Introduction
Sepsis, a systemic inflammatory response often
resulting from an invasive bacterial infection, leads to
multi-organ failure and mortality. In the United States, ~750,000
individuals develop sepsis each year (1,2). The
vast majority of these individuals are at least 65 years old
(3). The mechanism by which sepsis
exerts its detrimental effects in the elderly has not been fully
elucidated. A decline in immune function and an inadequate
inflammatory response has been previously reported (4); however, the more accepted mechanism
is an uncontrolled inflammatory response which, in turn, results in
an excessive release of pro-inflammatory mediators and severe
tissue injury. These observations are consistent with previous
studies that have demonstrated a marked increase in the release of
cytokines, particularly IL-6 and TNF-α, in association with more
severe tissue injuries (5–7).
Sphingosine kinase (Sphk) is an intracellular
signaling enzyme that catalyzes the phosphorylation of sphingosine
to sphingosine-1-phosphate (S1P) (8). It is has been implicated in the
regulation of immune cells, including neutrophils, monocytes and
macrophages (9–11). Sphk-1 is emerging as an important
mediator in inflammatory responses activated by various
inflammatory stimuli, including lipopolysaccharide (LPS), TNF-α and
IL-1β (12–15), and involves the toll-like receptor
(TLR) signaling pathways. It has also been reported that Sphk-1 is
upregulated in stimulated human phagocytes and peritoneal
phagocytes of patients with severe sepsis and potentially plays a
role in the development of sepsis (16). Blockade of Sphk-1 inhibits
phagocyte production of endotoxin-induced pro-inflammatory
cytokines, consistent with a study in which mice were pretreated
with a Sphk-1 inhibitor following LPS administration, which
revealed a decrease in TNF-α, IL-6, MCP-1 and HMGB1 (16). In addition, Sphk-1 and S1P have
been demonstrated as important inflammatory mediators in asthma,
rheumatoid arthritis and inflammatory bowel disease (IBD) (17–19).
Sphk-1 expression has been found to increase in IBD and in a mouse
model of colitis and Sphk-1 gene deficiency markedly decreased the
systemic inflammatory response (20). Recently, expression of Sphk-1 and
its activity were found to be markedly increased in peripheral
immune cells of patients in the early stages of severe acute
pancreatitis, indicating that the regulation of the Sphk-1 pathway
may represent a novel target in the treatment of this disease
(21).
Our previous studies demonstrated that levels of
pro-inflammatory mediators were significantly elevated in aged
animals following the induction of endotoxemia (6,7).
Therefore, in the aged population, alterations in the immune
response may contribute to a higher mortality rate following
bacterial infection. The molecular mediators associated with this
hyperinflammatory response are not clearly defined in the aged
population. To determine whether Sphk-1 represents a molecular
mediator associated with the observed hyperinflammatory response in
aging, Sphk-1 mRNA expression was examined by quantitative PCR in
hepatic tissues of young and aged rats subjected to endotoxemia,
and its expression was found to correlate with CD14, a known
component of the LPS receptor and the TLR signaling pathways.
Materials and methods
Experimental animals
Male Fischer-344 rats (young, 3 months old; aged, 24
months old) were obtained from the National Institute on Aging
(Bethesda, MD, USA), housed in a temperature-controlled room on a
12-h light/dark cycle and fed a standard Purina rat chow diet.
Experiments were performed in adherence with the National
Institutes of Health Guidelines for the Use of Experimental
Animals. This project was approved by the Institutional Animal Care
and Use Committee of The Feinstein Institute for Medical Research
(Manhasset, NY, USA).
Cell culture and treatment
Kupffer cells were isolated separately from
6–8-week-old normal Sprague-Dawley rats by collagenase perfusion of
the liver followed by Percoll gradient centrifugation as described
previously (22). Kupffer cells
were plated at 1×106 cells/well in 6-well culture plates
with DMEM containing 10% FBS and incubated overnight at 37°C. All
media were supplemented with 10 mM HEPES (pH 7.4), 2 mM
L-glutamine, 100 U/ml penicillin and 100 μg/ml streptomycin. Cells
were incubated overnight in a 37°C incubator with 5%
CO2. Cells were then washed with media and treated with
100 ng/ml LPS for 24 h.
Induction of endotoxemia in rats
Endotoxemia was induced by intravenous
administration of LPS, as described previously (6,7).
Prior to the induction of severe endotoxemia, rats were fasted
overnight, but allowed water ad libitum. Rats were then
anesthetized with isoflurane inhalation, inguinal regions were
shaved and washed with 10% povidone-iodine and a short subinguinal
incision was made. The femoral vein was carefully separated from
the artery and cannulated with a catheter (PE-50 tubing). A bolus
injection of LPS (15 mg/kg; E. coli 055:B5 in 200 μl normal
saline; Sigma-Aldrich, St. Louis, MO, USA) was administered through
the femoral vein catheter. The same surgery was performed on the
vehicle control animals, but the control was injected with normal
saline instead of LPS. Tissue samples were collected at 4 h
following LPS injection.
Real-time PCR (Q-PCR) analysis
Sphk-1, TNF-α and CD14 gene expression was
determined by Q-PCR. Total RNA was extracted from hepatic tissue
using TRIzol reagent (Invitrogen Life Technologies, Carlsbad, CA,
USA). Q-PCR was performed on cDNA samples reverse transcribed from
2 μg RNA using murine leukemia virus reverse transcriptase (Applied
Biosystems, Foster City, CA, USA). Using a SYBR Green PCR Master
mix (Applied Biosystems, Foster City, CA, USA), reactions were
performed in a 24-μl final volume containing 0.08-μmol each forward
and reverse primer, 2 μl cDNA, 9.2 μl H2O and 12 μl SYBR
Green PCR Master mix. Amplification was performed using the Applied
Biosystems 7300 Real-Time PCR machine under the thermal profile of
50°C for 2 min and 95°C for 10 min followed by 55 cycles at 95°C
for 15 sec and 60°C for 1 min. Rat GAPDH mRNA expression was used
to normalize each sample and analysis of each specific mRNA was
conducted in duplicate. Relative expression of mRNA was calculated
by the 2−ΔΔCt method and results were expressed as fold
change with respect to the corresponding experimental control. The
rat primers used in the experiment are listed in Table I. To assess the specificity of the
PCR products, a melting curve analysis was performed in each Q-PCR
experiment. No non-specific products from any of the primers used
in our experiments were detected.
 | Table IRat primers used in the present
study. |
Table I
Rat primers used in the present
study.
| Gene | Forward | Reverse |
|---|
| Sphk-1 |
TGCCTTCTCATTGGACTGTGG |
GTAGCAGCACCAGCACCAG |
| CD14 |
TGGGCGAGAAAGGACTGATC |
GGAGGGTCGGGAATTTGTG |
| TNF-α |
TGATCGGTCCCAACAAGGA |
GGGCCATGGAACTGATGAGA |
| GAPDH |
ATGACTCTACCCACGGCAAG |
CTGGAAGATGGTGATGGGTT |
Measurement of TNF-α protein levels
Supernatants from Kupffer cells treated with LPS
were measured for TNF-α protein levels using specific ELISA kits
(BD Pharmingen, Franklin Lakes, NJ, USA).
Statistical analysis
Data are expressed as the mean ± SEM and compared by
one-way analysis of variance and the Student-Newman-Keuls Method
for multiple groups and Student’s t-test for two groups. P<0.05
was considered to indicate a statistically significant
difference.
Results
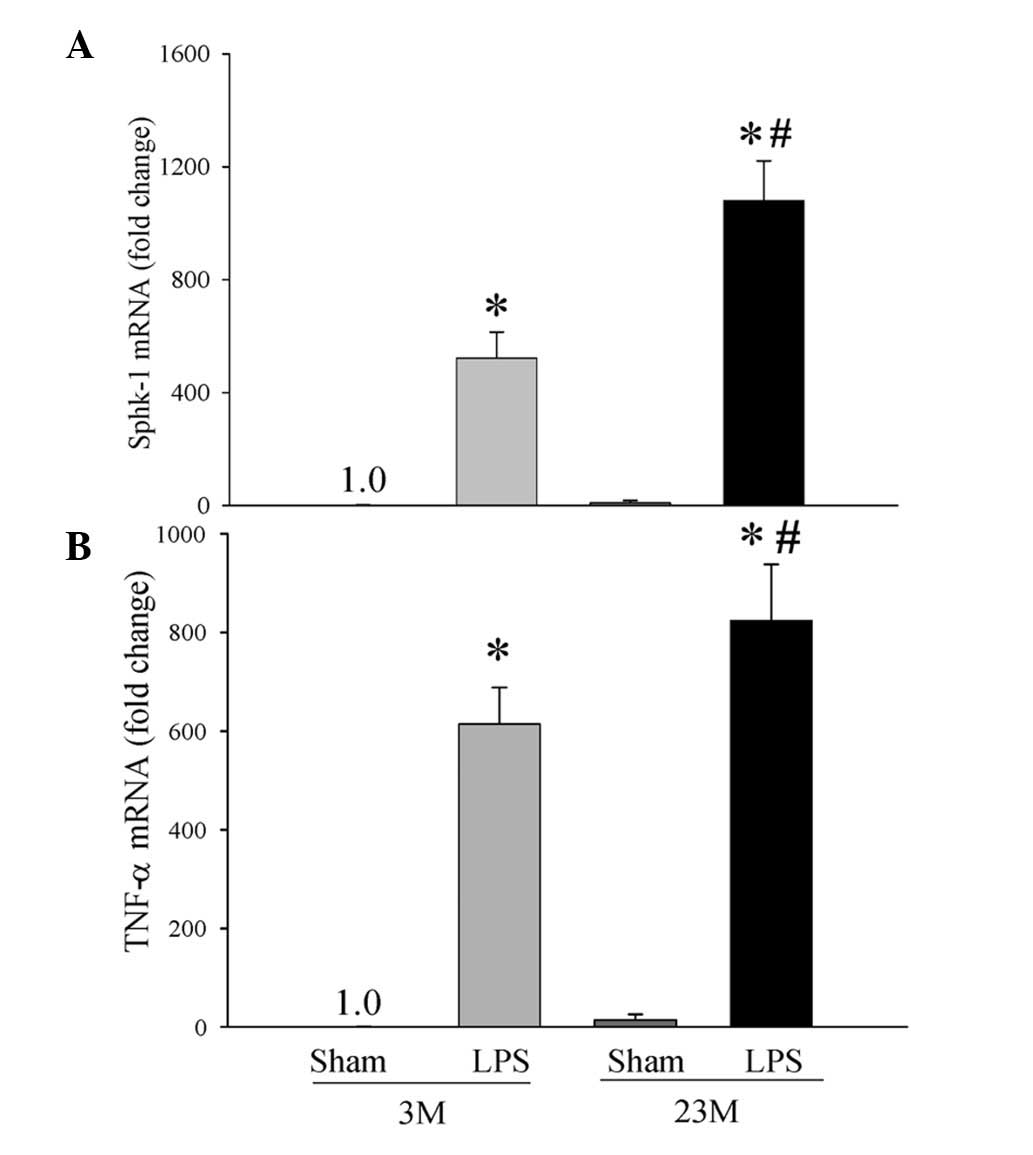
Sphk-1, TNF-α and CD14 mRNA expression in
the liver of young and aged endotoxemic rats
We have previously demonstrated that the
inflammatory response is exacerbated in a rodent endotoxemia model
of sepsis in aged rats compared with young rats (6). To determine the mechanism by which
the inflammatory response is exacerbated in aged rats in comparison
with young rats, we examined gene expression of Sphk-1 in hepatic
tissues of the young and aged animals during endotoxemia. As
revealed in Fig. 1, gene
expression of Sphk-1 in hepatic tissues was significantly increased
following endotoxemia when compared with their respective Sham
groups. There was a significant 2-fold increase in Sphk-1 mRNA
expression in endotoxemic aged rats compared with that of young
rats (1,080±141 vs. 522±92; P<0.001, Fig. 1A). The increase in Sphk-1 was
correlated with a significant 1.3-fold increase in TNF-α gene
expression during endotoxemia in the aged animals compared with
young rats (824±113 vs. 614±74; P<0.05, Fig. 1B).
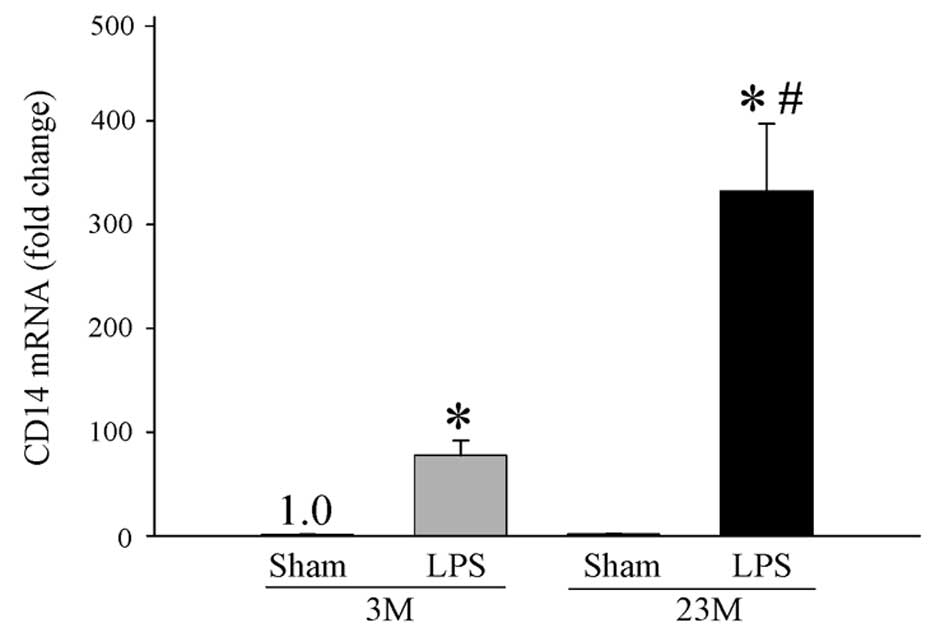
CD14, a known LPS receptor component, is important
for the production of proinflammatory cytokines, including TNF-α,
IL-1β and IL-6 (23,24). Gene expression of CD14 was
significantly increased in young and aged endotoxemic rats compared
with their respective Sham groups (Fig. 2). Of note, gene expression of CD14
was significantly increased by 4-fold during endotoxemia in the
aged animals in comparison with the young animals (332±65 vs.
77±14; P<0.001, Fig. 2). These
observations indicate a direct positive correlation among elevated
Sphk-1, TNF-α and CD14 gene expression in hepatic tissues of
endotoxemic aged rats.
Sphk-1, CD14 and TNF-α mRNA expression in
LPS-treated Kupffer cells
To further delineate the signaling pathway
responsible for the increased susceptibility of inflammation in the
aged animals, mRNA from Kupffer cells was examined for Sphk-1, CD14
and TNF-α expression. Kupffer cells are specialized macrophages
located in the liver. These cells line the walls of the sinusoids
that form the reticuloendothelial system. Kupffer cells are a
critical component of the innate immune system. As demonstrated in
Fig. 3A, gene expression of Sphk-1
was increased by 3-fold while gene expression of CD14 was increased
by 6.6-fold following treatment with LPS (Fig. 3B). TNF-α mRNA and protein
expression was also significantly increased. These results indicate
that increased expression of CD14 upregulates Sphk-1 in Kupffer
cells and leads to increased TNF-α production, and thus mediates
the hyperinflammatory response during endotoxemia in aging.
Discussion
Sepsis is a major cause of morbidity and mortality
in the geriatric population and ~60% of all sepsis cases occur in
patients older than 65 years of age (3). Aging studies in human and animal
models have revealed changes in a number of aspects of immunity
(25–27). It has been hypothesized that the
increased mortality rate of sepsis in the geriatric population is a
direct result of an impaired immune response. We have previously
reported that endotoxemia causes an in vivo
hyperinflammatory state in aged animals (6,7).
This was characterized by an increase in circulating and splenic
levels of proinflammatory cytokines. A previous study demonstrated
that Sphk-1 regulates proinflammatory responses associated with
endotoxin and polymicrobial sepsis (16). In the current study, we focused on
the potential role of Sphk-1 in the hyperinflammatory response
observed in aged animals subjected to endotoxemia.
Sphk is an intracellular signaling enzyme that
catalyzes the phosphorylation of sphingosine to S1P (8). A number of growth factors and
cytokines have been demonstrated to activate Sphk, including
platelet-derived growth factor, TNF-α and IL-1β (12,13,28).
In the present study, the hepatic tissues of young and aged animals
were examined to determine if Sphk-1 mRNA expression differed in
these groups. LPS-treated aged animals were found to exhibit
increased expression of Sphk-1 mRNA in hepatic tissues indicating a
role for Sphk-1 in the hyperinflammatory response observed in the
aged. Consistent with previous studies, increased Sphk-1 mRNA
levels were also observed in LPS-treated animals in comparison with
respective Sham groups. The exact signaling pathway by which Sphk-1
functions in association with inflammation has been studied by
Puneet et al and has been hypothesized to involve TLR
signaling. The authors found that LPS and bacterial lipoprotein
markedly upregulates Sphk-1 expression. Sphk-1 acts through the
second messenger, S1P, ultimately leading to the activation of
NF-κB. NF-κB then leads to the release of TNF-α, IL-6 and IL-1β,
and systemic inflammation (16).
This previous study indicated a direct correlation between Sphk-1
and proinflammatory cytokines. Results of the present study are
consistent with this proposed signaling pathway, whereby
concomitant increases in Sphk-1, CD14 and TNF-α were identified in
hepatic tissues of animals subjected to endotoxemia. Of note,
increased levels of these proinflammatory mediators were found in
the aged animal population. Thus, it appears that Sphk-1 may play a
significant role in sepsis in the elderly by increasing the
expression of proinflammatory cytokines.
To determine the cell type in the liver responsible
for the observed increase in Sphk-1, we focused on Kupffer cells,
which form part of the reticuloendothelial system and resident
macrophages of the liver. These cells play a vital role in the
innate immune response through the upregulation and release of
proinflammatory cytokines (5).
Increased Sphk-1 mRNA expression was identified in LPS-treated
Kupffer cells compared with control samples. To further
characterize the mechanism of action of Sphk-1, the expression of
CD14 mRNA was studied in these cells. A statistically significant
increase in expression of CD14 was found in LPS-treated cells.
These results correlate with our in vivo observations with
regards to a potential role of Sphk-1 in sepsis in the aged.
The role of CD14 as a key LPS signaling component
has been well documented in a number of cell systems, including
monocytes and macrophages (29–31).
In the liver tissue, it has been previously reported that CD14
transcription rates significantly increased in the hepatocytes of
LPS-treated rats (32,33). In addition, CD14 has been found to
be expressed in an LPS-inducible manner in Kupffer cells and
sinusoidal endothelial cells (34). Consistent with our observations, a
previous study revealed that hepatic CD14 upregulation led to
increased endotoxin sensitivity and host proinflammatory reactions,
causing organ failure and mortality in a rat model of cholestasis
(34,35). In addition, it has been
hypothesized that the high levels of CD14 expression observed in
Kupffer cells may increase proinflammatory responses and lead to
increased endotoxin-induced mortality (36).
In summary, in the present study, increased
expression of Sphk-1 was identified in young and aged animals
subjected to endotoxemia compared with their respective Sham
groups. Of note, the aged population was found to exhibit a
significantly higher level of expression. This increased expression
of Sphk-1 in the aged population corresponded with significantly
higher levels of CD14 and TNF-α in the hepatic tissues. Increased
expression of Sphk-1 and CD14 was also observed in vitro in
Kupffer cells treated with LPS. While there was a 4-fold increase
in CD14 expression, Sphk-1 only increased by 2-fold indicating that
additional signaling components are likely to be involved in the
hyperinflammatory state associated with endotoxemia in the aged.
However, results of the current study collectively indicate that
the hyperinflammatory state previously observed during endotoxemia
in the aged may, in part, be due to the increase in Kupffer cell
CD14 expression, leading to increased Sphk-1 and subsequent
increases in TNF-α. Therefore, it is indicated that Sphk-1
contributes to age-related hyperinflammation in endotoxemia.
Acknowledgements
The present study was supported by grants from the
National Institutes of Health (nos. R01 AG028352-05, R01 GM053008
and R01 GM057468). The authors thank Weifeng Dong, MS, Monowar
Aziz, PhD, Rongqian Wu, MD, PhD, and Michael A. Ajakaiye, MD, for
their expert assistance.
References
|
1
|
Angus DC, Linde-Zwirble WT, Lidicker J,
Clermont G, Carcillo J and Pinsky MR: Epidemiology of severe sepsis
in the United States: analysis of incidence, outcome and associated
costs of care. Crit Care Med. 29:1303–1310. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Martin GS, Mannino DM, Eaton S and Moss M:
The epidemiology of sepsis in the United States from 1979 through
2000. N Engl J Med. 348:1546–1554. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Martin GS, Mannino DM and Moss M: The
effect of age on the development and outcome of adult sepsis. Crit
Care Med. 34:15–21. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Renshaw M, Rockwell J, Engleman C, Gewirtz
A, Katz J and Sambhara S: Cutting edge: impaired Toll-like receptor
expression and function in aging. J Immunol. 169:4697–4701. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Tateda K, Matsumoto T, Miyazaki S and
Yamaguchi K: Lipopolysaccharide-induced lethality and cytokine
production in aged mice. Infect Immun. 64:769–774. 1996.PubMed/NCBI
|
|
6
|
Wu R, Zhou M, Dong W, et al: Ghrelin
hyporesponsiveness contributes to age-related hyperinflammation in
septic shock. Ann Surg. 250:126–133. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Zhou M, Wu R, Dong W, Leong J and Wang P:
Accelerated apoptosis contributes to aging-related
hyperinflammation in endotoxemia. Int J Mol Med. 25:929–935.
2010.PubMed/NCBI
|
|
8
|
Spiegel S and Milstien S:
Sphingosine-1-phosphate: an enigmatic signalling lipid. Nat Rev Mol
Cell Biol. 4:397–407. 2003. View
Article : Google Scholar : PubMed/NCBI
|
|
9
|
Allende ML, Sasaki T, Kawai H, et al: Mice
deficient in sphingosine kinase 1 are rendered lymphopenic by
FTY720. J Biol Chem. 279:52487–52492. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Egger M, Beer AG, Theurl M, et al:
Monocyte migration: a novel effect and signaling pathways of
catestatin. Eur J Pharmacol. 598:104–111. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Matloubian M, Lo CG, Cinamon G, et al:
Lymphocyte egress from thymus and peripheral lymphoid organs is
dependent on S1P receptor 1. Nature. 427:355–360. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Xia P, Gamble JR, Rye KA, et al: Tumor
necrosis factor-alpha induces adhesion molecule expression through
the sphingosine kinase pathway. Proc Natl Acad Sci USA.
95:14196–14201. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Billich A, Bornancin F, Mechtcheriakova D,
Natt F, Huesken D and Baumruker T: Basal and induced sphingosine
kinase 1 activity in A549 carcinoma cells: function in cell
survival and IL-1beta and TNF-alpha induced production of
inflammatory mediators. Cell Signal. 17:1203–1217. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Alvarez SE, Milstien S and Spiegel S:
Autocrine and paracrine roles of sphingosine-1-phosphate. Trends
Endocrinol Metab. 18:300–307. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Melendez AJ: Sphingosine kinase signalling
in immune cells: potential as novel therapeutic targets. Biochim
Biophys Acta. 1784:66–75. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Puneet P, Yap CT, Wong L, et al: SphK1
regulates proinflammatory responses associated with endotoxin and
polymicrobial sepsis. Science. 328:1290–1294. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Deguchi Y, Andoh A, Yagi Y, et al: The S1P
receptor modulator FTY720 prevents the development of experimental
colitis in mice. Oncol Rep. 16:699–703. 2006.PubMed/NCBI
|
|
18
|
Kitano M, Hla T, Sekiguchi M, et al:
Sphingosine 1-phosphate/sphingosine 1-phosphate receptor 1
signaling in rheumatoid synovium: regulation of synovial
proliferation and inflammatory gene expression. Arthritis Rheum.
54:742–753. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Lai WQ, Goh HH, Bao Z, Wong WS, Melendez
AJ and Leung BP: The role of sphingosine kinase in a murine model
of allergic asthma. J Immunol. 180:4323–4329. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Snider AJ, Kawamori T, Bradshaw SG, et al:
A role for sphingosine kinase 1 in dextran sulfate sodium-induced
colitis. Faseb J. 23:143–152. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Li Q, Wang C, Zhang Q, Tang C, Li N and Li
J: The role of sphingosine kinase 1 in patients with severe acute
pancreatitis. Ann Surg. 255:954–962. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Jacob A, Zhou M, Wu R, Halpern VJ,
Ravikumar TS and Wang P: Pro-inflammatory cytokines from Kupffer
cells downregulate hepatocyte expression of adrenomedullin binding
protein-1. Biochim Biophys Acta. 1772:766–772. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Lu YC, Yeh WC and Ohashi PS: LPS/TLR4
signal transduction pathway. Cytokine. 42:145–151. 2008. View Article : Google Scholar
|
|
24
|
Scott MJ, Liu S, Su GL, Vodovotz Y and
Billiar TR: Hepatocytes enhance effects of lipopolysaccharide on
liver nonparenchymal cells through close cell interactions. Shock.
23:453–458. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Weiskopf D, Weinberger B and
Grubeck-Loebenstein B: The aging of the immune system. Transpl Int.
22:1041–1050. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Grolleau-Julius A, Ray D and Yung RL: The
role of epigenetics in aging and autoimmunity. Clin Rev Allergy
Immunol. 39:42–50. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Sawhney M, Mathew M, Valarmathi MT and Das
SN: Age related changes in Fas (CD95) and Fas ligand gene
expression and cytokine profiles in healthy Indians. Asian Pac J
Allergy Immunol. 24:47–56. 2006.PubMed/NCBI
|
|
28
|
Olivera A and Spiegel S:
Sphingosine-1-phosphate as second messenger in cell proliferation
induced by PDGF and FCS mitogens. Nature. 365:557–560. 1993.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Gluck T, Silver J, Epstein M, Cao P,
Farber B and Goyert SM: Parameters influencing membrane CD14
expression and soluble CD14 levels in sepsis. Eur J Med Res.
6:351–358. 2001.PubMed/NCBI
|
|
30
|
Pan Z, Zhou L, Hetherington CJ and Zhang
DE: Hepatocytes contribute to soluble CD14 production and CD14
expression is differentially regulated in hepatocytes and
monocytes. J Biol Chem. 275:36430–36435. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Schutt C: Cd14. Int J Biochem Cell Biol.
31:545–549. 1999. View Article : Google Scholar
|
|
32
|
LeVan TD, Bloom JW, Bailey TJ, et al: A
common single nucleotide polymorphism in the CD14 promoter
decreases the affinity of Sp protein binding and enhances
transcriptional activity. J Immunol. 167:5838–5844. 2001.
View Article : Google Scholar
|
|
33
|
Liu S, Shapiro RA, Nie S, Zhu D, Vodovotz
Y and Billiar TR: Characterization of rat CD14 promoter and its
regulation by transcription factors AP1 and Sp family proteins in
hepatocytes. Gene. 250:137–147. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Chou MH, Chuang JH, Eng HL, et al:
Endotoxin and CD14 in the progression of biliary atresia. J Transl
Med. 8:1382010. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Chou MH, Chuang JH, Eng HL, et al: Effects
of hepatocyte CD14 upregulation during cholestasis on endotoxin
sensitivity. PLoS One. 7:e349032012. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Sewnath ME, Levels HH, Oude Elferink R, et
al: Endotoxin-induced mortality in bile duct-ligated rats after
administration of reconstituted high-density lipoprotein.
Hepatology. 32:1289–1299. 2000. View Article : Google Scholar : PubMed/NCBI
|