (138). The
terminal stage of cancer is also associated with decreased food
intake in cancer patients, which is called anorexia (139). IL6 seems to be linked to the
control of food intake, where it inhibits the appetite and
participates in the development of anorexia (140). TNFa and IL6 can cross the
blood-brain barrier (141). TNFa,
IL1 and IL6 can interact with hypothalamic neurons and affect the
serotoninergic metabolism, which can be reflected by decreased food
intake (142). Patients with
advanced cancer frequently suffer from depression that seems to be
associated with elevation of IL6, IL10 and TNFa (143). On the other hand, the serum
levels of IL6 (and also IL8) are significantly elevated in
tumour-free patients with bipolar disease, but not with major
depressive disorder (144). This
section demonstrates that factors produced by the cancer ecosystem
have a strong systemic effect by which they influence the metabolic
functions of cancer patients, resulting in wasting and death.
CMM, similarly to the majority of cancers, can be
characterised as a genetic abnormality and a regulation failure in
which cancer cells employ predestined pathophysiological pathways
that are normally activated in the course of organism growth,
tissue regeneration and repair. This deregulation is typically
associated with accumulation of mutations acquired during the
ageing of the individual.
The progression of CMM from tumour initiation to the
systemic effect on the patient's metabolism is organised according
to a quite uniform scenario (Fig.
2A-D). The intercellular crosstalk within this ecosystem is
mediated either directly by intercellular contacts, or indirectly
by paracrine secretion of numerous active molecules. This
interconnection strengthens the malignant potential of cancer
cells, and it can inhibit the anticancer immune response or protect
malignant cells from the harmful effect of oncological therapy. A
plethora of bioactive factors are transported via vessels and
significantly influence even the distant tissue. Collectively,
these factors prepare a suitable microenvironment for the malignant
cell extravasation and metastasising, the premetastatic niche. The
increasing mass of CMM cells in the body of patients generates a
cancer-induced profile of inflammatory mediators in the patients'
serum. The systemic availability of these bioactive molecules
triggers mental disorders, depression, and mental
anorexia-associated problems with food intake. Wasting leads to
cancer cachexia and death. Based on this scenario, it is evident
that besides the conventional anticancer therapy, it would be
necessary to influence the migration of CMM cells and their
metastasising - the concept of migrastatics (115). Because the CMM microenvironment
stimulates malignant cell invasiveness (145), targeting both cancerous and
non-cancerous cells of the tumour ecosystem and their products
seems to be a promising approach. IL6 and its signalling pathway
influence CMM cell growth and migration, but it can also positively
affect the entire patient metabolism and mental status (82). Therefore, targeting the
IL6/IL6R/STAT3 axis as a new therapeutic modality was
enthusiastically expected, but, unfortunately, the reality did not
meet this expectation (146). The
progress in the detection of clinically relevant markers using a
robust omics approach that includes stromal factors can be
translated into personalised therapy of CMM (147). For example, a combination of
blocking the anti-IL6 axis with drugs blocking other signalling
pathways seems to be promising for future trials (48). It can be hypothesised that the
progress in diagnostics and therapy covering the complex ecosystem
of melanoma can bring some benefit to CMM patients.
Authors are grateful to Dr Sarka Takacova for her
assistance in English language editing of this revised version.
The project 'Centre for Tumour Ecology-Research of
the Cancer Microenvironment Supporting Cancer Growth and Spread'
(reg. no. CZ.02.1.01/0.0/0.0/16_019/00007 85) is supported by the
Operational Programme 'Research, Development and Education'.
Additional support was received from the Ministry of Education,
Youth and Sports of CR within the National Sustainability Programme
II (Project BIOCEV-FAR, reg. no. LQ1604), project BIOCEV
(CZ.1.05/1.1.00/02.0109), grant no. CZ.1.05/2.1.00/19.0400
supported by the Research and Development for Innovations
Operational Programme, co-financed by the European Regional
Development Fund and the state budget of the Czech Republic, from
the Grant Agency of the Czech Republic, project no. 19-05048S and
from Charles University project PROGRES Q28.
All data and information are supported by relevant
references.
Conceptualisation, data collection and manuscript
preparation were carried out by KSm, OK and LL Manuscript
preparation was conducted by JK, JŠ, KSt and BD.
Not applicable.
The authors declare no competing interests.
|
1
|
Sacchetto L, Zanetti R, Comber H,
Bouchardy C, Brewster DH, Broganelli P, Chirlaque MD, Coza D,
Galceran J, Gavin A, et al: Trends in incidence of thick, thin and
in situ melanoma in Europe. Eur J Cancer. 92:108–118. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Rabbie R, Ferguson P, Molina-Aguilar C,
Adams DJ and Robles-Espinoza CD: Melanoma subtypes: Genomic
profiles, prognostic molecular markers and therapeutic
possibilities. J Pathol. 247:539–551. 2019. View Article : Google Scholar :
|
|
3
|
Lorentzen HF: Targeted therapy for
malignant melanoma. Curr Opin Pharmacol. 46:116–121. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Paget S: The distribution of secondary
growths in cancer of the breast. Lancet. 133:571–573. 1889.
View Article : Google Scholar
|
|
5
|
Faries MB, Thompson JF, Cochran AJ,
Andtbacka RH, Mozzillo N, Zager JS, Jahkola T, Bowles TL, Testori
A, Beitsch PD, et al: Completion dissection or observation for
sentinel-node metastasis in melanoma. N Engl J Med. 376:2211–2222.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Nathanson SD: Insights into the mechanisms
of lymph node metastasis. Cancer. 98:413–423. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Barth A, Wanek LA and Morton DL:
Prognostic factors in 1,521 melanoma patients with distant
metastases. J Am Coll Surg. 181:193–201. 1995.PubMed/NCBI
|
|
8
|
Damsky WE Jr and Bosenberg M: Mouse
melanoma models and cell lines. Pigment Cell Melanoma Res.
23:853–859. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Abdel-Ghany M, Cheng HC, Elble RC and
Pauli BU: The breast cancer ß 4 integrin and endothelial human
CLCA2 mediate lung metastasis. J Biol Chem. 276:25438–25446. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Tawbi HA, Boutros C, Kok D, Robert C and
McArthur G: New era in the management of melanoma brain metastases.
Am Soc Clin Oncol Educ Book. 38:741–750. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Menter DG, Herrmann JL and Nicolson GL:
The role of trophic factors and autocrine/paracrine growth factors
in brain metastasis. Clin Exp Metastasis. 13:67–88. 1995.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Balch CM, Houghton AN, Sober AJ and Soong
S: Cutaneous Melanoma, 4th Edition. Dermatologic Surg. 31:1715.
2005. View Article : Google Scholar
|
|
13
|
Murakami T, Cardones AR and Hwang ST:
Chemokine receptors and melanoma metastasis. J Dermatol Sci.
36:71–78. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Martinez-Rodriguez M, Thompson AK and
Monteagudo C: High CCL27 immunoreactivity in 'supratumoral'
epidermis correlates with better prognosis in patients with
cutaneous malignant melanoma. J Clin Pathol. 70:15–19. 2017.
View Article : Google Scholar
|
|
15
|
Ben-Baruch A: Organ selectivity in
metastasis: Regulation by chemokines and their receptors. Clin Exp
Metastasis. 25:345–356. 2008. View Article : Google Scholar
|
|
16
|
Marcoval J, Ferreres JR, Martrn C, Gomez
S, Penrn RM, Ochoa de Olza M and Fabra À: Patterns of Visceral
Metastasis in Cutaneous Melanoma: A Descriptive Study. Actas
Dermosifiliog. 104:593–597. 2013. View Article : Google Scholar
|
|
17
|
Adler NR, Wolfe R, Kelly JW, Haydon A,
McArthur GA, McLean CA and Mar VJ: Tumour mutation status and sites
of metastasis in patients with cutaneous melanoma. Br J Cancer.
117:1026–1035. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Holt JB, Sangueza OP, Levine EA, Shen P,
Bergman S, Geisinger KR and Creager AJ: Nodal melanocytic nevi in
sentinel lymph nodes. Correlation with melanoma-associated
cutaneous nevi. Am J Clin Pathol. 121:58–63. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Ji Y, Hao H, Reynolds K, McMahon M and
Zhou CJ: Wnt signaling in neural crest ontogenesis and oncogenesis.
Cells. 8:11732019. View Article : Google Scholar :
|
|
20
|
Lim J, Thiery JP, Kassem Y, Kalcheim C,
Moens CB, Burden SJ and Granato M: Epithelial-mesenchymal
transitions: Insights from development. Development. 139:3471–3486.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Mayor R and Theveneau E: The neural crest.
Development. 140:2247–2251. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Hall BK: The neural crest and neural crest
cells: Discovery and significance for theories of embryonic
organization. J Biosci. 33:781–793. 2008. View Article : Google Scholar
|
|
23
|
Mort RL, Jackson IJ and Patton EE: The
melanocyte lineage in development and disease. Development.
142:620–632. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Duband JL, Monier F, Delannet M and
Newgreen D: Epithelium-mesenchyme transition during neural crest
development. Acta Anat (Basel). 154:63–78. 1995. View Article : Google Scholar
|
|
25
|
Vega-Lopez GA, Cerrizuela S and Aybar MJ:
Trunk neural crest cells: Formation, migration and beyond. Int J
Dev Biol. 61:5–15. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Larribère L and Utikal J: Stem
cell-derived models of neural crest are essential to understand
melanoma progression and therapy resistance. Front Mol Neurosci.
12:1112019. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Gallik KL, Treffy RW, Nacke LM, Ahsan K,
Rocha M, Green-Saxena A and Saxena A: Neural crest and cancer:
Divergent travelers on similar paths. Mech Dev. 148:89–99. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Sieber-Blum M and Grim M: The adult hair
follicle: Cradle for pluripotent neural crest stem cells. Birth
Defects Res C Embryo Today. 72:162–172. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Person F, Wilczak W, Hube-Magg C,
Burdelski C, Moller-Koop C, Simon R, Noriega M, Sauter G, Steurer
S, Burdak-Rothkamm S, et al: Prevalence of pIII-tubulin (TUBB3)
expression in human normal tissues and cancers. Tumour Biol.
39:10104283177121662017. View Article : Google Scholar
|
|
30
|
Stasiak M, Boncela J, Perreau C, Karamanou
K, Chatron-Colliet A, Proult I, Przygodzka P, Chakravarti S,
Maquart FX, Kowalska MA, et al: Lumican inhibits SNAIL-induced
melanoma cell migration specifically by blocking MMP-14 activity.
PLoS One. 11:e01502262016. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Yang X, Liang R, Liu C, Liu JA, Cheung
MPL, Liu X, Man OY, Guan XY, Lung HL and Cheung M: SOX9 is a
dose-dependent metastatic fate determinant in melanoma. J Exp Clin
Cancer Res. 38:172019. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Lee H, Torres FX, McLean SA, Chen R and
Lee MW: Immunophenotypic heterogeneity of primary sinonasal
melanoma with aberrant expression of neuroendocrine markers and
calponin. Appl Immunohistochem Mol Morphol. 19:48–53. 2011.
View Article : Google Scholar
|
|
33
|
Tudrej KB, Czepielewska E and
Kozlowska-Wojciechowska M: SOX10-MITF pathway activity in melanoma
cells. Arch Med Sci. 13:1493–1503. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Iwakami Y, Yokoyama S, Watanabe K and
Hayakawa Y: STAM-binding protein regulates melanoma metastasis
through SLUG stabilization. Biochem Biophys Res Commun.
507:484–488. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Goding CR and Arnheiter H: MITF-the first
25 years. Genes Dev. 33:983–1007. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Campbell K, Kumarapeli AR, Gokden N, Cox
RM, Hutchins L and Gardner JM: Metastatic melanoma with
dedifferentiation and extensive rhabdomyosarcomatous heterologous
component. J Cutan Pathol. 45:360–364. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Krejci E and Grim M: Isolation and
characterization of neural crest stem cells from adult human hair
follicles. Folia Biol (Praha). 56:149–157. 2010.
|
|
38
|
Marzagalli M, Raimondi M, Fontana F,
Montagnani Marelli M, Moretti RM and Limonta P: Cellular and
molecular biology of cancer stem cells in melanoma: Possible
therapeutic implications. Semin Cancer Biol. 59:221–235. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Kasemeier-Kulesa JC, Teddy JM, Postovit
LM, Seftor EA, Seftor REB, Hendrix MJC and Kulesa PM: Reprogramming
multipotent tumor cells with the embryonic neural crest
microenvironment. Dev Dyn. 237:2657–2666. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Kaufman CK, Mosimann C, Fan ZP, Yang S,
Thomas AJ, Ablain J, Tan JL, Fogley RD, van Rooijen E, Hagedorn EJ,
et al: A zebrafish melanoma model reveals emergence of neural crest
identity during melanoma initiation. Science. 351:aad21972016.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Agnoletto C, Corrà F, Minotti L,
Baldassari F, Crudele F, Cook WJJ, Di Leva G, d'Adamo AP, Gasparini
P and Volinia S: Heterogeneity in circulating tumor cells: The
relevance of the stem-cell subset. Cancers (Basel). 11:112019.
View Article : Google Scholar
|
|
42
|
Gkountela S and Aceto N: Stem-like
features of cancer cells on their way to metastasis. Biol Direct.
11:332016. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Empringham B, Chiang KY and Krueger J:
Collection of hematopoietic stem cells and immune effector cells in
small children. Transfus Apheresis Sci. 57:614–618. 2018.
View Article : Google Scholar
|
|
44
|
Feehan J, Nurgali K, Apostolopoulos V, Al
Saedi A and Duque G: Circulating osteogenic precursor cells:
Building bone from blood. EBioMedicine. 39:603–611. 2019.
View Article : Google Scholar :
|
|
45
|
Ratajczak MZ, Bujko K, Mack A, Kucia M and
Ratajczak J: Cancer from the perspective of stem cells and
misappropriated tissue regeneration mechanisms. Leukemia.
32:2519–2526. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Lacina L, Plzak J, Kodet O, Szabo P,
Chovanec M, Dvorankova B and Smetana K Jr: Cancer microenvironment:
What can we learn from the stem cell niche. Int J Mol Sci.
16:24094–24110. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Hill BS, Pelagalli A, Passaro N and
Zannetti A: Tumor-educated mesenchymal stem cells promote
pro-metastatic phenotype. Oncotarget. 8:73296–73311. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Plzak J, Boucek J, Bandurova V, Kolar M,
Hradilova M, Szabo P, Lacina L, Chovanec M and Smetana K Jr: The
head and neck squamous cell carcinoma microenvironment as a
potential target for cancer therapy. Cancers (Basel). 11:112019.
View Article : Google Scholar
|
|
49
|
Lacina L, Kodet O, Dvorankova B, Szabo P
and Smetana K Jr: Ecology of melanoma cell. Histol Histopathol.
33:247–254. 2018.
|
|
50
|
Preisner F, Leimer U, Sandmann S, Zoernig
I, Germann G and Koellensperger E: Impact of human adipose
tissue-derived stem cells on malignant melanoma cells in an in
vitro co-culture model. Stem Cell Rev Rep. 14:125–140. 2018.
View Article : Google Scholar
|
|
51
|
Dvorankova B, Smetana K Jr, Rihova B,
Kucera J, Mateu R and Szabo P: Cancer-associated fibroblasts are
not formed from cancer cells by epithelial-to-mesenchymal
transition in nu/nu mice. Histochem Cell Biol. 143:463–469. 2015.
View Article : Google Scholar
|
|
52
|
Inada M, Takita M, Yokoyama S, Watanabe K,
Tominari T, Matsumoto C, Hirata M, Maru Y, Maruyama T, Sugimoto Y,
et al: Direct melanoma cell contact induces stromal cell autocrine
prostaglandin E2-EP4 receptor signaling that drives tumor growth,
angiogenesis, and metastasis. J Biol Chem. 290:29781–29793. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Ziani L, Safta-Saadoun TB, Gourbeix J,
Cavalcanti A, Robert C, Favre G, Chouaib S and Thiery J:
Melanoma-associated fibroblasts decrease tumor cell susceptibility
to NK cell-mediated killing through matrix-metalloproteinases
secretion. Oncotarget. 8:19780–19794. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Kodet O, Lacina L, Krejci E, Dvorankova B,
Grim M, Stork J, Kodetova D, Vlcek C, Sachova J, Kolar M, et al:
Melanoma cells influence the differentiation pattern of human
epidermal keratinocytes. Mol Cancer. 14:12015. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Van Kilsdonk JWJ, Bergers M, Van Kempen
LCLT, Schalkwijk J and Swart GWM: Keratinocytes drive melanoma
invasion in a reconstructed skin model. Melanoma Res. 20:372–380.
2010.PubMed/NCBI
|
|
56
|
Ciolczyk-Wierzbicka D and Laidler P: The
inhibition of invasion of human melanoma cells through N-cadherin
knock-down. Med Oncol. 35:422018. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Chung H, Jung H, Jho EH, Multhaupt HAB,
Couchman JR and Oh ES: Keratinocytes negatively regulate the
N-cadherin levels of melanoma cells via contact-mediated calcium
regulation. Biochem Biophys Res Commun. 503:615–620. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Nikkola J, Vihinen P, Vlaykova T,
Hahka-Kemppinen M, Heino J and Pyrhonen S: Integrin chains |31 and
alphav as prognostic factors in human metastatic melanoma. Melanoma
Res. 14:29–37. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Van Belle PA, Elenitsas R, Satyamoorthy K,
Wolfe JT, Guerry D IV, Schuchter L, Van Belle TJ, Albelda S, Tahin
P, Herlyn M, et al: Progression-related expression of |33 integrin
in melanomas and nevi. Hum Pathol. 30:562–567. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Li G, Satyamoorthy K, Meier F, Berking C,
Bogenrieder T and Herlyn M: Function and regulation of
melanoma-stromal fibroblast interactions: When seeds meet soil.
Oncogene. 22:3162–3171. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Brandner JM and Haass NK: Melanoma's
connections to the tumour microenvironment. Pathology. 45:443–452.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Peitsch WK, Doerflinger Y, Fischer-Colbrie
R, Huck V, Bauer AT, Utikal J, Goerdt S and Schneider SW:
Desmoglein 2 depletion leads to increased migration and
upregulation of the chemoattractant secretoneurin in melanoma
cells. PLoS One. 9:e894912014. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Tan LY, Mintoff C, Johan MZ, Ebert BW,
Fedele C, Zhang YF, Szeto P, Sheppard KE, McArthur GA, Foster-Smith
E, et al: Desmoglein 2 promotes vasculogenic mimicry in melanoma
and is associated with poor clinical outcome. Oncotarget.
7:46492–46508. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Klemke M, Weschenfelder T, Konstandin MH
and Samstag Y: High affinity interaction of integrin a4pi (VLA-4)
and vascular cell adhesion molecule 1 (VCAM-1) enhances migration
of human melanoma cells across activated endothelial cell layers. J
Cell Physiol. 212:368–374. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Lacaria L, Lange JR, Goldmann WH, Rico F
and Alonso JL: av|33 integrin expression increases elasticity in
human melanoma cells. Biochem Biophys Res Commun. 525:836–840.
2020. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Bedogni B: Notch signaling in melanoma:
Interacting pathways and stromal influences that enhance Notch
targeting. Pigment Cell Melanoma Res. 27:162–168. 2014. View Article : Google Scholar
|
|
67
|
Jobe NP, Rosel D, Dvorankova B, Kodet O,
Lacina L, Mateu R, Smetana K and Brabek J: Simultaneous blocking of
IL-6 and IL-8 is sufficient to fully inhibit CAF-induced human
melanoma cell invasiveness. Histochem Cell Biol. 146:205–217. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Jobe NP, Zivicova V, Mifkova A, Rosel D,
Dvorankova B, Kodet O, Strnad H, Kolar M, Sedo A, Smetana K Jr, et
al: Fibroblasts potentiate melanoma cells in vitro invasiveness
induced by UV-irradiated keratinocytes. Histochem Cell Biol.
149:503–516. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Strnadova K, Sandera V, Dvorankova B,
Kodet O, Duskova M, Smetana K and Lacina L: Skin aging: The dermal
perspective. Clin Dermatol. 37:326–335. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Payne AS and Cornelius LA: The role of
chemokines in melanoma tumor growth and metastasis. J Invest
Dermatol. 118:915–922. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Gebhardt C, Averbeck M, Viertel A, Kauer
F, Saalbach A, Anderegg U and Simon JC: Ultraviolet-B irradiation
enhances melanoma cell motility via induction of autocrine
interleukin 8 secretion. Exp Dermatol. 16:636–643. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Araki K, Shimura T, Yajima T, Tsutsumi S,
Suzuki H, Okada K, Kobayashi T, Raz A and Kuwano H: Phosphoglucose
isomerase/autocrine motility factor promotes melanoma cell
migration through ERK activation dependent on autocrine production
of interleukin-8. J Biol Chem. 284:32305–32311. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Kemp DM, Pidich A, Larijani M, Jonas R,
Lash E, Sato T, Terai M, De Pizzol M, Allegretti M, Igoucheva O, et
al: Ladarixin, a dual CXCR1/2 inhibitor, attenuates experimental
melanomas harboring different molecular defects by affecting
malignant cells and tumor microenvironment. Oncotarget.
8:14428–14442. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Brennecke S, Deichmann M, Naeher H and
Kurzen H: Decline in angiogenic factors, such as interleukin-8,
indicates response to chemotherapy of metastatic melanoma. Melanoma
Res. 15:515–522. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Gabellini C, Trisciuoglio D, Desideri M,
Candiloro A, Ragazzoni Y, Orlandi A, Zupi G and Del Bufalo D:
Functional activity of CXCL8 receptors, CXCR1 and CXCR2, on human
malignant melanoma progression. Eur J Cancer. 45:2618–2627. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
76
|
Nürnberg W, Tobias D, Otto F, Henz BM and
Schadendorf D: Expression of interleukin-8 detected by in situ
hybridization correlates with worse prognosis in primary cutaneous
melanoma. J Pathol. 189:546–551. 1999. View Article : Google Scholar
|
|
77
|
Ortega-Bernal D, La Rosa CHG,
Arechaga-Ocampo E, Alvarez-Avitia MA, Moreno NS and Rangel-Escareno
C: A meta-analysis of transcriptome datasets characterizes
malignant transformation from melanocytes and nevi to melanoma.
Oncol Lett. 16:1899–1911. 2018.PubMed/NCBI
|
|
78
|
Adamski V, Mentlein R, Lucius R, Synowitz
M, Held-Feindt J and Hattermann K: The chemokine receptor CXCR6
evokes reverse signaling via the transmembrane chemokine CXCL16.
Int J Mol Sci. 18:182017. View Article : Google Scholar
|
|
79
|
da Silva WC, Oshiro TM, de Sa DC, Franco
DDGS, Festa Neto C and Pontillo A: Genotyping and differential
expression analysis of inflammasome genes in sporadic malignant
melanoma reveal novel contribution of CARD8, IL1B and IL18 in
melanoma susceptibility and progression. Cancer Genet. 209:474–480.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
80
|
Lacina L, Brabek J, Kral V, Kodet O and
Smetana K Jr: Interleukin-6: A molecule with complex biological
impact in cancer. Histol Histopathol. 34:125–136. 2019.
|
|
81
|
Armstrong CA, Murray N, Kennedy M, Koppula
SV, Tara D and Ansel JC: Melanoma-derived interleukin 6 inhibits in
vivo melanoma growth. J Invest Dermatol. 102:278–284. 1994.
View Article : Google Scholar : PubMed/NCBI
|
|
82
|
Lu C and Kerbel RS: Interleukin-6
undergoes transition from paracrine growth inhibitor to autocrine
stimulator during human melanoma progression. J Cell Biol.
120:1281–1288. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
83
|
Elias EG, Hasskamp JH and Sharma BK:
Cytokines and growth factors expressed by human cutaneous melanoma.
Cancers (Basel). 2:794–808. 2010. View Article : Google Scholar
|
|
84
|
Linnskog R, Jönsson G, Axelsson L, Prasad
CP and Andersson T: Interleukin-6 drives melanoma cell motility
through p38a-MAPK-dependent up-regulation of WNT5A expression. Mol
Oncol. 8:1365–1378. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
85
|
Karst AM, Gao K, Nelson CC and Li G:
Nuclear factor kappa B subunit p50 promotes melanoma angiogenesis
by upregulating interleukin-6 expression. Int J Cancer.
124:494–501. 2009. View Article : Google Scholar
|
|
86
|
Nagai H, Oniki S, Fujiwara S, Xu M,
Mizoguchi I, Yoshimoto T and Nishigori C: Antitumor activities of
interleukin-27 on melanoma. Endocr Metab Immune Disord Drug
Targets. 10:41–46. 2010. View Article : Google Scholar
|
|
87
|
Shinozaki Y, Wang S, Miyazaki Y, Miyazaki
K, Yamada H, Yoshikai Y, Hara H and Yoshida H: Tumor-specific
cytotoxic T cell generation and dendritic cell function are
differentially regulated by interleukin 27 during development of
anti-tumor immunity. Int J Cancer. 124:1372–1378. 2009. View Article : Google Scholar
|
|
88
|
Chiba Y, Mizoguchi I, Mitobe K, Higuchi K,
Nagai H, Nishigori C, Mizuguchi J and Yoshimoto T: IL-27 enhances
the expression of TRAIL and TLR3 in human melanomas and inhibits
their tumor growth in cooperation with a TLR3 agonist poly(I:C)
partly in a TRAIL-dependent manner. PLoS One. 8:e761592013.
View Article : Google Scholar : PubMed/NCBI
|
|
89
|
Bisevac JP, Stanojevic I, Mijuskovic Z,
Banovic T, Djukic M and Vojvodic D: High interleukin 27 production
is associated with early clinical stage and localized disease in
patients with melanoma. J Med Biochem. 35:443–450. 2016. View Article : Google Scholar
|
|
90
|
Onoue K, Kusubashi H, Sato Y, Wakitani S
and Takagi M: Quantitative correlation between production rate of
melanoma inhibitory activity and aggrecan gene expression level
during differentiation from mesenchymal stem cells to chondrocytes
and redifferentiation of chondrocytes. J Biosci Bioeng.
111:594–596. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
91
|
Decarlo K, Yang S, Emley A, Wajapeyee N,
Green M and Mahalingam M: Oncogenic BRAF-positive dysplastic nevi
and the tumor suppressor IGFBP7 - challenging the concept of
dysplastic nevi as precursor lesions? Hum Pathol. 41:886–894. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
92
|
Yotsumoto F, Yagi H, Suzuki SO, Oki E,
Tsujioka H, Hachisuga T, Sonoda K, Kawarabayashi T, Mekada E and
Miyamoto S: Validation of HB-EGF and amphiregulin as targets for
human cancer therapy. Biochem Biophys Res Commun. 365:555–561.
2008. View Article : Google Scholar
|
|
93
|
Marchetti D and Nicolson GL: Neurotrophin
stimulation of human melanoma cell invasion: Selected enhancement
of heparanase activity and heparanase degradation of specific
heparan sulfate subpopulations. Adv Enzyme Regul. 37:111–134. 1997.
View Article : Google Scholar : PubMed/NCBI
|
|
94
|
Antunes LCM, Cartell A, de Farias CB,
Bakos RM, Roesler R and Schwartsmann G: Tropomyosin-related kinase
receptor and neurotrophin expression in cutaneous melanoma is
associated with a poor prognosis and decreased survival. Oncology.
97:26–37. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
95
|
Li JR, Wang JQ, Gong Q, Fang RH and Guo
YL: MicroRNA-328 inhibits proliferation of human melanoma cells by
targeting TGFp2. Asian Pac J Cancer Prev. 16:1575–1579. 2015.
View Article : Google Scholar
|
|
96
|
Hutchenreuther J, Vincent K, Norley C,
Racanelli M, Gruber SB, Johnson TM, Fullen DR, Raskin L, Perbal B,
Holdsworth DW, et al: Activation of cancer-associated fibroblasts
is required for tumor neovascularization in a murine model of
melanoma. Matrix Biol. 74:52–61. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
97
|
Straussman R, Morikawa T, Shee K,
Barzily-Rokni M, Qian ZR, Du J, Davis A, Mongare MM, Gould J,
Frederick DT, et al: Tumour micro-environment elicits innate
resistance to RAF inhibitors through HGF secretion. Nature.
487:500–504. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
98
|
Capparelli C, Rosenbaum S, Berger AC and
Aplin AE: Fibroblast-derived neuregulin 1 promotes compensatory
ErbB3 receptor signaling in mutant BRAF melanoma. J Biol Chem.
290:24267–24277. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
99
|
Ruivo CF, Adem B, Silva M and Melo SA: The
biology of cancer exosomes: Insights and new perspectives. Cancer
Res. 77:6480–6488. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
100
|
Hood JL: Melanoma exosome induction of
endothelial cell GM-CSF in pre-metastatic lymph nodes may result in
different M1 and M2 macrophage mediated angiogenic processes. Med
Hypotheses. 94:118–122. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
101
|
Xiao D, Barry S, Kmetz D, Egger M, Pan J,
Rai SN, Qu J, McMasters KM and Hao H: Melanoma cell-derived
exosomes promote epithelial-mesenchymal transition in primary
melanocytes through paracrine/autocrine signaling in the tumor
microenvironment. Cancer Lett. 376:318–327. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
102
|
Gowda R, Robertson BM, Iyer S, Barry J,
Dinavahi SS and Robertson GP: The role of exosomes in metastasis
and progression of melanoma. Cancer Treat Rev. 85:1019752020.
View Article : Google Scholar : PubMed/NCBI
|
|
103
|
Gajos-Michniewicz A and Czyz M: Role of
mirnas in melanoma metastasis. Cancers (Basel). 11:112019.
View Article : Google Scholar
|
|
104
|
Clayton A, Mitchell JP, Court J, Mason MD
and Tabi Z: Human tumor-derived exosomes selectively impair
lymphocyte responses to interleukin-2. Cancer Res. 67:7458–7466.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
105
|
Gerloff D, Lützkendorf J, Moritz RKC,
Wersig T, Mader K, Müller LP and Sunderkotter C: Melanoma-derived
exosomal mir-125b-5p educates tumor associated macrophages (TAMs)
by targeting lysosomal acid lipase A (LIPA). Cancers (Basel).
12:122020. View Article : Google Scholar
|
|
106
|
Bardi GT, Smith MA and Hood JL: Melanoma
exosomes promote mixed M1 and M2 macrophage polarization. Cytokine.
105:63–72. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
107
|
Gershenwald JE and Scolyer RA: Melanoma
Staging: American Joint Committee on Cancer (AJCC) 8th Edition and
Beyond. Ann Surg Oncol. 25:2105–2110. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
108
|
Michielin O, van Akkooi ACJ, Ascierto PA,
Dummer R and Keilholz U; ESMO Guidelines Committee: Electronic
address: simpleclinicalguidelines@esmo.org:
Cutaneous melanoma: ESMO Clinical Practice Guidelines for
diagnosis, treatment and follow-up. Ann Oncol. 30:1884–1901. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
109
|
Riechers A and Bosserhoff AK: Melanoma
inhibitory activity in melanoma diagnostics and therapy - a small
protein is looming large. Exp Dermatol. 23:12–14. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
110
|
Forgber M, Trefzer U, Sterry W and Walden
P: Proteome serological determination of tumor-associated antigens
in melanoma. PLoS One. 4:e51992009. View Article : Google Scholar : PubMed/NCBI
|
|
111
|
Muqaku B, Eisinger M, Meier SM, Tahir A,
Pukrop T, Haferkamp S, Slany A, Reichle A and Gerner C: Multi-omics
analysis of serum samples demonstrates reprogramming of organ
functions via systemic calcium mobilization and platelet activation
in metastatic melanoma. Mol Cell Proteomics. 16:86–99. 2017.
View Article : Google Scholar :
|
|
112
|
Weber JS, Sznol M, Sullivan RJ, Blackmon
S, Boland G, Kluger HM, Halaban R, Bacchiocchi A, Ascierto PA,
Capone M, et al: A serum protein signature associated with outcome
after anti-PD-1 therapy in metastatic melanoma. Cancer Immunol Res.
6:79–86. 2018. View Article : Google Scholar
|
|
113
|
Hoejberg L, Bastholt L, Johansen JS,
Christensen IJ, Gehl J and Schmidt H: Serum interleukin-6 as a
prognostic biomarker in patients with metastatic melanoma. Melanoma
Res. 22:287–293. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
114
|
Correa D, Somoza RA, Lin P, Schiemann WP
and Caplan AI: Mesenchymal stem cells regulate melanoma cancer
cells extravasation to bone and liver at their perivascular niche.
Int J Cancer. 138:417–427. 2016. View Article : Google Scholar :
|
|
115
|
Gandalovicova A, Rosel D, Fernandes M,
Vesely P, Heneberg P, Cermak V, Petruzelka L, Kumar S, Sanz-Moreno
V and Brabek J: Migrastatics-anti-metastatic and anti-invasion
Drugs: Promises and challenges. Trends Cancer. 3:391–406. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
116
|
Herman H, Fazakas C, Hasko J, Molnar K,
Mészaros A, Nyul-Toth A, Szabo G, Erdélyi F, Ardelean A, Hermenean
A, et al: Paracellular and transcellular migration of metastatic
cells through the cerebral endothelium. J Cell Mol Med.
23:2619–2631. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
117
|
Kucera R, Topolcan O, Treskova I,
Kinkorova J, Windrichova J, Fuchsova R, Svobodova S, Treska V,
Babuska V, Novak J, et al: Evaluation of IL-2, IL-6, IL-8 and IL-10
in malignant melanoma diagnostics. Anticancer Res. 35:3537–3541.
2015.PubMed/NCBI
|
|
118
|
Sanmamed MF, Carranza-Rua O, Alfaro C,
Onate C, Martin-Algarra S, Perez G, Landazuri SF, Gonzalez A, Gross
S, Rodriguez I, et al: Serum interleukin-8 reflects tumor burden
and treatment response across malignancies of multiple tissue
origins. Clin Cancer Res. 20:5697–5707. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
119
|
Kucera J, Strnadova K, Dvorankova B,
Lacina L, Krajsova I, Stork J, Kovarova H, Skalmkova HK, Vodicka P,
Motlik J, et al: Serum proteomic analysis of melanoma patients with
immunohistochemical profiling of primary melanomas and cultured
cells: Pilot study. Oncol Rep. 42:1793–1804. 2019.PubMed/NCBI
|
|
120
|
Pelletier F, Bermont L, Puzenat E, Blanc
D, Cairey-Remonnay S, Mougin C, Laurent R, Humbert P and Aubin F:
Circulating vascular endothelial growth factor in cutaneous
malignant melanoma. Br J Dermatol. 152:685–689. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
121
|
Ugurel S, Rappl G, Tilgen W and Reinhold
U: Increased serum concentration of angiogenic factors in malignant
melanoma patients correlates with tumor progression and survival. J
Clin Oncol. 19:577–583. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
122
|
Krasagakis K, Tholke D, Farthmann B,
Eberle J, Mansmann U and Orfanos CE: Elevated plasma levels of
transforming growth factor (TGF)-pi and TGF-|32 in patients with
disseminated malignant melanoma. Br J Cancer. 77:1492–1494. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
123
|
Tuccitto A, Tazzari M, Beretta V, Rini F,
Miranda C, Greco A, Santinami M, Patuzzo R, Vergani B, Villa A, et
al: Immunomodulatory factors control the fate of melanoma tumor
initiating cells. Stem Cells. 34:2449–2460. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
124
|
Tung KH, Ernstoff MS, Allen C and Shu S: A
Review of exosomes and their role in the tumor microenvironment and
host-tumor 'macroenvironment'. J Immunol Sci. 3:4–8. 2019.
View Article : Google Scholar :
|
|
125
|
Shu S, Yang Y, Allen CL, Maguire O,
Minderman H, Sen A, Ciesielski MJ, Collins KA, Bush PJ, Singh P, et
al: Metabolic reprogramming of stromal fibroblasts by melanoma
exosome microRNA favours a pre-metastatic microenvironment. Sci
Rep. 8:129052018. View Article : Google Scholar : PubMed/NCBI
|
|
126
|
Kodet O, Dvorankova B, Bendlova B,
Sykorova V, Krajsova I, Stork J, Kucera J, Szabo P, Strnad H, Kolar
M, et al: Microenvironment driven resistance to B Raf inhibition in
a melanoma patient is accompanied by broad changes of gene
methylation and expression in distal fibroblasts. Int J Mol Med.
41:2687–2703. 2018.PubMed/NCBI
|
|
127
|
Jouve N, Bachelier R, Despoix N, Blin MG,
Matinzadeh MK, Poitevin S, Aurrand-Lions M, Fallague K, Bardin N,
Blot-Chabaud M, et al: CD146 mediates vEGF-induced melanoma cell
extravasation through FAK activation. Int J Cancer. 137:50–60.
2015. View Article : Google Scholar
|
|
128
|
Hamilla SM, Stroka KM and Aranda-Espinoza
H: VE-cadherin-independent cancer cell incorporation into the
vascular endothelium precedes transmigration. PLoS One.
9:e1097482014. View Article : Google Scholar : PubMed/NCBI
|
|
129
|
Kim KJ, Kwon SH, Yun JH, Jeong HS, Kim HR,
Lee EH, Ye SK and Cho CH: STAT3 activation in endothelial cells is
important for tumor metastasis via increased cell adhesion molecule
expression. Oncogene. 36:5445–5459. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
130
|
Borgenström M, Wärri A, Hiilesvuo K,
Käkönen R, Käkönen S, Nissinen L, Pihlavisto M, Marjamäki A,
Vlodavsky I, Naggi A, et al: O-sulfated bacterial polysaccharides
with low anticoagulant activity inhibit metastasis. Semin Thromb
Hemost. 33:547–556. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
131
|
Dange MC, Srinivasan N, More SK, Bane SM,
Upadhya A, Ingle AD, Gude RP, Mukhopadhyaya R and Kalraiya RD:
Galectin-3 expressed on different lung compartments promotes organ
specific metastasis by facilitating arrest, extravasation and organ
colonization via high affinity ligands on melanoma cells. Clin Exp
Metastasis. 31:661–673. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
132
|
Desch A, Strozyk EA, Bauer AT, Huck V,
Niemeyer V, Wieland T and Schneider SW: Highly invasive melanoma
cells activate the vascular endothelium via an MMP-2/integrin
αvβ5-induced secretion of VEGF-A. Am J Pathol. 181:693–705. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
133
|
Tsukamoto H, Fujieda K, Miyashita A,
Fukushima S, Ikeda T, Kubo Y, Senju S, Ihn H, Nishimura Y and
Oshiumi H: Combined blockade of IL6 and PD-1/PD-L1 signaling
abrogates mutual regulation of their immunosuppressive effects in
the tumor microenvironment. Cancer Res. 78:5011–5022. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
134
|
Anker MS, Holcomb R, Muscaritoli M, von
Haehling S, Haverkamp W, Jatoi A, Morley JE, Strasser F, Landmesser
U, Coats AJS, et al: Orphan disease status of cancer cachexia in
the USA and in the European Union: A systematic review. J Cachexia
Sarcopenia Muscle. 10:22–34. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
135
|
Loumaye A and Thissen JP: Biomarkers of
cancer cachexia. Clin Biochem. 50:1281–1288. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
136
|
Weidle UH, Klostermann S, Eggle D and
Krüger A: Interleukin 6/interleukin 6 receptor interaction and its
role as a therapeutic target for treatment of cachexia and cancer.
Cancer Genomics Proteomics. 7:287–302. 2010.PubMed/NCBI
|
|
137
|
Zimmers TA, Fishel ML and Bonetto A: STAT3
in the systemic inflammation of cancer cachexia. Semin Cell Dev
Biol. 54:28–41. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
138
|
Cehreli R, Yavuzsen T, Ates H, Akman T,
Ellidokuz H and Oztop I: Can inflammatory and nutritional serum
markers predict chemotherapy outcomes and survival in advanced
stage nonsmall cell lung cancer patients? BioMed Res Int.
2019:16480722019. View Article : Google Scholar : PubMed/NCBI
|
|
139
|
Johannes CM and Musser ML: Anorexia and
the cancer patient. Vet Clin North Am Small Anim Pract. 49:837–854.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
140
|
Pisetsky DS, Trace SE, Brownley KA, Hamer
RM, Zucker NL, Roux-Lombard P, Dayer JM and Bulik CM: The
expression of cytokines and chemokines in the blood of patients
with severe weight loss from anorexia nervosa: An exploratory
study. Cytokine. 69:110–115. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
141
|
Rochfort KD and Cummins PM: The
blood-brain barrier endothelium. A target for pro-inflammatory
cytokines. Biochem Soc Trans. 43:702–706. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
142
|
Dwarkasing JT, Witkamp RF, Boekschoten MV,
Ter Laak MC, Heins MS and van Norren K: Increased hypothalamic
serotonin turnover in inflammation-induced anorexia. BMC Neurosci.
17:262016. View Article : Google Scholar : PubMed/NCBI
|
|
143
|
Liu WJ, Wang XD, Wu W and Huang X:
Relationship between depression and blood cytokine levels in lung
cancer patients. Med Sci (Paris). 34(Focus issue F1): 113–115.
2018. View Article : Google Scholar
|
|
144
|
Lu YR, Rao YB, Mou YJ, Chen Y, Lou HF,
Zhang Y, Zhang DX, Xie HY, Hu LW and Fang P: High concentrations of
serum interleukin-6 and interleukin-8 in patients with bipolar
disorder. Medicine (Baltimore). 98:e144192019. View Article : Google Scholar
|
|
145
|
Ju RJ, Stehbens SJ and Haass NK: The role
of melanoma cell-stroma interaction in cell motility, invasion, and
metastasis. Front Med (Lausanne). 5:3072018. View Article : Google Scholar
|
|
146
|
Johnson DE, O'Keefe RA and Grandis JR:
Targeting the IL-6/JAK/STAT3 signalling axis in cancer. Nat Rev
Clin Oncol. 15:234–248. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
147
|
Donnelly D III, Aung PP and Jour G: The
'-OMICS' facet of melanoma: Heterogeneity of genomic, proteomic and
metabolomic biomarkers. Semin Cancer Biol. 59:165–174. 2019.
View Article : Google Scholar : PubMed/NCBI
|