Introduction
Glioma is a type of central nervous system (CNS)
cancer that is associated with a high mortality rate. Glioma has a
particularly poor prognosis, the worst of all CNS cancer types,
aside from brain stem glioma. It is considered that the various
characteristics of glioma, including high invasion and wide range
of metastasis, are associated with epithelial-mesenchymal
transition (EMT) (1,2).
EMT is a basic physiological process in
multicellular organism morphology. It refers to the
transdifferentiation phenomenon of epithelial cells to mesenchymal
cells. In this process, epithelial cells gradually obtain the
mesenchymal phenotype by losing polarity and acquiring mobility.
EMT is mainly characterized by epithelial cell loss of polarity and
the acquisition of mesenchymal characteristics. Specifically, this
process includes: Cellular adhesion-related gene expression was
reduced, causing epithelial cells to lose their intercellular
adhesion capability. This induces structural changes in cellular
keratin, which in turn transforms the morphology of the epithelial
cells to a spindle fibroblast shape. The cells then acquire the
characteristics of mesenchymal or fibroblast cells, including
expression of vimentin, N-cadherin and fibronectin. EMT is involved
in the occurrence and development of epithelial origin tumors,
particularly in the process of cancer invasion and metastasis.
Therefore, investigating the role of EMT in the formation and
metastasis of tumors, as well as its regulatory mechanisms, has
attracted notable attention in recent years.
microRNA (miRNA) is a type of non-coding
single-stranded small molecule RNA with 21–25 nucleotides. The main
function of microRNA is to regulate the expression of genes through
translation inhibition, mRNA lysis and deadenylation. Furthermore,
it has been reported to have numerous effects on cell
proliferation, survival, apoptosis and cell cycle. With the
advancement of the studies on miR-16, numerous antitumor
characteristics of this miRNA have been revealed, including as a
genetic therapy tool for tumor treatment. The present study
investigated the inhibitory effects of miR-16 on EMT-related gene
expression of U87 and U251 glioma cells and provided evidence that
has potential applications for the development of novel genetic
tumor therapies using miR-16.
Materials and methods
Cell culture and transfection
The U87 and U251 human glioma cell line were
obtained from the American Type Culture Collection (Manassas, VA,
USA). The cells were cultured in Dulbecco’s modified Eagle’s medium
(DMEM containing 10% fetal bovine serum) under the conditions of 5%
CO2, 37°C and saturated humidity. The cells were
digested by 0.25% trypsin-EDTA.
Pre-miR-16 and scrambled negative control RNA
(pre-miR-NC) were purchased from GenePharma and transfected by
Lipofectamine 2000 (Invitrogen Life Technologies, Carslbad, CA,
USA) according to the manufacturer’s instructions. The miR-16
quantification assays were conducted using an mirVana™ qRT-PCR
miRNA Detection kit (Ambion, Austin, TX, USA) according to the
manufacturer’s instructions. The groups were as follows: (i) U87 or
U215 cells without treatment (control group); (ii) U87 or U215
cells transfected with negative control RNA (pre-miR-NC group);
(iii) U87 or U215 cells transfected with pre-miR-16 (pre-miR-16
group).
Transwell assay to detect the cell
invasion potential of human glioma cell lines
A total of 3×105 cells were seeded into
the upper compartment of the Transwell kit (Coster, South Elgin,
IL, USA) with Matrigel (BD Biosciences, Franklin Lakes, NJ, USA),
600 μl 0.1% bovine serum albumin medium was added to the lower
compartment and cells were cultured with 5% CO2 at 37°C
for 24 h. Then, the cells were washed three times with cold
phosphate-buffered saline, fixed, stained with hematoxylin and
eosin and counted in at least five areas using a CX31 microscope
(Olympus Corporation, Shimadzu, Japan) at a magnification of
×100.
Cell adhesion assay to detect the cell
adhesion potential of human glioma cell lines
A total of 5×105 cells were seeded into
96-well plates and incubated with 5% CO2 in 37°C for 2
h. The medium was discarded and the wells were washed four times in
PBS. MTS dye (Promega Corporation, Madison, WI, USA) was added to
the wells and the adhering cell rate was measured at 490 nm by a
Bio-Rad microplate-reader 4 h later (Bio-Rad Laboratories,
Hercules, CA, USA).
Flow cytometric assay to detect the cell
cycle of human glioma cell lines
A total of 3×105 cells were seeded into
6-well plates and incubated with 5% CO2 at 37°C for 24
h. The cells were then collected and fixed with 75% cold alcohol at
4°C overnight. Then, the cells were centrifuged at 1,000 × g for 5
min and incubated with RNase A and PI for 30 min. The cell cycle
stage of the tumor cells was measured by flow cytometry (FACSAria;
BD Biosciences) at 488 nm.
Enzyme-linked immunosorbent assay (ELISA)
assay to detect the interleukin (IL)-6, IL-8 and transforming
growth factor (TGF)-β production in human glioma cell lines
A total of 3×105 cells were seeded into
6-well plates and incubated with 5% CO2 at 37°C for 24
h. The medium was replaced with serum free DMEM and was incubated
for an additional 24 h. Then, the medium was collected and
centrifuged at 1,000 × g for 5 min. The IL-6, IL-8 and TGF-β
production in tumor cells was measured by a human ELISA kit
(R&D Systems, Inc., Minneapolis, MN, USA) at 490 nm.
Western blot analysis
The cells were lysed and centrifuged at 4°C and
14,000 × g for 5 min, and the concentration of the extracted
protein was measured using a Coomassie (Bradford) protein assay kit
(Pierce Biotechnology, Inc., Rockford, IL, USA). The protein was
separated by 12% SDS-PAGE and transferred onto a nitrocellulose
membrane. The monoclonal antibodies against E-cadherin, vimentin,
β-catenin, p-FAK and p-Akt were diluted at a ratio of 1:500 (Santa
Cruz Biotechnology, Inc., Santa Cruz, CA, USA), and the β-actin
monoclonal antibody was diluted at a ratio of 1:2,000 (Sigma, St.
Louis, MO, USA) as an internal control at 4°C overnight. The
membrane was washed using Tris-buffered saline with 0.1% Tween-20
and was added to 1:1,000 diluted horseradish peroxidase
(HRP)-conjugated goat anti-rabbit secondary antibody (Santa Cruz
Biotechnology, Inc.) at room temperature for 1 h. Following
washing, the membrane was visualized with the Phototope-HRP Western
Blot Detection system (Cell Signaling Technology, Inc., Danvers,
MA, USA).
Quantitative (q)PCR assay
The cells were collected and the total RNA was
extracted by TRIzol (Invitrogen Life Technologies). GAPDH was used
as the internal control. The total RNA was amplified using the
following primers: Sense: 5′-ACAGCCCCGCCTTATGATT-3′ and antisense:
5′-TCGGAACCGCTTCCTTCA-3′ for E-cadherin; sense:
5′-TGGCACGTCTTGACCTTGAA-3′ and antisense: 5′-GGT
CATCGTGATGCTGAGAA-3′ for vimentin; sense: 5′-TTT
GAAGGCAGTCTGTCGTA-3′ and antisense: 5′-TTATGC
TTTGCACTACGTCCCTCCA-3′ for β-catenin; and sense:
5′-ATGGAAATCCCATCACCATCTT-3′ and antisense: 5′-CGCCCCACTTGATTTGG-3′
for GAPDH. The amplification conditions were: Pre-denaturation,
94°C for 5 min, a total of 35 cycles of amplification were
conducted and each cycle consisted of 5 sec at 95°C and 35 sec at
65°C.
Report gene assay
pNF-κB-luc, pSlug-luc and the CMV promoter
Renilla luciferase internal standard (Beyotime, Shanghai,
China) were transfected by Lipofectamine 2000 (Invitrogen Life
Technologies, Carlsbad, CA, USA) to detect the transcriptional
activity of nuclear factor (NF)-κB and Slug according to
manufacturer’s instructions. The ratio between the number of
firefly luciferase excitation photon and the number of
Renilla luciferase excitation photons was calculated.
Statistical analysis
The experimental data is expressed as the mean ±
standard deviation and was analyzed using SPSS 13.0 software (SPSS,
Inc., Chicago, IL, USA). One-way analysis of variance was applied
to compare the data of different groups and P<0.05 was
considered to indicate a statistically significant difference. Each
experiment was repeated three times to obtain similar results.
Result
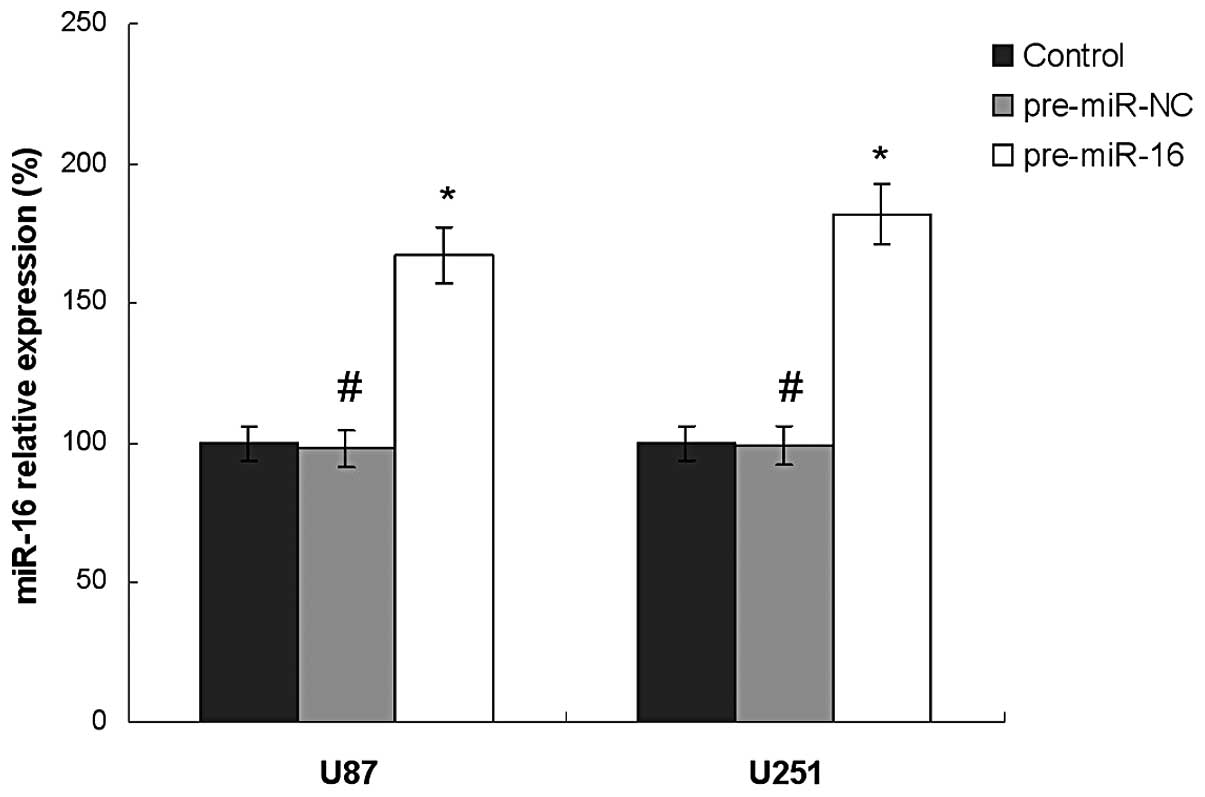
Expression of miR-16 in human glioma cell
lines
The miR-16 expression levels in these cell lines
were measured by qPCR (Fig. 1).
The results showed that there was an increase in the expression
levels of miR-16 in the pre-miR-16 group compared with those in the
control group in U87 and U251 cell lines. The expression of miR-16
in U87 pre-miR-16 and U251 pre-miR-16 groups were 173.2 and 187.6%
compared with that of the control group, respectively.
miR-16 regulates the invasion and
adhesion potential of human glioma cell lines in vitro
As demonstrated in Fig.
2, miR-16 overexpression reduced the invasion and adhesion
potential of U87 and U251 cells. The invasion rate of the U87
pre-miR-16 and U251 pre-miR-16 groups were 50.0 and 41.6% compared
with the control group, respectively. The adhesion rate of the U87
pre-miR-16 and U251 pre-miR-16 groups were 52.6 and 48.2% compared
with the control group, respectively. miR-16 overexpression also
arrested the cell cycle in the G0/G1 phase, as revealed by the
significant increase in the percentage of cells in the G0/G1 phase
in the U87 pre-miR-16 and U251 pre-miR-16 groups, at 61.5 and 66.3%
compared with the control group values of 53.2 and 47.3%,
respectively.
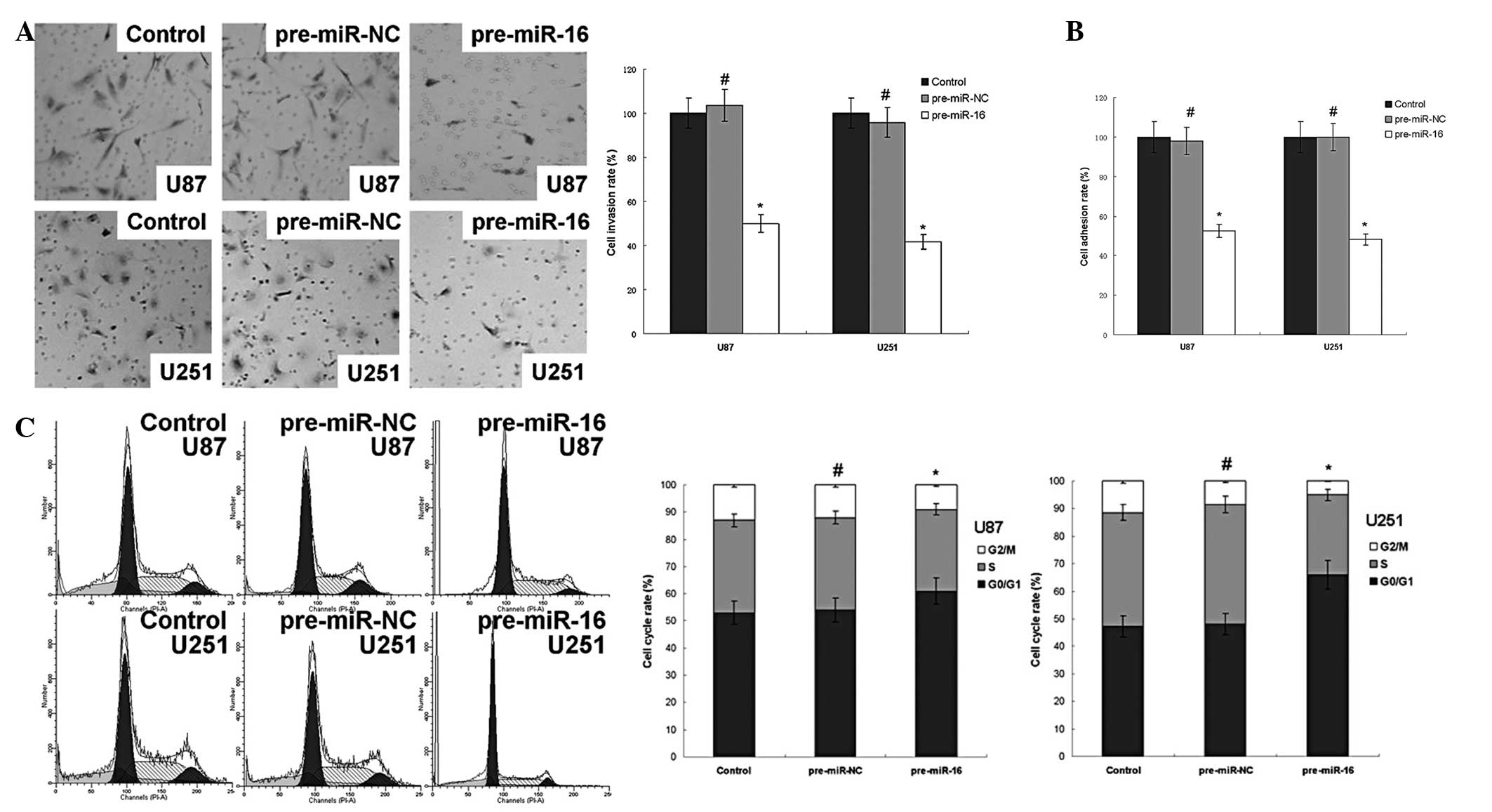 | Figure 2miR-16 regulates invasion and adhesion
potential of human glioma cell lines in vitro. (A) The
transwell assay results demonstrated that the invasion rate of the
U87 pre-miR-16 and U251 pre-miR-16 groups were 50.0 and 41.6%
compared with the control group, respectively. The values are
presented as the mean ± SD, n=3. (B) The adhesion rates of the U87
pre-miR-16 and U251 pre-miR-16 groups were 52.6 and 48.2% compared
with control group, respectively. The values are presented as the
mean ± SD, n=5. (C) The G0/G1 phase of U87 pre-miR-16 and U251
pre-miR-16 groups were 61.5 and 66.3% and increased significantly
compared with the control group at 53.2 and 47.3%, respectively.
The values are presented as the mean ± SD, n=3.
#P<0.05 and *P<0.05, compared with the
control group. miR-16, microRNA-16; ELISA, enzyme-linked
immunosorbent assay; pre-miR-NC, scrambled negative control RNA;
SD, standard deviation. |
miR-16 regulates IL-6, IL-8 and TGF-β
production of human glioma cell lines in vitro
As demonstrated in Fig.
3, miR-16 overexpression reduced the IL-6, IL-8 and TGF-β
production of U87 and U251 cells. The IL-6 production of the U87
pre-miR-16 and U251 pre-miR-16 groups was 67.5 and 44.6% compared
with the control group, respectively. The IL-8 production of the
U87 pre-miR-16 and U251 pre-miR-16 groups were 62.3 and 46.8%, and
the TGF-β production of the U87 pre-miR-16 and U251 pre-miR-16
groups were 59.5 and 30.3% compared with the control group,
respectively.
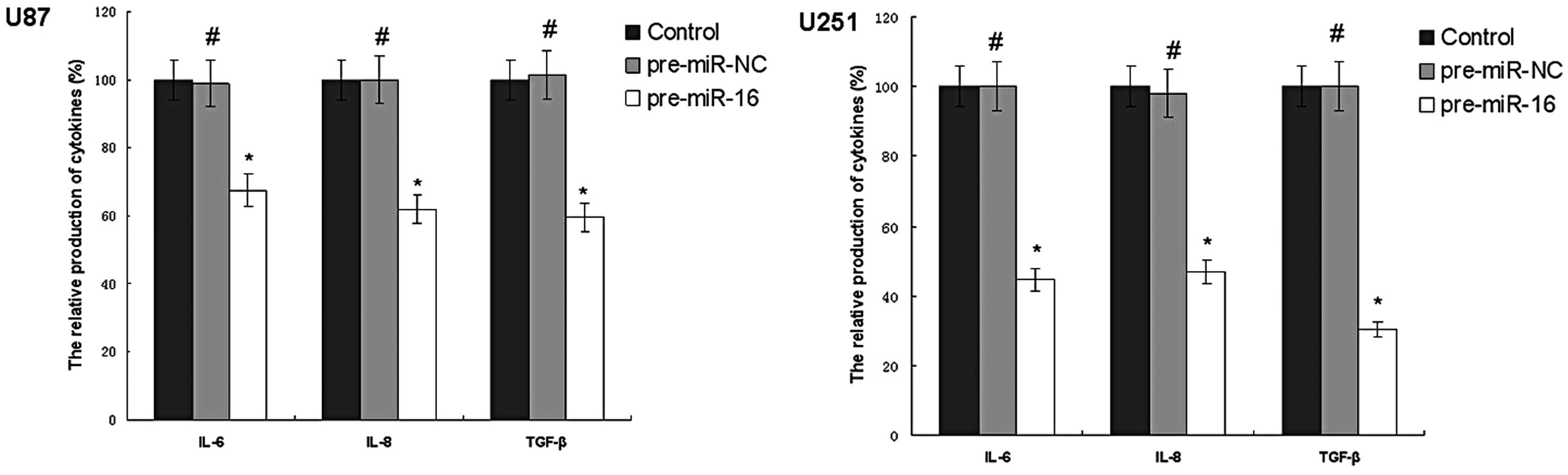 | Figure 3miR-16 regulates IL-6, IL-8 and TGF-β
production in human glioma cell lines in vitro. The ELISA
results demonstrated that overexpressing miR-16 reduced the IL-6,
IL-8 and TGF-β production of U87 cells. The IL-6 production of the
U87 pre-miR-16 and U251 pre-miR-16 groups was 67.5 and 44.6%, the
IL-8 production of the U87 pre-miR-16 and U251 pre-miR-16 groups
was 62.3 and 46.8%, the TGF-β production of the U87 pre-miR-16 and
U251 pre-miR-16 groups was 59.5 and 30.3% compared with control
group, respectively. The values are presented as the mean ±
standard deviation, n=5. #P<0.05 and
*P<0.05, compared with the control group. miR-16,
microRNA-16; IL, interleukin; TGF, transforming growth factor;
pre-miR-NC, scrambled negative control RNA; ELISA, enzyme-linked
immunosorbent assay. |
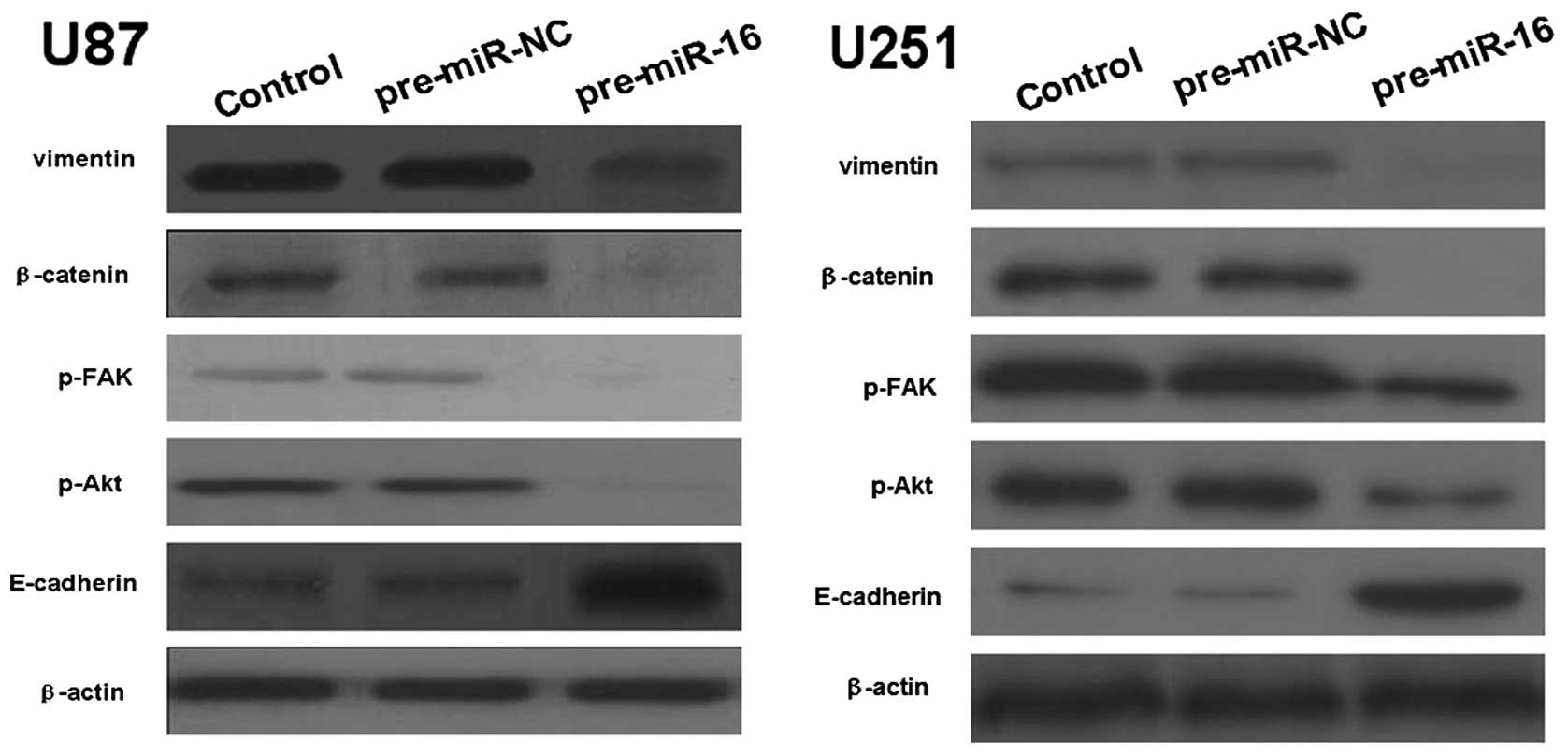
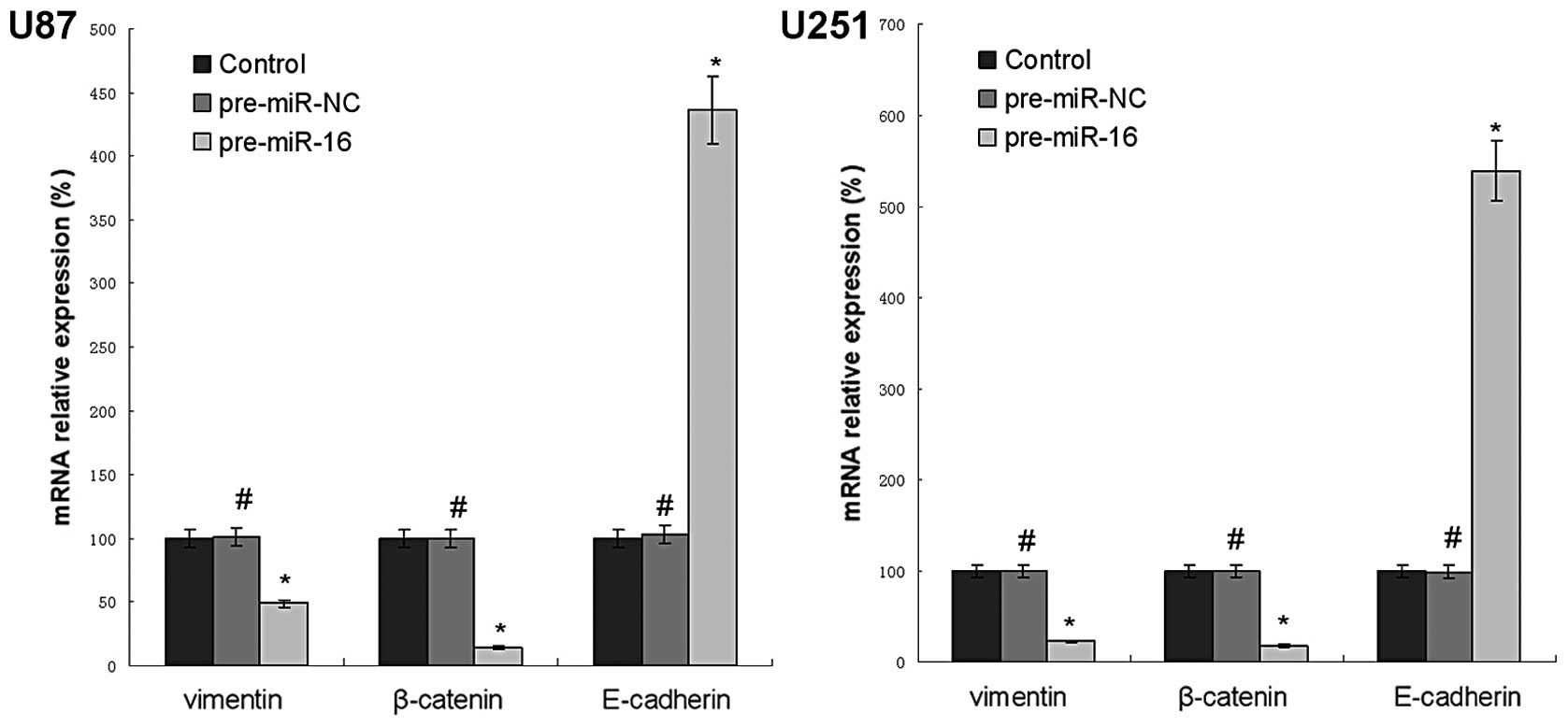
miR-16 regulates EMT-related gene
expression of human glioma cell lines in vitro
The EMT-related gene expression of human glioma cell
lines in vitro was measured by western blotting and qPCR.
The results revealed that the U87 and U251 miR-16 overexpressing
groups had a significantly lower protein expression levels of
vimentin, β-catenin, p-Akt and p-FAK, compared with control groups.
By contrast, the protein expression levels of E-cadherin in the U87
and U251 miR-16 overexpressing groups were higher than the control
groups (Fig. 4). In addition, the
mRNA expression levels of vimentin, β-catenin and E-cadherin
reflected the same trend in the U87 and U251 cell lines (Fig. 5).
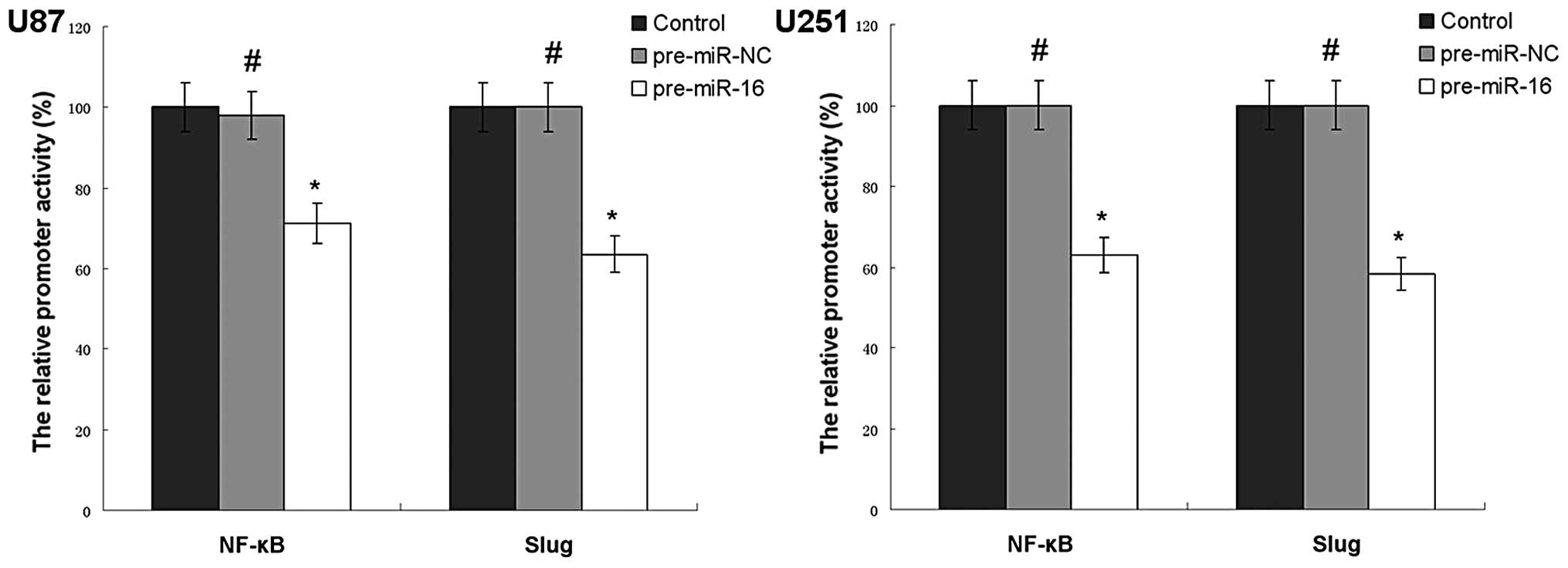
miR-16 regulates NF-κB transcriptional
activity of human glioma cell lines in vitro
As demonstrated in Fig.
6, miR-16 overexpression reduces the promoter activity of NF-κB
and Slug in U87 and U251 cells. The promoter activity of NF-κB in
the U87 pre-miR-16 and U251 pre-miR-16 groups were 71.3 and 62.9%
compared with the control group, respectively. The promoter
activity of Slug in the U87 pre-miR-16 and U251 pre-miR-16 groups
was 63.5 and 58.2% compared with the control group,
respectively.
Discussion
Previously, the correlation between EMT, and tumor
invasion and metastasis has attracted notable attention. EMT refers
to the process whereby epithelial cells interact with surrounding
mesenchymal cells, decreasing the expression of cellular adhesion
molecules, including E-cadherin, and leading to cell dispersion.
Subsequently, the main component of the cytoskeleton gradually
changes from keratin to vimentin resulting in its rearrangement, a
decrease in cellular adhesion and the enhancement of cell mobility.
Therefore, EMT enables non-invasive tumor cells to obtain invasion
ability, which then accelerates the formation of local tumor
invasion and distant metastasis (3).
E-cadherin and vimentin are important markers of EMT
and are key for maintaining the epithelial phenotype and
stabilizing inter-cellular contact (4–5). The
present study has demonstrated that subsequent to the
overexpression of miR-16, the mRNA and protein expression of
E-cadherin in tumor cells was significantly increased, while the
mRNA and the protein expression of vimentin was reduced in U87 and
U251 cells. These data indicate that the inhibitory effect of
miR-16 on the EMT process may be associated with the regulation of
the expression of E-cadherin and vimentin in tumor cells.
β-catenin, as a type of cellular adhesion and signal transduction
molecule, has an important role in the maintenance of cellular
structure and function. It may participate in cell adhesion as a
cytoskeletal component and participate in gene transcription inside
the nucleus. As an important member of the classic
E-cadherin-catenin complex, β-catenin bonds with E-cadherin to
participate in cell adhesion and connection.
Akt, FAK, NF-κB, Slug, IL-6, IL-8 and TGF-β also
have a key role in the signal transduction pathway and upon
activation may regulate downstream gene expression, which is
associated with EMT (6–9). The effect of EMT on p-Akt and p-FAK
protein expression, NF-κB and Slug transcriptional activity, and
IL-6, IL-8 and TGF-β production in tumor cells was investigated and
it was identified that miR-16 affected the EMT process by
preventing the excessive phosphorylation of FAK and Akt proteins,
the transcriptional activity of NF-κB and Slug, and IL-6, IL-8 and
TGF-β production in tumor cells in the EMT process.
In conclusion, miR-16 is closely associated with
EMT-related gene expression in human glioma and the present study
may have established a theoretical basis for a future clinical
treatment for human glioma.
References
|
1
|
Guo L, Qiu Y, Ge J and Zhou D:
Glioblastoma multiforme with subcutaneous metastases, case report
and literature review. J Korean Neurosurg Soc. 52:484–487. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Velpula KK, Dasari VR, Tsung AJ, Dinh DH
and Rao JS: Cord blood stem cells revert glioma stem cell EMT by
down regulating transcriptional activation of Sox2 and Twist1.
Oncotarget. 2:1028–1042. 2011.PubMed/NCBI
|
|
3
|
Techasen A, Loilome W, Namwat N, Dokduang
H, Jongthawin J and Yongvanit P: Cytokines released from activated
human macrophages induce epithelial mesenchymal transition markers
of cholangiocarcinoma cells. Asian Pac J Cancer Prev. 13(Suppl):
115–118. 2012.
|
|
4
|
Gheldof A and Berx G: Cadherins and
epithelial-to-mesenchymal transition. Prog Mol Biol Transl Sci.
116:317–336. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Liu T, Zhang X, Shang M, Zhang Y, Xia B,
Niu M, Liu Y and Pang D: Dysregulated expression of Slug, vimentin,
and E-cadherin correlates with poor clinical outcome in patients
with basal-like breast cancer. J Surg Oncol. 107:188–194. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Chen CH, Chuang SM, Yang MF, Liao JW, Yu
SL and Chen JJ: A novel function of YWHAZ/β-catenin axis in
promoting epithelial-mesenchymal transition and lung cancer
metastasis. Mol Cancer Res. 10:1319–1331. 2012.
|
|
7
|
Ho JN, Jun W, Choue R and Lee J: I3C and
ICZ inhibit migration by suppressing the EMT process and FAK
expression in breast cancer cells. Mol Med Rep. 7:384–388.
2013.PubMed/NCBI
|
|
8
|
Zhong D, Ran JH, Tang WY, Zhang XD, Tan Y,
Chen GJ, Li XS and Yan Y: Mda-9/syntenin promotes human brain
glioma migration through focal adhesion kinase (FAK)-JNK and
FAK-AKT signaling. Asian Pac J Cancer Prev. 13:2897–2901. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Takenouchi T, Iwamaru Y, Imamura M,
Fukuhara S, Sugama S, Sato M, Mochizuki N, Hashimoto M, Yokoyama T,
Mohri S and Kitani H: Cytochalasin D enhances the accumulation of a
protease-resistant form of prion protein in ScN2a cells:
involvement of PI3 kinase/Akt signalling pathway. Cell Biol Int.
36:1223–1231. 2012. View Article : Google Scholar : PubMed/NCBI
|