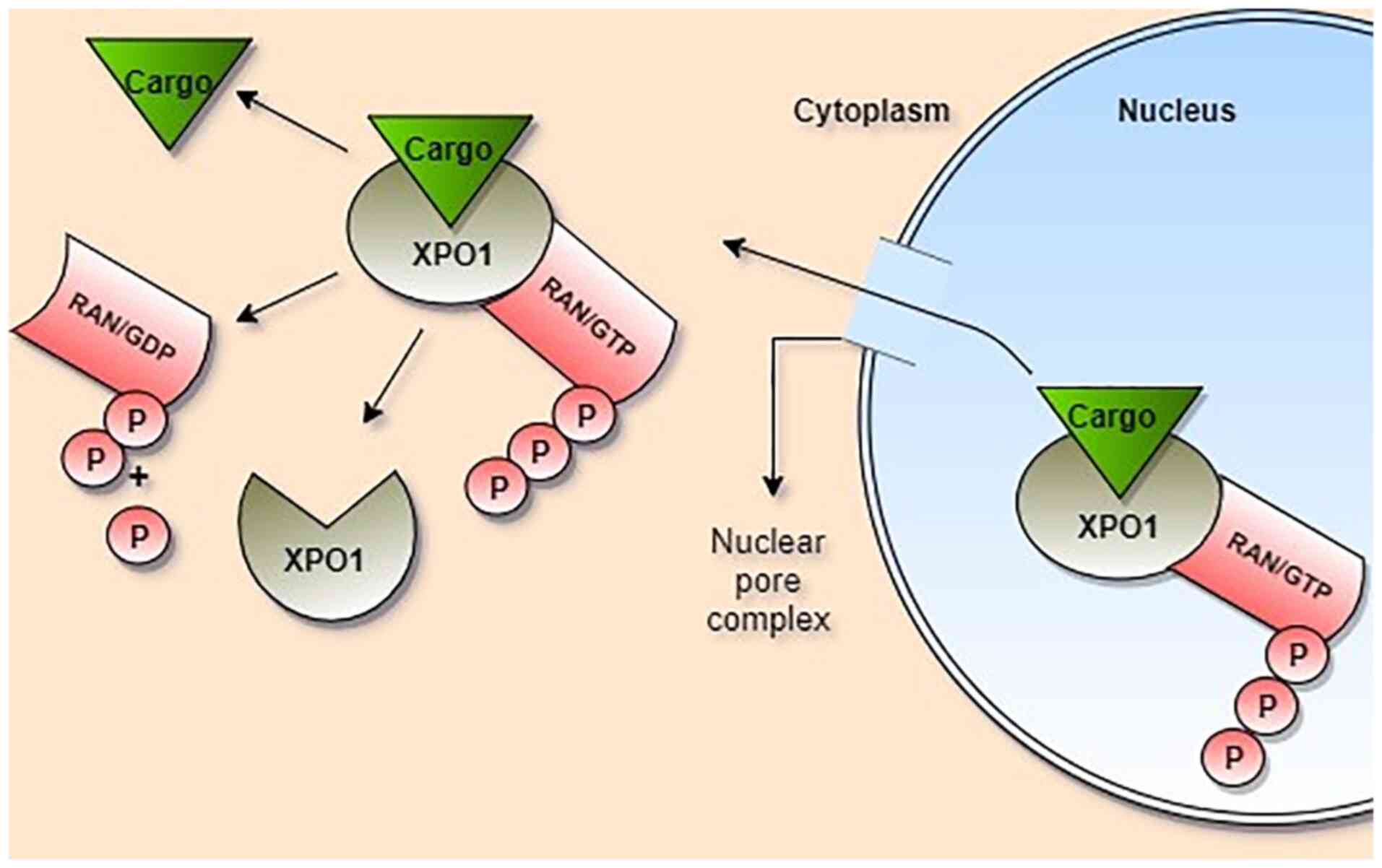
1. Nucleo-cytoplasmic transport
Nuclear pore complexes (NPCs) regulate the entry and
exit of molecules from the cell nucleus. Small molecules (≤30 kDa)
pass through NPCs by diffusion. However, larger molecules, such as
RNA and proteins, enter and exit the cell nucleus via proteins
called karyopherins, which are transport factors (1). Karyopherins (nuclear-cytoplasmic
transport receptors family) denote a family of receptors associated
with transport of molecules between the cytoplasm and nucleus of
eukaryotic cells and comprise >19 members (including importins,
exportins and transportins) (1,2). They
are called karyopherins because they are found inside the nucleus
in the karyoplasm (or nucleoplasm). Importins transport the cargo
molecule from the cytoplasm to the nucleus, while exportins
transport the cargo molecule from the nucleus to the cytosol.
Moreover, transportins can transport molecules both from the
nucleus to the cytoplasm and from the cytoplasm to the nucleus
(3,4).
Karyopherins recognise and select the cargo molecule
from its target sequences and transport it across the nuclear
membrane (1,2). The nuclear target sequence is a short
amino acid sequence, which enables the cargo molecule to be
recognised by carrier karyopherins for entry and exit to the cell
nucleus; it also determines the direction of transport (5). The nuclear target sequence recognised
by importins when transporting the cargo molecule from the
cytoplasm to the nucleus is referred to as the nuclear localization
signal (NLS). The NLS sequence commonly comprises hydrophilic amino
acids (especially lysine) (2,3). The
target sequence that is recognized by the exportins when
transporting the cargo molecule from the nucleus to the cytoplasm
is called the nuclear export signal (NES) (2-4,6).
The NES sequence is a short peptide of 10-15 amino acids with 3-4
regular leucine-rich hydrophobic amino acid repeats (generally
conserved sequence Φ1-X2,3-Φ2-X2,3-Φ3-X-Φ4,
where Φ is leucine, valine, isoleucine, phenylalanine or methionine
and X is any amino acid) (7).
Various mechanisms, such as post-translational modification
(phosphorylation, acetylation, sumoylation), mutation or protein
interaction, lead to changes in the intracellular location of the
cargo molecule via the creation of new NES/NLS sequences, thus
concealing the target sequence or changing the affinity of
karyopherins to the cargo molecule (8,9).
The transfer of molecules larger than 30 kDa from
NPCs, which cannot pass through the NPC by diffusion, is an
energy-dependent process, and the function of importins and
exportins is mediated by Ras-related nuclear protein (Ran). To
perform their function, importins bind to Ran/GDP, whereas
exportins bind to Ran/GTP (10).
Exportins creates a cargo-exportin-Ran/GTP ternary complex in the
nucleus by facilitating binding between cargo molecules and RanGTP.
As this complex passes from the NPC to the cytosol, Ran/GTP is
hydrolysed, Ran/GDP is formed and exportin releases the cargo
molecule in the cytoplasm and then returns to the nucleus (11,12). By
contrast, importins bind to RanGTP, releasing the cargo protein
(2).
2. Structure of XPO1
In the human genome, exportins are encoded by six
genes, one of the primary nuclear exporters is the Exportin-1
(XPO1) gene, also known as CRM1 (Ensembl no. ENSG000000828987). The
XPO1 protein functions as a shuttle as a nuclear transport receptor
between the cell nucleus and the cytoplasm (13,14). The
XPO1 gene was discovered by Adachi and Yanagida in Saccharomyces
cerevisiae and Schizosaccharomyces pombe cells in 1989.
Mutations in the XPO1 gene first observed in mutant yeast strains
deform the chromosome structure and the XPO1 gene is associated
with preservation of chromosome structure. Therefore, the gene
product has been named chromosome region maintenance 1 (CRM1)
(15). The S. pombe XPO1
protein is homologous to the human protein originally named CC112.
Given this homology, it became referred to as XPO1 instead of
CC112(16). The XPO1 gene is
localized in the 2p16 region of the second chromosome in humans
(17). It consists of 60,778 bases
organized into 25 exons and separated by 24 introns. This gene has
24 transcript variants. The XPO1 gene product is a 112 kDa protein.
This mature polypeptide consists of 1,071 amino acids (13); it is a modular protein consisting of
numerous functional domains and mediates the transport of ~220
proteins (18). Given that the
N-terminal region of XPO1 protein (UniProtKB no. O149803) interacts
with GTPase/Ran, it is believed that the affinity of its C-terminal
region (707-1,034 residues) with cargo molecules is increased
(19).
The C-terminal region (amino acids 707-1,027) of the
CRM1 protein is composed of two U-shaped antiparallel α-helices and
the HEAT repeat 15A-21A crystal structure was reported in 2004. In
the Protein Data Bank, 26 data show the crystal structure of XPO1
alone or its interaction complexes with different proteins
(20). Various methods, such as
X-ray crystallography and electron microscopy, have revealed that
XPO1 displays conformational flexibility as a transporter protein
(21). They also shed light on the
X-ray structure of the XPO1-RanGTP-Snurportin 1 and
XPO1-RanGTP-RanBP1 (Ran binding protein 1) complexes. HEAT 9
interacts with the NES sequence of cargo proteins, serving as an
allosteric inhibitor and controlling formation of the complex
(22,23).
3. Physiological function of XPO1 in
cells
XPO1 demonstrates weak binding with RanGTP in the
nucleus and with the NES sequence of cargo molecules (24) However, when XPO1 simultaneously binds
to RanGTP and to cargo molecules, its affinity for both increases
by 500-1,000 times, and it passes from the NPC to the cytosol by
forming a ternary complex (25). The
hydrolysis of Ran/GTP to Ran/GDP in the cytoplasm decreases the
affinity of XPO1 to cargo molecules by causing conformational
changes in the protein structure, facilitating the release of cargo
molecules (10-12)
(Fig. 1). XPO1 and RanGDP pass
through the NPC and return to the nucleus for a new transport cycle
(2,25).
The human XPO1 gene is expressed in a cell
cycle-dependent manner; mRNA transcription begins during the
G1 phase of the cell cycle and increases during the
G2/M phase (26). During
interphase, the XPO1 protein is localized inside the nucleus near
the nuclear membrane (26,27). High XPO1 expression levels have been
observed in the brain, thymus, lung, spleen, liver, heart,
pancreas, skeletal muscle, prostate, testis, placenta, ovary, small
intestine, colon and peripheral blood leukocytes (14,19).
XPO1 mediates the transport of certain types of RNA,
including viral/cellular mRNA, ribosomal RNA, transfer RNA and
small nuclear RNAs (snRNAs), as well as various macromolecules,
such as ribosomal subunit and NES-containing proteins rich in
leucine, short peptide stretches containing hydrophobic residues,
shuttle proteins, tumour suppressor proteins (TSPs), cellular or
non-spliced or incomplete spliced RNAs of various viruses [such as
human immunodeficiency virus (HIV)-1, human T-lymphotropic virus
type-1 (HTLV-1) and influenza A] and HIV-1 Rev protein and HTLV-1
Rex protein that interact with Ran/GTP in the nucleus and cytosol
(4,19,23,28-30).
4. XPO1 export of protein
In the proteomic analysis of yeast cells, 285
proteins regulated by XPO1 were detected, with ~45% of these
containing known NES sequences (31). NES sequences for XPO1 comprise
hydrophobic amino acids, including isoleucine, leucine, methionine,
valine and phenylalanine (32). The
NES sequences have a common conserved motif containing 10-15 amino
acids with hydrophobic character [HX2-3HX2-3HXH, where H is a
hydrophobic amino acid (such as isoleucine, leucine, methionine,
valine and phenylalanine), X is any amino acid and the number is
the potential number of repeats] (33). These hydrophobic amino acids form an
α-helix or the entire loop structure, thus allowing XPO1 to attach
to the hydrophobic pocket (25).
However, NES sequences are yet to be sufficiently defined, as
evidenced by the fact that <40% are known (7). The change in the cargo molecule's
three-dimensional structure by mutation, phosphorylation and
dephosphorylation results in the formation of a new NES to which
XPO1 binds (8,34,35). It
also causes NES sequences to be lost or masked or leads to the
emergence of new sequences by additional modifications, including
ubiquitination (33-35),
acetylation (34), 10 sumoylation
(9) or binding protein to specific
cofactors such as RanGTP. Therefore, it alters the affinity of XPO1
to the cargo molecule (36,37).
Nuclear export of proteins, including STAT, NF-κB,
nucleophosmin (NPM)1, Ras association domain family member SF2,
Merlin, TSPs, such as APC, p53, p73, forkhead box O (FOXO), IKB,
BCR-ABL, eukaryotic translation initiation factor 4E, BRCA1, viral
proteins, regulatory/pro-inflammatory p21CIP, p27Kip1,
retinoblastoma, anti-apoptotic proteins, such as NPM and AP-1 and
oncogenic proteins, such as Cox-2, c-MYC, epidermal growth factor
receptor and hypoxia inducible factor-1, is key for cell cycle and
cell proliferation. In addition, their nuclear export is mediated
by XPO1(38). Furthermore, XPO1
contributes to carcinogenesis by regulating the activity of TSPs
and oncogenes. XPO1 controls multiple intracellular processes by
regulating the localization of cyclin B, MPAK, MAPK-associated
protein kinase 2, p21, p33, p27 and Survivin (39,40).
Moreover, the overexpression of XPO1 leads to transport of Rev and
U snRNA from the nucleus to the cytoplasm (13). It has been reported that the
transcription factors nuclear transcription factor Y/CBP
(CREB-binding protein), Sp1 and p53 are associated with the
promoter of the XPO1 gene and serve an important role in the
transformation of cancer cells by activating the promoter of
XPO1(41). The XPO1 gene plays a
role in the control of cell proliferation and affects the loss of
control in cancer cell proliferation via various pathways. As a
nuclear export factor, XPO1 regulates direct intracellular
localization of cell cycle regulators, TSPs and pro-apoptotic
proteins; therefore, the displacement of these proteins containing
nuclear export sequences contributes to carcinogenesis and the
development of drug-resistance mechanisms by regulating the
activity of oncogenes.
5. Single nucleotide polymorphisms (SNPs) of
XPO1
Genotypical variations in XPO1 may affect the
function of the nuclear-cytoplasmic transport by altering
expression levels of XPO1, resulting in emergence of certain
disease phenotypes. The SNP c.1871A> G in XPO1 (pD624G) has been
found to be associated with oesophageal squamous cell carcinoma and
chronic lymphocytic leukaemia (42,43).
Structural modelling study have shown that it is necessary to
create a critical salt bridge with lysine at position 144 in the
area where aspartic acid amino acid located at 624 of the XPO1
protein attaches to the target cargo molecule of XPO1. It has been
reported that the conversion of the amino acid at the position 624
to glycine may cause the loss of this salt bridge and increase the
nuclear-cytoplasmic transport efficiency by changing the affinity
of XPO1 to the cargo molecule (42).
SNP rs6735330 in XPO1 is associated with autism (44). In addition, SNP rs7600515 in XPO1 has
been reported as a prognostic factor for Crohn's disease (45). SNP rs4430924 in XPO1 is also
associated with the risk of developing hepatotoxicity of
anti-tuberculosis drugs (46).
6. XPO1 in cancer
XPO1 is reported to be overexpressed in certain
types of cancer (Table I).
 | Table ICancer types associated with exportin
1. |
Table I
Cancer types associated with exportin
1.
| Cell line | Study design | Cancer | (Refs.) |
|---|
| OVCAR-3, SKOV-3,
CAOV-3, ES-2, A2780, Mdah2744, OAW42, EFO21, EFO27, FU-OV-1,
PA-1 | In
vitro | Ovarian | (47) |
| HeLa (HPV18), SiHa
(HPV16), CaSki (HPV16 and 18), ME180, MS751 (HPV18), C33A | In
vitro | Cervical | (48) |
| U251, SHG-44 U118,
U87 | In vitro and
in vivo | Glioma | (49) |
| U2OS, HOS, Saos2,
MG63 | In
vitro | Osteosarcoma | (50) |
| MiaPaCa-2, HPAC,
AsPC-1, PANC-1 | In vitro and
in vivo | Pancreatic | (51) |
| WHCO1, WHCO5, WHCO,
ECA109, TE1, TE8, KYSE306 | In vitro and
in vivo | Esophageal | (52,53) |
| A549, H460 | In
vitro | Lung | (54) |
| NCI-N87, SNU-1,
SNU-16 | In vitro and
in vivo | Gastric | (55) |
| UT-SCC-16A,
UT-SCC-16B, UT-SCC-60A, UT-SCC-60B, UT-SCC-74, UT-SCC-74B, | In
vitro | Head and neck | (56,57) |
| e ACHN, Caki-1,
786-O | In vitro and
in vivo | Renal cell
carcinoma | (58) |
| SK-HEP-1, Huh7,
MHCC97H, SNU-182, SNU-387, HepG2 | In vitro and
in vivo | Hepatocellular
carcinoma | (59) |
| Plat-E, 293,
OCI-AML-3, MOLM-13, and MV4 | In
vitro | Acute
myeloid/lymphoid leukemia (AML/ALL) | (60,61) |
| HS-5 | In vitro and
in vivo | Chronic
myeloid/lymphoid leukemia | (62) |
| HS-5 | In vitro and
in vivo | Multiple
myeloma | (63) |
| JVM-2, Maver-1,
NCEB-1, Jeko-1, DBsp53, Granta519, JVM-13, Z-138, Rec-1 | In vitro and
in vivo | Mantle cell
lymphoma | (64,65) |
| MM1S, MM1R, INA6,
INA6GFP, ANBL6 KMS18, KMS20 | In vitro and
in vivo | Plasma cell
leukemia | (66) |
| RPMI8226, MOLP8,
28BM, 12PE, XG1, U266 | | | |
| K562 | In
vitro | Acute myeloid
leukemia | (67) |
In ovarian (47),
cervical (48), glioma (49), osteosarcoma (50), pancreatic (51), oesophageal (52,53),
lung (54), gastric (55), head and neck (56,57),
renal cell carcinoma (58),
hepatocellular carcinoma (59),
acute lymphoid leukaemia (ALL) (60,61),
chronic myeloid/lymphoid leukaemia (CML/CLL), multiple myeloma (MM)
(62), mantle cell lymphoma (MCL)
(63,64) plasma cell leukaemia (65), acute myeloid leukaemia (AML)
(66) and breast cancer (67), an increase in the expression level of
XPO1 was detected and it has been reported that this increase was
associated with metastasis, increased tumour size, histological
grade and decreased overall survival. The increased expression of
XPO1 causes accumulation or mislocalisation of TSPs, cell cycle
regulator and/or pro-apoptotic proteins in the cytoplasm with
excessive nucleo-cytoplasmic transport, as well as deregulating
ribosomal biogenesis, increasing carcinogenesis and development of
resistance to chemotherapy (10). In
addition, Crm1-dependent pathways serve a role in cancer
pathogenesis since it is active in the control of mitosis and the
dispersion of chromosomes and is important in maintaining and
chromosome protected structure (68). One study reported that XPO1
overexpression in all solid tumour types except liver cancer and
haematological malignancies (69).
Higher XPO1 expression has been associated with worse patient
prognosis in ovarian, pancreatic, oesophageal, thymic epithelial
and breast tumour, as well as glioma. On the other hand, high XPO1
expression has been associated with a better prognosis in patients
with osteosarcoma. Given XPO1 overexpression and its association
with negative clinical outcome in various types of malignancy, it
has become an attractive potential therapeutic target molecule in
oncology (47,49-51,64).
7. Inhibitors of XPO1
It has been reported that suppressing XPO1-mediated
nuclear export with specific agents or suppressing gene expression
by XPO1-specific small interfering RNA activates apoptotic pathways
and increases the sensitivity of tumour cells to chemotherapy
drugs, such as doxorubicin, etoposide (70), cisplatin (71) and imatinib mesylate (72). Table
II summarizes compounds that have been described as XPO1
inhibitors.
| Table IIExportin 1 nuclear export
inhibitors. |
Table II
Exportin 1 nuclear export
inhibitors.
| Inhibitor | Compound | (Refs.) |
|---|
| Leptomycin B | Antibiotic | (14,74-76) |
| Anguinomycins | Antibiotic | (77) |
| Goniothalamin | Natural | (78,79) |
| 15d-PGJ2 | Natural | (80) |
| Plumbagin | Natural | (81,82) |
| Piperlongumine | Natural | (83,84) |
| Ratjadon | Antibiotic | (85,86,87) |
| Curcumin | Natural | (90,91) |
| SINE | Synthetic | (92-100) |
|
(R)-4'-methylklavuzon | Synthetic | (101) |
| KOS-2464 | Synthetic | (33,59) |
| CBS9106 | Synthetic | (33) |
| Compound S109 | Synthetic | (84,85,102) |
| PKF050-638 | Synthetic | (33,73,103) |
| Compound 1l | Synthetic | (104) |
Numerous inhibitors of nuclear export derived from
natural and synthetic sources have been identified (73). Leptomycin B (LMB), a natural
compound, emerged as the first inhibitory molecule to block the
function of the XPO1 protein (14,74). LMB
is covalently bound at the specific cysteine (Cys528) residue
located in the NES-binding cleft of the XPO1 protein; this inhibits
binding of XPO1 to the target cargo molecule (74). LMB has been examined in multiple
cancer cell lines and murine xenograft tumour models (75). Phase-I clinical LMB studies are
continuing to investigate its use as an anti-cancer agent (70,75).
However, LMB has not yet been used in clinical practice due to its
low therapeutic index and high toxicity (76).
Other natural inhibitors of XPO1 include
anguinomycins, isolated from Streptomyces species with
selective cytotoxicity to transformed cells (77), and goniothalamin, obtained from
Goniothalamus macrophyllus. Goniothalamin has been reported
to induce G2/M phase cell cycle arrest and apoptosis in
breast cancer cells (78,79). Moreover, 15d-PGJ2, a prostaglandin
family member with both anti- and pro-inflammatory properties
(80) and plumbagin, derived from
Plumbago zeylanica, have been shown to have a suppressive
role in XPO1 nuclear export function. In the presence of plumbagin,
interactions between XPO1 and Foxo1, p21, p53 and p73 are disrupted
(81). In particular, plumbagin
serves an anti-tumour effect via suppression of nuclear export
(82). Furthermore, piperlongumine,
an alkaloid of the long pepper, induces nuclear retention of major
TSPs including Foxo1, p53, p21 and IкB-α and blocks interactions
between XPO1 and these proteins (83,84). The
natural inhibitor ratjadon, isolated from Sorangium
cellulosum, has anti-proliferative effects (85). In a previous study, ratjadon exerted
anti-HIV activity (86). In one
study, ratjadon was conjugated to small-molecule targeted ligands,
which induced the inhibition of nuclear export in extracellular
targeted cancer therapy. These conjugates retain their inhibitory
activity by binding to XPO1(87).
These aforementioned natural inhibitors with
molecular mechanisms similar to LMB bind covalently to Cys528 in
the reactive XPO1 region. XPO1-mediated nuclear export is inhibited
by such binding, resulting in nuclear accumulation of TSPs and
growth regulatory proteins (GRPs) (88,89).
Contrary to the other natural XPO1 inhibitors,
curcumin, a natural polyphenol product, also suppresses p53 nuclear
export (90). Curcumin has various
anti-inflammatory, antimicrobial, antioxidative and anti-cancer
properties. Phase I/II studies of curcumin in multiple myeloma (MM)
are ongoing (91).
Besides natural inhibitors, several synthetic
compounds are also available to inhibit XPO1, including selective
inhibitors of nuclear export (SINEs), KOS-2464,
(R)-4'-methylklavuzon, CBS9106, Compound S109, PKF050-638 and
Compound 1l. SINE compounds are produced based on an in
silico molecular modelling strategy (82-95).
SINEs are covalently bound to Cys528 residue of XPO1 and hinder
XPO1 binding to target cargo molecules (92). In in vivo studies, oral use of
SINEs was observed to cause mild gastrointestinal symptoms
(93). The potential of SINEs to
inhibit XPO1-mediated nuclear export has been demonstrated.
Apoptosis of cancer cells and arrest of cells in the G1
phase of the cell cycle are induced by SINEs (62,93). The
effectiveness of KPT-330 (selinexor), SINE compound, has been
evaluated in clinical studies of solid tumours and haematological
malignancy. The results of these clinical trials have shown it to
be a promising therapeutic candidate (10,94).
KPT-330 has been found to have positive effects in clinical studies
of hematopoietic malignancy, such as MM, AML and non-Hodgkin
lymphoma (NHL). KPT-330 has been subjected to phase I/II studies in
patients with AML (92,95). Notable decreases in tumour size have
been demonstrated in a preclinical animal study with KPT-330. It
has been demonstrated that KPT-330 has high efficacy in combination
with various standard therapies, including selinexor/doxorubicin
and selinexor/dexamethasone combinations (96,97).
Moreover, a decrease in XPO1 levels was observed in studies
conducted with other SINEs, including KPT-185 and KPT-251(98). Thus, the nuclear localisation of
tumour suppressors may be preserved. Therefore, preclinical studies
of XPO1 inhibition using SINE compounds may lead to a novel
treatment for various types of cancer, including breast cancer
(18,97,99). In
addition, activity of KPT-185 and KPT-276 was investigated in NHL
using in vitro and in vivo study. A high level of
anti-tumour activity was observed in mouse models in which KPT-276
was orally applied. Therefore, KPT-276 is a promising candidate for
NHL treatment (100).
Another synthetic XPO1 inhibitor is
(R)-4'-methylklavuzon, which can retain tumour suppressor proteins
in the nucleus by inhibiting the XPO1 protein.
(R)-4'-methylklavuzon has been shown to be a novel drug candidate
for treating hepatocellular carcinoma (101). KOS-2464, another synthetic
molecule, is the most effective LMB analogue and has been reported
to induce apoptosis at low nanomolar concentrations. Low toxicity
and high anti-tumour activity of KOS-2464 have been demonstrated in
various cancer cell lines and xenograft mouse models (33,59).
CBS9106 binds to the XPO1 reactive site, causing
degradation, and its anti-tumor activity has been demonstrated in
in vitro in various cancer cell lines and in vivo in
xenograft animal models (33).
Compound S109 is a derivative of CBS9106 that causes cell cycle
arrest of large TSPs mediated by XPO1. Its anti-tumour activity has
been investigated in colorectal and kidney cancer cells and it has
been proven to inhibit proliferation and induce cell cycle arrest
in these cells (84,85,102).
Furthermore, PKF050-638 is a XPO1-inhibitor used in HIV treatment
to inhibit the nuclear export of HIV-1 Rev protein; however, its
anti-cancer effect has not been investigated yet (33,103).
PKF050-638 interacts with cysteine in the NES-binding groove and
prevents binding of the NES, similar to the mechanism of LMB
(73).
(R)-6-[(2-isopropyl-5-methylphenoxy) methyl]-5,6-dihydro-2-Pyron
(compound 11) is a colourless oily liquid. The cytotoxic effects of
compound 1l in HGC27 and MGC803 gastric cancer cell lines have been
investigated; compound 1l was reported to degrade XPO1, inducing
apoptosis in both MGC803 and HGC27 cell lines, exhibiting strong
cytotoxic and anti-tumour effects against these cells (104).
8. Conclusion and future directions
Studies have demonstrated the key role of XPO1 in
carcinogenesis and its potential as a therapeutic target (10,69). It
has thus become the focus of efforts to develop new tumour
treatment strategies. The clinical use of novel specific XPO1
inhibitors and their combination with other agents is promising.
Suppressing gene expression with specific inhibitors or
interference techniques have identified the biological function and
intracellular mechanisms of XPO1 in malignancy, as well as drug
resistance. Targeting XPO1 offers advantages in treatment
strategies by activating various apoptotic pathways to avoid the
development of drug resistance.
Acknowledgements
Not applicable.
Funding
Funding: No funding was received.
Availability of data and materials
Not applicable.
Authors' contributions
SÖ and İC contributed to acquisition and
interpretation of data and drafted and wrote the manuscript. Data
authentication is not applicable. All authors have read and
approved the final version of the manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Authors' information
Sibel Özdaş, ORCID no. 0000-0003-4610-2785; İpek
Canatar, ORCID no. 0000-0001-9448-8112.
Competing interests
The authors declare they have no competing
interests.
References
|
1
|
Wente SR: Gatekeepers of the nucleus.
Science. 288:1374–1377. 2000.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Watson J, Baker T, Bell S, Gann A, Levine
M and Losick R: Molecular Biology of the Gene. 5th ed.
Pearson/Benjamin Cummings: San Francisco Cold Spring Harbor, NY,
912 p. 2004.
|
|
3
|
Izaurralde E and Adam S: Transport of
macromolecules between the nucleus and the cytoplasm. RNA.
4:351–364. 1998.PubMed/NCBI
|
|
4
|
Misteli T: Physiological importance of RNA
and protein mobility in the cell nucleus. Histochem Cell Biol.
129:5–11. 2008.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Chook YM and Blobel G: Karyopherins and
nuclear import. Curr Opin Struct Biol. 11:703–715. 2001.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Alberts B, Johnson A, Lewis J, Raff M,
Roberts K and Walter P: Drosophila and the Molecular Genetics of
Pattern Formation: Genesis of the Body Plan. 4th edition. Garland
Science, New York, NY, 2002.
|
|
7
|
La Cour T, Kiemer L, Mølgaard A, Gupta R,
Skriver K and Brunak S: Analysis and prediction of leucine-rich
nuclear export signals. Protein Eng Des Sel. 17:527–536.
2004.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Powell SM, Zilz N, Beazer-Barclay Y, Bryan
TM, Hamilton SR, Thibodeau SN, Vogelstein B and Kinzler KW: APC
mutations occur early during colorectal tumorigenesis. Nature.
359:235–237. 1992.PubMed/NCBI View
Article : Google Scholar
|
|
9
|
Pichler A and Melchior F:
Ubiquitin-related modifier SUMO1 and nucleocytoplasmic transport.
Traffic. 3:381–387. 2002.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Azmi S, Uddin MH and Mohammad RM: The
nuclear export protein XPO1 - from biology to targeted therapy. Nat
Rev Clin Oncol. 18:152–169. 2021.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Pemberton LF and Paschal BM: Mechanisms of
receptor-mediated nuclear import and nuclear export. Traffic.
6:187–198. 2005.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Kau TR, Way JC and Silver PA: Nuclear
transport and cancer: From mechanism to intervention. Nat Rev
Cancer. 4:106–117. 2004.PubMed/NCBI View
Article : Google Scholar
|
|
13
|
Fornerod M, Ohno M, Yoshida M and Mattaj
IW: CRM1 is an export receptor for leucine-rich nuclear export
signals. Cell. 90:1051–1060. 1997.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Kudo N, Khochbin S, Nishi K, Kitano K,
Yanagida M, Yoshida M and Horinouchi S: Molecular cloning and cell
cycle-dependent expression of mammalian CRM1, a protein involved in
nuclear export of proteins. J Biol Chem. 272:29742–29751.
1997.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Adachi Y and Yanagida M: Higher order
chromosome structure is affected by cold-sensitive mutations in a
Schizosaccharomyces pombe gene crm1+ which encodes a 115-kD protein
preferentially localized in the nucleus and its periphery. J Cell
Biol. 108:1195–1207. 1989.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Ruggiero A, Giubettini M and Lavia P: XPO1
(exportin 1 (CRM1 homolog, yeast)). Atlas of Genetics and
Cytogenetics in Oncology and Haematology. INIST-CNRS 16: 2011.
|
|
17
|
Fornerod M, van Baal S, Valentine V,
Shapiro DN and Grosveld G: Chromosomal localization of genes
encoding CAN/Nup214-interacting proteins-human CRM1 localizes to
2p16, whereas Nup88 localizes to 17p13 and is physically linked to
SF2p32. Genomics. 42:538–540. 1997.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Parikh K, Cang S, Sekhri A and Liu D:
Selective inhibitors of nuclear export (SINE)-a novel class of
anti-cancer agents. J Hematol Oncol. 7(78)2014.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Fornerod M, Van Deursen J, Van Baal S,
Reynolds A, Davis D, Murti KG, Fransen J and Grosveld G: The human
homologue of yeast CRM1 is in a dynamic subcomplex with CAN/Nup214
and a novel nuclear pore component Nup88. EMBO J. 16:807–816.
1997.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Petosa C, Schoehn G, Askjaer P, Bauer U,
Moulin M, Steuerwald U, Soler-Lopez M, Baudin F, Mattaj IW and
Müller CW: Architecture of CRM1/Exportin1 suggests how
cooperativity is achieved during formation of a nuclear export
complex. Mol Cell. 16:761–775. 2004.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Dölker N, Blanchet CE, Voss B, Haselbach
D, Kappel C, Monecke T, Svergun DI, Stark H, Ficner R, Zachariae U,
et al: Structural determinants and mechanism of mammalian CRM1
allostery. Structure. 21:1350–1360. 2013.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Saito N and Matsuura YA: 2.1-Å-resolution
crystal structure of unliganded CRM1 reveals the mechanism of
autoinhibition. J Mol Biol. 425:350–364. 2013.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Koyama M and Matsuura Y: An allosteric
mechanism to displace nuclear export cargo from CRM1 and RanGTP by
RanBP1. EMBO J. 29:2002–1013. 2010.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Lee SH and Hannink M: The N-terminal
nuclear export sequence of IkappaBalpha is required for
RanGTP-dependent binding to CRM1. J Biol Chem. 276:23599–23606.
2001.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Dong X, Biswas A, Süel KE, Jackson LK,
Martinez R, Gu H and Chook YM: Structural basis for leucine-rich
nuclear export signal recognition by CRM1. Nature. 458:1136–1141.
2009.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Ptasznik A, Nakat Y, Kalota A, Emerson SG
and Gewirtz AM: Short interfering RNA (siRNA) targeting the Lyn
kinase induces apoptosis in primary, and drug-resistant,
BCR-ABL1(+) leukemia cells. Nat Med. 10:1187–1189. 2004.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Grunewald TG, Kammerer U, Schulze E,
Schindler D, Honig A, Zimmer M and Butt E: Silencing of LASP-1
influences zyxin localization, inhibits proliferation and reduces
migration in breast cancer cells. Exp Cell Res. 312:974–982.
2006.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Fukuda M, Asano S, Nakamura T, Adachi M,
Yoshida M, Yanagida M and Nishida E: CRM1 is responsible for
intracellular transport mediated by the nuclear export signal.
Nature. 390:308–311. 1997.PubMed/NCBI View
Article : Google Scholar
|
|
29
|
Ossareh-Nazari B, Bachelerie F and
Dargemont C: Evidence for a role of CRM1 in signal-mediated nuclear
protein export. Science. 278:141–144. 1997.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Fei E, Ma X, Zhu C, Xue T, Yan J, Xu Y,
Zhou J and Wang G: Nucleocytoplasmic shuttling of dysbindin-1, a
schizophrenia-related protein, regulates synapsin I expression. J
Biol Chem. 285:38630–38640. 2010.PubMed/NCBI View Article : Google Scholar
|
|
31
|
Matsuyama A, Arai R, Yashiroda Y, Shirai
A, Kamata A, Sekido S, Kobyashi Y, Hashimoto A, Hamamoto M, Hiraoka
Y, et al: ORFeome cloning and global analysis of protein
localization in the fission yeast Schizosaccharomyces pombe. Nat
Biotechnol. 24:841–847. 2006.PubMed/NCBI View Article : Google Scholar
|
|
32
|
Kutay U and Güttinger S: Leucine-rich
nuclear-export signals: Born to be weak. Trends Cell Biol.
15:121–124. 2005.PubMed/NCBI View Article : Google Scholar
|
|
33
|
Turner JG, Dawson J and Sullivan DM:
Nuclear export of proteins and drug resistance in cancer. Biochem
Pharmacol. 83:1021–1032. 2012.PubMed/NCBI View Article : Google Scholar
|
|
34
|
Vogt PK, Jiang H and Aoki M: Triple layer
control: Phosphorylation, acetylation and ubiquitination of FOXO
proteins. Cell Cycle. 4:908–913. 2005.PubMed/NCBI View Article : Google Scholar
|
|
35
|
Craig E, Zhang ZK, Davies KP and Kalpana
GV: A masked NES in INI1/hSNF5 mediates hCRM1-dependent nuclear
export: Implications for tumorigenesis. EMBO J. 21:31–42.
2002.PubMed/NCBI View Article : Google Scholar
|
|
36
|
Yoneda Y, Hieda M, Nagoshi E and Miyamoto
Y: Nucleocytoplasmic protein transport and recycling of Ran. Cell
Struct Funct. 24:425–333. 1999.PubMed/NCBI View Article : Google Scholar
|
|
37
|
Poon IK and Jans DA: Regulation of nuclear
transport: Central role in development and transformation? Traffic.
6:173–186. 2005.PubMed/NCBI View Article : Google Scholar
|
|
38
|
Gandhi UH, Senapedis W, Baloglu E, Unger
TJ, Chari A, Vogl D and Cornell RF: Clinical implications of
targeting XPO1-mediated nuclear export in multiple myeloma. Clin
Lymphoma Myeloma Leuk. 18:335–345. 2018.PubMed/NCBI View Article : Google Scholar
|
|
39
|
Fabbro M and Henderson BR: Regulation of
tumor suppressors by nuclear-cytoplasmic shuttling. Exp Cell Res.
282:59–69. 2003.PubMed/NCBI View Article : Google Scholar
|
|
40
|
Rensen WM, Mangiacasale R, Ciciarello M
and Lavia P: The GTPase Ran: Regulation of cell life and potential
roles in cell transformation. Front Biosci. 13:4097–4121.
2008.PubMed/NCBI View
Article : Google Scholar
|
|
41
|
van der Watt PJ and Leaner VD: The nuclear
exporter, Crm1, is regulated by NFY and Sp1 in cancer cells and
repressed by p53 in response to DNA damage. Biochim Biophys Acta.
1809:316–326. 2011.PubMed/NCBI View Article : Google Scholar
|
|
42
|
Lin DC, Hao JJ, Nagata Y, Xu L, Shang L,
Meng X, Sato Y, Okuno Y, Varela AM, Ding LW, et al: Genomic and
molecular characterization of esophageal squamous cell carcinoma.
Nat Genet. 46:467–473. 2014.PubMed/NCBI View Article : Google Scholar
|
|
43
|
Landau DA, Carter SL, Stojanov P, McKenna
A, Stevenson K, Lawrence MS, Sougnez C, Stewart C, Sivachenko A,
Wang L, et al: Evolution and impact of subclonal mutations in
chronic lymphocytic leukemia. Cell. 152:714–726. 2013.PubMed/NCBI View Article : Google Scholar
|
|
44
|
Liu X, Malenfant P, Reesor C, Lee A,
Hudson ML, Harvard C, Qiao Y, Persico AM, Cohen IL, Chudley AE, et
al: 2p15-p16. 1 microdeletion syndrome: Molecular characterization
and association of the OTX1 and XPO1 genes with autism spectrum
disorders. Eur J Hum Genet. 19:1264–1270. 2011.PubMed/NCBI View Article : Google Scholar
|
|
45
|
Pernat Drobež C, Repnik K, Gorenjak M,
Ferkolj I, Weersma RK and Potocnik U: DNA polymorphisms predict
time to progression from uncomplicated to complicated Crohn's
disease. Eur J Gastroenterol Hepatol. 30:447–455. 2018.PubMed/NCBI View Article : Google Scholar
|
|
46
|
He X, Zhang H, Tao B, Yang M, Chen H, Lu
L, Yi H, Pan H and Tang S: The A/A genotype of XPO1 rs4430924 is
associated with higher risk of antituberculosis drug-induced
hepatotoxicity in Chinese patients. J Clin Pharmacol. 59:1014–1021.
2019.PubMed/NCBI View Article : Google Scholar
|
|
47
|
Noske A, Weichert W, Niesporek S, Röske A,
Buckendahl AC, Koch I, Sehouli J, Dietel M and Denkert C:
Expression of the nuclear export protein chromosomal region
maintenance/exportin 1/Xpo1 is a prognostic factor in human ovarian
cancer. Cancer. 112:1733–1743. 2008.PubMed/NCBI View Article : Google Scholar
|
|
48
|
Van der Watt PJ, Maske CP, Hendricks DT,
Parker MI, Denny L, Govender D, Birrer MJ and Leaner VD: The
Karyopherin proteins, Crm1 and Karyopherin β1, are overexpressed in
cervical cancer and are critical for cancer cell survival and
proliferation. Int J Cancer. 124:1829–1840. 2009.PubMed/NCBI View Article : Google Scholar
|
|
49
|
Liu X, Chong Y, Tu Y, Liu N, Yue C, Qi Z,
Liu H, Yao Y, Liu H, Gao S, et al: CRM1/XPO1 is associated with
clinical outcome in glioma and represents a therapeutic target by
perturbing multiple core pathways. J Hematol Oncol.
9(108)2016.PubMed/NCBI View Article : Google Scholar
|
|
50
|
Jiang Y, Hou J, Zhang X, Xu G, Wang Y,
Shen L, Wu Y, Li Y and Yao L: Circ-XPO1 upregulates XPO1 expression
by sponging multiple miRNAs to facilitate osteosarcoma cell
progression. Exp Mol Pathol. 117(104553)2020.PubMed/NCBI View Article : Google Scholar
|
|
51
|
Azmi AS, Li Y, Muqbil I, Aboukameel A,
Senapedis W, Baloglu E, Landesman Y, Shacham S, Kauffman MG, Philip
PA and Mohammad RM: Exportin 1 (XPO1) inhibition leads to
restoration of tumor suppressor miR-145 and consequent suppression
of pancreatic cancer cell proliferation and migration. Oncotarget.
8:82144–82155. 2017.PubMed/NCBI View Article : Google Scholar
|
|
52
|
Van der Watt PJ, Zemanay W, Govender D,
Hendricks DT, Parker MI and Leaner VD: Elevated expression of the
nuclear export protein, Crm1 (exportin 1), associates with human
esophageal squamous cell carcinoma. Oncol Rep. 32:730–738.
2014.PubMed/NCBI View Article : Google Scholar
|
|
53
|
Yang X, Cheng L, Yao L, Ren H, Zhang S,
Min X, Chen X, Zhang J and Li M: Involvement of chromosome region
maintenance 1 (CRM1) in the formation and progression of esophageal
squamous cell carcinoma. Med Oncol. 31(155)2014.PubMed/NCBI View Article : Google Scholar
|
|
54
|
Liu Z and Ga W: Leptomycin B reduces
primary and acquired resistance of gefitinib in lung cancer cells.
Toxicol Appl Pharmacol. 335:16–27. 2017.PubMed/NCBI View Article : Google Scholar
|
|
55
|
Sexton R, Mahdi Z, Chaudhury R, Beydoun R,
Aboukameel A, Khan HY, Baloglu E, Senapedis W, Landesman Y, Tesfaye
A, et al: Targeting nuclear exporter protein XPO1/CRM1 in gastric
cancer. Int J Mol Sci. 20(4826)2019.PubMed/NCBI View Article : Google Scholar
|
|
56
|
Özdaş S and Özdaş T: Crm1 knockdown by
specific small interfering RNA reduces cell proliferation and
induces apoptosis in head and neck cancer cell lines. Turk J Biol.
42:132–143. 2018.PubMed/NCBI View Article : Google Scholar
|
|
57
|
Özdaş S: Nuclear entrapment of p33ING1b by
inhibition of exportin-1: A trigger of apoptosis in head and neck
squamous cell cancer. Cell Mol Biol (Noisy-le-grand). 64:66–72.
2018.PubMed/NCBI
|
|
58
|
Inoue H, Kauffman M, Shacham S, Landesman
Y, Yang J, Evans CP and Weiss RH: CRM1 blockade by selective
inhibitors of nuclear export attenuates kidney cancer growth. J
Urol. 189:2317–2326. 2013.PubMed/NCBI View Article : Google Scholar
|
|
59
|
Zheng Y, Gery S, Sun H, Shacham S,
Kauffman M and Koeffler HP: KPT-330 inhibitor of XPO1-mediated
nuclear export has anti-proliferative activity in hepatocellular
carcinoma. Cancer Chemother Pharmacol. 74:487–495. 2014.PubMed/NCBI View Article : Google Scholar
|
|
60
|
Conway AE, Haldeman JM, Wechsler DS and
Lava CP: A critical role for CRM1 in regulating HOXA gene
transcription in CALM-AF10 leukemias. Leukemia. 29:423–432.
2015.PubMed/NCBI View Article : Google Scholar
|
|
61
|
Kojima K, Kornblau SM, Ruvolo V, Dilip A,
Duvvuri S, Davis RE, Zhang M, Wang Z, Coombes KR, Zhang N, et al:
Prognostic impact and targeting of CRM1 in acute myeloid leukemia.
Blood. 121:4166–4174. 2013.PubMed/NCBI View Article : Google Scholar
|
|
62
|
Lapalombella R, Sun Q, Williams K,
Tangeman L, Jha S, Zhong Y, Goettl V, Mahoney E, Berglund C, Gupta
S, et al: Selective inhibitors of nuclear export show that
CRM1/XPO1 is a target in chronic lymphocytic leukemia. Blood.
120:4621–4634. 2012.PubMed/NCBI View Article : Google Scholar
|
|
63
|
Zhang K, Wang M, Tamayo AT, Shacham S,
Kauffman M, Lee J, Zhang L, Ou Z, Li C, Sun L, et al: Novel
selective inhibitors of nuclear export CRM1 antagonists for therapy
in mantle cell lymphoma. Exp Hematol. 41:67–78.e4. 2013.PubMed/NCBI View Article : Google Scholar
|
|
64
|
Yoshimura M, Ishizawa J, Ruvolo V, Dilip
A, Quintás-Cardama A, McDonnell TJ, Neelapu SS, Kwak LW, Shacham S,
Kauffman M, et al: Induction of p53-mediated transcription and
apoptosis by exportin-1 (XPO 1) inhibition in mantle cell lymphoma.
Cancer Sci. 105:795–801. 2014.PubMed/NCBI View Article : Google Scholar
|
|
65
|
Tai YT, Landesman Y, Acharya C, Calle Y,
Zhong MY, Cea M, Tannenbaum D, Cagnetta A, Reagan M, Munshi AA, et
al: CRM1 inhibition induces tumor cell cytotoxicity and impairs
osteoclastogenesis in multiple myeloma: Molecular mechanisms and
therapeutic implications. Leukemia. 28:155–165. 2014.PubMed/NCBI View Article : Google Scholar
|
|
66
|
Wang AY, Weiner H, Green M, Chang H,
Fulton N, Larson RA, Odenike O, Artz AS, Bishop MR, Godley LA, et
al: A phase I study of selinexor in combination with high-dose
cytarabine and mitoxantrone for remission induction in patients
with acute myeloid leukemia. J Hematol Oncol. 11(4)2018.PubMed/NCBI View Article : Google Scholar
|
|
67
|
Yue L, Sun ZN, Yao YS, Shen Z, Wang HB,
Liu XP, Zhou F, Xiang JY, Yao RY and Niu HT: CRM1, a novel
independent prognostic factor overexpressed in invasive breast
carcinoma of poor prognosis. Oncol Lett. 15:7515–7522.
2018.PubMed/NCBI View Article : Google Scholar
|
|
68
|
Dasso M: Ran at kinetochores. Biochem Soc
Trans. 34:711–715. 2006.PubMed/NCBI View Article : Google Scholar
|
|
69
|
Sendino M, Omaetxebarria MJ and Rodríguez
JA: Hitting a moving target: Inhibition of the nuclear export
receptor XPO1/CRM1 as a therapeutic approach in cancer. Cancer Drug
Resistance. 1:139–163. 2018.
|
|
70
|
Turner JG, Marchion DC, Dawson JL, Emmons
MF, Hazlehurst LA, Washausen P and Sullivan DM: Human multiple
myeloma cells are sensitized to topoisomerase II inhibitors by CRM1
inhibition. Cancer Res. 69:6899–6905. 2009.PubMed/NCBI View Article : Google Scholar
|
|
71
|
Takenaka Y, Fukumori T, Yoshii T, Oka N,
Inohara H, Kim HR, Bresalier RS and Raz A: Nuclear export of
phosphorylated galectin-3 regulates its antiapoptotic activity in
response to chemotherapeutic drugs. Mol Cell Biol. 24:4395–4406.
2004.PubMed/NCBI View Article : Google Scholar
|
|
72
|
Aloisi A, Di Gregorio S, Stagno F,
Guglielmo P, Mannino F, Sormani MP, Bruzzi P, Gambacorti-Passerini
C, Saglio G, Venuta S, et al: BCR-ABL nuclear entrapment kills
human CML cells: Ex vivo study on 35 patients with the combination
of imatinib mesylate and leptomycin B. Blood. 107:1591–1598.
2006.PubMed/NCBI View Article : Google Scholar
|
|
73
|
Dickmanns A, Monecke T and Ficner R:
Structural basis of targeting the exportin CRM1 in cancer. Cells.
4:538–568. 2015.PubMed/NCBI View Article : Google Scholar
|
|
74
|
Kudo N, Matsumori N, Taoka H, Fujiwara D,
Schreiner EP, Wolff B, Yoshida M and Horinouchi S: Leptomycin B
inactivates CRM1/exportin 1 by covalent modification at a cysteine
residue in the central conserved region. Proc Natl Acad Sci USA.
96:9112–9117. 1999.PubMed/NCBI View Article : Google Scholar
|
|
75
|
Mutka SC, Yang WQ, Dong SD, Ward SL, Craig
DA, Timmermans PB and Murli S: Identification of nuclear export
inhibitors with potent anticancer activity in vivo. Cancer Res.
69:510–517. 2009.PubMed/NCBI View Article : Google Scholar
|
|
76
|
Newlands ES, Rustin GJ and Brampton MH:
Phase I trial of elactocin. Br J Cancer. 74:648–649.
1996.PubMed/NCBI View Article : Google Scholar
|
|
77
|
Hayakawa Y, Sohda KY, Shin-Ya K, Hidaka T
and Seto H: Anguinomycins C and D, new antitumor antibiotics with
selective cytotoxicity against transformed cells. J Antibiot
(Tokyo). 48:954–961. 1995.PubMed/NCBI View Article : Google Scholar
|
|
78
|
Wach JY, Güttinger S, Kutay U and Gademann
K: The cytotoxic styryl lactone goniothalamin is an inhibitor of
nucleocytoplasmic transport. Bioorg Med Chem Lett. 20:2843–2846.
2010.PubMed/NCBI View Article : Google Scholar
|
|
79
|
Sophonnithiprasert T, Mahabusarakam W,
Nakamura Y and Watanapokasin R: Goniothalamin induces
mitochondria-mediated apoptosis associated with endoplasmic
reticulum stress-induced activation of JNK in HeLa cells. Oncol
Lett. 13:119–128. 2017.PubMed/NCBI View Article : Google Scholar
|
|
80
|
Hilliard M, Frohnert C, Spillner C,
Marcone S, Nath A, Lampe T, Fitzgerald DJ and Kehlenbach RH: The
anti-inflammatory prostaglandin 15-deoxy-delta(12,14)-PGJ2 inhibits
CRM1-dependent nuclear protein export. J Biol Chem.
285:22202–22210. 2010.PubMed/NCBI View Article : Google Scholar
|
|
81
|
Muqbil I, Azmi AS and Mohammad RM: Nuclear
export inhibition for pancreatic cancer therapy. Cancers (Basel).
10(138)2018.PubMed/NCBI View Article : Google Scholar
|
|
82
|
Liu X, Niu M, Xu X, Cai W, Zeng L, Zhou X,
Yu R and Xu K: CRM1 is a direct cellular target of the natural
anti-cancer agent plumbagin. J Pharmacol Sci. 124:486–493.
2014.PubMed/NCBI View Article : Google Scholar
|
|
83
|
Niu M, Chong Y, Han Y and Liu X: Novel
reversible selective inhibitor of nuclear export shows that CRM1 is
a target in colorectal cancer cells. Cancer Biol Ther.
16:1110–1118. 2015.PubMed/NCBI View Article : Google Scholar
|
|
84
|
Niu M, Xu X, Shen Y, Yao Y, Qiao J, Zhu F,
Zeng L, Liu X and Xu K: Piperlongumine is a novel nuclear export
inhibitor with potent anticancer activity. Chem Biol Interact.
237:66–72. 2015.PubMed/NCBI View Article : Google Scholar
|
|
85
|
Meissner T, Krause E and Vinkemeier U:
Ratjadone and leptomycin B block CRM1-dependent nuclear export by
identical mechanisms. FEBS Lett. 576:27–30. 2004.PubMed/NCBI View Article : Google Scholar
|
|
86
|
Fleta-Soriano E, Martinez JP, Hinkelmann
B, Gerth K, Washausen P, Diez J, Frank R, Sasse F and Meyerhans A:
The myxobacterial metabolite ratjadone A inhibits HIV infection by
blocking the Rev/CRM1-mediated nuclear export pathway. Microb Cell
Fact. 13(17)2014.PubMed/NCBI View Article : Google Scholar
|
|
87
|
Klahn P, Fetz V, Ritter A, Collisi W,
Hinkelmann B, Arnold T, Tegge W, Rox K, Hüttel S, Mohr KI, et al:
The nuclear export inhibitor aminoratjadone is a potent effector in
extracellular-targeted drug conjugates. Chem Sci. 10:5197–5210.
2019.PubMed/NCBI View Article : Google Scholar
|
|
88
|
Camus V, Miloudi H, Taly A, Sola B and
Jardin F: XPO1 in B cell hematological malignancies: From recurrent
somatic mutations to targeted therapy. J Hematol Oncol.
10(47)2017.PubMed/NCBI View Article : Google Scholar
|
|
89
|
Azmi AS, Muqbil I, Wu J, Aboukameel A,
Senapedis W, Baloglu E, Bollig-Fischer A, Dyson G, Kauffman M,
Landesman Y, et al: Targeting the nuclear export protein XPO1/CRM1
reverses epithelial to mesenchymal transition. Sci Rep.
5(16077)2015.PubMed/NCBI View Article : Google Scholar
|
|
90
|
Theodoropoulos N, Lancman G and Chari A:
Targeting nuclear export proteins in multiple myeloma therapy.
Target Oncol. 15:697–708. 2020.PubMed/NCBI View Article : Google Scholar
|
|
91
|
Niu M, Wu S, Mao L and Yang Y: CRM1 is a
cellular target of curcumin: New insights for the myriad of
biological effects of an ancient spice. Traffic. 14:1042–1052.
2013.PubMed/NCBI View Article : Google Scholar
|
|
92
|
Etchin J, Montero J, Berezovskaya A, Le
BT, Kentsis A, Christie AL, Conway AS, Chen WC, Reed C, Mansour MR,
et al: Activity of a selective inhibitor of nuclear export,
selinexor (KPT-330), against AML-initiating cells engrafted into
immunosuppressed NSG mice. Leukemia. 30:190–199. 2016.PubMed/NCBI View Article : Google Scholar
|
|
93
|
Etchin J, Sanda T, Mansour MR, Kentsis A,
Montero J, Le BT, Christie AL, McCauley D, Rodig SJ, Kauffman M, et
al: KPT-330 inhibitor of CRM 1 (XPO 1)-mediated nuclear export has
selective anti-leukaemic activity in preclinical models of T-cell
acute lymphoblastic leukaemia and acute myeloid leukaemia. Br J
Haematol. 161:117–127. 2013.PubMed/NCBI View Article : Google Scholar
|
|
94
|
Subhash VV, Yeo MS, Wang L, Tan SH, Wong
FY, Thuya WL, Tan WL, Peethala PC, Soe MY, Tan DSP, et al:
Anti-tumor efficacy of Selinexor (KPT-330) in gastric cancer is
dependent on nuclear accumulation of p53 tumor suppressor. Sci Rep.
8(12248)2018.PubMed/NCBI View Article : Google Scholar
|
|
95
|
Gravina GL, Senapedis W, McCauley D,
Baloglu E, Shacham S and Festuccia C: Nucleo-cytoplasmic transport
as a therapeutic target of cancer. J Hematol Oncol.
7(85)2014.PubMed/NCBI View Article : Google Scholar
|
|
96
|
Azizian NG and Li Y: XPO1-dependent
nuclear export as a target for cancer therapy. J Hematol Oncol.
13(61)2020.PubMed/NCBI View Article : Google Scholar
|
|
97
|
Lewin J, Malone E, Al-Ezzi E, Fasih S,
Pedersen P, Accardi S, Gupta A and Abdul Razak A: A phase 1b trial
of selinexor, a first-in-class selective inhibitor of nuclear
export (SINE), in combination with doxorubicin in patients with
advanced soft tissue sarcomas (STS). Eur J Cancer. 144:360–367.
2021.PubMed/NCBI View Article : Google Scholar
|
|
98
|
Mendonca J, Sharma A, Kim HS, Hammers H,
Meeker A, De Marzo A, Carducci M, Kauffman M, Shacham S and Kachhap
S: Selective inhibitors of nuclear export (SINE) as novel
therapeutics for prostate cancer. Oncotarget. 5:6102–6112.
2014.PubMed/NCBI View Article : Google Scholar
|
|
99
|
Uddin MH, Zonder JA and Azmi AS: Exportin
1 inhibition as antiviral therapy. Drug Discov Today. 25:1775–1781.
2020.PubMed/NCBI View Article : Google Scholar
|
|
100
|
Han X, Wang J, Shen Y, Zhang N, Wang S,
Yao J and Shi Y: CRM1 as a new therapeutic target for non-Hodgkin
lymphoma. Leuk Res. 39:38–46. 2015.PubMed/NCBI View Article : Google Scholar
|
|
101
|
Delman M, Avcı ST, Akçok İ, Kanbur T,
Erdal E and Çağır A: Antiproliferative activity of
(R)-4'-methylklavuzon on hepatocellular carcinoma cells and
EpCAM+/CD133+ cancer stem cells via SIRT1 and
Exportin-1 (CRM1) inhibition. Eur J Med Chem. 180:224–237.
2019.PubMed/NCBI View Article : Google Scholar
|
|
102
|
Liu X, Chong Y, Liu H, Han Y and Niu M:
CRM1 inhibitor S109 suppresses cell proliferation and induces cell
cycle arrest in renal cancer cells. Korean J Physiol Pharmacol.
20:161–168. 2016.PubMed/NCBI View Article : Google Scholar
|
|
103
|
Daelemans D, Afonina E, Nilsson J, Werner
G, Kjems J, De Clercq E, Pavlakis GN and Vandamme AM: A synthetic
HIV-1 Rev inhibitor interfering with the CRM1-mediated nuclear
export. Proc Natl Acad Sci USA. 99:14440–14445. 2002.PubMed/NCBI View Article : Google Scholar
|
|
104
|
Xu HW, Jia S, Liu M, Li X, Meng X, Wu X,
Yu L, Wang M and Jin CY: A low toxic CRM1 degrader: Synthesis and
anti-proliferation on MGC803 and HGC27. Eur J Med Chem.
206(112708)2020.PubMed/NCBI View Article : Google Scholar
|