Introduction
Glioblastomas (GBM), the most common primary brain
tumors, carry a dismal prognosis despite aggressive surgery,
chemotherapy and radiation therapy (1,2).
Subsets of GBMs, defined by the activation of given signaling
pathways (for instance, c-Met) or distinct driver genetic changes
(for instance, IDH1/2 mutation), however present diverse outcomes
(3,4). Novel therapeutic targets and solid
markers of prognosis are thus of crucial importance in the fight
against these types of cancer.
STAT5 transcription factors are composed of
homologous protein dimers of either STAT5a or STAT5b, encoded by
two different genes, and that show both overlapping and distinct
regulation, transcriptional targets and biological effects
(5). By modulating the expression
of effectors such as Bcl-XL, Aurora A, FAK or VEGF, STAT5 was
reported to contribute to GBM growth, invasion and therapeutic
resistance (6-9). The transcriptional targets of STAT5a
and b present certain overlap but are also differentiated (10). In malignant gliomas, STAT5a was
revealed to be activated downstream of EGFRVIII
(11) and to promote cell
migration, survival and proliferation, notably via induction of the
long non-coding RNA LINC01198 (12,13).
Similarly, STAT5b is highly expressed and predominantly activated
in these tumors (6,7), notably as a result of mir-134
repression (14) and high tyrosine
kinase activity (11,15). A single nucleotide polymorphism of
STAT5b was reported to associate with the risk of GBM (16), and STAT5b can associate with
EGFRVIII in the nucleus in GBM that harbor this
mutation, which activates the transcription of Bcl-XL. STAT5b was
also reported to contribute to GBM cell proliferation (7). Univariable analysis in small series
of glioma patients suggested an inverse correlation between STAT5b
activation and patient survival (6,8,17).
It was also found that epileptogenicity in GBM associates with an
improved survival and with a decreased HIF-STAT5b activation
(18). As a result, STAT5b has
been proposed as a potential target for the treatment of patients
with GBM (16). The nature and the
specificity of the tumorigenic role of STAT5b in GBM however varies
between these reports, and its clinical value as a marker of
survival or a therapeutic target still needs to be confirmed in
large series of patients.
Therefore, the prognostic value of STAT5b expression
in a large institutional cohort of 392 GBM samples and its
association with tumor invasion on the MRIs of 167 of these tumors
were analyzed. The effect of STAT5b inhibition on the
proliferative, therapeutic resistance and migratory capacities of
cultures of human GBM was further evaluated.
Materials and methods
Genetic analysis
The GISTIC 2.0 copy number data and Agilent-based
mRNA expression data of 538 and 552 GBM samples respectively of the
The Cancer Genome Atlas (TCGA) repository were obtained from the
UCSC Cancer Genomics Browser (accessed in September 2015).
Threshold copy number (CN) values were used to perform the
correlations with mRNA expression data using Pearson's correlation
analyses.
Ethics statement
The present study was conducted following review by
the ethics committee of University Medical Center of Utrecht
(Utrecht, The Netherlands) and the institutional review board (IRB;
TC-Bio; approval no. 16-229). According to Dutch regulations, the
need for informed consent was waived for this retrospective
analysis of patient clinical data.
Tissue microarrays (TMAs),
immunohistochemistry (IHC) and MR assessment
Formalin-fixed, paraffin-embedded tumor tissues of a
series of 392 GBM (320 IDH 1 R132 wild-type, 18 IDH1 R132 mutant
and 54 IDH1 status unknown) and 10 non-tumoral epileptic brains
operated between 2005 and 2013 at the University Hospital Center of
Utrecht were included in TMAs. Details on TMA construction and IHC
have been previously described (19). The WHO 2016 classification was used
to define and characterize these tumors. Primary antibodies to
Phospho-STAT5b were used for the immunostaining, as previously
described (20) and revealed using
secondary antibodies and diaminobenzidine (DAB). Briefly, the
4-µm sectioned TMA slides, were deparaffinized in xylene and
rehydrated with graded alcohol solutions. After peroxidase blocking
with hydrogen peroxide (3%), antigen retrieval was achieved by
incubation in citrate buffer (pH 6) for 12 min at 126°C, blocked
for 10 min at room temperature in Dako Protein Block Serum
X090930-2 according to the manufacturer's protocol (Dako; Agilent
Technologies, Inc.), and incubated with 731SP-Stat5b (1/50; cat.
no. ab52211; Abcam) for 1 h at room temperature, and revealed with
the EnvVision + System HRP (Dako; Agilent Technologies, Inc.)
according to the manufacturer's protocol. The processed TMZ slides
were scanned at high resolution on a Hamamatsu NDP scanner and
visualized using the Hamamatsu NDP.view2 software for Mac OS
(Hamamatsu Photonics K.K.). Protein expression evaluation was
blinded to the clinical data and scored as negative (0% of cells
with Phospho-STAT5b staining) or positive (>0% of cells with
Phospho-STAT5b staining; IHC score 1 with 0-25% of cells with
Phospho-STAT5b staining, score 2 with 25-50%, score 3 with 50-75%
and score 4 with 75-100%). Clinical data of the patients were
retrieved form the charts of the patients, following ethical and
IRB approval at the UMC Utrecht (approval nos. 16-229 and 16-342)
and are provided in Table SI.
Preoperative MRI scans available for 167 of the first 196 patients
of our institutional cohort were analyzed by a trained radiologist
for the following signs of tumor dissemination: the presence (or
not) of tumor islets at a distance of the tumor mass, a ratio >2
between the maximal diameters of the T2-weighted extent of the
tumor and of the contrast-enhancing T1-weighted tumor area, and the
invasion of the corpus callosum by the enhancing component of the
tumor.
Cell cultures, reagents and small
interfering (si)RNA
The genetic profile of human LN18 GBM (cat. no.
CRL-2610; ATCC) and U87 malignant glioma cells of unknown origin
(cat. no. HTB-14; ATCC) was verified using CGH (Affymetrix SNP6.0
arrays) and TP53 sequencing (ion torrent). GM2 and GM3 primary GBM
cells were derived from fresh samples of human GBM and cultured as
previously described (20). They
were characterized using GFAP IHC, TP53, IDH1/2 (both wild-type),
and EGFRVIII (both negative) mRNA sequencing and by CGH
analysis, and maintained at low passages. U87VIII cells
were kindly provided by Dr M. Broekman (University Medical Center
of Utrecht) and their expression of EGFRVIII was
confirmed by next generation sequencing. Cells were cultured in 5%
CO2 in DMEM (Thermo Fisher Scientific, Inc.)
supplemented with 10% FBS (Gibco; Thermo Fisher Scientific, Inc.)
and 1% of 5 mg/ml penicillin-streptomycin (Gibco; Thermo Fisher
Scientific, Inc.) solution at 37°C, and maintained at low
passages.
For siRNA experiments, 70% confluent cultured cells
were transiently transfected with 25 nM of Control pool
non-targeting #1 (D-001810-10-05) or SMARTpool human STAT5b siRNA
(M-010539-02) from Dharmacon using the DharmaFECT transfection
reagent (Thermo Fisher Scientific, Inc.; DharmaFECT Transfection
Reagents) according to the manufacturer's protocol at 37°C.
Transfection time was 48 h, after which the cells were used for
subsequent experimentations.
Western blot analysis
Whole-cell lysate were obtained using lysis SDS 1%
buffer containing protease and phosphatase inhibitors. Protein
concentration was determined using BCA method (Pierce kit; Thermo
Fisher Scientific, Inc.), and 25 µg of protein were loaded
per lane. Western blot analysis was performed in polyacrylamide 10%
gels and run for 1 h and 30 min at 100 V, then transferred to PVDF
membrane (Roche Diagnostics) for 1 h at 100 V. Blocking was
performed for 1 h at room temperature, in the same buffers used for
the incubation of the respective antibodies. All primary antibodies
were incubated overnight at 4°C and the dilution recommended by the
manufacturer was used; STAT5b (Abcam; cat. no. ab194380; 1:5,000 in
4% BSA (VWR International, LLC), 1X TBS, 0.1% Tween® 20
buffer), GAPDH (Sigma-Aldrich; Merck KGaA; cat. no. PLA0125;
1:10,000 in 4% non-fat dry milk, 1X TBS, 0.1% Tween®
20), p27 (Cell Signaling Technology, Inc.; cat. no. 3688; 1:1,000
in 4% BSA 1X TBS, 0.1% Tween® 20 buffer), Cyclin D1
(Cell Signaling Technology, Inc.; cat. no. 2922; 1:1,000 in 4% BSA
1X TBS, 0.1% Tween®-20 buffer), BCL-XL (Cell Signaling
Technology, Inc.; cat. no. 2764; 1:1,000 in 4% BSA 1X TBS, 0.1%
Tween®-20 buffer) and PD-L1 (Cell Signaling Technology,
Inc.; cat. no. 13684; 1:1,000 in 4% BSA 1X TBS, 0.1%
Tween®-20 buffer). Appropriate HRP-linked secondary
antibody was used (Cell Signaling Technology, Inc.; cat. no. 7074;
1:3,000) for incubation at room temperature during 2 h with gentle
shaking. For detection, enhanced chemiluminescence method was used.
Clarity Western ECL Blotting Substrate (Bio-Rad Laboratories, Inc.)
was used coupled with film-based imaging following the
manufacturer's protocol.
Cell survival assays
After 24 h of transfection, 2,500 cells were seeded
in 96-well plate, then let to adhere overnight before to support
Temozolomide (TMZ; cat. no. T2577; Sigma-Aldrich; Merck KGaA)
treatment or gamma-radiation and later MTS assay (One solution cell
proliferation assay; cat. no. G3582; Promega Corporation) as
recommended by the manufacturer, and the absorbance was measured at
490 nm.
Clonogenic assays
After 48 h of siRNA transfection, 500 cells were
seeded in six-well plate followed or not with gamma-radiation, then
left to grow for 7 days and then fixed in 4% paraformaldehyde (45
min at room temperature) and stained with crystal violet (5 mg/ml)
for 10 min at room temperature before counting on a light
microscope (Olympus Corporation).
Migration/Invasion assays
For Boyden chamber assays, 48 h post-siRNA
transfection, a calculated number of cells (50,000 for U87, GM2 and
GM3; 25,000 for U87VIII and LN18) in serum-free medium with 0,1%
BSA (VWR International, LLC) were seeded into the upper chamber of
Transwell inserts (8 µm) coated with collagen type I (50
µg/ml) for migration and with Matrigel (500 µg/ml;
precoating for 30 min at 37°C) for invasion, whereas medium with
1,5% FBS and 1% BSA was applied in the lower chamber as
chemo-attractant. After 6 h of migration or 24 h of invasion, cells
were fixed with 4% paraformaldehyde and stained with 0.4% crystal
violet (ambiant temperature, 10 min), scanned on a Hamamatsu NDP
scanner, and counted. For wound healing (scratch) assays, 48 h
post-siRNA transfection cells were cultured until confluence and
treated for 1 h with mitomycin C (Sigma-Aldrich; Merck KGaA) and
then wounded using a 100-µl pipette tip. Images of the
migration distance were captured on a phase contrast Leica
microscope and measured at zero time and after 1, 4, 6, 20 and 24 h
and expressed in percentages of the original gap. For
quantification, the margins were plotted and 3 measurements were
made per scratch, and averaged to calculate the healing percentage.
Each condition was performed in quadruplicate.
Statistical analysis
Statistical analysis was performed using the Prism
5.0 (GraphPad Software, Inc.) and the SPSS 24 (IBM Corp.) software.
Kaplan-Meier survival estimates were obtained and multivariable Cox
regression analysis was performed taking the age, KPS (> or
<70), tumor volume (in cubic cm, measured with
Osirix® and based on the contrast-enhancing T1-weighted
tumor boundaries), type of surgery (biopsy vs. debulking) and IDH
R132H status into account to assess correlation with overall
survival. Survival data were censored at 1,000 days in order to
comply with the condition of proportional hazard for the survival
analyses (19). Multiple t-tests
and two-way ANOVA, with Dunnett's multiple comparisons post hoc
tests were performed as appropriate for non-survival data.
Correlations were assessed using Pearson's test. The association of
low or high Stat5b activation with MRI criteria of invasion was
assessed by Chi-square and Kruskall Wallis tests. Results are
expressed as the mean ± SD and a two-sided P<0.05 was considered
to indicate a statistically significant difference.
Results
STAT5b activation and GBM patient
survival
Agilent-based STAT5b mRNA expression values and gene
copy number (CN) were obtained for 552 patients of the TCGA
repository of GBM. There was a weak correlation between the mRNA
expression of STAT5b and its copy number (Pearson's correlation
0.091, P=0.033), but this CN did not correlate with patient
survival (data not shown). Similarly, the mRNA expression of STAT5b
mRNA did not correlate with patient survival in a Cox survival
model taking the Karnofsky performance score (KPS) and patient age
into account (P=0.696). TMAs were obtained for a series of 392
patients treated at our neuro-oncological center and for 10
non-tumor brain tissue samples obtained from temporal lobe epilepsy
surgery.
IHC showed a high level of nuclear expression of
Phospho-STAT5b (p-STAT5b) in a majority of the samples, including
in the non-tumor brain samples. In these non-tumor samples, 75% of
non-neuronal cells presented a positive staining of the nucleus for
STAT5b, i.e., an immunochemistry score of 3 (Fig. S1). In the GBM patients, there was
an inverse correlation between the nuclear staining score for
nuclear p-STAT5b (taken as a continuous variable) and overall
survival in a multivariable Cox model taking the Age, KPS, tumor
volume, type of surgery and IDH1R132 mutational status into account
[hazard ratio (HR), 1.22; 95% confidence interval (CI),
1.111-1.346; P<0.001]. Also significant in this analysis were
the patient age and KPS (Table
SII for). This multivariable analysis remained even significant
when the patients were dichotomized between two groups based on a
practical threshold of 3, corresponding to that of 'non-tumor'
p-STAT5b nuclear staining instead of a continuous variable (HR
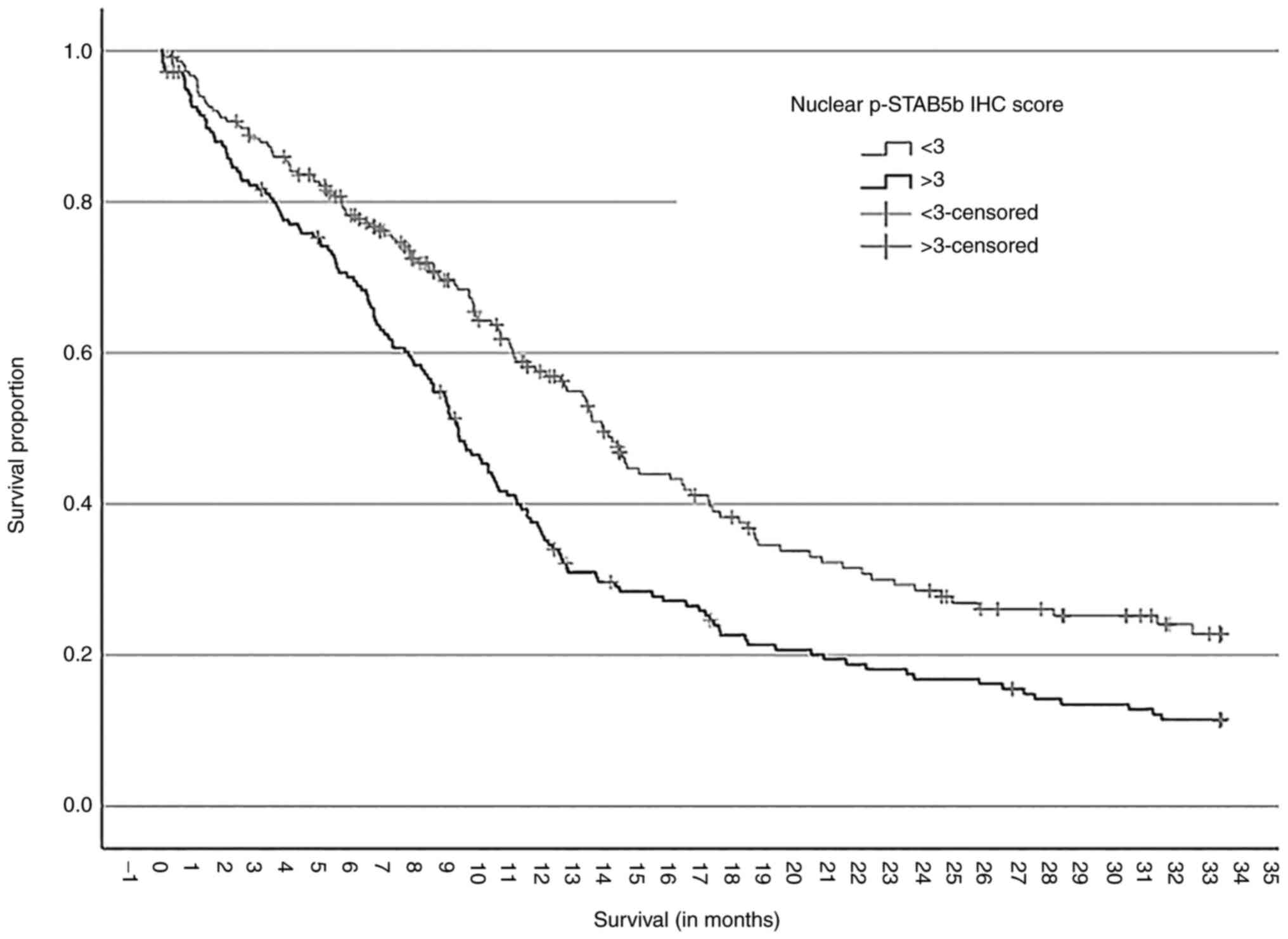
1.664; 95% CI 1.290-2.150; P<0.001; Table I). Kaplan Meier survival estimates
based on this threshold showed a median survival of 13.5 months in
the low STAT5b activation group vs. only 9.3 months in the high
STAT5b activation group of patients (P<0.001, Log Rank test,
Fig. 1).
 | Table IMultivariate Cox regression analysis
of the overall survival of 392 glioblastomapatients form our
institutional cohort. |
Table I
Multivariate Cox regression analysis
of the overall survival of 392 glioblastomapatients form our
institutional cohort.
| Variables in the
Cox model | P-value | HR | 95% CI for HR
|
|---|
| Lower | Upper |
|---|
| Nuclear p-STAT5b
IHC score (>3 vs. <3) | <0.001 | 1.664 | 1.290 | 2.150 |
| Age at
diagnosis | <0.001 | 1.037 | 1.024 | 1.05 |
| KPS (<70 vs.
>70) | <0.001 | 2.387 | 1.811 | 3.147 |
| Type of surgery
(Biopsy vs. debulking) | 0.191 | 1.349 | 0.862 | 2.111 |
| IDH1 R132H gene
(wild-type vs. mutant) | 0.06 | 2.207 | 0.967 | 5.037 |
STAT5b and GBM cell proliferation
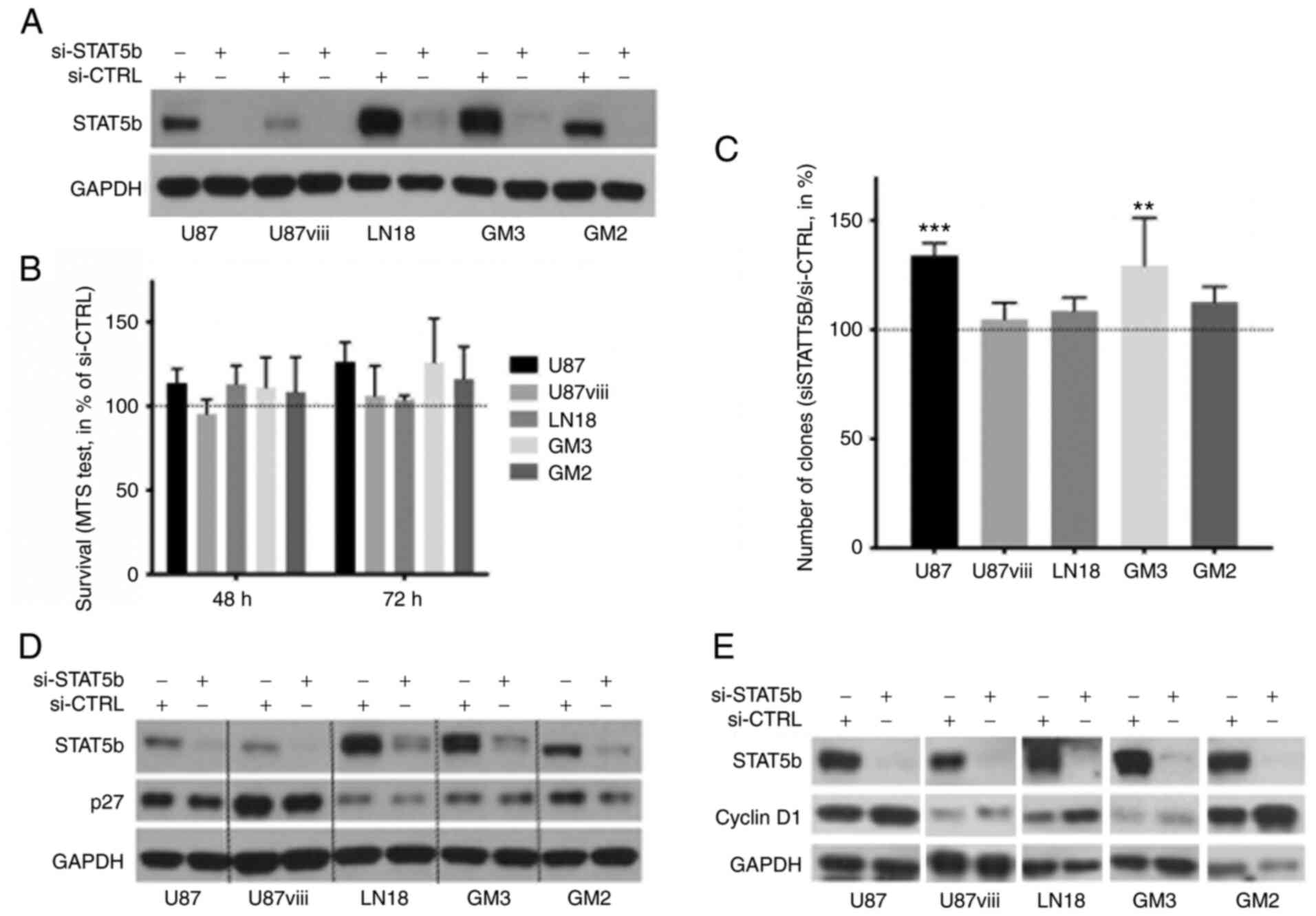
Transfection of human GBM cell cultures using a
SMARTpool® of human STAT5b siRNA resulted in a
significant and lasting STAT5b protein knockdown (80% protein
reduction minimum) within 24 h (Fig.
2A). This depletion of STAT5b did not prove cytotoxic to any of
the GBM cells as measured by a MTS test for 72 h (NS, ANOVA, n=3,
Fig. 2B). In clonogenic assays,
STAT5b knockdown even induced a slight but significant increase in
colony (minimum 20 cells) formation in U87 and GM3 cells
(P<0.01; n=3, Fig. 2C), and did
not alter that of U87VIII, LN18 and GM2 cells. STAT5b
depletion also resulted in a reproducible decrease of the cell
cycle inhibitor p27kip1 (Fig. 2D) and a reproducible increase of
Cyclin D1 in all cell types (Fig.
2E). In search for additional effects, STAT5b inhibition did
not affect expression the expression of Bcl-XL (Fig. S4), on GBM cells, and increased the
expression of the immune checkpoint ligand PD-L1 (Fig. S5) in these cells.
STAT5b and chemo/radio-sensitivity of GBM
cells
Given the inverse correlation between p-STAT5b
nuclear expression and survival in our cohort of GBM patients and a
previous study that STAT5b contributes to chemoresistance of of GBM
to cisplatin (12), it was
investigated whether this transcription factor would also
contribute to the resistance of these tumors to their conventional
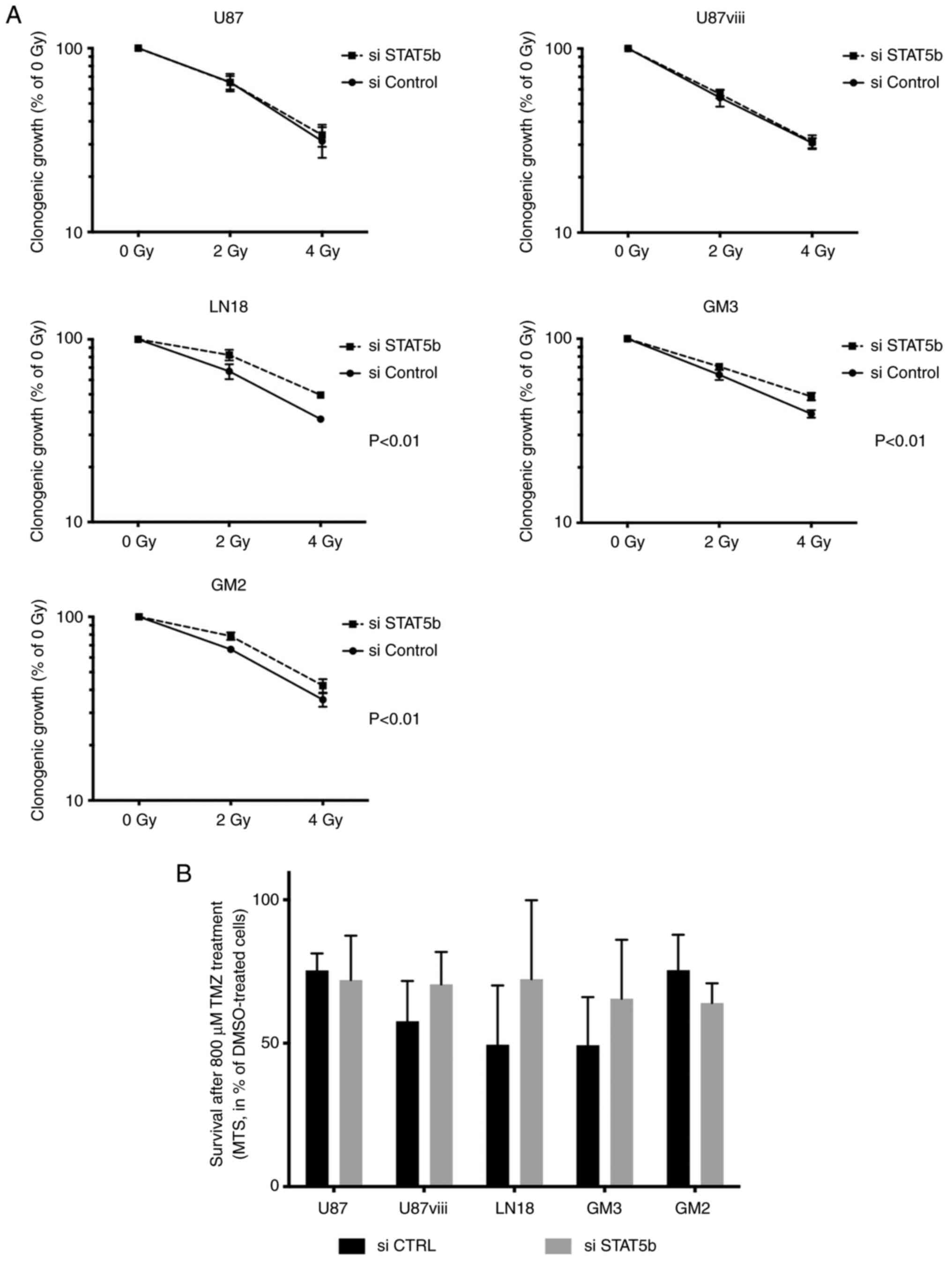
treatments, namely ionizing radiation and TMZ chemotherapy. In
clonogenic assays performed with increasing doses of radiation
(0-2-4 Gy), STAT5b knockdown did not affect the radiation
sensitivity of U87 and U87viii cells. STAT5b depletion even
slightly but significantly protected LN18, GM3 and GM2 cells
against radiation toxicity (P<0,01, two-ways ANOVA, n=4,
Fig. 3A). The sensitivity of GBM
cells to TMZ treatment (800 µM) was not significantly
affected by STAT5b depletion (Fig.
3B).
STAT5b and GBM dissemination
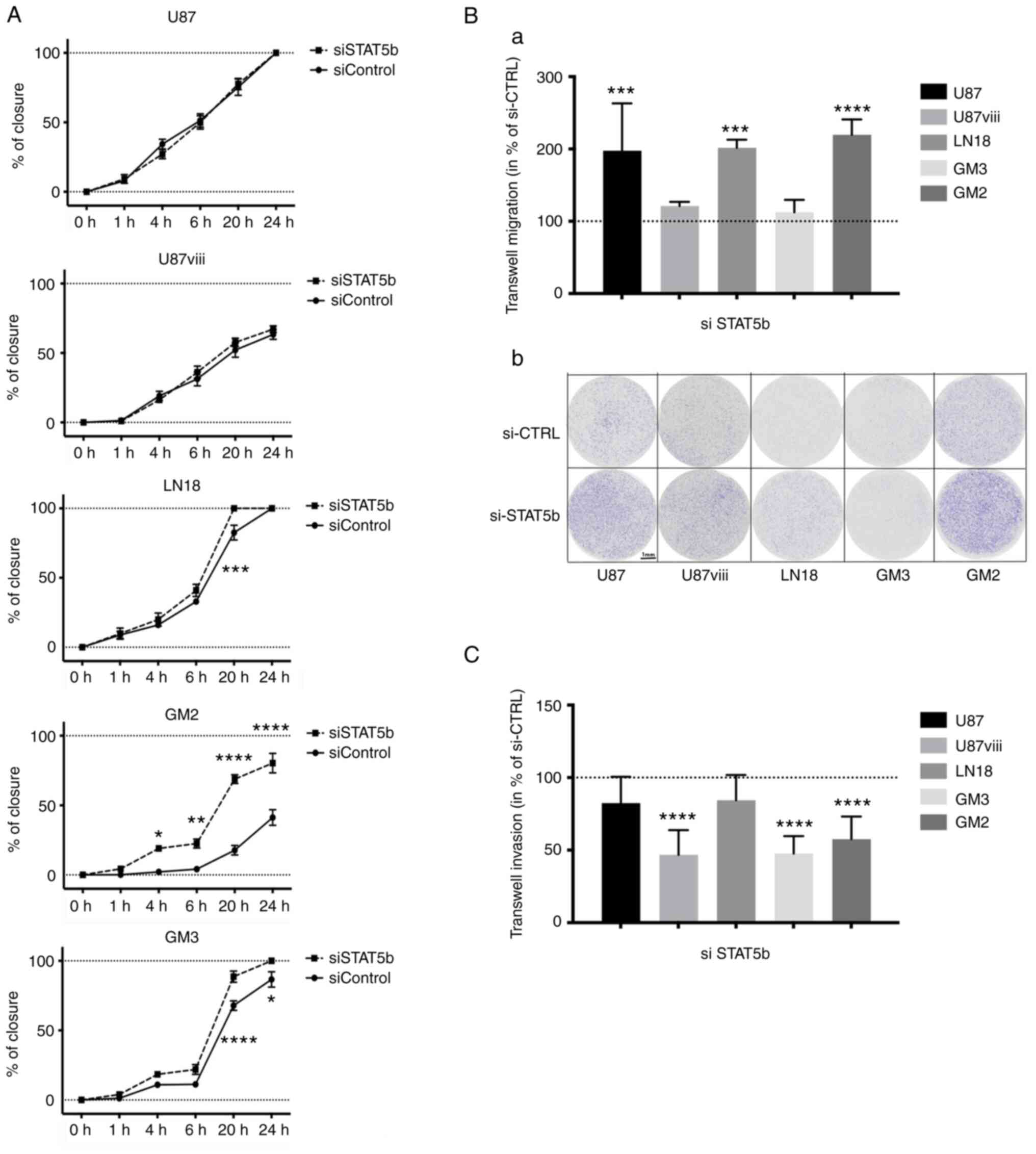
In scratch assays, the closure kinetic of the
monolayer gap was assessed for 24 h, and was similar between
siSTAT5b and siControl in both U87 and U87VIII cells.
The healing was however slightly faster in siSTAT5b-than in
siControl-treated LN18 cells, lasting 20 vs. 24 h. Similarly,
siSTAT5b-treated GM3 and GM2 cells healed significantly faster than
their siControl counterparts (Figs.
4A and S2). Following STAT5b
knockdown, the migration of U87, LN18 and GM2 cells through
collagen-coated membranes of Boyden chambers increased
significantly as well (by 197,7±65,4%, 201,6±11,2% and 219,7±21%
respectively; P<0,001; n=3), while that of U87VIII
and GM3 remained unaffected [Fig. 4B
(a) and (b)].
When the Boyden membranes were coated with Matrigel
however, the invasion of U87VIII, GM3 and GM2 cells
decreased by 53,2±17,1%, 52,2±11,9% and 43,5±15,6% upon STAT5b
depletion (P<0,0001; n=4 for GM3 and n=5 for U87VIII
and GM2; Figs. 4C and S3), while that of U87 and LN18 did not
change significantly.
Finally, in order to assess the resulting clinical
effect of these diverging effects of STAT5b on migration and
invasion of GBM, the available T1-weighted, gadolinium-enhanced MRI
images of the first half of our patients (available in 167/196
patients) were analyzed for signs of tumor dissemination. The
presence (or not) of tumor islets at a distance of the tumor mass,
a ratio >2 between the maximal diameters of the T2-weighted
extent of the tumor and of the contrast-enhancing T1-weighted tumor
area, or invasion of the corpus callosum by the enhancing component
of the tumor were assessed. There was neither any difference
between the level of nuclear STAT5b staining between tumors
presenting either of these dissemination features or not (N.S.,
Kruskall-Wallis non-parametric test), nor was there any association
between a high STAT5b IHC score (>3) and these parameters (N.S.,
Chi-square test).
Discussion
STAT5a and STAT5b present overlapping and distinct
regulatory pathways and transcriptional targets (5,8), and
while STAT5a has been shown to favor the proliferation of malignant
gliomas, STAT5b appears to be predominantly activated in GBM
(7,14) and has been proposed as a
therapeutic target against these tumors (16).
As previously described, indeed (6,7,17),
GBM from our cohort revealed a high level of STAT5b activation.
This activation was often higher than that of the glial cells of
non-tumoral brain samples, and is inversely correlated with patient
survival. The present Kaplan-Meier estimates demonstrated that
tumors with a high level of nuclear p-STAT5b staining had a median
overall survival more than 4 months shorter than those with a low
STAT5b activation level. This prognostic association remains highly
significant in multivariable analysis independent of the KPS, age,
tumor volume, type of surgery or IDH1 R132H mutation. The present
findings confirmed and extented those of previous survival analyses
that were performed in monovariable fashion and on much smaller
cohorts of patients (6,17). It was also observed that neither
the copy number nor the mRNA expression of STAT5b correlated with
survival in GBM patients, suggesting that it is truly the nuclear
activation of STAT5b, rather than its mere expression, that
correlates with the prognosis of patients with a GBM.
Despite its inverse association with patient
survival, STAT5b activation did not support GBM cell proliferation.
On the contrary in fact, the inhibition of STAT5b reduced the
expression of p27 and increased that of Cyclin D1 in our panel of
cell lines and primary cultures of GBM, and even slightly increased
the clonogenic potential of U87 and GM3 cells. This contrasts with
the proliferative actions of STAT5a (12,13),
and likely underscores the specificity of these two STAT family
members (10). Notably, Liang
et al (7) observed that a
less complete depletion of STAT5b than in our experiments decreased
the proliferation of some GBM cell lines. This suggested that the
effects of STAT5b could be concentration-dependent and non-linear,
or could depend on a more complex balance with other transcription
factors. It was also noted that the pro-clonogenic effect of STAT5b
knockdown observed in U87 cells, was absent in U87VIII
cells. EGFRvIII has been identified to form nuclear
STAT5b-EGFRvIII complexes in GBM (8,21),
which activate BCLXL and favor cell survival (8,22),
and could have contributed this difference. However, no alteration
of Bcl-XL expression was observed in our experimental conditions
following STAT5b knockdown, and other cell-specific mechanisms must
be at play. Altogether, and most importantly, the effects of STAT5b
on the proliferation of malignant gliomas remain variable from
tumor to tumor, and this should caution the targeting of STAT5b for
therapeutic goals.
In line with a previous study (9), it was observed that the invasion of
several GBM cell cultures decreased upon STAT5b inhibition.
However, and in contrast with a previous study (7), STAT5b inhibition increased the
migration of most of our GBM cells in wound healing and Transwell
migration experiments. Differential effects of a given signaling
pathways are uncommon, but have been observed previously, notably
for the focal adhesion kinase FAK, a target of STAT5 signaling
(7,23). As a likely result of these opposite
effects of STAT5b on invasion and migration, no correlation was
found between STAT5b activation and tumor invasion on the MRI scans
of our patients.
STAT5b has also been revealed to protect GBM cells
against the cytotoxic effects of DNA-damaging agents such as
cisplatin (8). However, no
sensitization of GBM cells to TMZ or ionizing radiations was
observed following STAT5b inhibition, and even a slight protection
against these conventional anti-GBM cytotoxic agents was identified
in some of the cultures of the present study. Finally, it was
revealed that STAT5b inhibition also increased the expression of
the immune checkpoint ligand PD-L1 on GBM cells, further casting
doubts on the potential of STAT5b as a potential therapeutic target
in GBM.
Collectively, the present findings clearly defined
the potential of activated nuclear p-STAT5b as a prognostic marker
of patient survival in GBM, independent on IDH1 R132H status and
other classical predictors of longevity. Given its lack of
significant proper oncogenic role however, our results do not
support anti-STAT5b strategies as a means to treat GBM. Rather,
nuclear p-STAT5b appears to be a surrogate marker of the activation
of true oncogenic pathways, possibly HIF-1α (18) or tyrosine kinase receptors, which
are frequently hyper-activated in GBM (8,11,24)
and can, besides the JAK/STAT cascade, activate known oncogenic
pathways such as the ERK/MAP kinases or the NF-kappaB pathways
(3,14,20,25).
Supplementary Data
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
Conceptualized the study. ND and PAR wrote the
prepared and wrote the draft. ND and PAR revised and edited the
manuscript. ND, TS, LS, KT, SB, WS and PAR performed data
acquisition, experiments and analysis. PAR and VB supervised the
study. PR, VB and TS acquired funding. All authors read and
approved the final manuscript. PAR, VB and ND confirm the
authenticity of all the raw data.
Ethics approval and consent to
participate
The present study was conducted following review by
the local ethical committee and the institutional review board
(TC-Bio; approval nos. 16-229 and 16-342). According to Dutch
regulations, the need for informed consent was waived for this
retrospective analysis of patient clinical data.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Acknowledgments
The authors would like to thank Dr J. Hendrikse (UMC
Utrecht) for his help assessing the MRI criteria of invasion.
Funding
The present study was supported by Televie grants (grant nos.
1.7.247.330.12 and 7.4567.15) from the FNRS of Belgium, the Belgian
National Cancer Plan (grant no. 20-044) and the T&P Bohnenn
Fund for Neuro-Oncology research.
References
|
1
|
Stupp R, Mason WP, van den Bent MJ, Weller
M, Fisher B, Taphoorn MJ, Belanger K, Brandes AA, Marosi C, Bogdahn
U, et al: Radiotherapy plus concomitant and adjuvant temozolomide
for glioblastoma. N Engl J Med. 352:987–996. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Stupp R, Taillibert S, Kanner AA, Kesari
S, Steinberg DM, Toms SA, Taylor LP, Lieberman F, Silvani A, Fink
KL, et al: Maintenance therapy with tumor-treating fields plus
temozolomide vs temozolomide alone for glioblastoma: A randomized
clinical trial. JAMA. 314:2535–2543. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Bell EH, Pugh SL, McElroy JP, Gilbert MR,
Mehta M, Klimowicz AC, Magliocco A, Bredel M, Robe P, Grosu AL, et
al: Molecular-based recursive partitioning analysis model for
glioblastoma in the temozolomide era: A correlative analysis based
on NRG oncology RTOG 0525. JAMA Oncol. 3:784–792. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Eckel-Passow JE, Lachance DH, Molinaro AM,
Walsh KM, Decker PA, Sicotte H, Pekmezci M, Rice T, Kosel ML,
Smirnov IV, et al: Glioma groups based on 1p/19q, IDH, and TERT
promoter mutations in tumors. N Engl J Med. 372:2499–2508. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Maurer B, Kollmann S, Pickem J,
Hoelbl-Kovacic A and Sexl V: STAT5A and STAT5B-twins with different
personalities in hematopoiesis and leukemia. Cancers (Basel).
11:17262019. View Article : Google Scholar
|
|
6
|
Kuo YH, Chen YT, Tsai HP, Chai CY and Kwan
AL: Nucleophosmin overexpression is associated with poor survival
in astrocytoma. APMIS. 123:515–522. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Liang QC, Xiong H, Zhao ZW, Jia D, Li WX,
Qin HZ, Deng JP, Gao L, Zhang H and Gao GD: Inhibition of
transcription factor STAT5b suppresses proliferation, induces G1
cell cycle arrest and reduces tumor cell invasion in human
glioblastoma multiforme cells. Cancer Lett. 273:164–171. 2009.
View Article : Google Scholar
|
|
8
|
Latha K, Li M, Chumbalkar V, Gururaj A,
Hwang Y, Dakeng S, Sawaya R, Aldape K, Cavenee WK, Bogler O and
Furnari FB: Nuclear EGFRvIII-STAT5b complex contributes to
glioblastoma cell survival by direct activation of the Bcl-XL
promoter. Int J Cancer. 132:509–520. 2013. View Article : Google Scholar :
|
|
9
|
Cao S, Wang C, Zheng Q, Qiao Y, Xu K,
Jiang T and Wu A: STAT5 regulates glioma cell invasion by pathways
dependent and independent of STAT5 DNA binding. Neurosci Lett.
487:228–233. 2011. View Article : Google Scholar
|
|
10
|
Lin JX and Leonard WJ: The role of Stat5a
and Stat5b in signaling by IL-2 family cytokines. Oncogene.
19:2566–2576. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Chumbalkar V, Latha K, Hwang Y, Maywald R,
Hawley L, Sawaya R, Diao L, Baggerly K, Cavenee WK, Furnari FB and
Bogler O: Analysis of phosphotyrosine signaling in glioblastoma
identifies STAT5 as a novel downstream target of ΔEGFR. J Proteome
Res. 10:1343–1352. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Roos A, Dhruv HD, Peng S, Tuncali S,
Pineda M, Millard N, Mayo Z, Eschbacher JM, Loftus JC, Winkles JA
and Tran NL: EGFRvIII-Stat5 signaling enhances glioblastoma cell
migration and survival. Mol Cancer Res. 16:1185–1195. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Tan C, Dai Y, Liu X, Zhao G, Wang W, Li J
and Qi L: STAT5A induced LINC01198 promotes proliferation of glioma
cells through stabilizing DGCR8. Aging (Albany NY). 12:5675–5692.
2020. View Article : Google Scholar
|
|
14
|
Zhang Y, Kim J, Mueller AC, Dey B, Yang Y,
Lee DH, Hachmann J, Finderle S, Park DM, Christensen J, et al:
Multiple receptor tyrosine kinases converge on microRNA-134 to
control KRAS, STAT5B, and glioblastoma. Cell Death Differ.
21:720–734. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Sawada T, Arai D, Jing X, Miyajima M,
Frank SJ and Sakaguchi K: Molecular interactions of EphA4, growth
hormone receptor, Janus kinase 2, and signal transducer and
activator of transcription 5B. PLoS One. 12:e01807852017.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Liu YL, Liu PF, Liu HE, Ma LX and Li G:
Association between STAT5 polymorphisms and glioblastoma risk in
Han Chinese population. Pathol Res Pract. 210:582–585. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Televantou D, Karkavelas G, Hytiroglou P,
Lampaki S, Iliadis G, Selviaridis P, Polyzoidis KS, Fountzilas G
and Kotoula V: DARPP32, STAT5 and STAT3 mRNA expression ratios in
glioblastomas are associated with patient outcome. Pathol Oncol
Res. 19:329–343. 2013. View Article : Google Scholar
|
|
18
|
Berendsen S, Spliet WGM, Geurts M, Van
Hecke W, Seute T, Snijders TJ, Bours V, Bell EH, Chakravarti A and
Robe PA: Epilepsy associates with decreased HIF-1α/STAT5b signaling
in glioblastoma. Cancers (Basel). 11:412019. View Article : Google Scholar
|
|
19
|
Berendsen S, Varkila M, Kroonen J, Seute
T, Snijders TJ, Kauw F, Spliet WG, Willems M, Poulet C, Broekman
ML, et al: Prognostic relevance of epilepsy at presentation in
glioblastoma patients. Neuro Oncol. 18:700–706. 2016. View Article : Google Scholar :
|
|
20
|
Robe PA, Bentires-Alj M, Bonif M, Rogister
B, Deprez M, Haddada H, Khac MT, Jolois O, Erkmen K, Merville MP,
et al: In vitro and in vivo activity of the nuclear factor-kappaB
inhibitor sulfasalazine in human glioblastomas. Clin Cancer Res.
10:5595–5603. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Fan QW, Cheng CK, Gustafson WC, Charron E,
Zipper P, Wong RA, Chen J, Lau J, Knobbe-Thomsen C, Weller M, et
al: EGFR phosphorylates tumor-derived EGFRvIII driving STAT3/5 and
progression in glioblastoma. Cancer Cell. 24:438–449. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Yang C, Huang W, Yan L, Wang Y, Wang W,
Liu D and Zuo X: Downregulation of the expression of B-cell
lymphoma-extra large by RNA interference induces apoptosis and
enhances the radiosensitivity of nonsmall cell lung cancer cells.
Mol Med Rep. 12:449–455. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Hsia DA, Mitra SK, Hauck CR, Streblow DN,
Nelson JA, Ilic D, Huang S, Li E, Nemerow GR, Leng J, et al:
Differential regulation of cell motility and invasion by FAK. J
Cell Biol. 160:753–767. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Gressot LV, Doucette TA, Yang Y, Fuller
GN, Heimberger AB, Bögler O, Rao A, Latha K and Rao G: Signal
transducer and activator of transcription 5b drives malignant
progression in a PDGFB-dependent proneural glioma model by
suppressing apoptosis. Int J Cancer. 136:2047–2054. 2015.
View Article : Google Scholar :
|
|
25
|
Bredel M, Scholtens DM, Yadav AK, Alvarez
AA, Renfrow JJ, Chandler JP, Yu IL, Carro MS, Dai F, Tagge MJ, et
al: NFKBIA deletion in glioblastomas. N Engl J Med. 364:627–637.
2011. View Article : Google Scholar
|