Introduction
Oral cavity cancer is among the leading 10 most
commonly diagnosed cancers. Squamous cell carcinoma of the tongue
is one of the most common tumors in head and neck cancers (1). During the past several decades, great
progress has been made in surgery, radiation therapy and
chemotherapy, yet the 5-year survival rate of oral cancer has
remained approximately 50% (2). In
addition, these patients are at high risk for tumor recurrence and
second primary tumors after regular treatment. Furthermore, the
molecular mechanisms underlying the pathogenesis of oral cancer
remain elusive.
Cancer stem cells (CSCs) were first discovered in
1997 in acute myeloid leukemia, and have recently been found in
several tumor types, including lung, breast, brain, liver, pancreas
and colon cancers (3,4). This unique subpopulation of cancer
cells possesses the ability to self-renew, proliferate and
differentiate through developmental signaling pathways in aberrant
ways (4). CSCs have also been
reported to be involved in tumor metastasis and recurrence.
Accumulating data indicate that tongue squamous cell carcinoma
harbors tumor-initiating cells or CSCs (5–7).
Certain CSC-related genes, such as bone morphogenetic protein-4
(BMP-4), octamer-binding transcription factor 4 (Oct4), the
homeobox protein Nanog, CD133 and B-cell-specific Moloney murine
leukemia virus insertion site 1 (Bmi-1), have been identified and
characterized in tongue squamous cell carcinoma (5,7–10).
Aldehyde dehydrogenase (ALDH) enzymes play a
critical role in the metabolism of many molecules, and in the
detoxification of external and internal substances, such as alcohol
and toxins. Studies have shown that ALDH is a CSC marker in certain
solid tumors, including colon, breast and lung cancers (11–13).
In addition, the 19 ALDH family members may function differently,
depending on the type of tissue and tumors (14). In this study, we observed that the
Tca8113 tongue squamous cell carcinoma cell line harbored 1.3% of
ALDH+ cells, which displayed CSC characteristics. The
ALDH+ subpopulation possessed an elevated capacity to
proliferate, differentiate and self-renew in comparison to their
ALDH− counterparts. Our data suggest that ALDH activity
is a CSC marker for tongue squamous cell carcinoma; therefore,
targeting ALDH may be a potential therapeutic strategy.
Materials and methods
Cell line and reagents
The Tca8113 human tongue squamous cell carcinoma
cell line was provided by the State Key Laboratory of Oral Diseases
of Sichuan University, China. Tca8113 cells were cultured in
RPMI-1640 (Life Technologies Inc.) and supplemented with 10% fetal
calf serum (Gibco) at 37°C in a humidified atmosphere containing 5%
CO2. Epidermal growth factors and basic fibroblast
growth factors were purchased from PeproTech.
Aldefluor assay and flow cytometry
The Aldefluor kit (Stem Cell Technologies) was
utilized to profile and isolate cells with high and low ALDH
activity as previously described (15). Cells were incubated in Aldefluor
assay buffer containing the ALDH substrate,
BODIPY-aminoacetaldehyde (BAAA), at 37°C for 45 min. Cells that
were able to catalyze BAAA to its fluorescent product,
BODIPY-aminoacetate (BAA), were considered ALDH+ cells.
The enzymatic activity of ALDH was blocked by a specific inhibitor,
DEAB. Sorting gates for fluorescence-activated cell sorting (FACS)
were drawn relative to baseline fluorescence, which was determined
by DEAB-treated samples. After incubation, cells were resuspended
in fresh assay buffer. ALDH+ and ALDH− cells
were sorted by a BD Aria (BD Biosciences).
Proliferation assay
Sorted ALDH+, ALDH− cells and
their parental cells were seeded in 96-well plates with a density
of 1,000 cells/ml. Cell proliferation was detected after culturing
for 1, 3, 5 and 7 days using the Cell Counting kit-8 (Dojindo)
according to the manufacturer’s instructions. Data are shown as the
means ± SD.
Differentiation assay
Unsorted and sorted single cells were cultured in
regular medium. ALDH activity was measured when the cells became
85% confluent.
Tumorsphere formation assay
Sorted ALDH+ and ALDH− single
cells were cultured in serum-free medium in 6-well plates at a
density of 20,000 cells/ml. Tumorspheres were detected and images
were captured using a Zeiss inverted microscope.
Suppression subtractive hybridization
(SSH)
Total RNA from sorted ALDH+ and
ALDH− cells was extracted with TRIzol (Invitrogen)
according to the manufacturer’s instructions. Double-stranded cDNA
was generated using PCR-Select cDNA Subtraction kit (Clontech).
cDNAs were digested with RsaI (New England Biolabs). The
tester cDNA (ALDH+ cells) was subdivided into two
portions and each was ligated to a different double-stranded
adaptor (1 and 2). Enrichment of differentially expressed
transcripts was achieved in two successive rounds of PCR
amplifications that employed different adapters (1 and 2), and
sequential subtractive hybridization of the tester to an excess of
driver cDNA (ALDH− cells). Purified cDNA was ligated
into the pGEM-T easy vector system (Promega) and DH5α-competent
cells were transformed. Selected clones were grown in LB medium
with 1 μg/ml ampicillin. For sequencing, the plasmids were
extracted and sequenced using BigDye Terminator Cycle Sequencing
Ready Reaction kit (Applied Biosystems) with M13 primers on an
ABI-PRISM 3100 (Applied Biosystems) automated gene analyzer.
Statistical methods
Two-way ANOVA was performed to determine the
significant differences in cell growth between sorted and unsorted
cells in vitro.
Results
Identification of ALDH+
subpopulation in Tca8113 tongue squamous cell carcinoma cells
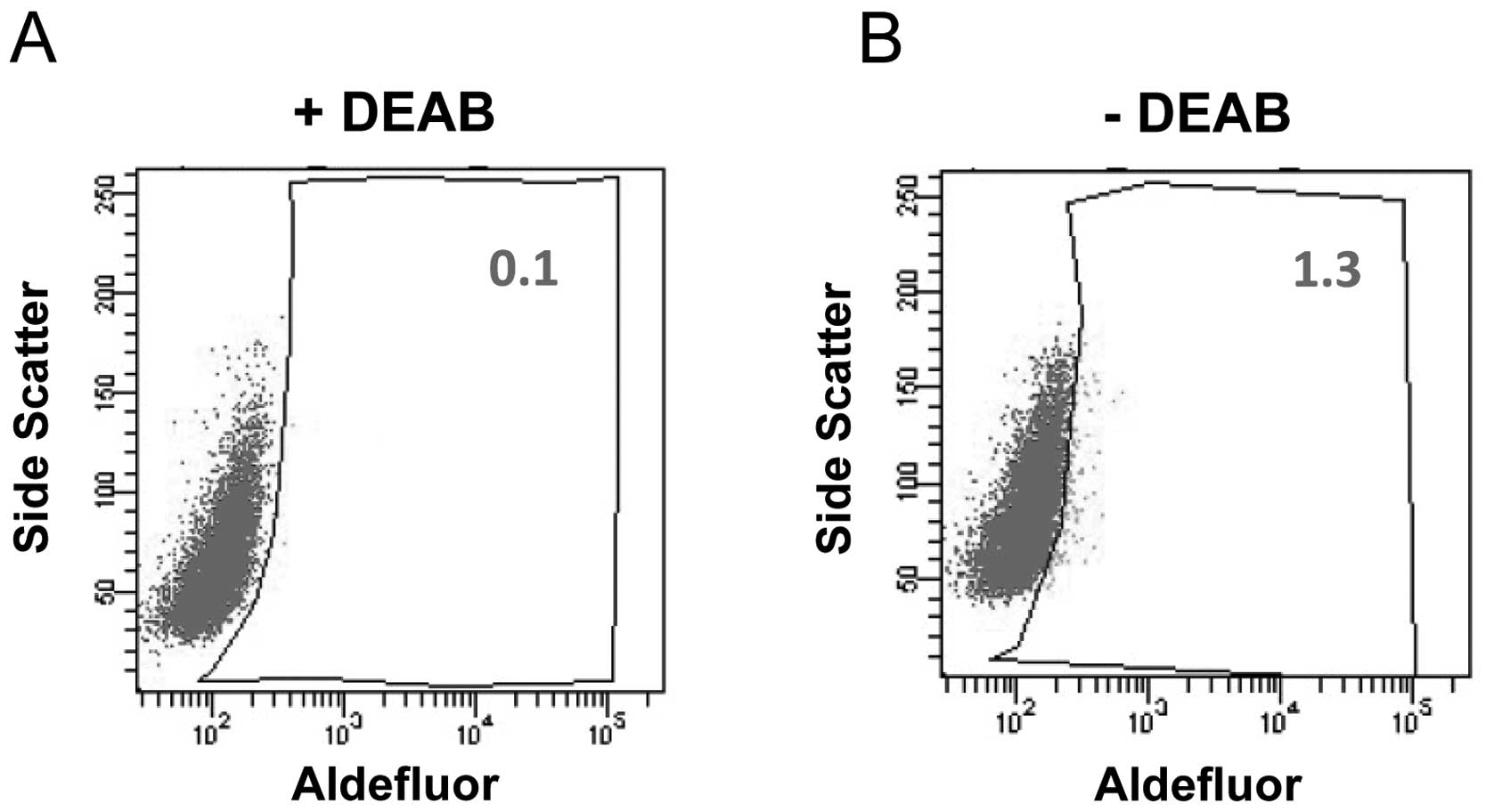
The enzymatic activity of ALDH has been demonstrated
to be a CSC marker in several cancer cell lines and patient tumor
samples (4). To examine whether
human tongue squamous cell carcinoma contains a subpopulation with
high ALDH activity, Aldefluor assays were performed to identify
ALDH+ cells in the Tca8113 cell line. In the presence of
the ALDH-specific inhibitor DEAB, ALDH+ cells were
barely observed. Notably, ~1.3% of Tca8113 cells displayed very
bright fluorescence in the absence of DEAB (Fig. 1). This result indicates that the
Tca8113 human tongue squamous cell carcinoma cell line contains a
subset of cells with high ALDH enzymatic activity. Our data are
consistent with a previous report that the Tca8113 cell line
harbors ~1% of CD133+ cells, which is another
well-studied CSC marker (7).
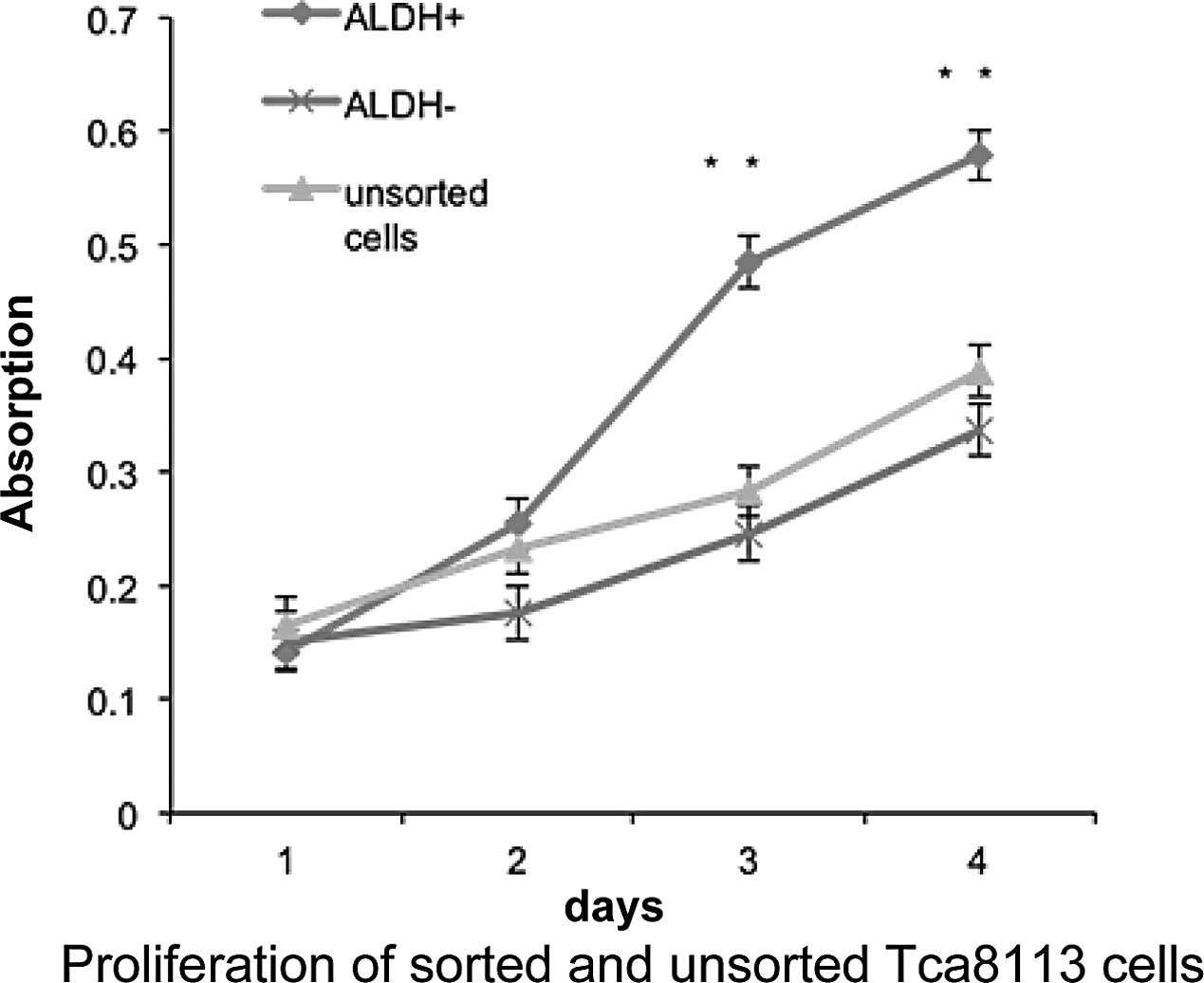
Enhanced proliferation of
ALDH+ tongue squamous cell carcinoma cells
CSCs possess an increased capacity for
proliferation, differentiation and tumorigenicity. We hypothesized
that ALDH+ cells in the Tca8113 cell line are CSCs. To
test this hypothesis, we compared the proliferation rate of sorted
ALDH+, ALDH− cells and parental Tca8113 cells
using Cell Counting Kit-8. After a 5-day culture, ALDH+
cells proliferated significantly faster than their ALDH−
counterparts, whereas the unsorted cells exhibited a similar
proliferation rate as that of the ALDH− cells (Fig. 2). This result indicates that
ALDH+ cells in the Tca8113 cell line have enhanced
capacity to proliferate relative to ALDH− cells in
vitro.
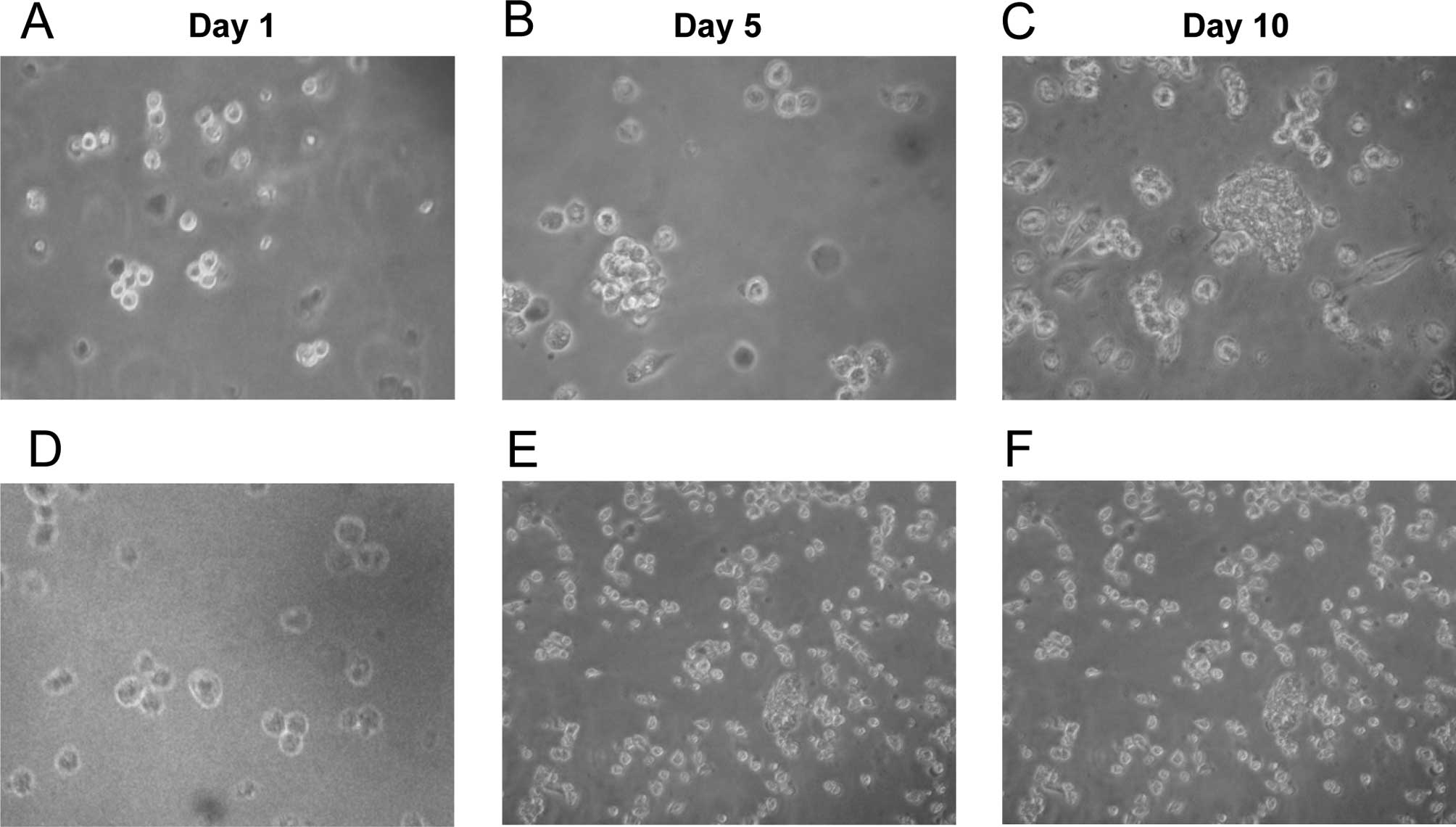
ALDH+ tongue squamous cell
carcinoma cells form tumorspheres
Similar to normal stem cells, CSCs are able to
self-renew through asymmetric cell divisions. Tumorsphere formation
assays are widely used to evaluate self-renewal and differentiation
at the single-cell level in vitro. Separated
ALDH+ and ALDH− single cells were maintained
in serum-free medium, in which non-CSCs cannot survive. On day 5,
ALDH+ cells started to aggregate with each other. Clear
tumorspheres were observed on day 10, while most single cells were
already dead (Fig. 3B and C). By
contrast, tumorspheres were barely found in the plates seeded with
ALDH− cells (Fig. 3E and
F).
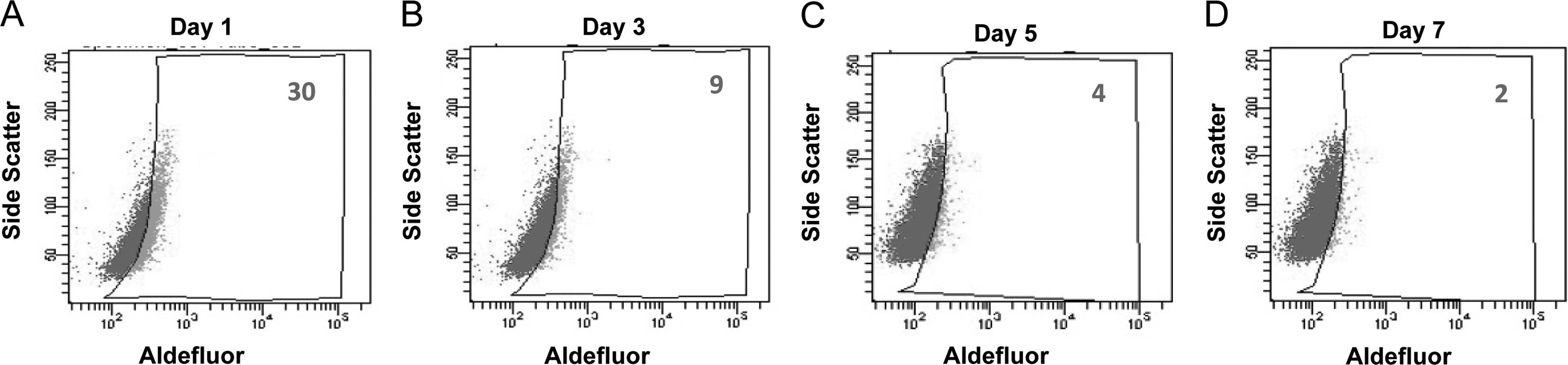
Differentiation of ALDH+
tongue squamous cell carcinoma cells
One critical characteristic of CSCs is the ability
to differentiate and form all types of tumor cells. To evaluate the
differentiation capacity of ALDH+ cells, the percentage
of ALDH+ cells was continuously measured during the
culture of isolated ALDH+ cells in regular medium. We
observed that the percentage of ALDH+ cells gradually
reduced to the level of parental Tca8113 cells, whereas the amount
of ALDH− counterparts increased (Fig. 4). Taken together, these data
demonstrate that ALDH+ cells self-renew, differentiate
and generate ALDH− subpopulations in vitro.
ALDH+ tongue squamous cell
carcinoma cells express CSC-related genes
Previous studies have reported that certain
signaling pathways, such as Notch, Wnt and Hedgehog pathways, play
critical roles in regulating CSCs for self-renewal, differentiation
and proliferation (4,12). Therefore, we compared gene
expression profiles between ALDH+ and ALDH−
fractions using the suppression subtraction hybridization assay.
Based on sequencing and bioinformatics analyses, 62 genes were
found to be differentially expressed in the two fractions isolated
from Tca8113 cells, among which 9 genes are involved in CSC biology
(Table I). For example, the Notch2
signaling pathway is known to be essential for the growth of tumor
neurospheres and xenografts (16).
In summary, there were 28 upregulated and 34 downregulated genes in
the ALDH+ subpopulation relative to the ALDH−
fraction. Our results provide insight into understanding the
molecular mechanisms of CSCs in tongue squamous cell carcinoma.
 | Table IDifferentially expressed cancer stem
cell-related genes. |
Table I
Differentially expressed cancer stem
cell-related genes.
| Gene name | Accession no. | Identified | E-value | Gene ID | Gene symbol | Chromosome
location |
|---|
| Homo sapiens
inositol 1,4,5-triphosphate receptor, type 1 (ITPR1), RefSeqGene on
chromosome 3 | NG_016144.1 | 253/254 (99%) | 1.00E-127 | 269954693 | ITPR1 | 3p26-p25 |
| Homo sapiens notch
2 (NOTCH2), RefSeqGene on chromosome 1 | NG_008163.1 | 512/514 (99%) | 0 | 4853 | NOTCH2 | 1p13-p11 |
| Homo sapiens
nuclear receptor corepressor 1 (NCOR1), transcript variant 3,
mRNA | NM_001190440.1 | 213/214 (99%) | 1.00E-106 | 9611 | NCOR1 | 17p11.2 |
| Homo sapiens
retinoblastoma 1 (RB1), mRNA | NM_000321.2 | 269/270 (99%) | 1.00E-137 | 5925 | RB1 | 13q14.2 |
| Homo sapiens CREB
binding protein (CREBBP), RefSeqGene on chromosome | NG_009873.1 | 197/198 (99%) | 1.00E-96 | 1387 | CREBBP | 16p13.3 |
| Homo sapiens SMAD
family member 1 (SMAD1), transcript variant 2, mRNA | NM_001003688.1 | 262/263 (99%) | 1.00E-133 | 4086 | SMAD1 | 7p15 |
| Homo sapiens
Niemann-Pick disease, type C1 (NPC1), RefSeqGene on chromosome
18 | NG_012795.1 | 174/175 (99%) | 3.00E-83 | 4864 | NPC1 | 18q11-q12 |
| Homo sapiens
epidermal growth factor receptor (EGFR), RefSeqGene on chromosome
7 | NG_007726.1 | 207/208 (99%) | 3.00E-102 | 1956 | EGFR | 7p12 |
| Homo sapiens septin
9 (SEPT9), transcript variant 4, mRNA | NM_001113495.1 | 379/380 (99%) | 0 | 10801 SEPT9 | SEPT9 | 17q25 |
Discussion
Cancer is a complex disease that results from dozens
of genetic and epigenetic mutations. Although it is known that
cancer cells are heterogeneous, the mechanism by which cancer cells
become heterogeneous during the mulitstep process of tumorigenesis
is largely unknown. The potential role of CSCs in this process has
been the source of great interest in cancer biology and
translational cancer research since the first identification of
CSCs in leukemia by John Dick in 1997 (3), and in solid tumors by Michael Clarke
in 2003 (17). The CSC model
states that only a small subpopulation of cancer cells functions as
cancer-initiating cells with unlimited proliferation potential,
whereas the majority of cancer cells are differentiated cells with
limited proliferation capacity. Thereby, the CSC hypothesis
provides a rational explanation for the recurrence of tumors after
chemotherapy or radiation therapy, and drug resistance. Targeting
CSCs may be the key for complete inhibition or killing of cancer
cells. Thus, identification and characterization of CSCs in
different types of cancers can help our understanding of cancer
biology and anticancer drug development.
Identification of CSC markers is the first step in
CSC research. Many molecules have been used to identify a
subpopulation of cancer cell lines or tumor samples which possess
CSC properties. For example, in breast cancer both CD44 high/CD24
low and ALDH are CSC markers. CD133 is also a promising CSC marker
for brain tumors and colon cancers (4). The ALDH superfamily of
NAD(P)+-dependent multifunctional enzymes catalyze the
oxidation of various aldehydes to their corresponding carboxylic
acids. ALDH isozymes are widely distributed in tissues and organs.
ALDH was first demonstrated to be highly enriched in hematopoietic
stem cells. Stem cells isolated using Aldefluor assays have been
utilized in regenerative medicine, such as bone marrow
transplantation (14). Recently,
ALDH activity has been reported to be a CSC marker in certain solid
tumors, including breast, lung, liver, pancreas, prostate and colon
cancers. In this study, Aldefluor assays were applied to enrich a
subset of cells with CSC characteristics from the Tca8113 tongue
squamous cell carcinoma cell line. To determine the functional
differences between sorted ALDH+ and ALDH−
fractions, we compared their proliferation and found that
ALDH+ Tca8113 cells proliferated faster than
ALDH− cells in vitro.
Self-renewal and differentiation are essential
properties of CSCs. Our in vitro differentiation assays
showed that ALDH+ cells were able to differentiate into
ALDH− cells, and to maintain undifferentiated
ALDH+ parts. By contrast, ALDH− cells did not
generate ALDH+ cells under the same conditions.
Non-adherent sphere formation assays have been accepted to evaluate
putative CSC activity. This assay predicts that a CSC can be
serially passaged for many cycles and generates a tumorsphere
resembling the primary sphere. Our sphere formation and
differentiation results demonstrated that ALDH+ cells
could self-renew and differentiate to ALDH− cells. We
also observed that a small portion of sorted ALDH− cells
survived in serum-free medium and formed spheres, which may be
caused by the contamination during FACS sorting procedure.
Together, these data confirmed that ALDH activity is valuable in
identifying tongue squamous cell carcinoma stem cells. Kang et
al (7) recently reported that
CD133 may be a potential tumor-initiating marker for the Tca183
cell line. They carried out similar experiments and showed that
CD133+ cells possessed a higher ability of
proliferation, differentiation and sphere formation than
CD133− cells. It will be interesting to test whether
these two markers identify the same subpopulation of Tca183
cells.
In this study, we not only showed that ALDH activity
can serve as a marker to identify CSCs in tongue squamous cell
carcinoma, but we also compared the gene expression patterns
between isolated fractions using the suppression subtractive
hybridization assay. It was interesting to find that CD44 was one
of the 28 overexpressed genes in the ALDH+ fraction,
indicating that ALDH+ cells may overlap with the
CD44+ subpopulation. Notch2 is another upregulated gene
in ALDH+ cells. It has been demonstrated that knockdown
of Notch2 sensitizes glioma stem cells to radiation and impairs
xenograft tumor formation (16).
Further analysis of these differentially expressed genes in CSCs is
required to identify new therapeutic targets of tongue squamous
cell carcinoma.
Acknowledgements
This study was supported by grants from the
ChongQing Science and Technology Commission Project (no.
2008-2-232). The authors thank Medjaden Bioscience Limited for
assisting in the preparation of this manuscript.
References
|
1
|
RI HaddadDM ShinRecent advances in head
and neck cancerN Engl J
Med35911431154200810.1056/NEJMra070797518784104
|
|
2
|
D SanoJN MyersMetastasis of squamous cell
carcinoma of the oral tongueCancer Metastasis
Rev26645662200710.1007/s10555-007-9082-y17768600
|
|
3
|
D BonnetJE DickHuman acute myeloid
leukemia is organized as a hierarchy that originates from a
primitive hematopoietic cellNat
Med3730737199710.1038/nm0797-7309212098
|
|
4
|
JE VisvaderGJ LindemanCancer stem cells in
solid tumours: accumulating evidence and unresolved questionsNat
Rev Cancer8755768200810.1038/nrc249918784658
|
|
5
|
ME PrinceR SivanandanA
KaczorowskiIdentification of a subpopulation of cells with cancer
stem cell properties in head and neck squamous cell carcinomaProc
Natl Acad Sci USA104973978200710.1073/pnas.061011710417210912
|
|
6
|
B QiaoNW JohnsonX ChenDisclosure of a stem
cell phenotype in an oral squamous cell carcinoma cell line induced
by BMP-4 via an epithelial-mesenchymal transitionOncol
Rep26455461201121567100
|
|
7
|
FW KangK WangM WuBiological
characteristics of CD133+ subpopulation in tongue
squamous cell carcinoma Tca8113 cell lineHua Xi Kou Qiang Yi Xue Za
Zhi285605642010(In Chinese).
|
|
8
|
L DuY YangX XiaoSox2 nuclear expression is
closely associated with poor prognosis in patients with
histologically node-negative oral tongue squamous cell
carcinomaOral
Oncol47709713201110.1016/j.oraloncology.2011.05.01721689966
|
|
9
|
V HayryLK MakinenT AtulaBmi-1 expression
predicts prognosis in squamous cell carcinoma of the tongueBr J
Cancer102892897201010.1038/sj.bjc.660554420145620
|
|
10
|
B MackO GiresCD44s and CD44v6 expression
in head and neck epitheliaPLoS
One3e3360200810.1371/journal.pone.000336018852874
|
|
11
|
C GinestierMH HurE Charafe-JauffretALDH1
is a marker of normal and malignant human mammary stem cells and a
predictor of poor clinical outcomeCell Stem
Cell1555567200710.1016/j.stem.2007.08.01418371393
|
|
12
|
JP SullivanM SpinolaM DodgeAldehyde
dehydrogenase activity selects for lung adenocarcinoma stem cells
dependent on notch signalingCancer
Res7099379948201010.1158/0008-5472.CAN-10-088121118965
|
|
13
|
RW StormsAP TrujilloJB SpringerIsolation
of primitive human hematopoietic progenitors on the basis of
aldehyde dehydrogenase activityProc Natl Acad Sci
USA9691189123199910.1073/pnas.96.16.911810430905
|
|
14
|
P MarcatoCA DeanCA GiacomantonioAldehyde
dehydrogenase: its role as a cancer stem cell marker comes down to
the specific isoformCell
Cycle1013781384201110.4161/cc.10.9.1548621552008
|
|
15
|
DJ PearceD TaussigC
SimpsonCharacterization of cells with a high aldehyde dehydrogenase
activity from cord blood and acute myeloid leukemia samplesStem
Cells23752760200510.1634/stemcells.2004-029215917471
|
|
16
|
X FanL KhakiTS ZhuNOTCH pathway blockade
depletes CD133-positive glioblastoma cells and inhibits growth of
tumor neurospheres and xenograftsStem Cells28516201019904829
|
|
17
|
M Al-HajjMS WichaA
Benito-HernandezProspective identification of tumorigenic breast
cancer cellsProc Natl Acad Sci
USA10039833988200310.1073/pnas.053029110012629218
|