Introduction
Ovarian cancer is one of the most common three
malignant tumors of the female reproductive system. Ovarian cancer
has been recently reported as the leading cause of mortality in
gynecological cancers in the United States (1) and other countries around the world.
There have been pooled studies investigating the pathogenesis of
ovarian cancer in order to acquire new approaches for alternative
treatment. However, there have been minor changes in the survival
of patients with ovarian cancer since platinum-based anticancer
drugs were introduced more than three decades ago (2). Therefore, novel therapeutic agents
are required in order to improve the prognosis of ovarian
cancer.
Ceramide, a derivative of sphingolipid breakdown
products, was initially identified as a regulator of apoptosis
(3) and cellular senescence
(4) in the 1990s and has since
become a well-established mediator of cell death. Dysregulated
ceramide has been documented in cancer development and prevention,
since ceramide acts as a tumor-suppressor lipid in numerous tumor
cells. However, a recent study, which investigated the effects of
ceramide on the biomechanical properties of murine ovarian cancer
cells, has not indicated that suppression results from exogenous
ceramide administration (5).
Paradoxically, paclitaxel and ceramide co-administration
effectively inhibited the growth of multidrug resistant ovarian
cancer cell xenografts (6).
Exogenous ceramide administration has been shown to successfully
suppress tumor growth in both sensitive and resistant ovarian
cancer xenograft models. An increase in apoptosis was observed in
both the ceramide-only and combined treatment groups, not only in
sensitive tumors but also in multidrug resistant tumors. These data
have raised the question as to whether ceramide induces ovarian
cancer cell death. In the present study, exogenous C2-ceramide was
used to explore the possible response and related mechanism in
A2780 ovarian cancer cells.
The purpose of the present study was to understand
the effects of exogenous C2-ceramide on ovarian cancer cells and to
establish the possible therapeutic value of using C2-ceramide in
treating this disease. Experiments were designed to reveal whether
C2-ceramide induced programmed cell death in ovarian cancer cells.
Furthermore, the death mechanism was investigated, to establish
whether C2-ceramide could initiate apoptosis as well as cause
autophagy.
Materials and methods
Reagents and antibodies
C2-ceramide and 3-methyladenine (3-MA) were
purchased from Sigma-Aldrich (St. Louis, MO, USA) and the former
was prepared as a 50 mm stock solution in dimethyl sulfoxide (DMSO;
Sigma-Aldrich). The final concentration of DMSO in the culture
medium was <0.2% volume. Forkhead box O3 (FOXO3), phosphorylated
(P)-FOXO3, adenosine monophosphate-activated protein kinase (AMPK)
and P-AMPK (Thr172) antibodies were purchased from Cell Signaling
Technology, Inc. (Danvers, MA, USA). Akt and P-Akt (Ser473)
antibodies were purchased from Abcam (Cambridge, MA, USA). Beclin 1
and microtubule-associated protein 1 light chain 3 (LC3) antibodies
were both purchased from Novus Biologicals LLC (Littleton, CO,
USA). GAPDH antibody and horseradish peroxidase-secondary
antibodies were obtained from Santa Cruz Biotechnology, Inc. (Santa
Cruz, CA, USA) and Crystal Biotech Company, (Northborough, MA,
USA), respectively.
Cell culture and culture conditions
The A2780 human ovarian cancer cell line was
purchased from Wuhan University (Wuhan, China). Cells were
maintained in RPMI-1640 (Gibco-BRL, Carlsbad, CA, USA) supplemented
with 10% fetal bovine serum (Gibco-BRL), and incubated at 37°C with
5% CO2.
MTT assay and trypan blue exclusion
test
For the MTT assay, A2780 cells were plated
(1×104 cells/well) in 96-wells. Following 24 h of
incubation, the cells were treated with C2-ceramide at
concentrations ranging from 0 to 100 μmol/l and grown over a 24-h
period. DMSO was used as a control. Cell viability was measured
using an MTT assay according to the manufacturer’s instructions.
For the trypan blue exclusion assay, A2780 cells were plated onto
6-well plates (1×105 cells/well) and then treated with
25 μmol/l C2-ceramide at different time points. 3-MA, at
concentration of 2 mmol/l, was added 1 h prior to C2-ceramide
treatment. Cells were then collected and mixed 1:1 (w/v) with
trypan blue dye. Cells that excluded the dye were counted. Data are
presented as the means ± standard error of the mean, derived from
triplicate samples of three independent experiments.
Analysis of apoptosis
A2780 cells were plated, and after 12 h were treated
with vehicle control (DMSO) or C2-ceramide at the indicated
concentrations. Following 24 h incubation, the cells were washed
twice in phosphate-buffered saline (PBS) and then resuspended in
500 μl binding buffer and stained with 5 μl Annexin V-fluorescein
isothiocyanate (FITC) and 5 μl propidium iodide for 15 min at room
temperature in the dark. Samples were measured using a FACScan flow
cytometer within 60 min following staining. The rate of apoptosis
was analyzed using CellQuest software (BD Biosciences, Franklin
Lakes, NJ, USA).
In addition to Annexin V-FITC staining, an in
situ Cell Death Detection kit (Roche Diagnostics, Mannheim,
Germany) was used to confirm the induction of apoptosis according
to the manufacturer’s instructions. A2780 cells were plated onto 60
mm tissue culture plates (4×105 cells/plate) and treated
with a vehicle control (DMSO) or C2-ceramide at the indicated
concentrations for 24 h. Adherent cells were harvested by trypsin
treatment and were washed with PBS once, added to polylysine
treated glass slides, and then fixed with 4% paraformaldehyde
before permeabilization with 0.2% Triton X-100. The cells were then
treated with terminal deoxynucleotidyl transferase (TUNEL) in the
presence of fluorescein-labeled nucleotide polymers. TUNEL-positive
cells were analyzed by fluorescence microscopy to quantify the
number of apoptotic cells.
Transmission electron microscopy
A2780 cells were plated in 6-well plates and
incubated overnight. The cells were then treated with C2-ceramide
for 24 h. DMSO was used as a control. Samples were fixed in 4%
glutaraldehyde for 1 h at room temperature and then treated with 1%
osmium tetroxide (OsO4) for 1 h. The samples were then
dehydrated with increasing concentrations of ethanol, and gradually
infiltrated with araldite resin. Ultrathin sections were obtained
using an ultramicrotome (Leica, Mannheim, Germany). Sections were
stained with uranyl acetate and lead citrate, and examined using a
Tecnai™ G2 20 transmission electron microscope (FEI, Hillsboro, OR,
USA).
Quantitative polymerase chain reaction
(qPCR)
Total RNA was extracted from cells by using
TRIzol® (Invitrogen Life Technologies, Carlsbad, CA,
USA) reagent, according to the manufacturer’s instructions. The
reverse transcription reaction was performed with ReverTra Ace
(Toyobo, Osaka, Japan), Oligo(dT)20, RNase inhibitor, 5X RT buffer
and dNTP mixture (Invitrogen Life Technologies). The PCR primers
were designed using Premier Primer 5.0 software (Premier Biosoft
International, Palo Alton, CA, USA). Primer sequences are shown in
Table I. PCR amplification of cDNA
was performed by using SYBR green I (Biotium, Hayward, CA, USA) and
ABI Prism 7700 Sequence Detector (Applied Biosystems, Foster City,
CA, USA). Relative quantification of the targets was normalized
with an endogenous housekeeping gene (GAPDH) and data analysis was
performed by using the comparative (ΔΔCt) method.
 | Table IPrimer sequences for quantitative
polymerase chain reaction. |
Table I
Primer sequences for quantitative
polymerase chain reaction.
| Human gene | Forward primer | Reverse primer | Product length
(bp) |
|---|
| BECN1 |
CTCCCGAGGTGAAGAGCATC |
AATGGAGCTGTGAGTTCCTGG | 169 |
| MAP1LC3A |
CTCAGACCGGCCTTTCAAGC |
CGATGATCACCGGGATTTTGC | 101 |
| GABARAPL1 |
GGGCCAACTGTATGAGGACAA |
CAAGTCCAGGTGCTCCCATC | 120 |
| GABARAP |
TGCCTTCTGATCTCACAGTTGG |
CACTGGTGGGTGGAATGACA | 114 |
| BNIP3 |
GCCATCGGATTGGGGATCTA |
CCACCCCAGGATCTAACAGC | 149 |
| BNIP3L |
AATGTCGTCCCACCTAGTCG |
TCCACCCAGGAACTGTTGAG | 114 |
| BIM |
ATCCTCCCTGCTGTCTCGAT |
ATTTCTCTAACCATTGCACTGAGA | 150 |
| PUMA |
GAAACTGAAAAAGAAACGGAATGGA |
CTCCCTGGGGCCACAAATC | 139 |
| GAPDH |
CTATAAATTGAGCCCGCAGCC |
ACCAAATCCGTTGACTCCGA | 142 |
Western blot analysis
A2780 cells were seeded in 6 well plates and
incubated overnight, followed by the addition of 25 μmol/l ceramide
and incubation for the indicated time points. The cells were then
lysed in lysis buffer (50 mm Tris, 150 mm NaCl, 1% NP-40, EDTA,
β-glycerophosphate and protease inhibitor cocktail). The protein
extracts were quantified using a bovine serum albumin protein assay
kit. Equal amounts of protein (50 μg) were separated by 10%
SDS-PAGE (beclin 1, Akt, P-Akt, FOXO3, P-FOXO3, AMPK, and P-AMPK)
and 15% SDS-PAGE (LC3), respectively.
Statistical analysis
The statistical significance of the differences were
analyzed by t test in SPSS 13.0 (SPSS, Inc., Chicago, IL, USA).
P<0.05 was considered to indicate a statistically significant
difference. All experiments were repeated three times.
Results
C2-ceramide inhibits ovarian cancer cell
growth
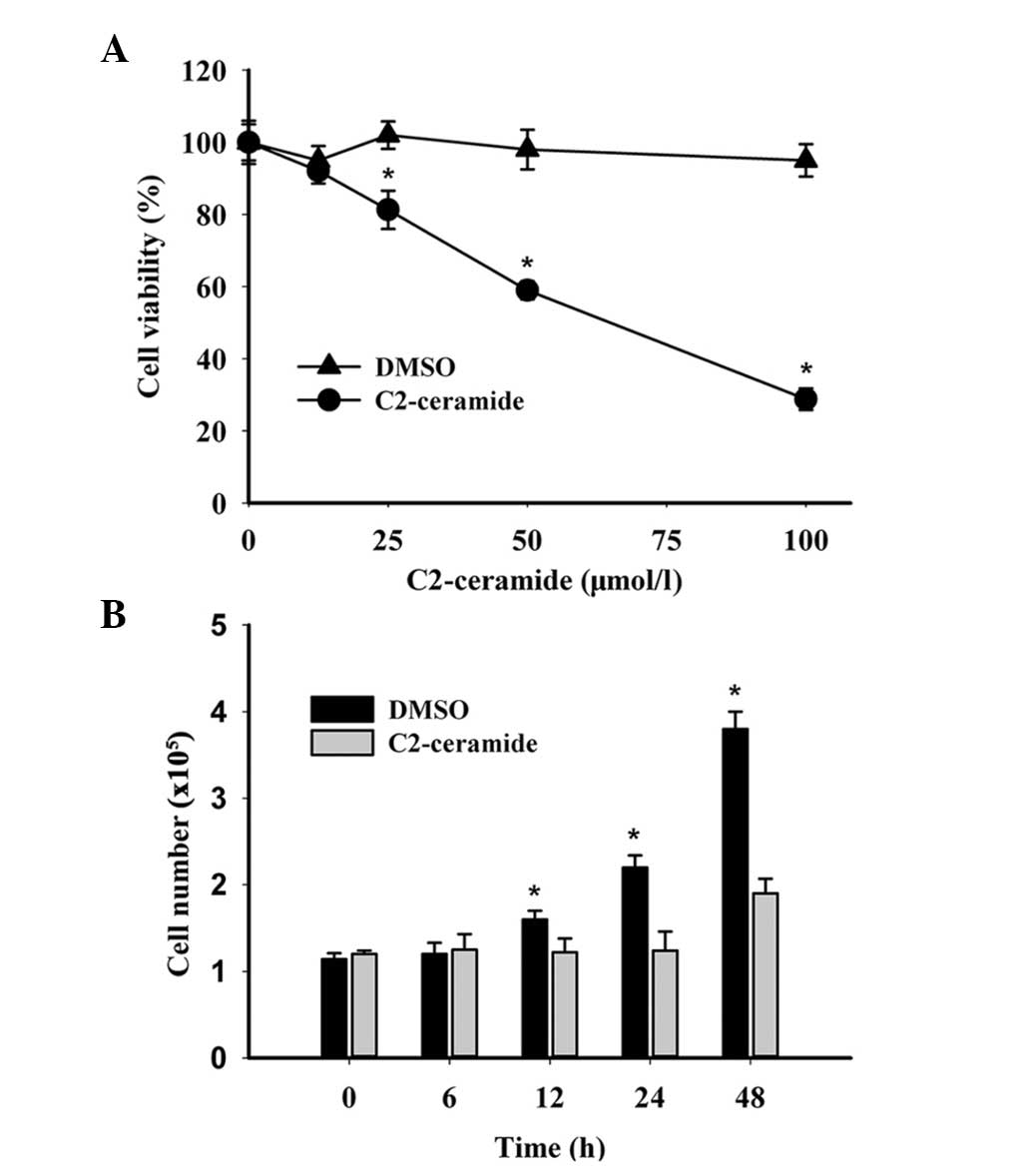
To assess the effects of C2-ceramide on ovarian
cancer cell growth, A2780 cells were treated with different
concentrations (0, 12.5, 25, 50, 100 μmol/l) of C2-ceramide.
Following 24 h treatment, the cells were analyzed by MTT assay. As
shown in Fig. 1A, increasing
concentrations of C2-ceramide significantly enhanced the inhibition
of cell growth. The half maximal inhibitory concentration
(IC50) of C2-ceramide was found to be 55.719 μmol/l. The
live cell numbers were analyzed by trypan blue test in the presence
of 25 μmol/l C2-ceramide, over a period of time. As shown in
Fig. 1B, the number of live cells
decreased over time following C2-caremide treatment, whereas the
number of live cells treated with DMSO remained relatively constant
over time. The results suggested that C2-ceramide could inhibit
ovarian cancer cell growth in a dose- and time-dependent
manner.
C2-ceramide induces apoptosis in ovarian
cancer cells
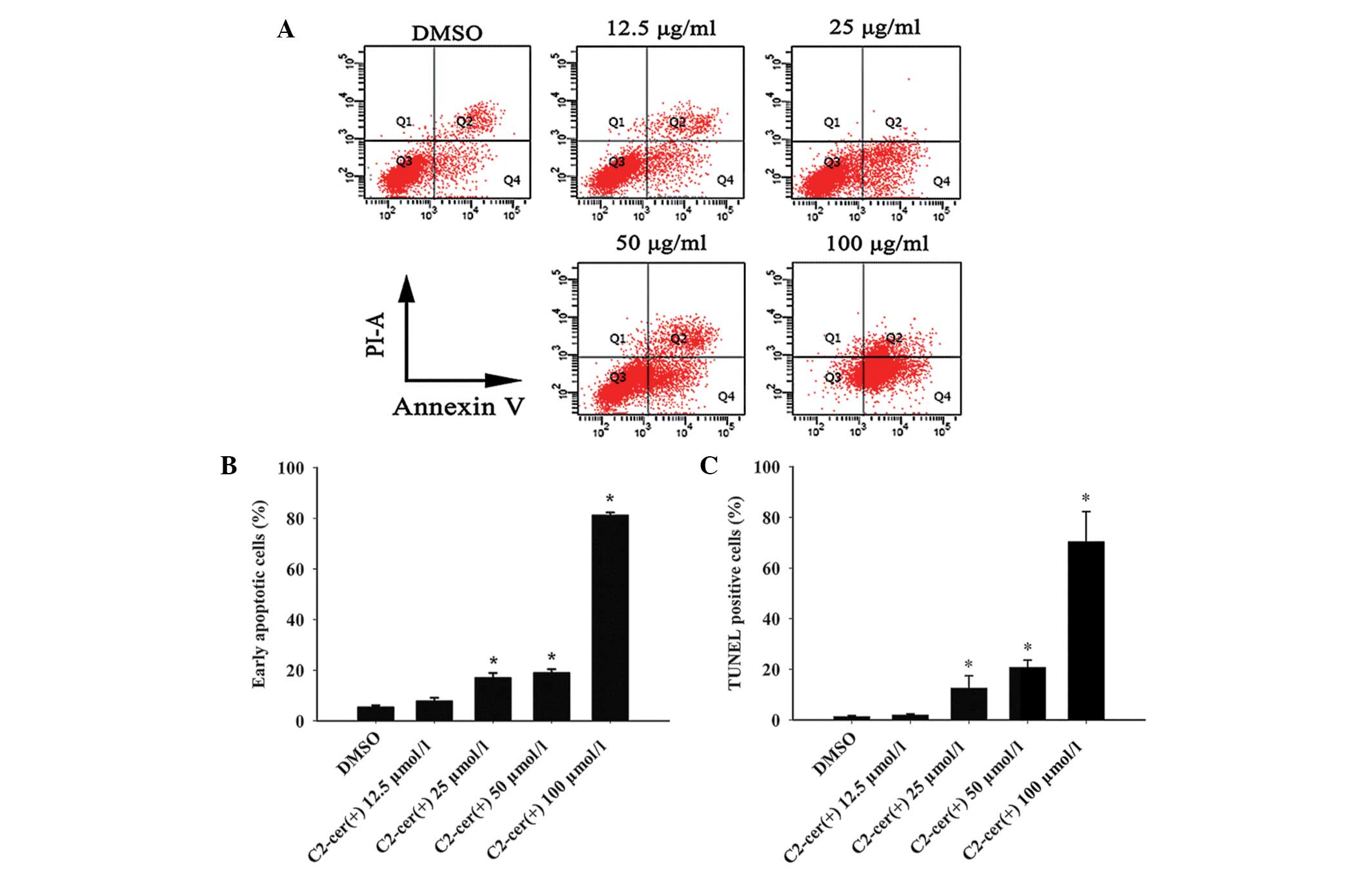
To understand the underlying mechanism of
C2-ceramide cytotoxicity in A2780 cells, flow cytometric analysis
and TUNEL assay were used to detect apoptosis. For the flow
cytometric analysis, cells were cultured under standard conditions
and treated with either DMSO as a control or the designated
concentration of C2-ceramide, for 24 h. The basal early apoptotic
rate in the DMSO group was 5.3%. The apoptotic rate was
dose-dependent when the concentration of C2-ceramide was >25
μmol/l (Fig. 2A and B). In
addition, the TUNEL assay confirmed the induction of apoptosis by
C2-ceramide (Fig. 2C). These
results indicated that C2-ceramide induced apoptosis in A2780
cells. On the basis of these results, 25 μmol/l C2-ceramide was
used for the subsequent studies.
C2-ceramide induced autophagy in ovarian
cancer cells
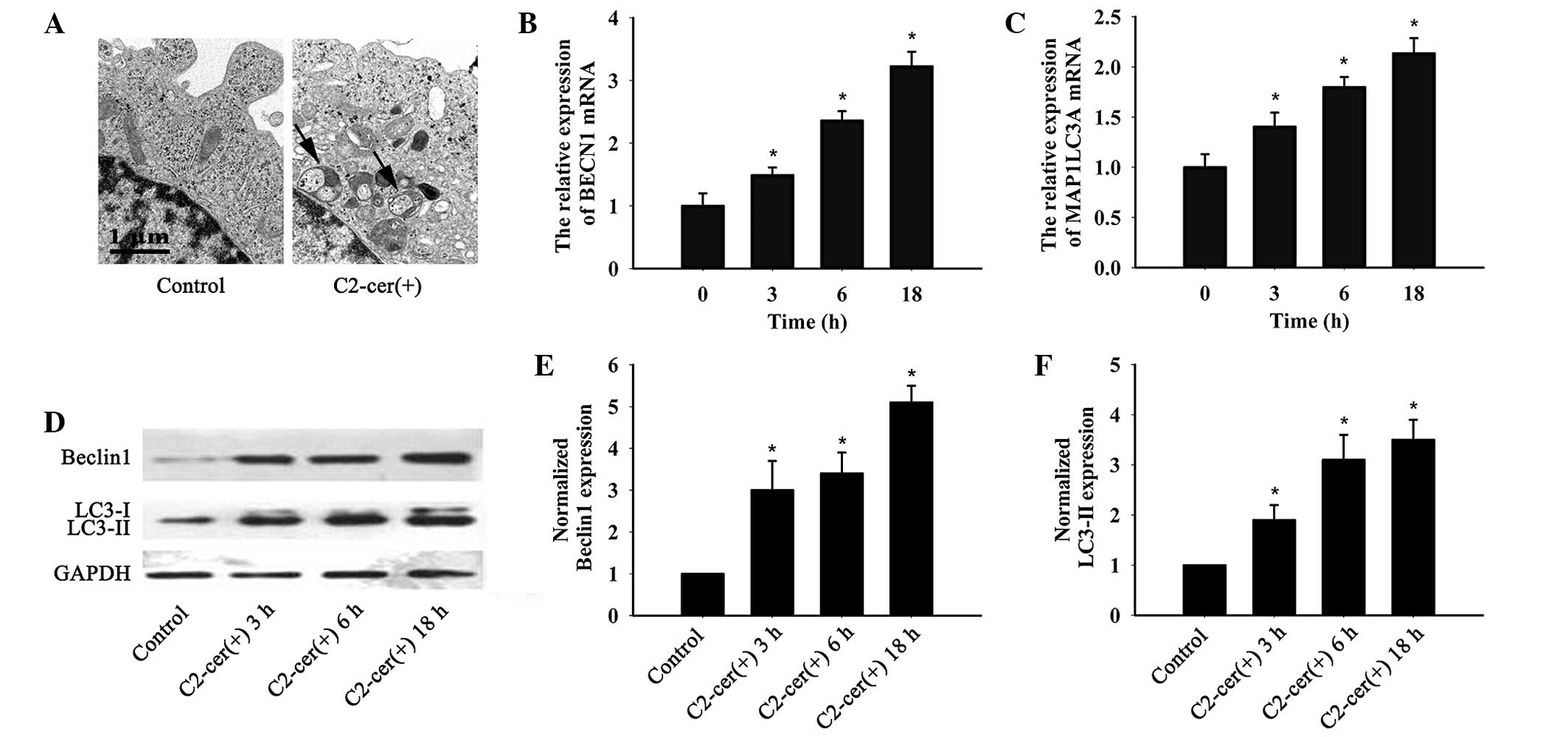
To determine whether C2-ceramide induced autophagy
in ovarian cancer cells, transmission electron microscopy was used,
which successfully confirmed that C2-ceramide induced the
accumulation of autolysosome structures (Fig. 3A). The effects of C2-ceramide on
beclin 1 and LC3 messenger (m)RNA and protein were analyzed by
quantitative polymerase chain reaction and western blotting,
respectively. As shown in Fig.
3B–F, C2-ceramide treatment increased beclin 1 and LC3 mRNA
level in a time dependent manner (Fig.
3B and C). The same results were confirmed by western blotting
(Fig. 3D–F). C2-ceramide increased
both LC3-I and LC3-II protein in A2780 cells, while the LC3-II
protein level was much higher than the LC3-I level. Thus,
C2-ceramide induced typical autophagy in A2780 ovarian cancer
cells.
C2-ceramide does not induce
autophagy-related cell death
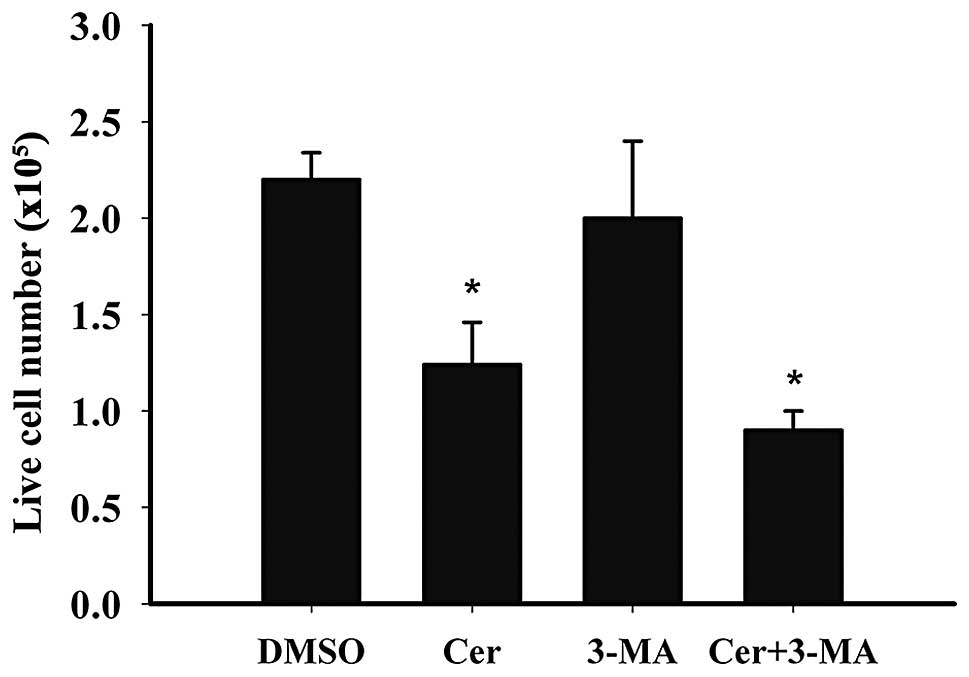
Ceramide has been previously shown to induce
autophagic cell death in malignant glioma cells (7). Since C2-ceramide treatment induces
apoptosis and autophagy, it was examined whether
C2-ceramide-dependent autophagy induced cell death in ovarian
cancer cells. A2780 cells were treated with autophagy inhibitor
3-MA with or without C2-ceramide for 24 h. Live cells were then
monitored by trypan blue test. As shown in Fig. 4, the presence of 3-MA reduced the
number of live cells following C2-ceramide treatment. This
suggested that in A2780 cells, autophagy may serve as a protective
mechanism against C2-ceramide-induced apoptosis. A similar
mechanism has been previously shown in MCF-7 cells treated with
C6-ceramide (8).
FOXO3-dependent transcription is
activated by C2-ceramide treatment
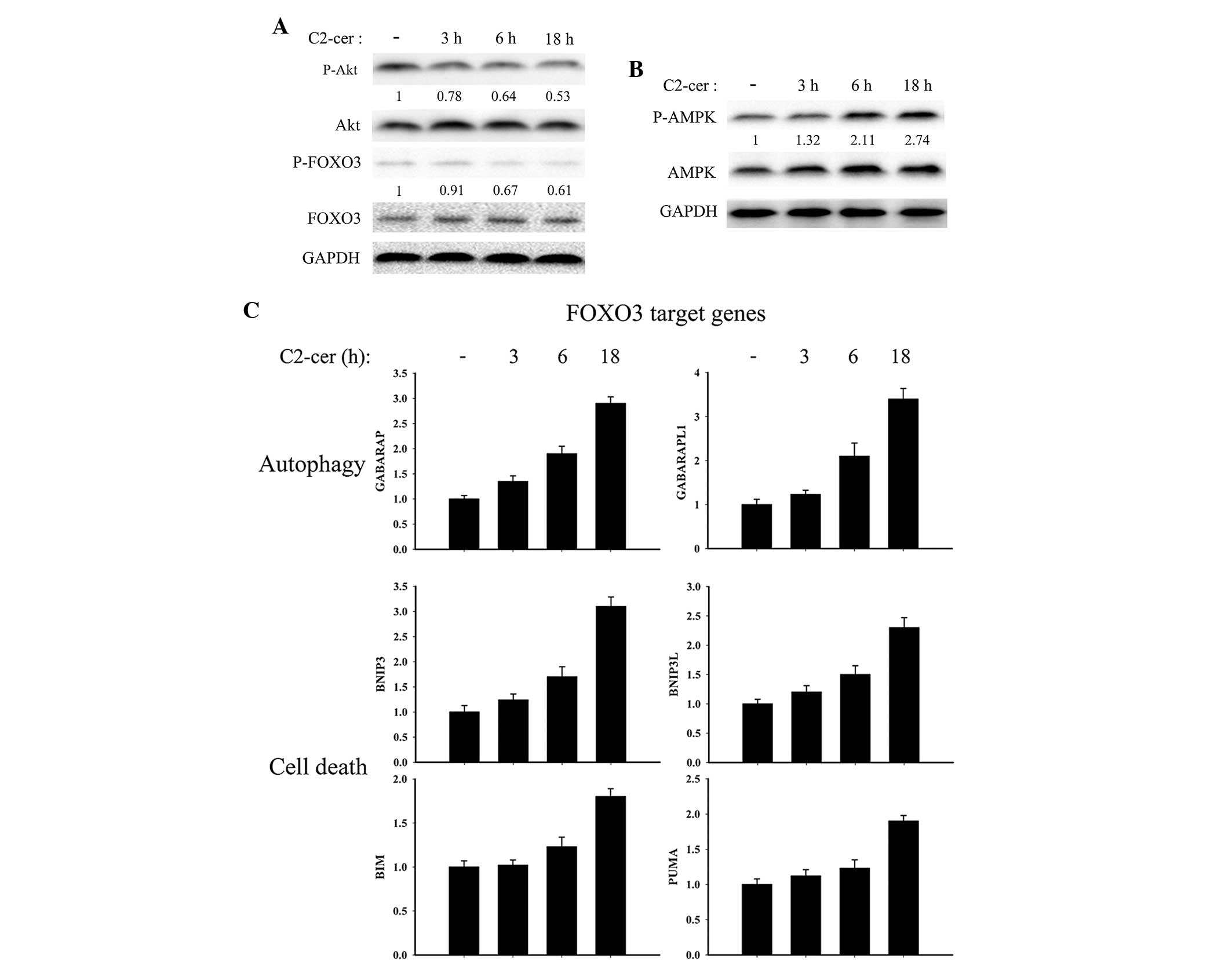
Ceramide has been show to inhibit Akt
phosphorylation (9). FOXO3 is
phosphorylated and inactivated by activated Akt, and translocates
from the nucleus thus decreasing the transcription of the target
genes (10). In the present study,
the Akt, P-Akt (Ser473), FOXO3, P-FOXO3 (Thr32) status was
investigated following C2-ceramide treatment in A2780 cells.
C2-ceramide treatment significantly inhibited Akt phosphorylation
in a time dependent manner and FOXO3 phosphorylation was reduced,
thus causing an increase in the unphosphorylated active pool of
FOXO3 (Fig. 5A). In addition, a
previous study has reported that AMPK was associated with FOXO3
phosphorylation (10). Given this
data, the AMPK phosphorylation status was monitored. It was
identified that AMPK was significantly phosphorylated in A2780
cells following C2-ceramide treatment. Given this, it was
investigated whether FOXO3 target genes were modulated during
C2-ceramide treatment. Consistently, the FOXO3 target genes, which
were associated with autophagy and cell death, were significantly
upregulated by C2-ceramide (Fig.
5C). GABARAP, GABARAPL1 and MAP1LC3 (Fig. 3C) belong to the ATG8 gene family
and code for proteins involved in autophagosome biogenesis
(11). Their expression was
consistent with the formation of autophagic vesicles in
C2-ceramide-treated A2780 cells (Fig.
3A). BNIP3, BNIP3L, BIM and PUMA are proapoptotic genes.
Altogether, these data indicate that C2-ceramide promotes FOXO3
transcription in A2780 ovarian cancer cells.
Discussion
In the present study, it was identified that
C2-ceramide inhibited the proliferation of A2780 ovarian cancer
cells in vitro. C2-ceramide not only induced apoptosis in
A2780 cells apoptosis, but also autophagy. However,
C2-ceramide-associated autophagy did not induce cell death but
protected cells from apoptosis. In addition, FOXO3 transcription
was activated by C2-ceramide in A2780 ovarian cancer cells and its
target genes were associated with apoptosis and autophagy.
Ovarian cancer is a particularly challenging disease
attributed to late stage diagnosis and development of resistance to
chemotherapy. Although the majority of the patients respond to the
first-line chemotherapy, they will relapse and eventually succumb
to the disease. The relapse is predominantly due to
chemotherapeutic resistance (2).
Therefore, identification of novel compounds that circumvent this
resistance mechanism is required to improve the management of
ovarian cancer.
The mechanism of ceramide-induced cell death through
apoptosis is well established (12), inducing apoptosis through extrinsic
and intrinsic pathways (13). The
extrinsic apoptosis pathway is often triggered by the activation of
tumor necrosis factor receptors, whereas the intrinsic pathway is
initiated by mitochondrial dysfunction. Furthermore, ceramide has
also been found to cause autophagy which results in either cell
survival or death (14). Previous
research has reported that ceramide caused COX-2-dependent
apoptosis in ovarian cancer OVCAR-3 cells (15). In the present study, C2-ceramide
did not only induce cell apoptosis, but also autophagy. C2-ceramide
treatment of A2780 cells significantly enhanced the formation of
autophagosomes and elevated the expression of LC3 and beclin 1 at
both the mRNA and protein level.
Autophagy is a homeostatic and evolutionarily
conserved process that regulates the cellular levels of long-lived
proteins and organelles. Under several conditions, autophagy
constitutes a stress adaptation that avoids cell death, whereas in
other circumstances, autophagy constitutes an alternative pathway
to cell death (16). LC3 (17) and mammalian homologue of yeast
autophagy-related (ATG) gene 8 (ATG8), are associated with the
autophagosome membranes. Beclin 1 (18), a mammalian homolog of the yeast
ATG6, functions as a scaffold for the formation of the
phosphoinositide 3-kinase (PI3K) complex, which is essential for
the recruitment of other Atg proteins during the development of
autophagosomes (19). Therefore,
LC3-II together with beclin 1 was monitored in the present study as
markers for autophagosomes. The expression of LC3 and beclin 1 has
been previously shown to be decreased in malignant epithelial
ovarian cancers (20).
Overexpression of beclin 1 in SKOV3 ovarian cancer cells has
additionally been shown to inhibit proliferation and induce
apoptosis (21). The decrease of
autophagic capacity may be related to tumorigenesis and the
development of epithelial ovarian cancer. Therefore, inducers of
autophagy could be applied for the treatment of epithelial ovarian
cancer (22). C2-ceramide-induced
autophagy partially attenuated C2-ceramide-induced A2780 cell
apoptosis in the present study. Mathew et al (23) identified that reduced autophagy can
promote chromosome instability, which associated with tumor
progression and poor prognosis. It is possible that paclitaxel and
ceramide co-administration could promote autophagy, to limit genome
damage thus resulting in inhibition of the growth of multidrug
resistant ovarian cancer cells (6). Further studies are needed to confirm
whether C2-ceramide increases the chemosensitivity of ovarian
cancers.
Interestingly, apoptosis and autophagy are two major
opposing pathways that regulate cellular outcomes (24). They are often co-regulated by
common upstream signaling components (16). FOXO3, one of the FOXO transcription
factors, is the downstream target of the PI3K-Akt pathway, which
functions in tumorigenesis and cancer progression (10). Inhibition of Akt causes
translocation of FOXO3 to the nucleus, while Akt overexpression
causes FOXO3 protein reduction in prostate cancer cells (25). FOXO3 acts as a suppressor of
follicular activation at the earliest stages of follicular growth
in mice (26). Low expression of
FOXO3 has been significantly associated with poor prognosis in
ovarian cancer patients (27).
Detainment of FOXO3 could therefore be one option to treat ovarian
cancer. In the present study, it was found that Akt activity was
inhibited by C2-ceramide, accompanied by a decrease in FOXO3
phosphorylation and non-phosphorylated-FOXO3 accumulation. It has
been recently shown that AMPK phosphorylation is associated with
C6-ceramide-induced autophagy cell death in colorectal cancer cells
(28), consistent with these
results that AMPK was activated by C2-ceramide. It was shown that
FOXO3 targeted genes, which were related to apoptosis (BNIP3,
BNIP3L, BIM and PUMA) and autophagy (MAP1LC3, GABARAP and
GABARAPL1) were upregulated by C2-ceramide in A2780 cells.
BNIP3, BNIP3L, BIM and PUMA are all members of the
BH3-only subfamily which belongs to the Bcl-2 gene family (29). These proteins are well
characterized pro-apoptotic proteins, that also regulate autophagy.
BINP3 is activated by C2-ceramide in malignant gliomas and
functions in C2-ceramide-induced autophagic cell death (7). Beclin 1, BINP3 and BINP3L
transcriptions were shown in the present study to be activated by
C2-ceramide. However, C2-ceramide did not cause A2780 autophagic
cell death. Interestingly, a previous study has shown that a
hypoxic microenvironment promoted BNIP3 and BNIP3L competing with
beclin-1-Bcl-2 and beclin 1-Bcl-XL complexes, releasing
beclin-1 from the complex and enhancing autophagy (30). There is currently little known
regarding the expression and activity of BIM and PUMA in ceramide
treated cells, however they are known to function in mitochondrial
apoptosis (31). It is therefore
hypothesized that C2-ceramide may regulate apoptosis and autophagy
through FOXO3 transcription in A2780 cells. The specific molecular
mechanism for C2-ceramide regulating A2780 cells death and survival
remains to be elucidated.
In summary, the present study has demonstrated that
C2-ceramide can induce cell apoptosis and autophagy in ovarian
cancer cells. FOXO3 target genes, which related to cell death and
autophagy, were upregulated by C2-ceramide. These findings provide
a novel concept to ceramide-induced cell death mechanism, as well
as a potential role of ceramide in anti-ovarian cancer therapy.
Acknowledgements
The authors would like to thank Professor Qinghua Hu
form Key Laboratory of Pulmonary Diseases of Ministry of Health of
China, Tongji Medical College, Huazhong Science and Technology
University for his valuable suggestions and lab assistance.
References
|
1
|
Siegel R, Naishadham D and Jemal A: Cancer
statistics, 2013. CA Cancer J Clin. 63:11–30. 2013. View Article : Google Scholar
|
|
2
|
Vaughan S, Coward JI, Bast RC Jr, et al:
Rethinking ovarian cancer: recommendations for improving outcomes.
Nat Rev Cancer. 11:719–725. 2011. View
Article : Google Scholar : PubMed/NCBI
|
|
3
|
Obeid LM, Linardic CM, Karolak LA and
Hannun YA: Programmed cell death induced by ceramide. Science.
259:1769–1771. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Venable ME, Lee JY, Smyth MJ, Bielawska A
and Obeid LM: Role of ceramide in cellular senescence. J Biol Chem.
270:30701–30708. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Babahosseini H, Roberts PC, Schmelz EM and
Agah M: Roles of bioactive sphingolipid metabolites in ovarian
cancer cell biomechanics. Conf Proc IEEE Eng Med Biol Soc.
2012:2436–2439. 2012.PubMed/NCBI
|
|
6
|
Devalapally H, Duan Z, Seiden MV and Amiji
MM: Paclitaxel and ceramide co-administration in biodegradable
polymeric nanoparticulate delivery system to overcome drug
resistance in ovarian cancer. Int J Cancer. 121:1830–1838. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Daido S, Kanzawa T, Yamamoto A, et al:
Pivotal role of the cell death factor BNIP3 in ceramide-induced
autophagic cell death in malignant glioma cells. Cancer Res.
64:4286–4293. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Hou Q, Jin J, Zhou H, et al:
Mitochondrially targeted ceramides preferentially promote
autophagy, retard cell growth, and induce apoptosis. J Lipid Res.
52:278–288. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Scarlatti F, Bauvy C, Ventruti A, et al:
Ceramide-mediated macroautophagy involves inhibition of protein
kinase B and up-regulation of beclin 1. J Biol Chem.
279:18384–18391. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Chiacchiera F and Simone C: The
AMPK-FOXO3A axis as a target for cancer treatment. Cell Cycle.
9:1091–1096. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Weidberg H, Shvets E, Shpilka T, et al:
LC3 and GATE-16/GABARAP subfamilies are both essential yet act
differently in autophagosome biogenesis. EMBO J. 29:1792–1802.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Young MM, Kester M and Wang HG:
Sphingolipids: regulators of crosstalk between apoptosis and
autophagy. J Lipid Res. 54:5–19. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Morad SA and Cabot MC:
Ceramide-orchestrated signalling in cancer cells. Nat Rev Cancer.
13:51–65. 2013. View
Article : Google Scholar : PubMed/NCBI
|
|
14
|
Pattingre S, Bauvy C, Levade T, Levine B
and Codogno P: Ceramide-induced autophagy: to junk or to protect
cells? Autophagy. 5:5582009. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Lin HX, Qiu HJ, Zeng F, et al: Decreased
expression of Beclin 1 correlates closely with Bcl-xL expression
and poor prognosis of ovarian carcinoma. PLoS One. 8:e605162013.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Maiuri MC, Zalckvar E, Kimchi A and
Kroemer G: Self-eating and self-killing: crosstalk between
autophagy and apoptosis. Nat Rev Mol Cell Biol. 8:741–752. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Kabeya Y, Mizushima N, Ueno T, et al: LC3,
a mammalian homologue of yeast Apg8p, is localized in autophagosome
membranes after processing. EMBO J. 19:5720–5728. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Aita VM, Liang XH, Murty VV, et al:
Cloning and genomic organization of beclin 1, a candidate tumor
suppressor gene on chromosome 17q21. Genomics. 59:59–65. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Cao Y and Klionsky DJ: Physiological
functions of Atg6/Beclin 1: a unique autophagy-related protein.
Cell Res. 17:839–849. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Shen Y, Li DD, Wang LL, Deng R and Zhu XF:
Decreased expression of autophagy-related proteins in malignant
epithelial ovarian cancer. Autophagy. 4:1067–1068. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Duan ZL, Peng ZL, Wang ZH and Yan NH:
Correlation of autophagy gene Beclin1 to tumorigenesis and
development of epithelial ovarian cancer. Ai Zheng. 26:258–263.
2007.(In Chinese).
|
|
22
|
Bast RC Jr, Hennessy B and Mills GB: The
biology of ovarian cancer: new opportunities for translation. Nat
Rev Cancer. 9:415–428. 2009. View
Article : Google Scholar : PubMed/NCBI
|
|
23
|
Mathew R, Kongara S, Beaudoin B, et al:
Autophagy suppresses tumor progression by limiting chromosomal
instability. Genes Dev. 21:1367–1381. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Eisenberg-Lerner A, Bialik S, Simon HU and
Kimchi A: Life and death partners: apoptosis, autophagy and the
cross-talk between them. Cell Death Differ. 16:966–975. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Shukla S, Shukla M, Maclennan GT, Fu P and
Gupta S: Deregulation of FOXO3A during prostate cancer progression.
Int J Oncol. 34:1613–1620. 2009.PubMed/NCBI
|
|
26
|
Castrillon DH, Miao L, Kollipara R, Horner
JW and DePinho RA: Suppression of ovarian follicle activation in
mice by the transcription factor FOXO3a. Science. 301:215–218.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Fei M, Zhao Y, Wang Y, et al: Low
expression of Foxo3a is associated with poor prognosis in ovarian
cancer patients. Cancer Invest. 27:52–59. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Huo HZ, Wang B, Qin J, et al:
AMP-activated protein kinase (AMPK)/Ulk1-dependent autophagic
pathway contributes to C6 ceramide-induced cytotoxic effects in
cultured colorectal cancer HT-29 cells. Mol Cell Biochem.
378:171–181. 2013. View Article : Google Scholar
|
|
29
|
Levine B, Sinha S and Kroemer G: Bcl-2
family members: dual regulators of apoptosis and autophagy.
Autophagy. 4:600–606. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Mazure NM and Pouysségur J: Atypical
BH3-domains of BNIP3 and BNIP3L lead to autophagy in hypoxia.
Autophagy. 5:868–869. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Kim H, Tu HC, Ren D, et al: Stepwise
activation of BAX and BAK by tBID, BIM, and PUMA initiates
mitochondrial apoptosis. Mol Cell. 36:487–499. 2009. View Article : Google Scholar : PubMed/NCBI
|