Introduction
The progressive wasting of skeletal muscle tissue
plays a crucial pathophysiological role in frailty, fragility and
loss of independence in aged people (1). In the majority of humans, skeletal
muscle mass and contractile strength decline during aging (2), making sarcopenia a striking feature
of the senescent organism (3). A
sedentary life style clearly promotes muscle degeneration in
elderly patients (4). Functional
impairments of the neuromuscular system are believed to be
multi-factorial, but it is not clear whether a pathobiochemical
hierarchy exists within these aberrant pathways (5,6). The
exact molecular mechanisms underlying primary muscle degeneration,
secondary effects, impaired repair cycles and physiological
adaptations of aging muscle are still elusive. Although
considerable inter-individual differences exist in elderly people
with respect to the degree of muscle wasting (7,8), the
age-related impairment of innervation and concomitant
excitation-contraction uncoupling appears to be an intractable
consequence of advanced age (9).
Although it is encouraging that a protein-rich diet
and regular physical activity can at least partially diminish
severe fibre degeneration (10–12),
the symptoms of sarcopenia may be aggravated by unrelated
pathologies. This includes cancer, traumatic injury, type 2
diabetes, metabolic syndrome, renal failure, chronic obstructive
pulmonary disease and congestive heart failure (13–15).
Thus, the clinical prevalence of sarcopenia in the aging population
requires detailed physiological, biochemical and genetic
investigations into the molecular pathogenesis of age-related
muscle degeneration. In the present study, we used
electrophoresis-based proteomics to study potential changes in
muscle protein abundance in an established animal model of
sarcopenia, the aged Wistar rat (16). Over the last few years, a variety
of proteomic approaches have determined changes in animal and human
muscle proteins during aging (17). This includes the mass spectrometric
(MS) identification of altered protein nitration (18), protein phosphorylation (19), protein carbonylation (20) and protein glycosylation (21), as well as changes in the abundance
of muscle proteins using crude tissue extracts (22–27)
and subcellular fractions, such as myofibrils (28), integral membrane proteins (29) and mitochondria (30).
Modifications in mitochondria have previously been
implicated in the natural aging process (31), including cellular senescence in
contractile tissues (32).
Functional changes and altered expression patterns of mitochondrial
proteins in aged skeletal muscle have been confirmed by proteomic
investigations, as reviewed by Staunton et al (33). However, a potential problem with
the gel electrophoretic separation of the urea-soluble portion of
the skeletal muscle proteome is that the presence of proteins that
exhibit an isoelectric point in the basic range is often
underestimated (34). To address
this technical problem, we carried out an offgel electrophoretic
pre-fractionation step prior to standard 2D gel electrophoresis.
The offgel approach fractionates protein populations in solution by
isoelectric focusing (IEF) (35)
and can therefore be employed to separate proteins with small
differences in their isoelectric point (36). Offgel electrophoresis has been
successfully applied to studying extraocular muscle specimens
(37) and was used here for a
comparative subproteomic study of young adult versus aged rat
muscle. Our proteomic survey has focused on potential changes in
proteins with a basic isoelectric point and showed an increase in
mitochondrial creatine kinase (CK) and ubiquinol cytochrome-c
reductase and a decrease in glyceraldehyde-3-phosphate
dehydrogenase (GAPDH). These findings agree with the idea that
muscle aging is associated with a glycolytic-to-oxidative shift and
that changes in key metabolic enzymes are involved in age-related
muscle weakness.
Materials and methods
Materials
Chemicals and materials for the offgel and 2D gel
electrophoretic analysis of muscle proteins, including IEF pH
gradient strips and ampholytes, were obtained from Agilent
Technologies (Santa Clara, CA, USA) and Amersham Bioscience/GE
Healthcare (Little Chalfont, Bucks, UK). Ultrapure Protogel
acrylamide stock solutions were purchased from National Diagnostics
(Atlanta, GA, USA). For the generation of peptides, sequencing
grade-modified trypsin was obtained from Promega (Madison, WI,
USA). Protease inhibitors, nitrocellulose sheets and
chemiluminescence substrate were from Roche Diagnostics (Manheim,
Germany), Millipore (Bedford, MA, USA) and Pierce and Warriner
(Chester, UK), respectively. For immunoblotting, primary antibodies
were purchased from Abcam (Cambridge, UK), (pAb to GAPDH; and mAb
46A1 to CK) and Upstate Biotechnology (Lake Placid, NY) [mAb SKB1
to protein kinase B (Akt/PKB)]. Secondary peroxidase-conjugated
antibodies were from Chemicon International (Temecula, CA). All
other chemicals were of analytical grade and obtained from Sigma
Chemical Company (Dorset, UK).
Animal model of sarcopenia
The gastrocnemius muscle from young adult versus
aged Wistar rats was used for the subproteomic analysis of aging,
focusing on proteins with an isoelectric point in the basic range.
Aged Wistar rat muscle is an internationally established model
system to study age-dependent changes in sarcopenia (16). Both human and rodent muscles share
many common age-related alterations, including the incomplete
recruitment of distinct fiber groupings, altered proportions
between slow and fast fibers, altered fiber size, tissue
degeneration and contractile weakness (5). This makes rat muscle an ideal tissue
source for detailed proteomic studies of muscle wasting associated
with aging. Freshly dissected muscle specimens from 3- and
30-month-old rats were obtained from the Animal Facility of the
Department of Physiology, Trinity College Dublin, Ireland. For one
set of comparative proteomic analyses, a combination of 6
biological repeats was carried out for each age group. All rats
used in this study were fed ad libitum and kept at a
standard light-dark cycle with unrestricted movement in standard
animal house cages (26). The
activity level of the younger rat population was enhanced as
compared to the older cohort.
Preparation of crude skeletal muscle
extracts for electrophoretic analysis
Fresh muscle tissue (100 mg) was quick-frozen in
liquid nitrogen and ground into a fine powder using a pestle and
mortar. The pulverized muscle tissue was resuspended in 1 ml of
ice-cold buffer (7 M urea, 2 M thiourea, 65 mM Chaps, 10 mM Trizma
base, 1% ampholytes, 100 mM DTT). To avoid protein degradation and
eliminate excessive viscosity of the extract due to DNA, the
solution was supplemented with a protease inhibitor cocktail (Roche
Diagnostics), and 2 μl of DNase-I (200 units, Sigma Chemical
Company) was added per 100 μl buffer (26). The tissue homogenate was mixed by
vortexing and was then placed on a bench top shaker for 2.5 h at
room temperature to precipitate the total protein fraction. The
suspension was centrifuged at 20,000 × g in an Eppendorf 5417R
centrifuge (Eppendorf, Hamburg, Germany) for 20 min. The resulting
protein pellet was washed in 5 ml of ice-cold 100% acetone and
thoroughly broken up by vortexing and sonication. The
centrifugation and washing step was repeated twice more with 80%
acetone and the final protein precipitate was collected by
centrifugation and resuspended in 1 ml of the above described
buffer by gentle pipetting and vortexing. The protein lysate was
then incubated for 1 h at room temperature, whereby samples were
vortexed every 10 min for 5 sec, and then subjected to IEF.
Offgel electrophoresis
The initial separation of the acidic versus the
basic protein cohort from rat gastrocnemius muscle was carried out
with a 3100 OFFGEL Fractionator from Agilent Technologies. Protein
samples were pre-fractionated on 24-well lanes across 24 cm IPG pH
3.0–10.0 strips (35). In order to
enrich the acidic and the basic portion of the muscle sample,
proteins from wells 1–12 (acidic samples) and wells 13–24 (basic
samples) were pooled, respectively, upon completion of IEF. The
Agilent 3100 OFFGEL Fractionator performs IEF of proteins in
immobilized pH gradient strips while the separated proteins can
then be recovered from the liquid phase (36). The offgel electrophoresis procedure
was carried out according to the manufacturer’s guideline using
reagents supplied with the OFFGEL pH 3.0–10.0 kit from Agilent.
Fifteen minutes prior to IEF, IPG strips were incubated with 40 μl
of 1X protein OFFGEL stock solution. Samples were run at 1 mg of
protein sample/lane for our comparative study of young adult versus
aged muscle. The sample protein volume was brought up to 0.72 ml
with ddH2O, and added to 2.88 ml of 1.25X protein OFFGEL
stock solution. A 150-μl aliquot of this solution was then added to
each well. IEF strips were focused in four stages, using 200 V for
90 min, 2,000 V for 90 min, 3,000 V for 90 min and a final 5,000 V
step until the procedure reached 64 kV/h. Total focusing time
depended mainly on the number of strips being focused and could
take up to 72 h when 12 strips were run simultaneously. Every 24 h
the wicks on each strip were removed and replaced with new wicks
wetted with distilled water. Upon completion of IEF, samples from
the two different pH-ranges were removed and pooled. Excess urea,
which was found to interfere with the acetone precipitation of
protein, was removed by repeated dialysis. Samples were acetone
precipitated (26) and resuspended
in 200 μl of rehydration buffer (7 M urea, 2 M thiourea, 65 mM
Chaps, 1% ampholytes pH 3.0 to 10.0 and 10 mg/ml DTT).
Two-dimensional (2D) gel electrophoretic
analysis
In order to analyze pre-fractionated proteins with
an isoelectric point in the basic range, 2D gel electrophoresis was
carried out as previously described in detail by our laboratory
(19,26,28).
Following IEF with an Amersham IPGphor system with the following
running conditions: 30 V for 2 h, 500 V for 2.5 h, 1,000 V for 1 h,
2,000 V for 1 h, 4,000 V for 1 h, 6,000 V for 1 h, 800 V for 3 h,
500 V for 1.5 h and finally 8,000 V for 2.5 h, samples were
equilibrated and washed in running buffer (26), and then separated in the second
dimension with 12% resolving gels in a Protean Xi-ll cell from
Bio-Rad Laboratories (Hemel-Hemstead, Hertz, UK). Sodium dodecyl
sulfate (SDS)-polyacrylamide gel electrophoresis (PAGE) was carried
out for 2 h at 30 V and a subsequent 80 V step until the
bromophenol blue dye front ran off the gel. Silver staining of 2D
gels was carried out by the method of Chevallet et al
(38). For image analysis, 6 gels
representing young adult muscle and 6 gels representing aged muscle
were scanned using Image Scanner II from Amersham Bioscience/GE
Healthcare. Images were analyzed with Progenesis SameSpots analysis
software from Nonlinear Dynamics (Newcastle upon Tyne, UK).
Mass spectrometric (MS) identification of
basic muscle proteins
Protein identification was carried out by MS
analysis of peptides with a Model 6340 Ion Trap LC/MS apparatus
from Agilent Technologies. Excision, washing, destaining and
treatment with sequencing-grade trypsin were performed using an
optimized method as previously described by our laboratory
(28,30). Trypsin-generated peptides were
resuspended in 30 μl of ultrapure ddH2O and 0.1% formic
acid for ion trap LC-MS analysis. Separation of peptides was
performed with a Nanoflow Agilent 1200 series system (Agilent
Technologies), equipped with a Zorbax 300SB C18 5 μm, 4 mm 40 nl
pre-column and an Zorbax 300SB C18 5 μm, 43 mm × 75 μm analytical
reversed phase column using HPLC-Chip technology. Mobile phases
utilized were A, 0.1% formic acid; B, 50% acetonitrile and 0.1%
formic acid. Samples (15 μl) were loaded into the enrichment at a
capillary flow rate set to 4 μl/min with a mix of A and B at a
ratio 19:1. Tryptic peptides were eluted with a linear gradient of
10–90% solvent B over 40 min with a constant nano pump flow of 0.60
ml/min. A 10 min post time of solvent A was used to remove sample
carryover. The capillary voltage was set to 1,700 V. The flow and
the temperature of the drying gas was 4 l/min and 300˚C,
respectively. For muscle protein identification, database searches
were carried out with Mascot MS/MS Ion search (Matrix Science,
London, UK). All searches used ‘Rattus norvegicus’ as
taxonomic category and the following parameters: i) two missed
cleavages by trypsin, ii) mass tolerance of precursor ions ± 2.5 Da
and product ions ± 0.7 Da, iii) carboxymethylated cysteine fixed
modification and iv) oxidation of methionine as variable
modification. In addition, the percentage coverage was set at over
10%, with at least 2 matched distinct peptide and a Mascot score of
over 60.
Immunoblot analysis
For the verification of changes in select marker
proteins, immunoblotting was used to determine potential changes in
proteins in young adult versus aged muscle preparations (26). The electrophoretic transfer of
proteins to Immobilon NC-Pure nitrocellulose membranes was carried
out with a Mini-Protean II transfer system from Bio-Rad
Laboratories (Hemel-Hempstead, Herts, UK). Following transfer for 1
h at 100 V and 4˚C, the efficiency of transfer was evaluated by
Ponceau-S-Red staining of membranes, followed by de-staining in
phosphate-buffered saline (PBS, 50 mM sodium phosphate, 0.9% (w/v)
NaCl, pH 7.4). Protein-containing nitrocellulose membranes were
blocked for 1 h in 5% (w/v) fat-free milk powder in PBS and
incubated for 3 h at room temperature with appropriately diluted
primary antibodies to GAPDH, CK or Akt/PKB. Following washing in
blocking solution, sheets were incubated with an appropriate
dilution of peroxidase-conjugated secondary antibody for 1 h at
room temperature, washed again in blocking solution and PBS, and
finally immuno-decorated protein bands were visualized using the
SuperSignal-Type enhanced chemiluminecence system from Pierce and
Warriner (Chester, Cheshire, UK). Densitometric scanning of
immunoblots was performed on a Molecular Dynamics 300S computing
densitometer (Sunnyvale, CA, USA) with Imagequant V3.0
software.
Results
Certain muscle proteins with an isoelectric point in
the basic range are underestimated in gel electrophoresis-based
proteomic studies. Therefore, to address this technical issue, we
performed a comparative subproteomic investigation of young adult
versus aged rat muscle focusing on potential changes in muscle
proteins with an alkaline isoelectric point. The combination of
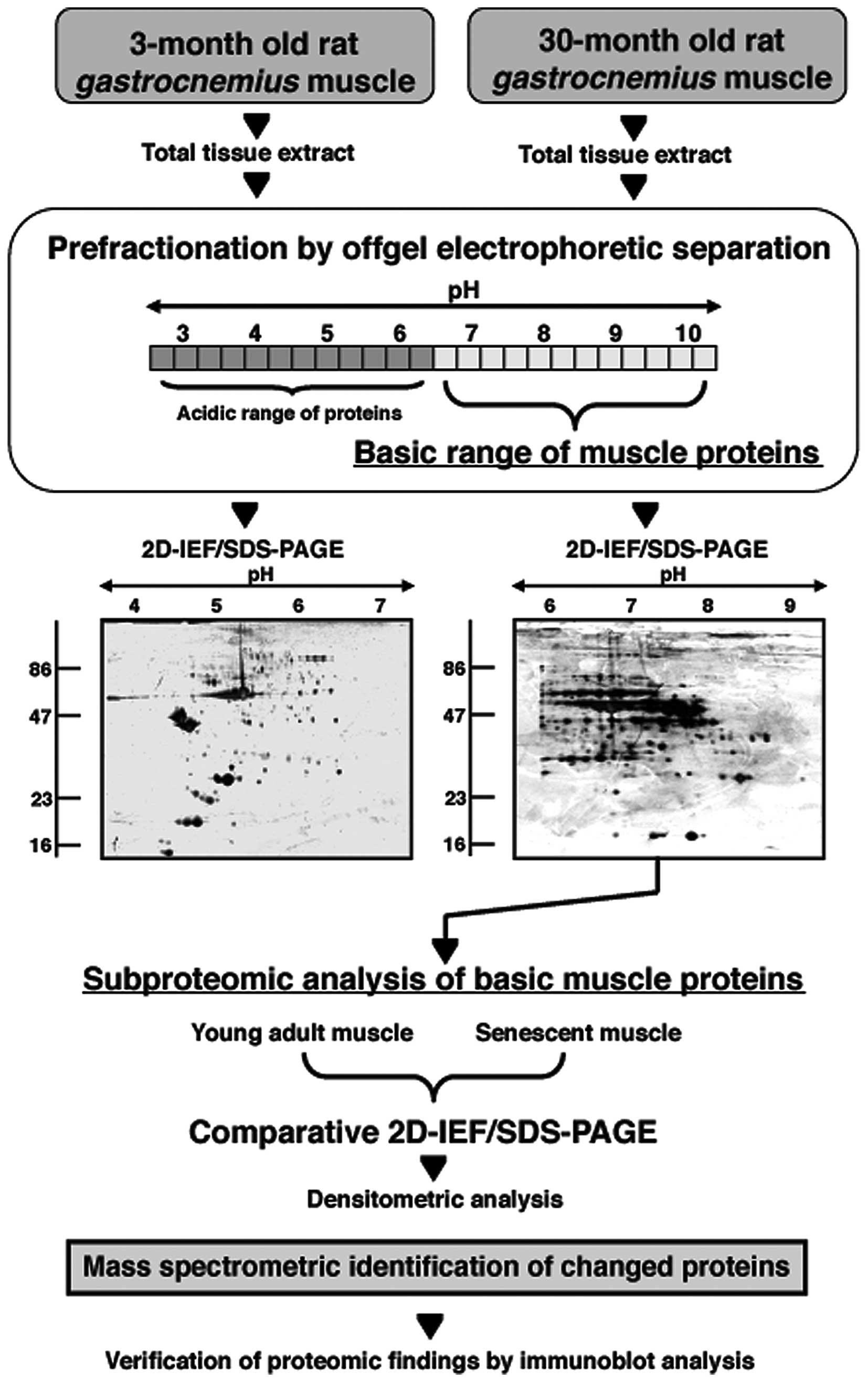
offgel electrophoresis and 2D gel electrophoresis is outlined in
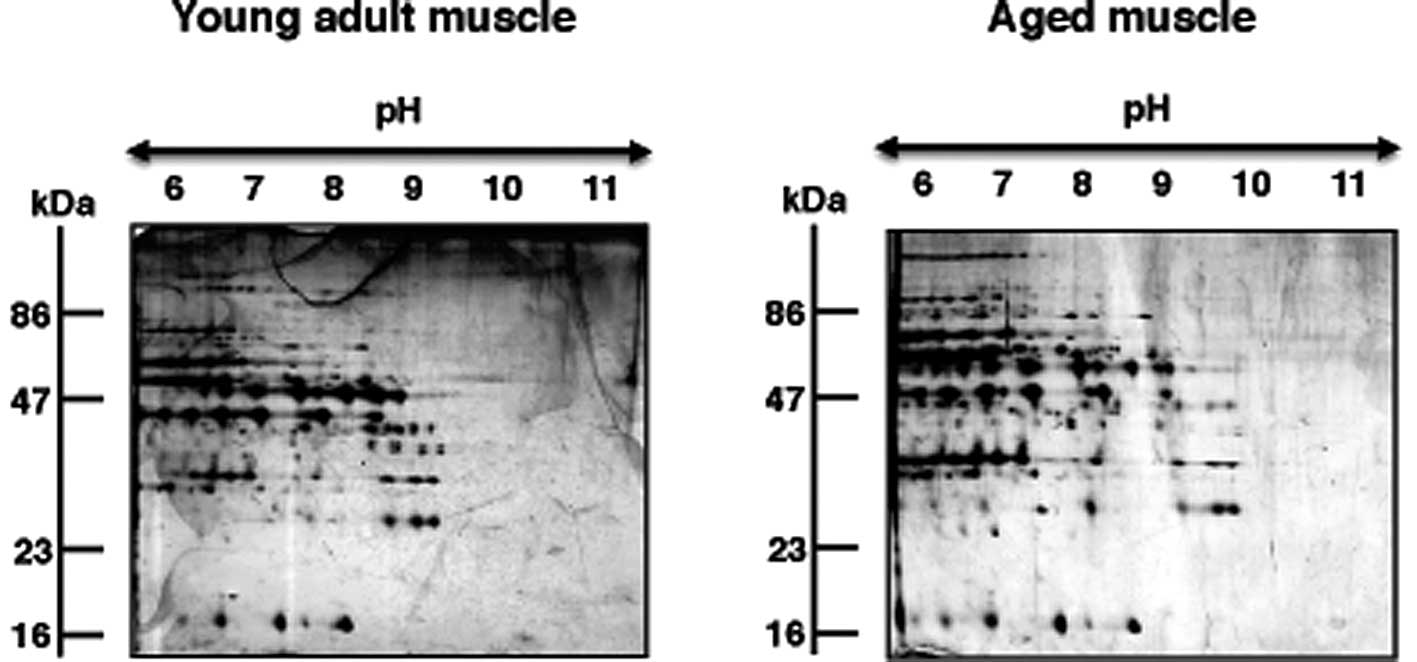
Fig. 1 and the separation patterns
in the basic pH-range are shown in Fig. 2 that compares young adult versus
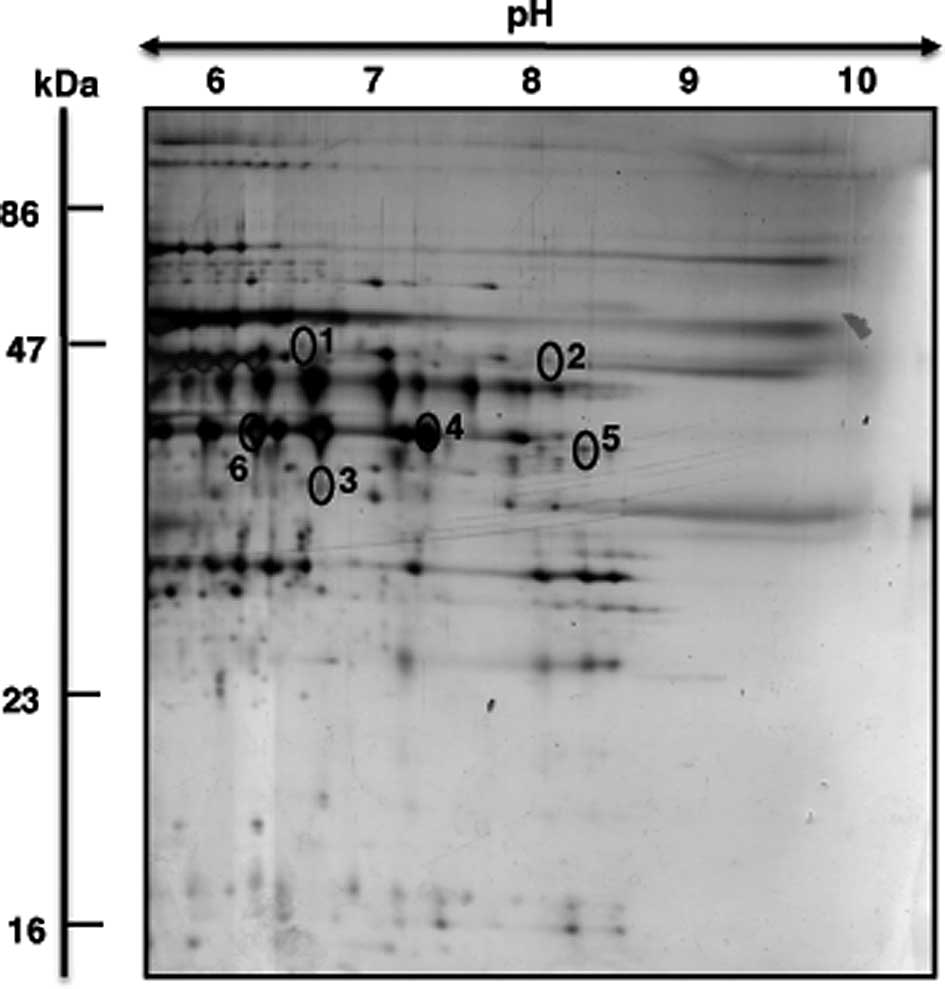
aged muscle specimens. A mastergel of proteins with age-related
changes in abundance and their mass spectrometric identification is
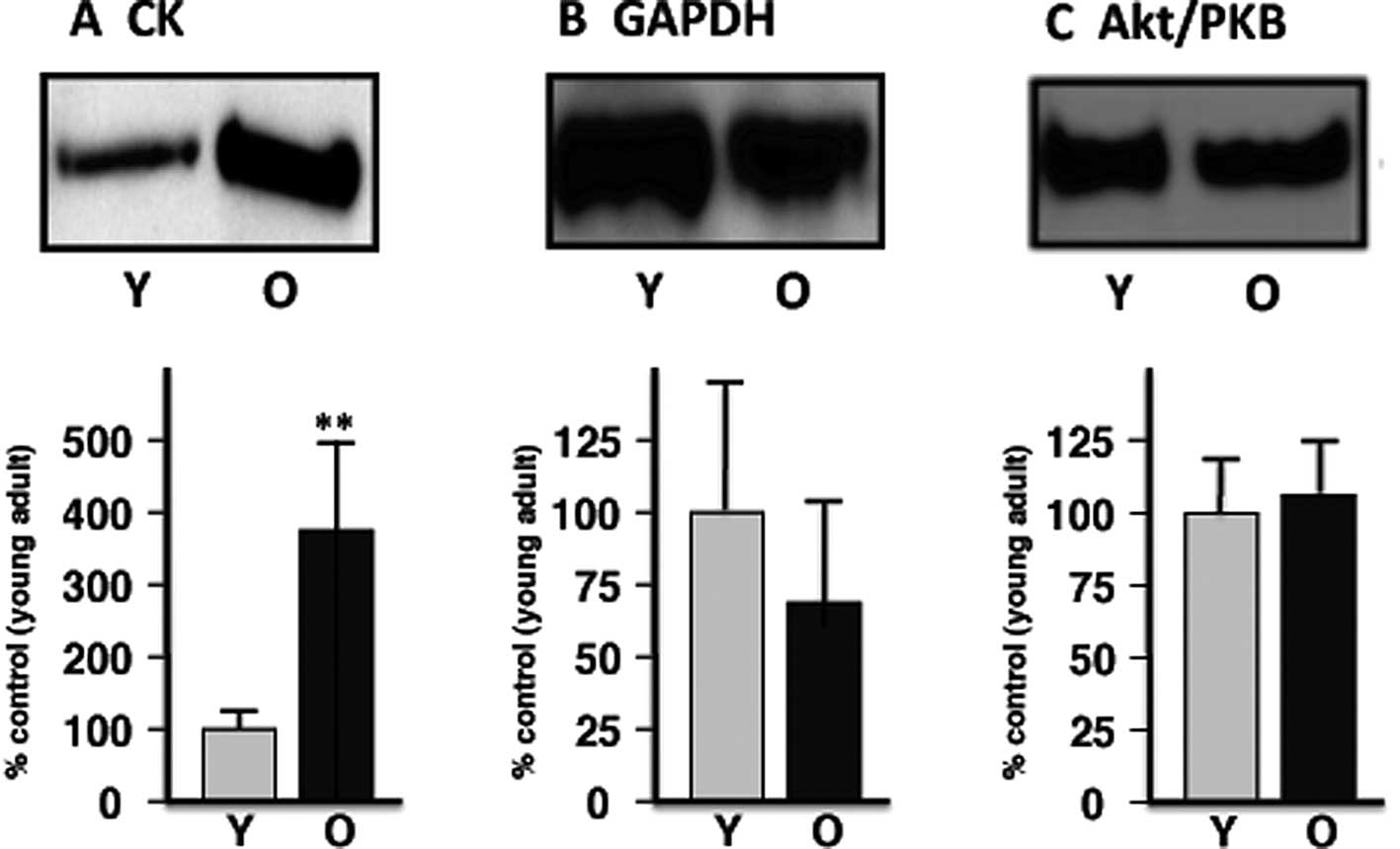
presented in Fig. 3 and Table I. Verification of proteomic data is
provided by comparative immunoblot analysis of select marker
proteins (Fig. 4).
 | Table IList of basic proteins with a changed
abundance in aged rat gastrocnemius muscle. |
Table I
List of basic proteins with a changed
abundance in aged rat gastrocnemius muscle.
| Spot no. | Accession no. | Name of identified
protein | Peptides
matched | Peptide
sequence | Coverage sequence
(%) | Molecular mass
(kDa) | Isoelectric point
(pI) | Mascot score | Fold-change |
|---|
| 1 | gi|38259206| | Mitochondrial
creatine kinase | 4 |
REVENVAITALEGLKG
KLSEMTEQDQQRL
RGIWHNYDKT
KVPPPLPQFGRK | 10 | 47.89 | 8.64 | 69 | 1.5 |
| 2 | gi|55741544| | Ubiquinol
cytochrome-c reductase core protein 2 | 7 |
KLPNGLVIASLENYAPLSRI
RRWEVAALRS
KAVAFQNPQTRI
RIIENLHDVAYKN
KEVAEQFLNIRG
RGGLGLAGAKA
KAVAQGNLSSADVQAAKN | 18 | 48.42 | 9.16 | 81 | 1.5 |
| 3 | gi|6671762| | Muscle creatine
kinase | 8 |
KLNYKPQEEYPDLSKH
KVLTPDLYNKL
KDLFDPIIQDRH
KTDLNHENLKG
KGGDDLDPNYVLSSRV
KFEEILTRL
RGTGGVDTAAVGAVFDISNADRL
RLGSSEVEQVQLVVDGVKL | 26 | 43.25 | 6.58 | 168 | −1.5 |
| 4 | gi|62664602| | Glyceraldehyde
3-phosphate-dehydrogenase (GAPDH) | 5 |
KLVINGKPITIFQERD
RGAAQNIIPASTGAVKAVGKV
KVIPELNGKL
KLISWYDNEYGYSNRV
RVVDLMAYMASKE | 19 | 36.18 | 7.6 | 84 | −1.6 |
| 5 | gi|56188| | GAPDH | 2 |
KVIHDNFGIVEGLMTTVHAITATQKT
KLISWYDNEYGYSNRV | 11 | 36.10 | 8.43 | 126 | −1.6 |
| 6 | gi|56188| | GAPDH | 10 |
KVGVNGFGRI
KLVINGKPITIFQERD
KIVSNASCTTNCLAPLAKV
RGAAQNIIPASTGAAKA
KVIPELNGKLTGMAFRV
KLTGMAFRV
RVPTPNVSVVDLTCRL
KLISWYDNEYGYSNRV
RVVDLMAYMASKE
RVVDLMAYMASKE | 32 | 36.09 | 8.43 | 185 | −1.9 |
Offgel electrophoretic separation of the
skeletal muscle proteome
In order to improve protein separation in the basic
pH-range, offgel electrophoresis was employed as a
pre-fractionation step. Running conditions were optimized with
respect to loading volume and protein amounts needed for a proper
offgel electrophoresis-based separation of muscle proteins (data
not shown). An amount of 1 mg protein/lane resulted in an
acceptable rate of protein recovery following offgel
electrophoresis, dialysis and acetone precipitation for protein
fractionation, removal of access urea and protein concentration,
respectively. Fig. 1 outlines the
various experimental steps used in the analysis of the basic
protein cohort from young versus aged gastrocnemius muscle. The
silver-stained 2D gels of the acidic complement versus the basic
cohort of urea-soluble muscle proteins show a comparable protein
separation for the pH 4.0–7.0 range and an improved separation
pattern for the pH 6.0–9.0 range, as compared to previously
published 2D maps of crude skeletal muscle preparations (24–26).
Gel electrophoretic comparison of young
adult versus aged skeletal muscle
The 2D gels presented in Fig. 2 illustrate the expression pattern
of basic proteins from young adult versus aged rat gastrocnemius
muscle. Both gels show a relatively comparable 2D protein spot
distribution in the pH 6.0–11.0 range. In order to determine
potential changes in abundance in individual protein spots, gels
were analyzed with Progenesis SameSpots analysis software. The
densitometric analysis of the 2D gels revealed 12 muscle-associated
proteins with an altered abundance. Of these differentially
expressed protein species, 6 protein isoforms could unequivocally
be identified by MS.
Mass spectrometric (MS) identification of
muscle proteins with an age-related change in abundance
The representative mastergel in Fig. 3 shows the 6 identified proteins
that differed in their abundance between young adult and senescent
muscle preparations. Proteins ranged in molecular mass from 36–48
kDa and exhibited an isoelectric point from pI 6.6–9.1. Table I lists spot numbers, protein
accession numbers, name of identified proteins, number of matched
peptides, peptide sequences, sequence coverage, molecular mass,
isolectric point, Mascot score and fold-change of individual
protein species during aging. Listed are muscle-associated protein
with a Mascot score above 60. Protein spots with an increased
density were identified as two key mitochondrial components, the
mitochondrial isoform of CK (spot 1) and ubiquinol cytochrome-c
reductase (spot 2). A reduction was shown for the muscle-type CK
(spot 3) and the glycolytic enzyme GAPDH (spots 4–6).
Immunoblot analysis of marker proteins in
aging skeletal muscle
In order to verify the differential expression of
muscle-associated components that was identified by our proteomic
survey of basic proteins, immunoblotting was carried out. Fig. 4 illustrates the immuno-decoration
of CK versus GAPDH, as well as Akt/PKB as a loading control. In
agreement with the proteomic establishment of an increased level of
mitochondrial CK and a decreased concentration of GAPDH, immunoblot
analysis showed a significant elevation of the mitochondrial enzyme
and suggests a reduced presence of the glycolytic protein in aged
skeletal muscle.
Discussion
As the primary site for energy generation via
oxidative phosphorylation, mitochondria play a key role in skeletal
muscle metabolism. In addition, this class of organelle is involved
in protein transportation, cell cycle progression, calcium
handling, intermediary metabolism and apoptotic regulation
(39). Alterations in the
abundance and/or post-translational modifications of mitochondrial
enzymes are believed to be involved in the natural aging process
(40). In addition, altered
concentration levels of mitochondrial enzymes from skeletal muscle
are often an indication of fiber type shifting during physiological
or pathophysiological muscle adaptations (41). In this respect, the findings
presented in this subproteomic survey of basic proteins in young
adult versus senescent rat gastrocnemius muscle support the idea of
a glycolytic-to-oxidative shift during skeletal muscle aging. The
mass spectrometric identification of altered basic proteins has
established an increase in two key mitochondrial enzymes, ubiquinol
cytochrome-c reductase core protein 2 and mitochondrial CK and a
concomitant decrease in the glycolytic marker enzyme GAPDH in
senescent rat muscle. This metabolic shift would imply a
fast-to-slow muscle transformation process during age-related
adaptations of contractile fibers. The mitochondrial and glycolytic
aging markers identified in this study are enzymes with
well-characterized metabolic functions. CK is a key metabolic
enzyme of the mitochondrial inter-membrane space and is involved in
the production of creatine phosphate from mitochondrial ATP and
cytosolic creatine (42).
Ubiquinol cytochrome-c reductase is an essential subunit of the
cytochrome-bc1 complex, making it a crucial element of the
mitochondrial respiratory chain (43). GAPDH is a key enzyme of the
glycolytic pathway that converts glyceraldehyde-3-phosphate to
1,3-bisphosphoglycerate and generates NADH (44).
Previous proteomic studies of both human and animal
muscles have shown a general trend of muscle transitions to more
aerobic-oxidative metabolism in a slower-twitching aged fiber
population (22–30). This would agree with the results of
the subproteomic analysis of basic muscle proteins presented in
this study. Aged muscle fibers are under considerable oxidative
stress and exhibit mitochondrial abnormalities (45–47).
Importantly, changes in the abundance of mitochondrial versus
glycolytic enzymes are often accompanied by shifts in fiber type
distribution patterns of individual muscles (48). For example, chronic low-frequency
stimulation triggers a gradual shift towards slower isoforms of
contractile and calcium handling proteins, as well as a drastic
increase in mitochondrial enzymes (49). The same alterations in metabolic
enzymes have been described in the case of pathological fiber type
transitions. The proteomic profiling of the ADR mouse model of
myotonia has clearly shown increased levels of mitochondrial
enzymes in hyper-excitable muscle tissue (50). With respect to sarcopenia, the
exact extent of a glycolytic-to-oxidative shift and accompanying
fast-to-slow muscle transformation process is still the subject of
intense debate in the field of biogerontology. Both human and
animal skeletal muscles show changes in their molecular and
cellular constellation and clearly exhibit alterations in their
glycolytic and aerobic capacity. However, large-scale histological
surveys of potential alterations in fiber type distributions with
aging have shown variable results (51–54).
The results from this report have clearly demonstrated differential
changes in the abundance of mitochondrial and glycolytic marker
proteins during rat skeletal muscle aging.
Muscle aging appears to be associated with a
plethora of cellular and functional abnormalities, including: i)
impaired neuromuscular transmission resulting in the loss of entire
motor units and cycles of denervation and impaired reinnervation
(4), ii) the uncoupling between
plasmalemmal excitation and fiber contraction (55), iii) altered functioning of the
actomyosin apparatus causing a decline in contractile efficiency
(56), iv) an altered equilibrium
of growth factors and hormones involved in the maintenance of
muscle contraction (57), v)
denervation-associated atrophy (2), vi) increased apoptosis (58), vii) impaired regulation of
bioenergetic processes (59),
viii) disturbed ion handling (60), ix) increased
Ca2+-dependent calpain activity triggering mitochondrial
and cytoskeletal dysfunction (61), x) a drastically decreased capacity
for cellular regeneration (62)
and xi) a diminished stress response by molecular chaperones
(63). Here, we confirm the idea
that a glycolytic-to-oxidative shift occurs in skeletal muscle
metabolism during the natural aging process. In the future, the
identified mitochondrial and glycolytic enzymes could be useful as
potential biomarkers of fiber aging. Since mitochondria are of
central importance for energy generation in contractile fibers and
because alterations in the density and/or function of mitochondrial
enzymes are generally involved in muscle development, muscle
differentiation, fiber adaptations and a variety of muscle
diseases, the establishment of reliable biomarker signatures is
crucial. As shown here, MS-based subproteomics represents a
convenient large-scale approach to identify mitochondrial markers
of muscle aging.
In conclusion, since large-scale surveys of fiber
type distributions during muscle aging have resulted in variable
findings, it was of interest to determine potential changes in the
abundance of mitochondrial and glycolytic marker proteins in aged
skeletal muscle by subproteomics. We showed here that a
glycolytic-to-oxidative shift occurs in an animal model of
sarcopenia, the 30-month-old Wistar rat. In the long-term, the
systematic cataloguing of age-related alterations in the
concentration, post-translational modifications, physiological
functioning and subcellular localization of mitochondrial enzymes
might be useful for the establishment of a comprehensive biomarker
signature of skeletal muscle aging. International working groups on
skeletal muscle aging have not yet fully established a definition
and agreed clinical assessment of sarcopenia as a common geriatric
syndrome (64–66). It is therefore crucial to identify
and characterize reliable biomarkers of sarcopenia of old age
(67). Once suitable combinations
of marker enzymes have been characterized, they might be useful for
improving diagnostic procedures to differentiate mild versus severe
forms of sarcopenia, advance biomedical approaches for monitoring
disease progression, expand the set of bioanalytical tools for
testing potential side effects of new drug regimes and shed new
light on the molecular pathogenesis of sarcopenia of old age.
Acknowledgements
Research in the author’s laboratory was supported by
grants from the Science Foundation Ireland and the Higher Education
Authority. We thank Anne Kirwan for the excellent technical
support.
Abbreviations:
|
2D
|
two-dimensional
|
|
Akt/PKB
|
protein kinase B
|
|
CK
|
creatine kinase
|
|
GAPDH
|
glyceraldehyde-3-phosphate
dehydrogenase
|
|
IEF
|
isoelectric focusing
|
|
MS
|
mass spectrometry
|
|
SDS
|
sodium dodecyl sulfate
|
|
PAGE
|
polyacrylamide gel electrophoresis
|
|
PBS
|
phosphate-buffered saline
|
References
|
1
|
DR ThomasSarcopeniaClin Geriatr
Med26331346201010.1016/j.cger.2010.02.012
|
|
2
|
AA VandervoortAging of the human
neuromuscular systemMuscle
Nerve251725200210.1002/mus.121511754180
|
|
3
|
LV ThompsonAge-related muscle
dysfunctionExp
Gerontol44106111200910.1016/j.exger.2008.05.00318657920
|
|
4
|
JA FaulknerLM LarkinDR ClaflinSV
BrooksAge-related changes in the structure and function of skeletal
musclesClin Exp Pharmacol
Physiol3410911096200710.1111/j.1440-1681.2007.04752.x17880359
|
|
5
|
E EdstromM AltunE BergmanH JohnsonS
KullbergV Ramirez-LeonB UlfhakeFactors contributing to
neuromuscular impairment and sarcopenia during agingPhysiol
Behav92129135200710.1016/j.physbeh.2007.05.04017585972
|
|
6
|
JG RyallJD SchertzerGS LynchCellular and
molecular mechanisms underlying age-related skeletal muscle wasting
and
weaknessBiogerontology9213228200810.1007/s10522-008-9131-018299960
|
|
7
|
WR FronteraKF ReidEM PhillipsLS
KrivickasVA HughesR RoubenoffRA FieldingMuscle fiber size and
function in elderly humans: a longitudinal studyJ Appl
Physiol105637642200810.1152/japplphysiol.90332.200818556434
|
|
8
|
MC KostekMJ DelmonicoAge-related changes
in adult muscle morphologyCurr Aging SciApril292011(Epub ahead of
print)
|
|
9
|
AA VandervoortTB SymonsFunctional and
metabolic consequences of sarcopeniaCan J Appl
Physiol2690101200110.1139/h01-00711173671
|
|
10
|
JS KimJM WilsonSR LeeDietary implications
on mechanisms of sarcopenia: roles of protein, amino acids and
antioxidantsJ Nutr
Biochem21113201010.1016/j.jnutbio.2009.06.01419800212
|
|
11
|
Y RollandC DupuyG Abellan van KanS
GilletteB VellasTreatment strategies for sarcopenia and frailtyMed
Clin North Am95427438201110.1016/j.mcna.2011.02.00821549870
|
|
12
|
BF HurleyED HansonAK SheaffStrength
training as a countermeasure to aging muscle and chronic
diseaseSports
Med41289306201110.2165/11585920-000000000-0000021425888
|
|
13
|
M PahorS KritchevskyResearch hypotheses on
muscle wasting, aging, loss of function and disabilityJ Nutr Health
Aging297100199810993575
|
|
14
|
DR ThomasLoss of skeletal muscle mass in
aging: examining the relationship of starvation, sarcopenia and
cachexiaClin Nutr26389399200710.1016/j.clnu.2007.03.00817499396
|
|
15
|
JE MorleyDiabetes, sarcopenia, and
frailtyClin Geriatr Med24455469200810.1016/j.cger.2008.03.004
|
|
16
|
P DoranJ GannonK O’ConnellK
OhlendieckProteomic profiling of animal models mimicking skeletal
muscle disordersProteomics Clin
Appl111691184200710.1002/prca.20070004221136766
|
|
17
|
P DoranP DonoghueK O’ConnellJ GannonK
OhlendieckProteomics of skeletal muscle
agingProteomics99891003200910.1002/pmic.20080036519180535
|
|
18
|
J KanskiSJ HongC SchoneichProteomic
analysis of protein nitration in aging skeletal muscle and
identification of nitrotyrosine-containing sequences in vivo by
nanoelectrospray ionization tandem mass spectrometryJ Biol
Chem2802426124266200510.1074/jbc.M501773200
|
|
19
|
J GannonL StauntonK O’ConnellP DoranK
OhlendieckPhosphoproteomic analysis of aged skeletal muscleInt J
Mol Med2233422008
|
|
20
|
J FengH XieDL MeanyLV ThompsonEA ArriagaTJ
GriffinQuantitative proteomic profiling of muscle type-dependent
and age-dependent protein carbonylation in rat skeletal muscle
mitochondriaJ Gerontol A Biol Sci Med
Sci6311371152200810.1093/gerona/63.11.113719038828
|
|
21
|
K O’ConnellP DoranJ GannonK
OhlendieckLectin-based proteomic profiling of aged skeletal muscle:
decreased pyruvate kinase isozyme M1 exhibits drastically increased
levels of N-glycosylationEur J Cell Biol877938052008
|
|
22
|
I PiecA ListratJ AlliotC ChambonRG TaylorD
BechetDifferential proteome analysis of aging in rat skeletal
muscleFASEB J1911431145200515831715
|
|
23
|
C GelfiA ViganoM RipamontiA PontoglioS
BegumMA PellegrinoB GrassiR BottinelliR WaitP CerretelliThe human
muscle proteome in agingJ Proteome
Res513441353200610.1021/pr050414x16739986
|
|
24
|
K O’ConnellJ GannonP DoranK
OhlendieckProteomic profiling reveals a severely perturbed protein
expression pattern in aged skeletal muscleInt J Mol
Med20145153200717611631
|
|
25
|
P DoranJ GannonK O’ConnellK
OhlendieckAging skeletal muscle shows a drastic increase in the
small heat shock proteins alphaB-crystallin/HspB5 and
cvHsp/HspB7Eur J Cell
Biol86629640200710.1016/j.ejcb.2007.07.003
|
|
26
|
P DoranK O’ConnellJ GannonM KavanaghK
OhlendieckOpposite pathobiochemical fate of pyruvate kinase and
adenylate kinase in aged rat skeletal muscle as revealed by
proteomic DIGE
analysisProteomics8364377200810.1002/pmic.20070047518050275
|
|
27
|
D CapitanioM VassoC FaniaM MoriggiA
ViganoP ProcacciV MagnaghiC GelfiComparative proteomic profile of
rat sciatic nerve and gastrocnemius muscle tissues in ageing by 2-D
DIGEProteomics920042020200910.1002/pmic.20070116219333999
|
|
28
|
J GannonP DoranA KirwanK OhlendieckDrastic
increase of myosin light chain MLC-2 in senescent skeletal muscle
indicates fast-to-slow fibre transition in sarcopenia of old ageEur
J Cell Biol88685700200910.1016/j.ejcb.2009.06.00419616867
|
|
29
|
P DonoghueL StauntonE MullenG ManningK
OhlendieckDIGE analysis of rat skeletal muscle proteins using
nonionic detergent phase extraction of young adult versus aged
gastrocnemius tissueJ
Proteomics7314411453201010.1016/j.jprot.2010.01.014
|
|
30
|
K O’ConnellK OhlendieckProteomic DIGE
analysis of the mitochondria-enriched fraction from aged rat
skeletal muscleProteomics955095524200919834913
|
|
31
|
I BraticA TrifunovicMitochondrial energy
metabolism and ageingBiochim Biophys
Acta1797961967201010.1016/j.bbabio.2010.01.00420064485
|
|
32
|
G PariseM De LisioMitochondrial theory of
aging in human age-related sarcopeniaInterdiscip Top
Gerontol37142156201010.1159/00031999920703060
|
|
33
|
L StauntonK O’ConnellK OhlendieckProteomic
profiling of mitochondrial enzymes during skeletal muscle agingJ
Aging Res2011908035201110.4061/2011/90803521437005
|
|
34
|
PG RighettiA CastagnaP AntonioliE
BoschettiPrefractio nation techniques in proteome analysis: the
mining tools of the third
millenniumElectrophoresis26297319200510.1002/elps.20040618915657944
|
|
35
|
P HorthCA MillerT PreckelC WenzEfficient
fractionation and improved protein identification by peptide OFFGEL
electrophoresisMol Cell
Proteomics519681974200610.1074/mcp.T600037-MCP20016849286
|
|
36
|
EM KeidelD DoschA BrunnerJ KellermannF
LottspeichEvaluation of protein loading techniques and improved
separation in OFFGEL isoelectric
focusingElectrophoresis3216591666201121563181
|
|
37
|
S FratermanU ZeigerTS KhuranaNA
RubinsteinM WilmCombination of peptide OFFGEL fractionation and
label-free quantitation facilitated proteomics profiling of
extraocular
muscleProteomics734043416200710.1002/pmic.20070038217708596
|
|
38
|
M ChevalletS LucheT RabilloudSilver
staining of proteins in polyacrylamide gelsNat
Protoc118521858200610.1038/nprot.2006.28817487168
|
|
39
|
HM McBrideM NeuspielS WasiakMitochondria:
more than just a powerhouseCurr
Biol16R551R560200610.1016/j.cub.2006.06.05416860735
|
|
40
|
DC ChanMitochondria: dynamic organelles in
disease, aging, and
developmentCell12512411252200610.1016/j.cell.2006.06.01016814712
|
|
41
|
DA HoodI IrrcherV LjubicicAM
JosephCoordination of metabolic plasticity in skeletal muscleJ Exp
Biol20922652275200610.1242/jeb.0218216731803
|
|
42
|
U SchlattnerM Tokarska-SchlattnerT
WallimannMitochondrial creatine kinase in human health and
diseaseBiochim Biophys
Acta1762164180200610.1016/j.bbadis.2005.09.00416236486
|
|
43
|
NV DudkinaR KourilK PetersHP BraunEJ
BoekemaStructure and function of mitochondrial
supercomplexesBiochim Biophys
Acta1797664670201010.1016/j.bbabio.2009.12.01320036212
|
|
44
|
K OhlendieckProteomics of skeletal muscle
glycolysisBiochim Biophys
Acta180420892101201010.1016/j.bbapap.2010.08.00120709194
|
|
45
|
A HionaC LeeuwenburghThe role of
mitochondrial DNA mutations in aging and sarcopenia: implications
for the mitochondrial vicious cycle theory of agingExp
Gerontol432433200810.1016/j.exger.2007.10.00117997255
|
|
46
|
B ChabiV LjubicicKJ MenziesJH HuangA
SaleemDA HoodMitochondrial function and apoptotic susceptibility in
aging skeletal muscleAging
Cell7212200810.1111/j.1474-9726.2007.00347.x18028258
|
|
47
|
PA FigueiredoSK PowersRM FerreiraHJ
AppellJA DuarteAging impairs skeletal muscle mitochondrial
bioenergetic functionJ Gerontol A Biol Sci Med
Sci642133200910.1093/gerona/gln04819196905
|
|
48
|
D PetteThe adaptive potential of skeletal
muscle fibersCan J Appl
Physiol27423448200210.1139/h02-02312442355
|
|
49
|
P DonoghueP DoranK WynneK PedersenMJ DunnK
OhlendieckProteomic profiling of chronic low-frequency stimulated
fast
muscleProteomics734173430200710.1002/pmic.20070026217708595
|
|
50
|
L StauntonH JockuschC WiegandT AlbrechtK
OhlendieckIdentification of secondary effects of hyperexcitability
by proteomic profiling of myotonic mouse muscleMol
Biosyst724802489201110.1039/c1mb05043e21629954
|
|
51
|
MA AlnaqeebG GoldspinkChanges in fiber
type, number and diameter in developing and ageing skeletal muscleJ
Anat153314519873429325
|
|
52
|
MR DeschenesEffects of aging on muscle
fiber type and sizeSports
Med34809824200410.2165/00007256-200434120-0000215462613
|
|
53
|
J LexellCC TaylorM SjostromWhat is the
cause of the ageing atrophy? Total number, size and proportion of
different fiber types studied in whole vastus lateralis muscle from
15- to 83-year-old menJ Neurol Sci842752941988
|
|
54
|
J LexellHuman aging, muscle mass, and
fiber type compositionJ Gerontol A Biol Sci Med Sci5011161995
|
|
55
|
O DelbonoExpression and regulation of
excitation-contraction coupling proteins in aging skeletal
muscleCurr Aging SciApril292011(Epub ahead of print)
|
|
56
|
E ProchniewiczLV ThompsonDD
ThomasAge-related decline in actomyosin structure and functionExp
Gerontol42931938200710.1016/j.exger.2007.06.01517706387
|
|
57
|
CE LeeA McArdleRD GriffithsThe role of
hormones, cytokines and heat shock proteins during age-related
muscle lossClin
Nutr26524534200710.1016/j.clnu.2007.05.00517590243
|
|
58
|
E MarzettiC LeeuwenburghSkeletal muscle
apoptosis, sarcopenia and frailty at old ageExp
Gerontol4112341238200610.1016/j.exger.2006.08.01117052879
|
|
59
|
DW RussIR LanzaThe impact of old age on
skeletal muscle energetics: supply and demandCurr Aging
Sci2011Apr29(in press)
|
|
60
|
K O’ConnellJ GannonP DoranK
OhlendieckReduced expression of sarcalumenin and related
Ca2+-regulatory proteins in aged rat skeletal muscleExp
Gerontol43958961200818762239
|
|
61
|
C BruleE DargelosR DialloA ListratD
BechetP CottinS PoussardProteomic study of calpain interacting
proteins during skeletal muscle
agingBiochimie9219231933201010.1016/j.biochi.2010.09.00320850499
|
|
62
|
P LorenzonE BandiF de GuarriniT
PietrangeloR SchaferM ZweyerA WernigF RuzzierAgeing affects the
differentiation potential of human myoblastsExp
Gerontol3915451554200410.1016/j.exger.2004.07.00815501025
|
|
63
|
AC KayaniJP MortonA McArdleThe
exercise-induced stress response in skeletal muscle: failure during
agingAppl Physiol Nutr
Metab3310331041200810.1139/H08-08918923581
|
|
64
|
T LangT StreeperP CawthonK BaldwinDR
TaaffeTB HarrisSarcopenia: etiology, clinical consequences,
intervention, and assessmentOsteoporos
Int21543559201010.1007/s00198-009-1059-y19779761
|
|
65
|
AJ Cruz-JentoftJP BaeyensJM BauerY BoirieT
CederholmF LandiFC MartinJP MichelY RollandSM SchneiderSarcopenia:
European consensus on definition and diagnosis: report of the
European working group on sarcopenia in older peopleAge
Ageing39412423201010.1093/ageing/afq03420392703
|
|
66
|
AJ Cruz-JentoftF LandiE TopinkovaJP
MichelUnderstanding sarcopenia as a geriatric syndromeCurr Opin
Clin Nutr Metab
Care1317201010.1097/MCO.0b013e328333c1c119915458
|
|
67
|
M PahorT ManiniM CesariSarcopenia:
clinical evaluation, biological markers and other evaluation toolsJ
Nutr Health Aging13724728200910.1007/s12603-009-0204-919657557
|