Cancer endangers the health of the population
worldwide and its incidence is increasing annually with the
accumulation of risk factors, such as the aging of the population,
accelerated industrialization and poor lifestyle habits (1). Although the survival rate of cancer
patients has improved due to treatments such as surgery, targeted
therapy, radiotherapy and immunotherapy, cancer remains a major
challenge due to its highly aggressive and drug-resistant
nature.
The long-recognized modes of cell death comprise
apoptosis and necrosis. Apoptosis is characterized by cell
shrinkage and the formation of apoptotic bodies. The surrounding
phagocytic cells engulf these bodies without causing an
inflammatory response. Necrosis was previously thought to be an
unregulated and passive death process; however, with further
research, it has now been indicated that certain forms of necrosis
may be controlled and are called programmed necrosis (2), of which pyroptosis is one of the main
forms. Pyroptosis may intensify the inflammatory response by
activating the inflammasome and inflammatory cytokines in the
extracellular space in addition to rupturing cells and releasing
inflammatory cytokines that promote inflammation.
The gasdermin (GSDM) family of proteins are key
effector molecules that mediate the onset of pyroptosis and include
GSDMA-GSDME and Pejvakin (PJVK), also known as deafness, autosomal
recessive 59 (DFNB59). The GSDM family of proteins have similar
structures and are characterized by a conserved domain called the
GSDM-N domain (GSDM-N), which is responsible for executing the
process of pyroptosis. The GSDM-N domain consists of a six-stranded
β-sheet flanked by α-helices and is crucial for the pore-forming
activity of the protein. In addition to the GSDM-N domain, the GSDM
family proteins possess a C-terminal domain known as GSDM-C, which
aids in membrane binding (3). Most
of the GSDM family proteins exert their effects via inducing
pyroptosis, and all of them influence the incidence, development
and prognosis of multiple cancer types. The purpose of the present
study was to summarize the molecular mechanisms of GSDM-mediated
pyroptosis and the latest progress in its application in cancer
research, and to provide new strategies for cancer prevention and
treatment.
The gene encoding GSDMA is located on human
chromosome region 17q21 and mouse chromosome region 11. There is
only one copy of the gene in humans, whereas mice have three copies
of the gene (GSDMA1-3) (4). GSDMA
expression patterns differ between humans and mice. Human GSDMA is
mainly expressed in epithelial cells of the skin, tongue, mammary
glands, bladder, umbilical cord and gastrointestinal tract, such as
the esophagus and stomach (5).
Furthermore, GSDMA was detected in T lymphocytes (6). Mouse GSDMA1-3 is expressed in
distinct tissues. GSDMA1 is mainly expressed in the suprabasal
epidermis, cornea, hair follicles and forestomach (5); GSDMA2 is expressed in the stomach;
and GSDMA3 is mainly detected in the sebaceous glands of the skin
(7). Overexpression of the
N-terminal domain of GSDMA or GSDMA3 induces pore formation in the
plasma membranes and leads to significant pyroptosis (3,8).
However, the inflammatory caspase responsible for the cleavage of
GSDMA remains to be determined.
Limited evidence has demonstrated an association
between GSDMA and cancer. Saeki et al (9) detected the expression level in a
pan-cancer analysis and observed that GSDMA was amplified in 4/4
(100%) breast cancer cell lines and 2/8 (25%) gastric HER2-positive
cell lines. However, northern blot analysis showed no GSDMA
expression in any of the 24 cancer cell lines examined in this
study. GSDMA is expressed in the upper gastrointestinal tract but
is frequently silenced in gastric cancers. Reverse
transcription-quantitative PCR analysis was performed to determine
the expression level of GSDMA in esophageal and gastric cancer cell
lines and the results indicated that GSDMA was expressed in only
3/10 (33%) gastric cancer cell lines and 1/11 (9%) esophageal
cancer cell lines. GSDMA expression was observed in only 2 of the
60 primary cancer tissues, including esophageal and gastric cancer
(10). Saeki et al
(11) found that hypermethylation
of the promoters of GSDMA and its expression was able to be
restored by demethylation of the promoter. GSDMA overexpression
induces apoptosis in gastric epithelial cells. Peng et al
(12) found that the expression
level of GSDMA was upregulated in lung adenocarcinoma, whereas the
methylation level of GSDMA was reduced in cancer tissues compared
to that in normal tissues, which may account for the abnormal
expression in lung adenocarcinoma. Furthermore, the expression of
GSDMA is positively correlated with the infiltration of immune
cells in the TME, including B cells, CD8+ T cells,
CD4+ T cells, macrophages, neutrophils and dendritic
cells, suggesting an underlying association between GSDMA and the
effect of immunotherapy (12). A
similar trend was observed in ccRCC, in which the expression level
of GSDMA was significantly higher and positively associated with
the tumor grade, whereas the methylation level of GSDMA decreased.
The expression level of GSDMA was positively correlated with the
infiltration of macrophages, neutrophils and dendritic cells;
however, the GSDMA expression level was not associated with
relapse-free survival or overall survival (13). In luminal B breast cancer, Yang
et al (14) found that the
GSDMA expression level was higher than that in normal tissues.
Bioinformatics analysis showed that GSDMA was overexpressed in
ovarian cancer, and had a cancer-promoting role and a negative
impact on survival time (15);
however, the mechanism by which GSDMA acts in ovarian cancer
remains elusive (Table I). The
protease cleavage of GSDMA and its potential regulatory mechanisms
require further exploration to elucidate the association between
GSDMA and cancer.
GSDMB is located on 17q12 and the protein comprises
411 amino acids. Unlike other GSDM family members that are highly
homologous between humans and mice, GSDMB is absent in mice
(16). GSDMB has six splicing
variants in humans, each encoding a protein with a molecular weight
of 35-50 kDa. The expression levels and cellular localization of
the different isoforms vary. The domains of these isoforms are more
unstable than those of other GSDMs due to the unique sequence of
the connector between the C- and N-terminal domains, except for
isoform 5, which consists only of the C-terminal domain (17,18).
GSDMB may be cleaved by caspase-1, -3, -6 and -7, which promotes
the cleavage of GSDMB and the release of the N-terminal effector
domain, which oligomerizes in the cell membrane and forms pores,
resulting in pyroptosis (3). GSDMB
is not a substrate of caspase-4, -5 and -11 due to the absence of
the specific sequence (such as that in GSDMD) of the connector
between the C- and N-terminal domains (17). Although GSDMB cannot be cleaved by
caspase-4, it may induce the oligomerization of caspase-4 proteins,
thus increasing its activity and promoting the cleavage of GSDMD,
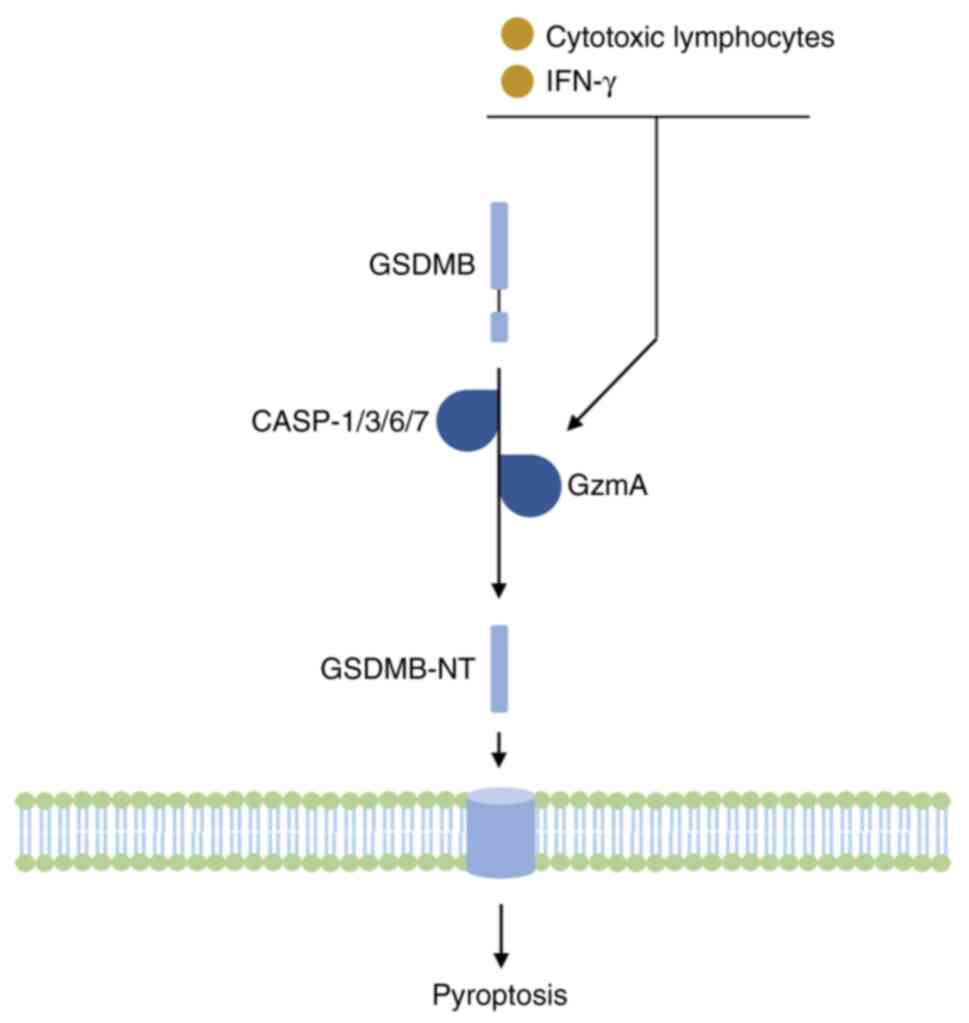
finally inducing non-canonical pyroptosis (19). In addition to specific caspases,
granzyme A from cytotoxic lymphocytes may directly cleave and
activate GSDMB to induce cell pyroptosis and promote cytotoxic T
lymphocyte-mediated tumor clearance in mice (20) (Fig.
1).
GSDMB is highly expressed in certain cancer types,
such as breast, cervical, uterine, and gastric cancers (21). Hergueta-Redondo et al
(22) found that GSDMB was
significantly upregulated in breast cancer compared to normal
controls and elucidated that isoform 2 of GSDMB induced invasion,
progression and metastasis of breast cancer cells and that GSDMB
may be a potential prognostic marker in breast cancer. Further
studies revealed that overexpression of GSDMB indicated invasive
behavior and poor prognosis, particularly in Erb-B2 receptor
tyrosine kinase 2 (HER2)-positive breast cancer, by increasing
therapy resistance (23-25), which also suggests that GSDMB may
be a potential therapeutic target. Molina-Crespo et al
(25) developed a GSDMB
antibody-based nanomedicine targeting intracellular oncoproteins
that inhibited the proliferation of breast cancer cells in
vitro, reduced tumor growth and metastasis in vivo and
enhanced therapy sensitivity. The mechanism of action of GSDMB in
breast cancer remains unclear and its potential therapeutic role
requires further study.
In addition to the above cancers, it was recently
found that GSDMB overexpression promotes bladder cancer progression
by interacting with STAT3, increasing its phosphorylation and
modulating glucose metabolism, thus promoting tumor proliferation
of bladder cancer (29).
Bioinformatics analysis revealed that the upregulated expression of
GSDMB is associated with immune infiltrates and poor survival in
ccRCC (13,30). The expression level of GSDMB is
upregulated in lung adenocarcinoma; accordingly, the methylation
level is lower in tumor tissues than in normal tissues.
Furthermore, the expression levels of GSDMB positively correlated
with the infiltration of B cells, CD4+ T cells and
dendritic cells, indicating that GSDMB may modulate the TME, as a
higher expression level of GSDMB suggests a poorer prognosis
(12) (Table II). Further studies are required
to determine the triggers underlying the mechanism by which GSDMB
participates in pyroptosis and the role of GSDMB in cancer.
GSDMC was originally identified as a
tumor-processing marker in human melanoma, which is located on 8q24
in humans, and there are four orthologous genes in mice (GSDMC1-4)
(5). GSDMC is expressed in the
trachea, spleen and gastrointestinal tract, such as the esophagus,
stomach, large and small intestines, cecum and colon (16,31,32).
There are differences in the expression patterns between humans and
mice, but the N-terminal domain has intrinsic cytotoxicity and may
induce pyroptosis in both human and mouse cells.
GSDMC has been found to be associated with the
development, stage, differentiation, therapeutic effect and
prognosis of multiple cancer types, which may or may not be
accompanied by pyroptosis; however, the underlying complex
mechanisms require further exploration.
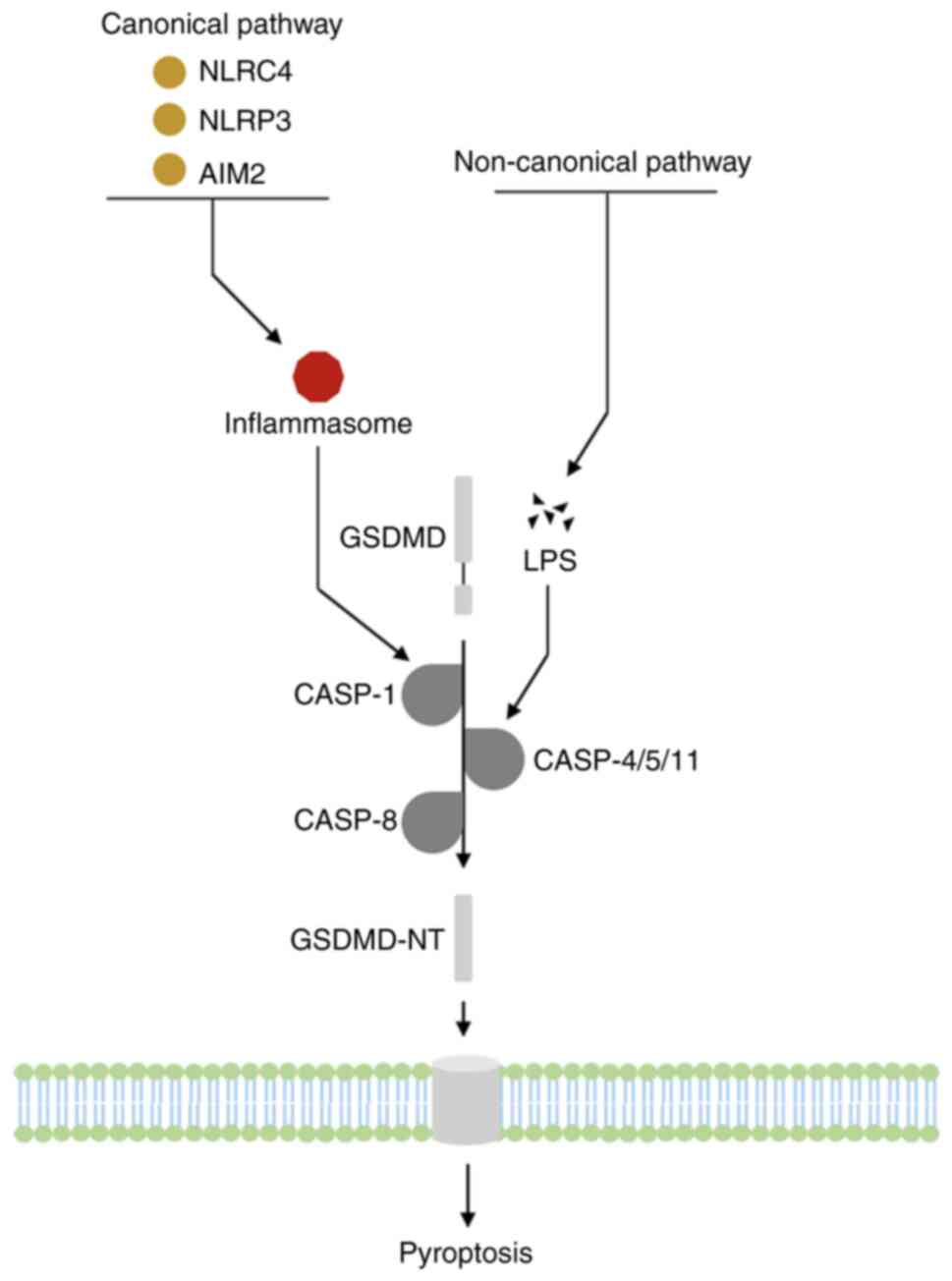
Almost all human organs and tissues, including
different subsets of leukocytes, express GSDMD at the mRNA and
protein levels (6,51). GSDMD participates in various
biological processes and has a role in numerous cancer types.
By contrast, GSDMD was observed to be downregulated
or silenced in several cancer types, such as colorectal, gastric
and ovarian cancers, in which pyroptosis-induced tumor
proliferation or metastasis is suppressed (44,64,65).
Saeki et al (10) reported
that GSDMD inhibited the proliferation of the gastric cancer cell
line MKN28 in a colony-formation assay and acted as a tumor
suppressor. In addition, inhibition of GSDMD expression affected
the cell cycle and accelerated the S/G2-phase transition by
activating the signal transducer and activator of transcription 3
(STAT3) and PI3K/AKT signaling pathways, indicating that the
inhibition of GSDMD may be a therapeutic target for gastric cancer
(66). Tanaka et al
(67) revealed that GSDMD
deficiency increases the development of colon cancer, partly due to
decreased apoptosis caused by the downregulation of IFN-γ-STAT1
signaling. Wang et al (68)
found that not only the expression level but also the subcellular
localization patterns were associated with colon cancer progression
and immune reactions; however, the underlying mechanisms remain
elusive (Table IV).
Therefore, despite the dual function (pro- and
anti-tumor) of GSDMD, it is closely involved in cancer incidence,
development and prognosis and may serve as a new and promising
target for cancer therapies.
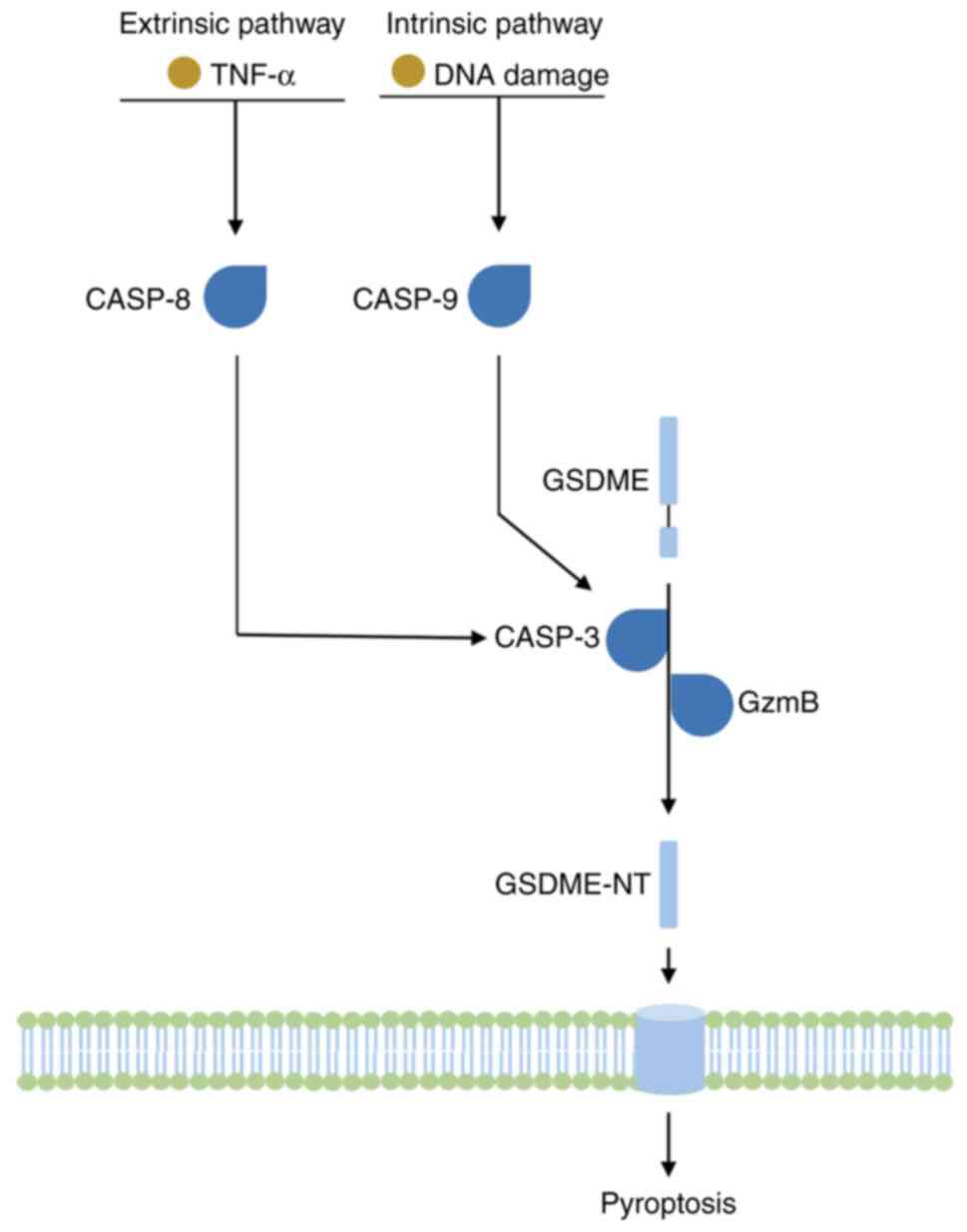
GSDME was primarily identified as a deafness-related
autosomal dominant gene, DFNA5, and an its encoding gene is located
on human chromosome 7q15, with the protein comprising 496 amino
acids (69). Unlike GSDMD,
GSDME-induced pyroptosis mainly relies on the activation of
caspase-3. Traditionally, caspase-3 is an apoptosis-related caspase
that may be activated under the treatment of TNF-α or chemotherapy
drugs, inducing cell apoptosis. However, when GSDME is present,
activated caspase-3 cleaves GSDME at Asp270 and induces pyroptosis
instead of apoptosis (70). In
addition to caspase-3, Zhang et al (71) found that granzyme B may also cleave
and activate GSDME directly at the same site as that cleaved by
caspase-3, and that the activation by granzyme B could not be
suppressed by a caspase-3 inhibitor (Fig. 4).
GSDME has an important role in regulating the
biological functions of tumors. In terms of tumor detection, GSDME
methylation may be an early detection biomarker. In terms of
treatment, immunotherapy and chemotherapy could kill tumor cells
through GSDME-induced pyroptosis. Combination with DNA methylation
inhibitors may improve its anti-tumor efficacy (92). However, GSDME-induced pyroptosis
may result in side effects such as nephrotoxicity (93) and lung injury (94). The related mechanisms are not fully
understood, and further research and support from clinical data are
required.
The gene encoding PJVK, also known as PVJK, is
located on human chromosome 2q31.2. Unlike other GSDMs, PJVK has a
truncated non-homologous C-terminal domain and lacks a cleavable
linker domain (31). PJVK was
reported to be the causative gene for autosomal recessive
nonsyndromic sensorineural hearing loss (95) and it was classified as a GSDM
member owing to its divergent expression patterns,
mutant-associated phenotypes and amino acid sequences compared with
those of other GSDMs (96). Human
PJVK is mainly expressed in the auditory system, including neurons,
hair cells, supporting cells and spiral ganglion cells in the inner
ear (96). However, whether PJVK
participates in membrane pore formation or the induction of
pyroptosis remains to be clarified. Only a small number of studies
have focused on the expression levels of PJVK in various cancer
types.
Studies have revealed that GSDMs are mainly cleaved
and activated by specific caspases, and have a crucial role in
pyroptosis. New mechanisms have also been proposed for mediating
pyroptosis. Demarco et al (48) reported that caspase-8-induced GSDMD
cleavage relies on caspase-8 dimerization and autoprocessing, and
that activated GSDMD imparts vulnerability to TNF-induced mortality
independent of caspase-1. Orning et al (97) found that in macrophages, caspase-8
was also able to act as a regulator of GSDMD-driven cell death, and
caspase-8-mediated GSDMD led to the NLRP3 inflammasome-dependent
release of IL-1β. Evavold et al (98) described macrophage hyperactivation,
which is a state in which the cell secretes IL-1 while retaining
viability, and GSDMD was indicated to have a key role and exert a
non-pyroptotic pore-forming function in the process, suggesting a
more complex nature of GSDMD. Certain GSDM family members have been
reported to exert synergistic effects. Rogers et al
(80) showed that GSDMD-mediated
pyroptosis facilitates the release of GSDME, which then amplifies
pyroptosis, leading to enhanced inflammation and immune responses.
Although the effects and mechanisms of pyroptosis in cancer
development remain to be fully elucidated, numerous studies have
indicated that the GSDM family of proteins have a role in the
incidence, development and prognosis of various cancer types. Most
GSDM proteins exhibit anti-tumor functions and are effective
targets for cancer therapy. Furthermore, the enhanced activity of
GSDM proteins increases the sensitivity to chemotherapy and
radiotherapy. Several studies have analyzed the relationship
between the expression levels of each GSDM family member. Mu et
al (99) systematically
analyzed the molecular characteristics and oncogenic role of the
GSDM family and found a correlation between the expression of the
GSDM genes and patient survival, indicating the prognostic value of
the expression of the GSDM genes. Zheng et al (100) systematically evaluated the gene
expression, genetic variations and prognostic values of GSDM family
members and found that expression levels were associated with
prognosis, clinical characteristics, TME features, and stemness
scores in several cancer types. Huo et al (101) confirmed that the risk or
protective effects of GSDM family members on prognosis depend on
the cancer type; the mutation frequency appeared to be high and the
mutation group had a worse prognosis. GSDMs may be upregulated,
downregulated or even silenced in cancers, and their expression
levels are mainly associated with the DNA methylation level of
their promoters. Higher methylation levels inhibited GSDM
expression, whereas demethylation was able to activate the
expression of GSDMs, indicating that GSDM methylation may be an
early diagnostic biomarker, a prognostic marker and a therapeutic
target for combined treatment.
Although pyroptosis may be harnessed to kill cancer
cells, it must be noted that during the process of pyroptosis,
large amounts of inflammatory substances, such as IL-1β and IL-18,
are released, resulting in a cascade of inflammatory reactions that
may be harmful to normal cells. Liu et al (102) also reported a complication after
chimeric antigen receptor (CAR) T-cell therapy called cytokine
release syndrome (CRS). CAR-T cells promote the release of granzyme
B, activate caspase-3-dependent cleavage of GSDME and induce the
release of pyroptosis-related inflammatory factor by macrophages,
further activating caspase-1-mediated GSDMD cleavage in macrophages
and causing the release of more cytokines and subsequent CRS.
However, CRS did not occur after the knockout of GSDME, reduction
in macrophages or inhibition of caspase-1 (102). Pyroptosis causes local
inflammation and acts as a highly immunogenic form of cell death
with the release of multiple chemokines, leading to the recruitment
and infiltration of immune cells, providing a great opportunity to
relieve the immunosuppression of the tumor immune microenvironment
and induce systemic immune responses in the treatment of solid
tumors. GSDMD-, GSDME-, GSDMC- and GSDMB-mediated pyroptosis has
been reported to enhance the immune response in various cancer
types (70). However, chronic
tumor pyroptosis may also suppress anti-tumor immunity and
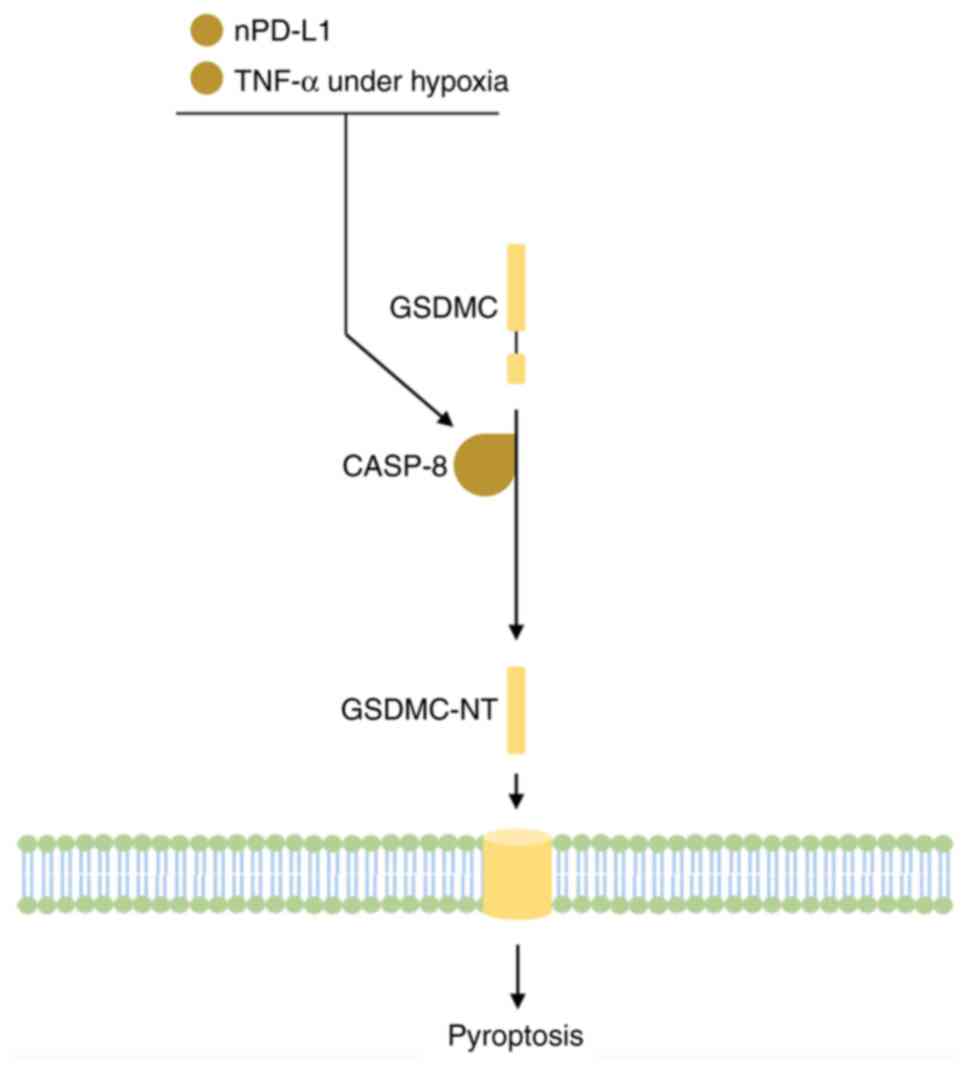
accelerate tumor growth. Hou et al (33) found that solid tumors had hypoxic
areas in which the expression of GSDMC was enhanced and cleaved by
TNFα-activated caspase-8, which led to pyroptosis and subsequent
chronic tumor necrosis while suppressing anti-tumor immune
responses. Therefore, GSDM-induced pyroptosis has a dual role in
tumorigenesis, regression and the tumor immune microenvironment
(103,104).
A pan-cancer analysis of the genetic variation in
GSDM genes revealed that in pan-cancer tissues, the overall
mutation frequency in GSDM genes was relatively low, and missense
mutations were the most frequent form of mutation. Studies have
partly elucidated the relationship between GSDM mutations and
specific diseases, including various cancers. Ruan et al
(105) analyzed the structure of
the GSDMA3 membrane pores using cryo-electron microscopy and partly
revealed the mechanisms by which the cleavage of GSDMA3 formed
membrane pores and the mechanisms of autoinhibition. Due to a loss
of autoinhibition, disease-related mutations in GSDMA3 and its N
terminus alone can initiate pyroptosis and are closely associated
with spontaneous alopecia and hyperkeratosis (3). SNPs in GSDMA and GSDMB have been
reported to be related to childhood asthma and, to a lesser extent,
adult asthma (96). The GSDMB SNPs
are also associated with cancer development and progression
(21,28). Somatic mutations in GSDMC have been
reported in breast cancer (34).
Mutations in GSDME and PJVK induce deafness through different
mechanisms. Mutations in GSDME result in its overexpression,
leading to pyroptosis in HeLa cells (72), whereas PJVK mutations exert
non-pyroptosis functions (95).
Xia et al (106) reported
the cryo-electron microscopy structures of the pores of GSDMD,
elucidating the process of GSDMD-dependent membrane pore formation
and the GSDMD-mediated release of IL-1β. Liu et al (107) revealed the mechanisms of
autoinhibition, lipid binding and oligomerization of the
GSDMD-N-terminus by virtue of its crystal structures. These
molecular structure studies provide an explanation for the mode of
action of several mutant GSDM family members, including mutants
linked to cancer (105).
Considering that pyroptosis signaling pathways are
deeply involved in the incidence and development of different
cancer types, certain agents have been developed or discovered to
exert protective effects against cancers by targeting pyroptosis
pathways, either by inducing or inhibiting pyroptosis.
Small-molecule inhibitors of di-peptidyl-peptidase 8/9, two serine
proteases in host cells, have been reported to activate NLRP1 and
caspase recruitment domain-containing protein 8, and induce
pyroptosis via the caspase-1/GSDMD pathway, thus inhibiting the
progression of acute myeloid leukemia (108,109). It was also revealed that FL118,
anthocyanin and docosahexaenoic acid have anti-tumor activity
against colorectal cancer (110),
oral squamous cell carcinoma (111) and breast cancer (58) partly by promoting
NLRP3/caspase1/GSDMD-mediated pyroptosis. Traditional chemotherapy
drugs, such as cisplatin, paclitaxel, doxorubicin and lobaplatin,
have also been found to induce pyroptosis via the caspase-3/GSDME
pathway to inhibit the development of lung cancer (112), ESCC (77), melanoma (113) and colon cancer (114). Previously, the mechanisms of the
above chemotherapy drugs in exerting anti-cancer effects were
thought to be mainly dependent on apoptosis, and updated evidence
has shown that it is important to identify whether the therapeutic
effects are due to the differential activation of apoptosis or
pyroptosis. Due to the double-edged sword effect of pyroptosis in
cancer development, pyroptosis inhibitors also show therapeutic
potential, most of which were developed to target the molecules
upstream of GSDMs. Rathkey et al (115) reported that necrosulfonamide
(NSA) was able to directly bind to GSDMD and inhibit the cleavage
of GSDMD, blocking pyroptosis cell death and IL-1β release. Zhou
et al (116) also found
that NSA represses NLRP3 inflammasome-mediated pyroptosis,
indicating that NSA may be an effective agent for targeting
pyroptosis. Disulfiram inhibits pyroptosis by blocking
GSDMD-induced membrane pore formation, indicating its potential as
a therapeutic method for cancer and other inflammatory illnesses
exacerbated by pyroptosis (117).
NLRP3 and caspase inhibitors have been explored for the treatment
of neurodegenerative diseases by targeting overactivated pyroptosis
and have shown certain effects (44,118,119). Recently, LDC7559, a newly
developed selective GSDMD inhibitor, was reported to promote brain
functional recovery by inhibiting over-activated pyroptosis by
directly blocking the GSDMD-N-terminus (120,121). However, whether these inhibitors
may also be used for cancer treatment remains to be investigated.
To the best of our knowledge, there are currently no other specific
inhibitors of the GSDM family members, and this would be a further
direction for exploring more potent anti-tumor treatments.
Regarding the effects of GSDMs and their regulatory
mechanisms, we are beginning to understand the molecular,
biological and pathological functions of GSDMs in pyroptosis.
Further research to elucidate the mechanisms of GSDM-mediated
pyroptosis will deepen our understanding of the roles of GSDM
family proteins in cancers and provide new ideas for developing
more potent anti-tumor methods.
Not applicable.
XY carried out the primary literature search and
drafted and revised the manuscript. ZT contributed to drafting and
revising the manuscript. XY and ZT performed literature analyses.
All the authors have read and approved the final version of the
manuscript. Data authentication is not applicable.
Not applicable.
Not applicable.
The authors declare that they have no competing
interests.
Not applicable.
No funding was received.
|
1
|
Xia C, Dong X, Li H, Cao M, Sun D, He S,
Yang F, Yan X, Zhang S, Li N and Chen W: Cancer statistics in China
and United States, 2022: Profiles, trends, and determinants. Chin
Med J (Engl). 135:584–590. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Tonnus W, Meyer C, Paliege A, Belavgeni A,
von Mässenhausen A, Bornstein SR, Hugo C, Becker JU and Linkermann
A: The pathological features of regulated necrosis. J Pathol.
247:697–707. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Ding J, Wang K, Liu W, She Y, Sun Q, Shi
J, Sun H, Wang DC and Shao F: Pore-forming activity and structural
autoinhibition of the gasdermin family. Nature. 535:111–116. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Tanaka S, Mizushina Y, Kato Y, Tamura M
and Shiroishi T: Functional conservation of Gsdma cluster genes
specifically duplicated in the mouse genome. G3 (Bethesda).
3:1843–1850. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Zou J, Zheng Y, Huang Y, Tang D, Kang R
and Chen R: The versatile gasdermin family: Their function and
roles in diseases. Front Immunol. 12:7515332021. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Rieckmann JC, Geiger R, Hornburg D, Wolf
T, Kveler K, Jarrossay D, Sallusto F, Shen-Orr SS, Lanzavecchia A,
Mann M and Meissner F: Social network architecture of human immune
cells unveiled by quantitative proteomics. Nat Immunol. 18:583–593.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Lunny DP, Weed E, Nolan PM, Marquardt A,
Augustin M and Porter RM: Mutations in gasdermin 3 cause aberrant
differentiation of the hair follicle and sebaceous gland. J Invest
Dermatol. 124:615–621. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Wang Q, Wang Y, Ding J, Wang C, Zhou X,
Gao W, Huang H, Shao F and Liu Z: A bioorthogonal system reveals
antitumour immune function of pyroptosis. Nature. 579:421–426.
2020. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Saeki N, Kuwahara Y, Sasaki H, Satoh H and
Shiroishi T: Gasdermin (Gsdm) localizing to mouse Chromosome 11 is
predominantly expressed in upper gastrointestinal tract but
significantly suppressed in human gastric cancer cells. Mamm
Genome. 11:718–724. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Saeki N, Usui T, Aoyagi K, Kim DH, Sato M,
Mabuchi T, Yanagihara K, Ogawa K, Sakamoto H, Yoshida T and Sasaki
H: Distinctive expression and function of four GSDM family genes
(GSDMA-D) in normal and malignant upper gastrointestinal
epithelium. Genes Chromosomes Cancer. 48:261–271. 2009. View Article : Google Scholar
|
|
11
|
Saeki N, Kim DH, Usui T, Aoyagi K, Tatsuta
T, Aoki K, Yanagihara K, Tamura M, Mizushima H, Sakamoto H, et al:
GASDERMIN, suppressed frequently in gastric cancer, is a target of
LMO1 in TGF-beta-dependent apoptotic signalling. Oncogene.
26:6488–6498. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Peng LS, Duan SL, Li RQ, Wang D, Han YY,
Huang T, Yu YP, Ou CL and Wang JP: Prognostic value and immune
infiltration of the gasdermin family in lung adenocarcinoma. Front
Oncol. 12:10438622022. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Yao L, Li J, Xu Z, Yan Y and Hu K: GSDMs
are potential therapeutic targets and prognostic biomarkers in
clear cell renal cell carcinoma. Aging (Albany NY). 14:2758–2774.
2022. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Yang C, Liu J, Zhao S, Ying J, Liu Y, Ma
L, Shang Q, Meng X, Feng K, Zheng B, et al: Establishment and
validation of a gasdermin signature to evaluate the immune status
and direct risk-group classification in luminal-B breast cancer.
Clin Transl Med. 11:e6142021. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Ye Y, Dai Q and Qi H: A novel defined
pyroptosis-related gene signature for predicting the prognosis of
ovarian cancer. Cell Death Discov. 7:712021. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Tamura M, Tanaka S, Fujii T, Aoki A,
Komiyama H, Ezawa K, Sumiyama K, Sagai T and Shiroishi T: Members
of a novel gene family, Gsdm, are expressed exclusively in the
epithelium of the skin and gastrointestinal tract in a highly
tissue-specific manner. Genomics. 89:618–629. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Chao KL, Kulakova L and Herzberg O: Gene
polymorphism linked to increased asthma and IBD risk alters
gasdermin-B structure, a sulfatide and phosphoinositide binding
protein. Proc Natl Acad Sci USA. 114:E1128–E1137. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Ruan J: Structural insight of gasdermin
family driving pyroptotic cell death. Adv Exp Med Biol.
1172:189–205. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Chen Q, Shi P, Wang Y, Zou D, Wu X, Wang
D, Hu Q, Zou Y, Huang Z, Ren J, et al: GSDMB promotes non-canonical
pyroptosis by enhancing caspase-4 activity. J Mol Cell Biol.
11:496–508. 2019. View Article : Google Scholar :
|
|
20
|
Zhou Z, He H, Wang K, Shi X, Wang Y, Su Y,
Wang Y, Li D, Liu W, Zhang Y, et al: Granzyme A from cytotoxic
lymphocytes cleaves GSDMB to trigger pyroptosis in target cells.
Science. 368:eaaz75482020. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Lutkowska A, Roszak A, Lianeri M, Sowinska
A, Sotiri E and Jagodzinski PP: Analysis of rs8067378 polymorphism
in the risk of uterine cervical cancer from a polish population and
its impact on gasdermin B expression. Mol Diagn Ther. 21:199–207.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Hergueta-Redondo M, Sarrio D,
Molina-Crespo A, Megias D, Mota A, Rojo-Sebastian A, García-Sanz P,
Morales S, Abril S, Cano A, et al: Gasdermin-B promotes invasion
and metastasis in breast cancer cells. PLoS One. 9:e900992014.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Hergueta-Redondo M, Sarrio D,
Molina-Crespo A, Vicario R, Bernadó-Morales C, Martínez L,
Rojo-Sebastián A, Serra-Musach J, Mota A, Martínez-Ramírez Á, et
al: Gasdermin B expression predicts poor clinical outcome in
HER2-positive breast cancer. Oncotarget. 7:56295–56308. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Gamez-Chiachio M, Molina-Crespo A,
Ramos-Nebot C, Martinez-Val J, Martinez L, Gassner K, Llobet FJ,
Soriano M, Hernandez A, Cordani M, et al: Gasdermin B
over-expression modulates HER2-targeted therapy resistance by
inducing protective autophagy through Rab7 activation. J Exp Clin
Cancer Res. 41:2852022. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Molina-Crespo A, Cadete A, Sarrio D,
Gámez-Chiachio M, Martinez L, Chao K, Olivera A, Gonella A, Díaz E,
Palacios J, et al: Intracellular delivery of an antibody targeting
gasdermin-B Reduces HER2 breast cancer aggressiveness. Clin Cancer
Res. 25:4846–4858. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Komiyama H, Aoki A, Tanaka S, Maekawa H,
Kato Y, Wada R, Maekawa T, Tamura M and Shiroishi T: Alu-derived
cis-element regulates tumorigenesis-dependent gastric expression of
GASDERMIN B (GSDMB). Genes Genet Syst. 85:75–83. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Saeki N, Komatsuzaki R, Chiwaki F,
Yanagihara K and Sasaki H: A GSDMB enhancer-driven HSV thymidine
kinase-expressing vector for controlling occult peritoneal
dissemination of gastric cancer cells. BMC Cancer. 15:4392015.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Li S, Li X, Zhang S, Feng Y, Jia T, Zhu M,
Fang L, Gong L, Dong S, Kong X, et al: Association Between GSDMB
gene polymorphism and cervical cancer in the Northeast Chinese Han
Population. Front Genet. 13:8607272022. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
He H, Yi L, Zhang B, Yan B, Xiao M, Ren J,
Zi D, Zhu L, Zhong Z, Zhao X, et al: USP24-GSDMB complex promotes
bladder cancer proliferation via activation of the STAT3 pathway.
Int J Biol Sci. 17:2417–2429. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Cui Y, Zhou Z, Chai Y and Zhang Y:
Upregulated GSDMB in clear cell renal cell carcinoma is associated
with immune infiltrates and poor prognosis. J Immunol Res.
2021:77535532021. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Orning P, Lien E and Fitzgerald KA:
Gasdermins and their role in immunity and inflammation. J Exp Med.
216:2453–2465. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Watabe K, Ito A, Asada H, Endo Y,
Kobayashi T, Nakamoto K, Itami S, Takao S, Shinomura Y, Aikou T, et
al: Structure, expression and chromosome mapping of MLZE, a novel
gene which is preferentially expressed in metastatic melanoma
cells. Jpn J Cancer Res. 92:140–151. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Hou J, Zhao R, Xia W, Chang CW, You Y, Hsu
JM, Nie L, Chen Y, Wang YC, Liu C, et al: PD-L1-mediated gasdermin
C expression switches apoptosis to pyroptosis in cancer cells and
facilitates tumour necrosis. Nat Cell Biol. 22:1264–1275. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Xu D, Ji Z and Qiang L: Molecular
characteristics, clinical implication, and cancer immunity
interactions of pyroptosis-related genes in breast cancer. Front
Med (Lausanne). 8:7026382021. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Sun K, Chen RX, Li JZ and Luo ZX:
LINC00511/hsa-miR-573 axis-mediated high expression of Gasdermin C
associates with dismal prognosis and tumor immune infiltration of
breast cancer. Sci Rep. 12:147882022. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Zhang JY, Zhou B, Sun RY, Ai YL, Cheng K,
Li FN, Wang BR, Liu FJ, Jiang ZH, Wang WJ, et al: The metabolite
α-KG induces GSDMC-dependent pyroptosis through death receptor
6-activated caspase-8. Cell Res. 31:980–997. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Wei J, Xu Z, Chen X, Wang X, Zeng S, Qian
L, Yang X, Ou C, Lin W, Gong Z and Yan Y: Overexpression of GSDMC
is a prognostic factor for predicting a poor outcome in lung
adenocarcinoma. Mol Med Rep. 21:360–370. 2020.PubMed/NCBI
|
|
38
|
Miguchi M, Hinoi T, Shimomura M, Adachi T,
Saito Y, Niitsu H, Kochi M, Sada H, Sotomaru Y, Ikenoue T, et al:
Gasdermin C is upregulated by inactivation of transforming growth
factor beta receptor type II in the presence of mutated Apc,
promoting colorectal cancer proliferation. PLoS One.
11:e01664222016. View Article : Google Scholar
|
|
39
|
Pereira BS, Wisnieski F, Calcagno DQ,
Santos LC, Gigek CO, Chen ES, Rasmussen LT, Payão SLM, Almeida RS,
Pinto CA, et al: Genetic and transcriptional analysis of 8q24.21
cluster in gastric cancer. Anticancer Res. 42:4381–4394. 2022.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Yan C, Niu Y, Li F, Zhao W and Ma L:
System analysis based on the pyroptosis-related genes identifies
GSDMC as a novel therapy target for pancreatic adenocarcinoma. J
Transl Med. 20:4552022. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Berkel C and Cacan E: Differential
expression and copy number variation of gasdermin (GSDM) family
members, pore-forming proteins in pyroptosis, in normal and
malignant serous ovarian tissue. Inflammation. 44:2203–2216. 2021.
View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Kayagaki N, Stowe IB, Lee BL, O'Rourke K,
Anderson K, Warming S, Cuellar T, Haley B, Roose-Girma M, Phung QT,
et al: Caspase-11 cleaves gasdermin D for non-canonical
inflammasome signalling. Nature. 526:666–671. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Shi J, Zhao Y, Wang K, Shi X, Wang Y,
Huang H, Zhuang Y, Cai T, Wang F and Shao F: Cleavage of GSDMD by
inflammatory caspases determines pyroptotic cell death. Nature.
526:660–665. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Burdette BE, Esparza AN, Zhu H and Wang S:
Gasdermin D in pyroptosis. Acta Pharm Sin B. 11:2768–2782. 2021.
View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Broz P and Dixit VM: Inflammasomes:
Mechanism of assembly, regulation and signalling. Nat Rev Immunol.
16:407–420. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Benaoudia S, Martin A, Puig Gamez M, Gay
G, Lagrange B, Cornut M, Krasnykov K, Claude JB, Bourgeois CF,
Hughes S, et al: A genome-wide screen identifies IRF2 as a key
regulator of caspase-4 in human cells. EMBO Rep. 20:e482352019.
View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Kayagaki N, Lee BL, Stowe IB, Kornfeld OS,
O'Rourke K, Mirrashidi KM, Haley B, Watanabe C, Roose-Girma M,
Modrusan Z, et al: IRF2 transcriptionally induces GSDMD expression
for pyroptosis. Sci Signal. 12:eaax49172019. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Demarco B, Grayczyk JP, Bjanes E, Le Roy
D, Tonnus W, Assenmacher CA, Radaelli E, Fettrelet T, Mack V,
Linkermann A, et al: Caspase-8-dependent gasdermin D cleavage
promotes antimicrobial defense but confers susceptibility to
TNF-induced lethality. Sci Adv. 6:eabc34652020. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Brubaker SW, Brewer SM, Massis LM, Napier
BA and Monack DM: A Rapid Caspase-11 Response Induced by IFNү
priming is independent of guanylate binding proteins. iScience.
23:1016122020. View Article : Google Scholar
|
|
50
|
Zhang C, Zhao C, Chen X, Tao R, Wang S,
Meng G, Liu X, Shao C and Su X: Induction of ASC pyroptosis
requires gasdermin D or caspase-1/11-dependent mediators and IFNβ
from pyroptotic macrophages. Cell Death Dis. 11:4702020. View Article : Google Scholar
|
|
51
|
Fagerberg L, Hallstrom BM, Oksvold P,
Kampf C, Djureinovic D, Odeberg J, Habuka M, Tahmasebpoor S,
Danielsson A, Edlund K, et al: Analysis of the human
tissue-specific expression by genome-wide integration of
transcriptomics and antibody-based proteomics. Mol Cell Proteomics.
13:397–406. 2014. View Article : Google Scholar :
|
|
52
|
Zhang J, Chen Y and He Q: Distinct
characteristics of dasatinib-induced pyroptosis in gasdermin
E-expressing human lung cancer A549 cells and neuroblastoma SH-SY5Y
cells. Oncol Lett. 20:145–154. 2020. View Article : Google Scholar
|
|
53
|
Xi G, Gao J, Wan B, Zhan P, Xu W, Lv T and
Song Y: GSDMD is required for effector CD8(+) T cell responses to
lung cancer cells. Int Immunopharmacol. 74:1057132019. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Traughber CA, Deshpande GM, Neupane K,
Bhandari N, Khan MR, McMullen MR, Swaidani S, Opoku E, Muppala S,
Smith JD, et al: Myeloid-cell-specific role of Gasdermin D in
promoting lung cancer progression in mice. iScience. 26:1060762023.
View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Gao J, Qiu X, Xi G, Liu H, Zhang F, Lv T
and Song Y: Downregulation of GSDMD attenuates tumor proliferation
via the intrinsic mitochondrial apoptotic pathway and inhibition of
EGFR/Akt signaling and predicts a good prognosis in non-small cell
lung cancer. Oncol Rep. 40:1971–1984. 2018.PubMed/NCBI
|
|
56
|
Peng J, Jiang H, Guo J, Huang J, Yuan Q,
Xie J and Xiao K: CD147 Expression is associated with tumor
proliferation in bladder cancer via GSDMD. Biomed Res Int.
2020:76389752020. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Wang L, Li K, Lin X, Yao Z, Wang S, Xiong
X, Ning Z, Wang J, Xu X, Jiang Y, et al: Metformin induces human
esophageal carcinoma cell pyroptosis by targeting the miR-497/PELP1
axis. Cancer Lett. 450:22–31. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Pizato N, Luzete BC, Kiffer LFMV, Corrêa
LH, de Oliveira Santos I, Assumpção JAF, Ito MK and Magalhães KG:
Omega-3 docosahexaenoic acid induces pyroptosis cell death in
triple-negative breast cancer cells. Sci Rep. 8:19522018.
View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Yan H, Luo B, Wu X, Guan F, Yu X, Zhao L,
Ke X, Wu J and Yuan J: Cisplatin induces pyroptosis via activation
of MEG3/NLRP3/caspase-1/GSDMD pathway in triple-negative breast
cancer. Int J Biol Sci. 17:2606–2621. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Yang Y, Liu PY, Bao W, Chen SJ, Wu FS and
Zhu PY: Hydrogen inhibits endometrial cancer growth via a
ROS/NLRP3/caspase-1/GSDMD-mediated pyroptotic pathway. BMC Cancer.
20:282020. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Shen X, Zhang Q, He Z, Xiao S, Li H and
Huang Z: Overexpression of gasdermin D promotes invasion of adenoid
cystic carcinoma. Int J Clin Exp Pathol. 13:1802–1811.
2020.PubMed/NCBI
|
|
62
|
Lv T, Xiong X, Yan W, Liu M, Xu H and He
Q: Targeting of GSDMD sensitizes HCC to anti-PD-1 by activating
cGAS pathway and downregulating PD-L1 expression. J Immunother
Cancer. 10:e0047632022. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Yamagishi R, Kamachi F, Nakamura M,
Yamazaki S, Kamiya T, Takasugi M, Cheng Y, Nonaka Y, Yukawa-Muto Y,
Thuy LTT, et al: Gasdermin D-mediated release of IL-33 from
senescent hepatic stellate cells promotes obesity-associated
hepatocellular carcinoma. Sci Immunol. 7:eabl72092022. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Fang Y, Tian S, Pan Y, Li W, Wang Q, Tang
Y, Yu T, Wu X, Shi Y, Ma P and Shu Y: Pyroptosis: A new frontier in
cancer. Biomed Pharmacother. 121:1095952020. View Article : Google Scholar
|
|
65
|
Qiao L, Wu X, Zhang J, Liu L, Sui X, Zhang
R, Liu W, Shen F, Sun Y and Xi X: α-NETA induces pyroptosis of
epithelial ovarian cancer cells through the GSDMD/caspase-4
pathway. FASEB J. 33:12760–12767. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Wang WJ, Chen D, Jiang MZ, Xu B, Li XW,
Chu Y, Zhang YJ, Mao R, Liang J and Fan DM: Downregulation of
gasdermin D promotes gastric cancer proliferation by regulating
cell cycle-related proteins. J Dig Dis. 19:74–83. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Tanaka S, Orita H, Kataoka T, Miyazaki M,
Saeki H, Wada R, Brock MV, Fukunaga T, Amano T and Shiroishi T:
Gasdermin D represses inflammation-induced colon cancer development
by regulating apoptosis. Carcinogenesis. 44:341–349. 2023.
View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Wang J, Kang Y, Li Y, Sun L, Zhang J, Qian
S, Luo K, Jiang Y, Sun L and Xu F: Gasdermin D in different
subcellular locations predicts diverse progression, immune
microenvironment and prognosis in colorectal cancer. J Inflamm Res.
14:6223–6235. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Masuda Y, Futamura M, Kamino H, Nakamura
Y, Kitamura N, Ohnishi S, Miyamoto Y, Ichikawa H, Ohta T, Ohki M,
et al: The potential role of DFNA5, a hearing impairment gene, in
p53-mediated cellular response to DNA damage. J Hum Genet.
51:652–664. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Ouyang X, Zhou J, Lin L, Zhang Z, Luo S
and Hu D: Pyroptosis, inflammasome, and gasdermins in tumor
immunity. Innate Immun. 29:3–13. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Zhang Z, Zhang Y, Xia S, Kong Q, Li S, Liu
X, Junqueira C, Meza-Sosa KF, Mok TMY, Ansara J, et al: Gasdermin E
suppresses tumour growth by activating anti-tumour immunity.
Nature. 579:415–420. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Wang Y, Gao W, Shi X, Ding J, Liu W, He H,
Wang K and Shao F: Chemotherapy drugs induce pyroptosis through
caspase-3 cleavage of a gasdermin. Nature. 547:99–103. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Rogers C, Fernandes-Alnemri T, Mayes L,
Alnemri D, Cingolani G and Alnemri ES: Cleavage of DFNA5 by
caspase-3 during apoptosis mediates progression to secondary
necrotic/pyroptotic cell death. Nat Commun. 8:141282017. View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Jia C, Zhang Z, Tang J, Cai MC, Zang J,
Shi K, Sun Y, Wu J, Shi H, Shi W, et al: Epithelial-Mesenchymal
transition induces GSDME transcriptional activation for
inflammatory pyroptosis. Front Cell Dev Biol. 9:7813652021.
View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Vernon M, Wilski NA, Kotas D, Cai W,
Pomante D, Tiago M, Alnemri ES and Aplin AE: Raptinal induces
gasdermin E-Dependent pyroptosis in naive and therapy-resistant
melanoma. Mol Cancer Res. 20:1811–1821. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
76
|
Hu J, Pei W, Jiang M, Huang Y, Dong F,
Jiang Z, Xu Y and Li Z: DFNA5 regulates immune cells infiltration
and exhaustion. Cancer Cell Int. 22:1072022. View Article : Google Scholar : PubMed/NCBI
|
|
77
|
Wu M, Wang Y, Yang D, Gong Y, Rao F, Liu
R, Danna Y, Li J, Fan J, Chen J, et al: A PLK1 kinase inhibitor
enhances the chemosensitivity of cisplatin by inducing pyroptosis
in oesophageal squamous cell carcinoma. EBioMedicine. 41:244–255.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
78
|
Liu Z, Liu H, Dong Q, Li H, Zhang B, Liu
Y, Zhong L and Tang H: Prognostic role of DFNA5 in head and neck
squamous cell carcinoma revealed by systematic expression analysis.
BMC Cancer. 21:9512021. View Article : Google Scholar : PubMed/NCBI
|
|
79
|
Yin J, Che G, Wang W, Chen S and Liu J:
Investigating the prognostic significance of pyroptosis-related
genes in gastric cancer and their impact on cells' biological
functions. Front Oncol. 12:8612842022. View Article : Google Scholar : PubMed/NCBI
|
|
80
|
Rogers C, Erkes DA, Nardone A, Aplin AE,
Fernandes-Alnemri T and Alnemri ES: Gasdermin pores permeabilize
mitochondria to augment caspase-3 activation during apoptosis and
inflammasome activation. Nat Commun. 10:16892019. View Article : Google Scholar : PubMed/NCBI
|
|
81
|
Liu Z, Li Y, Zhu Y, Li N, Li W, Shang C,
Song G, Li S, Cong J, Li T, et al: Apoptin induces pyroptosis of
colorectal cancer cells via the GSDME-dependent pathway. Int J Biol
Sci. 18:717–730. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
82
|
Zhang Z, Zhang H, Li D, Zhou X, Qin Q and
Zhang Q: Caspase-3-mediated GSDME induced Pyroptosis in breast
cancer cells through the ROS/JNK signalling pathway. J Cell Mol
Med. 25:8159–8168. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
83
|
An H, Heo JS, Kim P, Lian Z, Lee S, Park
J, Hong E, Pang K, Park Y, Ooshima A, et al: Tetraarsenic hexoxide
enhances generation of mitochondrial ROS to promote pyroptosis by
inducing the activation of caspase-3/GSDME in triple-negative
breast cancer cells. Cell Death Dis. 12:1592021. View Article : Google Scholar : PubMed/NCBI
|
|
84
|
de Beeck KO, Van Laer L and Van Camp G:
DFNA5, a gene involved in hearing loss and cancer: A review. Ann
Otol Rhinol Laryngol. 121:197–207. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
85
|
Akino K, Toyota M, Suzuki H, Imai T,
Maruyama R, Kusano M, Nishikawa N, Watanabe Y, Sasaki Y, Abe T, et
al: Identification of DFNA5 as a target of epigenetic inactivation
in gastric cancer. Cancer Sci. 98:88–95. 2007. View Article : Google Scholar
|
|
86
|
Ibrahim J, Op de Beeck K, Fransen E,
Fransen E, Croes L, Beyens M, Suls A, Vanden Berghe W, Peeters M
and Van Camp G: Methylation analysis of Gasdermin E shows great
promise as a biomarker for colorectal cancer. Cancer Med.
8:2133–2145. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
87
|
Liu X, Xia S, Zhang Z, Wu H and Lieberman
J: Channelling inflammation: Gasdermins in physiology and disease.
Nat Rev Drug Discov. 20:384–405. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
88
|
Guo J, Yu J, Mu M, Chen Z, Xu Z, Zhao C,
Yang K, Zheng J, Qin X, Zhao W and Sun X: DFNA5 inhibits colorectal
cancer proliferation by suppressing the mTORC1/2 signaling pathways
via upregulation of DEPTOR. Cell Cycle. 21:2165–2178. 2022.
View Article : Google Scholar : PubMed/NCBI
|
|
89
|
Tan G, Lin C, Huang C, Chen B, Chen J, Shi
Y and Zhi F: Radiosensitivity of colorectal cancer and
radiation-induced gut damages are regulated by gasdermin E. Cancer
Lett. 529:1–10. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
90
|
Di M, Miao J, Pan Q, Wu Z, Chen B, Wang M,
Zhao J, Huang H, Bai J, Wang Q, et al: OTUD4-mediated GSDME
deubiquitination enhances radiosensitivity in nasopharyngeal
carcinoma by inducing pyroptosis. J Exp Clin Cancer Res.
41:3282022. View Article : Google Scholar : PubMed/NCBI
|
|
91
|
Li F, Xia Q, Ren L, Nie Y, Ren H, Guo X,
Yu J, Xing Y and Chen Z: GSDME increases chemotherapeutic drug
sensitivity by inducing pyroptosis in retinoblastoma cells. Oxid
Med Cell Longev. 2022:23718072022.PubMed/NCBI
|
|
92
|
Xie B, Liu T, Chen S, Zhang Y, He D, Shao
Q, Zhang Z and Wang C: Combination of DNA demethylation and
chemotherapy to trigger cell pyroptosis for inhalation treatment of
lung cancer. Nanoscale. 13:18608–18615. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
93
|
Shen X, Wang H, Weng C, Jiang H and Chen
J: Caspase 3/GSDME-dependent pyroptosis contributes to chemotherapy
drug-induced nephrotoxicity. Cell Death Dis. 12:1862021. View Article : Google Scholar : PubMed/NCBI
|
|
94
|
Mai FY, He P, Ye JZ, Xu LH, Ouyang DY, Li
CG, Zeng QZ, Zeng CY, Zhang CC, He XH and Hu B: Caspase-3-mediated
GSDME activation contributes to cisplatin- and doxorubicin-induced
secondary necrosis in mouse macrophages. Cell Prolif.
52:e126632019. View Article : Google Scholar : PubMed/NCBI
|
|
95
|
Delmaghani S, del Castillo FJ, Michel V,
Leibovici M, Aghaie A, Ron U, Van Laer L, Ben-Tal N, Van Camp G,
Weil D, et al: Mutations in the gene encoding pejvakin, a newly
identified protein of the afferent auditory pathway, cause DFNB59
auditory neuropathy. Nat Genet. 38:770–778. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
96
|
Zheng Z, Deng W, Lou X, Bai Y, Wang J,
Zeng H, Gong S and Liu X: Gasdermins: Pore-forming activities and
beyond. Acta Biochim Biophys Sin (Shanghai). 52:467–474. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
97
|
Orning P, Weng D, Starheim K, Ratner D,
Best Z, Lee B, Brooks A, Xia S, Wu H, Kelliher MA, et al: Pathogen
blockade of TAK1 triggers caspase-8-dependent cleavage of gasdermin
D and cell death. Science. 362:1064–1069. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
98
|
Evavold CL, Ruan J, Tan Y, Xia S, Wu H and
Kagan JC: The pore-forming protein gasdermin D regulates
interleukin-1 secretion from living macrophages. Immunity. 48:35–44
e6. 2018. View Article : Google Scholar
|
|
99
|
Mu M, Yu Q, Zhang Q, Guo J, Wang X, Sun X
and Yu J: A pan-cancer analysis of molecular characteristics and
oncogenic role of gasdermins. Cancer Cell Int. 22:802022.
View Article : Google Scholar : PubMed/NCBI
|
|
100
|
Zheng Y, Yuan D, Zhang F and Tang R: A
systematic pan-cancer analysis of the gasdermin (GSDM) family of
genes and their correlation with prognosis, the tumor
microenvironment, and drug sensitivity. Front Genet. 13:9267962022.
View Article : Google Scholar : PubMed/NCBI
|
|
101
|
Huo CL, Deng Y and Sun ZG: A comprehensive
analysis of gasdermin family gene as therapeutic targets in
pan-cancer. Sci Rep. 12:133292022. View Article : Google Scholar : PubMed/NCBI
|
|
102
|
Liu Y, Fang Y, Chen X, Wang Z, Liang X,
Zhang T, Liu M, Zhou N, Lv J, Tang K, et al: Gasdermin E-mediated
target cell pyroptosis by CAR T cells triggers cytokine release
syndrome. Sci Immunol. 5:eaax79692020. View Article : Google Scholar : PubMed/NCBI
|
|
103
|
Xia X, Wang X, Cheng Z, Qin W, Lei L,
Jiang J and Hu J: The role of pyroptosis in cancer: Pro-cancer or
pro-'host'? Cell Death Dis. 10:6502019. View Article : Google Scholar
|
|
104
|
Karki R and Kanneganti TD: Diverging
inflammasome signals in tumorigenesis and potential targeting. Nat
Rev Cancer. 19:197–214. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
105
|
Ruan J, Xia S, Liu X, Lieberman J and Wu
H: Cryo-EM structure of the gasdermin A3 membrane pore. Nature.
557:62–67. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
106
|
Xia S, Zhang Z, Magupalli VG, Pablo JL,
Dong Y, Vora SM, Wang L, Fu TM, Jacobson MP, Greka A, et al:
Gasdermin D pore structure reveals preferential release of mature
interleukin-1. Nature. 593:607–611. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
107
|
Liu Z, Wang C, Yang J, Zhou B, Yang R,
Ramachandran R, Abbott DW and Xiao TS: Crystal structures of the
full-length murine and human gasdermin D reveal mechanisms of
autoinhibition, lipid binding, and oligomerization. Immunity.
51:43–49 e4. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
108
|
Johnson DC, Taabazuing CY, Okondo MC, Chui
AJ, Rao SD, Brown FC, Reed C, Peguero E, de Stanchina E and Kentsis
A and Bachovchin DA: DPP8/DPP9 inhibitor-induced pyroptosis for
treatment of acute myeloid leukemia. Nat Med. 24:1151–1156. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
109
|
Johnson DC, Okondo MC, Orth EL, Rao SD,
Huang HC, Ball DP and Bachovchin DA: DPP8/9 inhibitors activate the
CARD8 inflammasome in resting lymphocytes. Cell Death Dis.
11:6282020. View Article : Google Scholar : PubMed/NCBI
|
|
110
|
Li F, Aljahdali I and Ling X: Cancer
therapeutics using survivin BIRC5 as a target: what can we do after
over two decades of study? J Exp Clin Cancer Res. 38:3682019.
View Article : Google Scholar : PubMed/NCBI
|
|
111
|
Yue E, Tuguzbaeva G, Chen X, Qin Y, Li A,
Sun X, Dong C, Liu Y, Yu Y, Zahra SM, et al: Anthocyanin is
involved in the activation of pyroptosis in oral squamous cell
carcinoma. Phytomedicine. 56:286–294. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
112
|
Zhang CC, Li CG, Wang YF, Xu LH, He XH,
Zeng QZ, Zeng CY, Mai FY, Hu B and Ouyang DY: Chemotherapeutic
paclitaxel and cisplatin differentially induce pyroptosis in A549
lung cancer cells via caspase-3/GSDME activation. Apoptosis.
24:312–325. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
113
|
Yu P, Wang HY, Tian M, Li AX, Chen XS,
Wang XL, Zhang Y and Cheng Y: Eukaryotic elongation factor-2 kinase
regulates the cross-talk between autophagy and pyroptosis in
doxorubicin-treated human melanoma cells in vitro. Acta Pharmacol
Sin. 40:1237–1244. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
114
|
Yu J, Li S, Qi J, Chen Z, Wu Y, Guo J,
Wang K, Sun X and Zheng J: Cleavage of GSDME by caspase-3
determines lobaplatin-induced pyroptosis in colon cancer cells.
Cell Death Dis. 10:1932019. View Article : Google Scholar : PubMed/NCBI
|
|
115
|
Rathkey JK, Zhao J, Liu Z, Chen Y, Yang J,
Kondolf HC, Benson BL, Chirieleison SM, Huang AY, Dubyak GR, et al:
Chemical disruption of the pyroptotic pore-forming protein
gasdermin D inhibits inflammatory cell death and sepsis. Sci
Immunol. 3:eaat27382018. View Article : Google Scholar : PubMed/NCBI
|
|
116
|
Zhou Y, Zhang Y, Wang H, Zhang X, Chen Y
and Chen G: Microglial pyroptosis in hippocampus mediates
sevolfurane-induced cognitive impairment in aged mice via ROS-NLRP3
inf lammasome pathway. Int Immunopharmacol. 116:1097252023.
View Article : Google Scholar
|
|
117
|
Hu JJ, Liu X, Xia S, Zhang Z, Zhang Y,
Zhao J, Ruan J, Luo X, Lou X, Bai Y, et al: FDA-approved disulfiram
inhibits pyroptosis by blocking gasdermin D pore formation. Nat
Immunol. 21:736–745. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
118
|
Li J, Zhuang L, Luo X, Liang J, Sun E and
He Y: Protection of MCC950 against Alzheimer's disease via
inhibiting neuronal pyroptosis in SAMP8 mice. Exp Brain Res.
238:2603–2614. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
119
|
Tian D, Xing Y, Gao W, Zhang H, Song Y,
Tian Y and Dai Z: Sevoflurane aggravates the progress of
Alzheimer's disease through NLRP3/Caspase-1/Gasdermin D pathway.
Front Cell Dev Biol. 9:8014222022. View Article : Google Scholar : PubMed/NCBI
|
|
120
|
Cai W, Wu Z, Lai J, Yao J, Zeng Y, Fang Z,
Lin W, Chen J, Xu C and Chen X: LDC7559 inhibits microglial
activation and GSDMD-dependent pyroptosis after subarachnoid
hemorrhage. Front Immunol. 14:11173102023. View Article : Google Scholar : PubMed/NCBI
|
|
121
|
Yu E, Zhang E, Lv X, Yan L, Lin Z,
Siaw-Debrah F, Zhang Y, Yang S, Ruan L, ZhuGe Q and Ni H: LDC7559
Exerts neuroprotective effects by inhibiting GSDMD-Dependent
pyroptosis of microglia in mice with traumatic brain injury. J
Neurotrauma. 40:742–757. 2023. View Article : Google Scholar
|