Introduction
In the past decade, an increasing number of studies
have demonstrated the involvement of various adipokines in the
process of liver cirrhosis, of which the most important are leptin
(1), adiponectin (2), tumor necrosis factor-α, resistin and
interleukin-6 (3,4). Visfatin, a novel adipokine which has
insulin-like effects, regulates inflammatory and immunomodulatory
processes (5,6) and catalyzes the rate-limiting step in
the production of nicotinamide adenine dinucleotide from
nicotinamide. Previous studies have identified the potential
effects of visfatin on the proliferation and collagen synthesis of
rat cardiac fibroblasts (7).
Visfatin is involved in perivascular adipose tissue-induced
vascular smooth muscle cell proliferation (8) and the increased synthesis of
profibrotic molecules, including transforming growth factor-1
(TGF-1) and plasminogen activator inhibitor-1 (9). Therefore, there is convincing
evidence to suggest that visfatin is involved in the initiation and
progression of fibrosis.
Hepatic stellate cells (HSC) are liver mesenchymal
cells located in the space of Disse, in close contact with
sinusoidal endothelial cells and hepatocytes (10). The process of quiescent HSC
activation into contractile myofibroblasts is an important step
during liver fibrogenesis (11).
Quiescent HSC can transdifferentiate into α-smooth muscle actin
(α-SMA)-positive activated myofibroblast-like cells, which produce
a wide variety of collagenous and noncollagenous extracellular
matrix (ECM) proteins. Increased collagen deposition leads to the
accumulation of ECM, which is the major cause of liver dysfunction
during hepatic fibrosis (12).
The association between visfatin and the activation
of HSC, the key event in fibrogenesis, remains to be elucidated. In
the present study, culture-activated HSC were treated with
visfatin. The mRNA and protein expression levels of α-SMA, collagen
types I and III and connective tissue growth factor (CTGF) were
then determined using reverse transcription quantitative polymerase
chain reaction (RT-qPCR) and western blot analyses.
Materials and methods
Cell culture
Primary cultures of HSC were isolated from the
livers of eight healthy male Sprague-Dawley rats (10–12 weeks old,
weighing 450–500 g), which were obtained from the Animal Center of
Sichuan University (Sichuan, China), by in situ pronase and
collagenase perfusion (13) and a
single step Nycodenz gradient according to previous studies by our
group (10,14). The HSC were maintained in
Dulbecco’s modified Eagle’s medium (DMEM; Gibco-BRL, Grand Island,
NY, USA) supplemented with 10% fetal bovine serum (FBS; HyClone,
Logan, UT, USA), 2 mmol/l L-glutamine (Gibco-BRL) and 1%
penicillin-streptomycin (KeLong Chemical Co., Ltd., Chengdu,
China). All cell cultures were maintained in a humidified 5%
CO2/95% air incubator at 37°C. The primary HSC were
identified by assessing the expression of desmin and their
activation was determined by the level of expression of the
specific marker, α-smooth muscle actin (α-SMA). The present study
was approved by the Medical Ethics Committee of the West China
Hospital, Sichuan University (Chengdu, China).
Based on previous studies (7,9,15,16),
experimental concentrations of 0, 25, 50, 100 and 200 ng/ml
visfatin were selected for the present study. Subsequently, HSC at
90% confluence in the culture dishes were serum-starved for 24 h
followed by treatment with different concentrations of visfatin
(25, 50, 100 or 200 ng/ml; Enzo Biochemical Co., Inc., Farmingdale,
NY, USA) for 24 h with the exception of an untreated group as the
control. All animal investigations were approved by the Ethics
Committee for Animal Care of Sichuan University (Chengdu,
China).
Immunocytochemistry
For immunocytochemical staining, the HSC were
cultured on coverslips in six-well plates (1×105
cells/well). The cells were fixed by incubation with 4%
paraformaldehyde in phosphate-buffered saline (PBS) for 20 min
stored at 4°C followed by washing in PBS three times. The cells
were then rinsed in 0.1% Triton X-100 (Kelong Chemical Co., Ltd.)
in PBS for 15 min at room temperature (RT), washed three times in
PBS and incubated with 10% goat serum (Gibco-BRL) for 1 h to block
non-specific binding sites. The coverslips were incubated with the
primary antibodies monoclonal rabbit anti-rat α-SMA (Abcam,
Cambridge, UK) or polyclonal rabbit anti-rat desmin (Santa Cruz
Biotechnology, Inc., Dallas, TX, USA) at working dilutions of 1:100
at 4°C overnight. Negative controls were included by incubation
with goat serum (Gibco-BRL) corresponding to the primary antibody.
Following rinsing three times with PBS, the secondary biotinylated
goat anti-rabbit secondary antibody (1:200; Zhonshan Jinquai
Biotechnology Co., Ltd., Beijing, China) was applied and incubated
at room temperature for 2 h. Color development was performed using
a DBA kit (Zhonshan Jinquai Biotechnology Co., Ltd.) as previously
described (17). A microscope
(TH4-200; Olympus, Tokyo, Japan) was then used for image
capture.
RT-qPCR
Total RNA from the HSC was isolated using TRIzol
reagent (Invitrogen Life Technologies, Carlsbad, CA, USA) according
to the manufacturer’s instructions and the purity and content of
total RNA were determined by spectrophotography (SmartSpec Plus;
Bio-Rad, Hercules, CA, USA). The A260/A280 ratio of total RNA was
between 1.8 and 2.0. For the first-strand cDNA synthesis, 1 μg
total RNA was used in each sample using an RT kit (ReverTra Ace;
Toyobo, Osaka, Japan) according to the manufacturer’s instructions.
qPCR was performed on a Bio-Rad real-time PCR system (CFX96;
Bio-Rad, Hercules, CA, USA) using the fluorescent dye SYBR-Green I
(SYBR-Green Real-time PCR Master mix; Toyobo). The sequences of the
primers used were as follows: α-SMA forward, 5′-CCG AGA TCT CAC CGA
CTA CC-3′ and reverse, 5′-TCC AGA GCG ACA TAG CAC AG-3′; collagen I
(α1) forward, 5′-ACG TCC TGG TGA AGT TGG TC-3′ and reverse, 5′-TCC
AGC AAT ACC CTG AGG TC-3′ (18);
collagen III (α1) forward, 5′-TCC TCT GTG ATG ACA TAA TGT GTG-3′
and reverse, 5′-GTA GAA GGC TGT GGA CAT ATT GC-3′ (19); CTGF forward, 5′-CAG GGA GTA AGG GAC
ACG A-3′ and reverse, 5′-ACA GCA GTT AGG AAC CCA GAT-3′ (20) and β-actin forward, 5′-ACT ATC GGC
AAT GAG CGG TTC-3′ and reverse, 5′-ATG CCA CAG GAT TCC ATA CCC-3′.
The primers was provided by Invitrogen Life Technologies. The
amplification conditions for each sample were as follows: 95°C for
1 min and 40 cycles of 95°C for 15 sec and 60°C for 40 sec. The
fluorescence resulting from the incorporation of SYBR-Green I into
the double-stranded DNA was detected at the end of the elongation
phase of each PCR cycle using an RT-qPCR detection system (CFX96;
Bio-Rad). Each gene was standardized against the housekeeping gene
β-actin. The mRNA levels were expressed as a ratio, using the
2−ΔΔCT method with Bio-Rad software (Quantity One 4.62;
Bio-Rad), for comparing the relative mRNA expression levels between
different groups in the qPCR.
Western blot analysis
The treated HSC were lysed with
radioimmunoprecipitation assay buffer containing 50 mM Tris-HCl (pH
7.4), 150 mM NaCl, 1% NP-40, 0.1% SDS and the protease inhibitor
cocktail cOmplete Mini (Roche Diagnostics GmbH, Mannheim, Germany).
The protein concentrations were determined using a bicinchoninic
acid protein assay kit (Pierce Biotechnology, Inc., Rockford, IL,
USA). The total proteins from the cell lysate (30 μg) were
separated by 10% SDS-PAGE by electrophoresis and then transferred
onto a 0.45-μm nitrocellulose membrane (Roche Diagnostics). The
membranes were inhibited by agitation in 5% fat-free milk in
tris-buffered saline-Tween 20 (TBS-T; KeLong Chemical Co., Ltd.)
buffer for 2 h at RT and subsequently incubated overnight at 4°C
with the following primary antibodies: Rabbit anti-rat α-SMA
polyclonal antibody (1:1,000; Abcam), mouse anti-rat collagen type
I monoclonal antibody (1:200), goat anti-rat collagen type III
polyclonal antibody (1:200), goat anti-rat CTGF polyclonal antibody
(1:200) and mouse anti-rat β-actin monoclonal antibody (1:2,000)
all from Santa Cruz Biotechnology, Inc. The membranes were then
washed with TBS-T buffer and incubated with horseradish
peroxidase-conjugated secondary antibody (1:3,000 in TBST with 5%
skim milk) at room temperature for 2 h (Zhonshan Jinquai
Biotechnology Co., Ltd.). The bands were detected using an enhanced
chemiluminescence detection kit (ECL kit; Pierce Biotechnology,
Inc.,). Optical densities of the bands were scanned and quantified
using the Gel Doc 2000 system (Bio-Rad). β-actin was used as a
loading control.
Statistical analysis
Data are expressed as the mean ± standard error of
the mean. Each experiment was repeated three times. Group means
were compared by one-way analysis of variance using the statistical
software program SPSS 18.0 for Windows (International Business
Machines, Armonk, NY, USA). P<0.05 was considered to indicate a
statistically significant difference.
Results
HSC culture activation and
identification
By culturing quiescent HSC on uncoated plastic
culture wells, spontaneous activation was induced, mimicking the
process observed in vivo, and providing a suitable model for
investigating the relative factors involved in HSC activation and
its underlying mechanisms (21). A
number of techniques can be used to assist in the identification of
HSC. The expression of desmin and α-SMA remain useful methods for
identifying primary and activated HSC in vitro (22). In the present study, the cells were
stained for desmin, which is a marker of HSC that is positive in
rodent HSC regardless of activation status (23). The immunocytochemical staining by
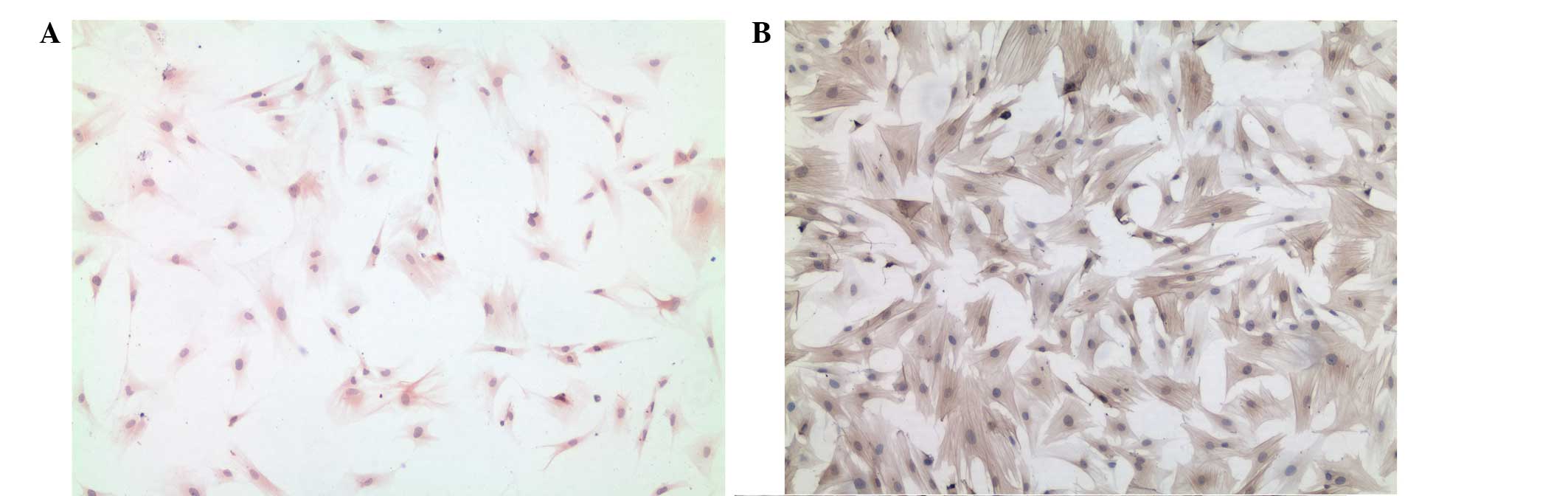
desmin suggested that, by day 7 of culturing, HSC accounted for
>90% of the cells in the cultures (Fig. 1A). The passaged cells were also
stained for α-SMA, a marker of activated stellate cell phenotypes.
The passaged cells were markedly positive for α-SMA (Fig. 1B).
Visfatin upregulates the mRNA and protein
expression of α-SMA in HSC
The expression of α-SMA is a reliable marker of HSC
activation. To determine whether visfatin had modulatory effects on
the expression of α-SMA in HSC, the rat HSC were treated with
different concentrations of visfatin for 24 h. The results
demonstrated that 100 and 200 ng/ml visfatin evoked significant
expression of α-SMA in the HSC compared with the control group.
Additionally, 200 ng/ml visfatin had a more marked effect on
inducing the gene and protein expression of α-SMA compared with 100
ng/ml visfatin (Fig. 2).
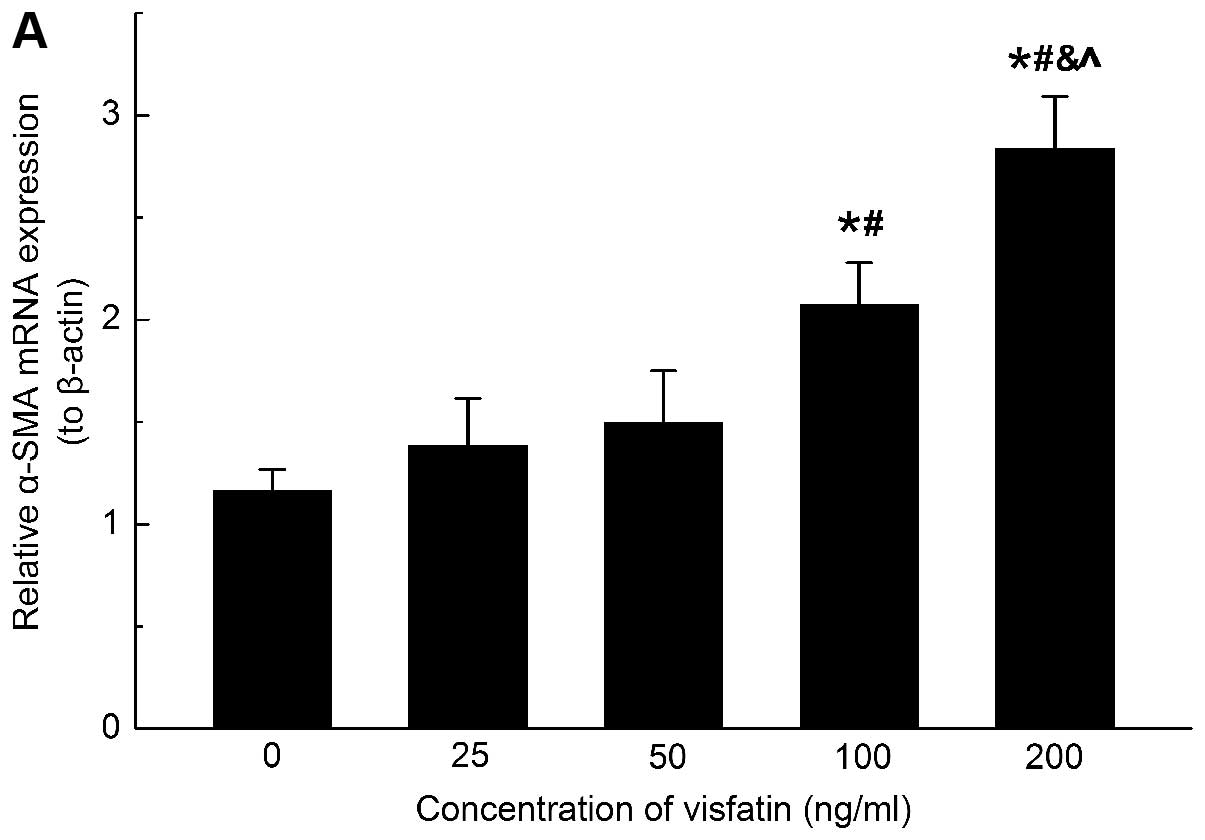 | Figure 2Visfatin increases the mRNA expression
and protein production of α-SMA in HSC. (A) Visfatin increases gene
and (B and C) protein expression of α-SMA in activated HSC in a
dose-dependent manner. Experiments were performed in triplicate.
For the quantitative polymerase chain reaction, values were
normalized to β-actin. For western blot analyses, β-actin was used
as an internal control. Error bars represent standard deviation.
*,#,&,^P<0.05, vs. 0, 25, 50 and 100 ng/ml
visfatin groups, respectively. α-SMA α-smooth muscle actin; HSC,
hepatic stellate cells. |
Visfatin increases collagen production in
HSC
When activated, HSC transform into
myofibroblast-like cells and produce substantial quantities of ECM
proteins, including collagen, fibronectin, undulin, elastin,
laminin, hyaluronan and proteoglycans (24). The accumulation of ECM proteins
distorts the hepatic architecture by forming fibrous scars and the
subsequent development of nodules in the regenerating hepatocytes
produces cirrhosis (25).
To determine whether visfatin affected the process
of liver fibrogenesis through upregulating the secretion of ECM,
the expression of collagen types I and III in the HSC was assessed
following treatment with different concentrations of visfatin.
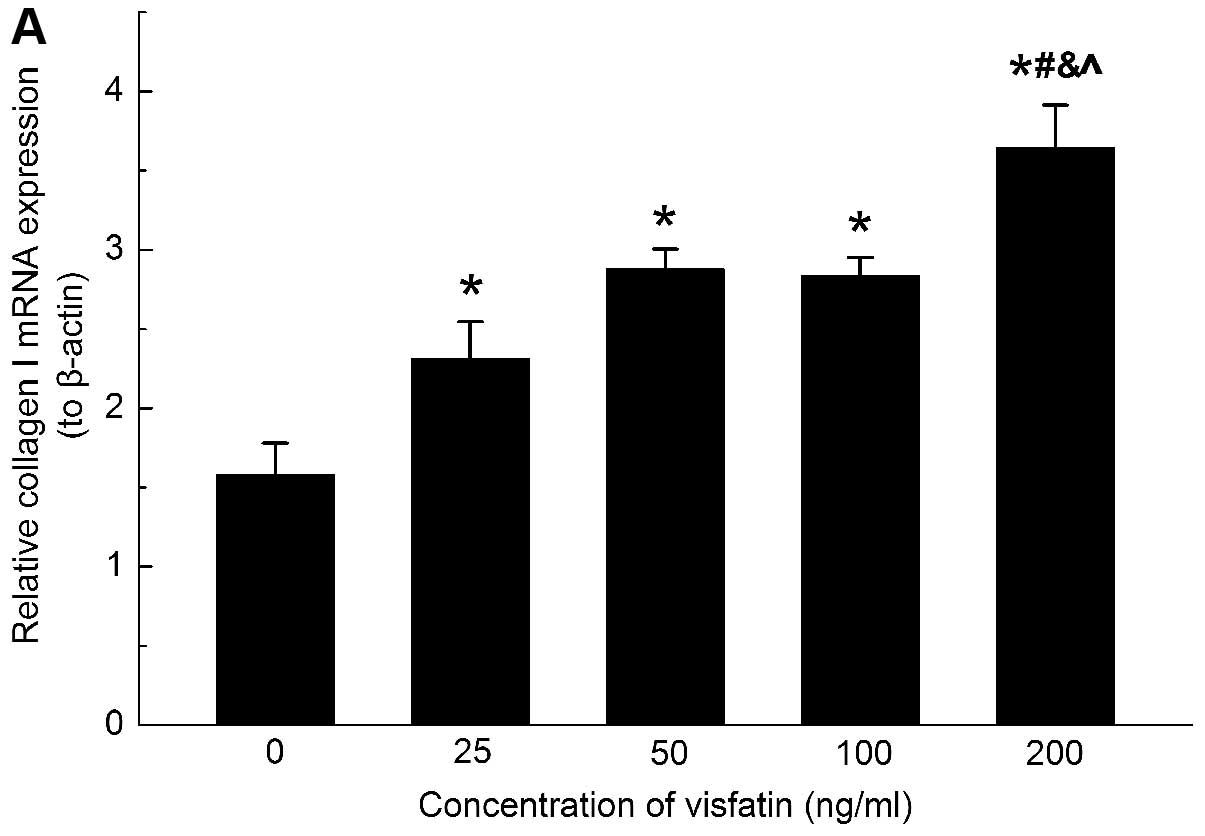
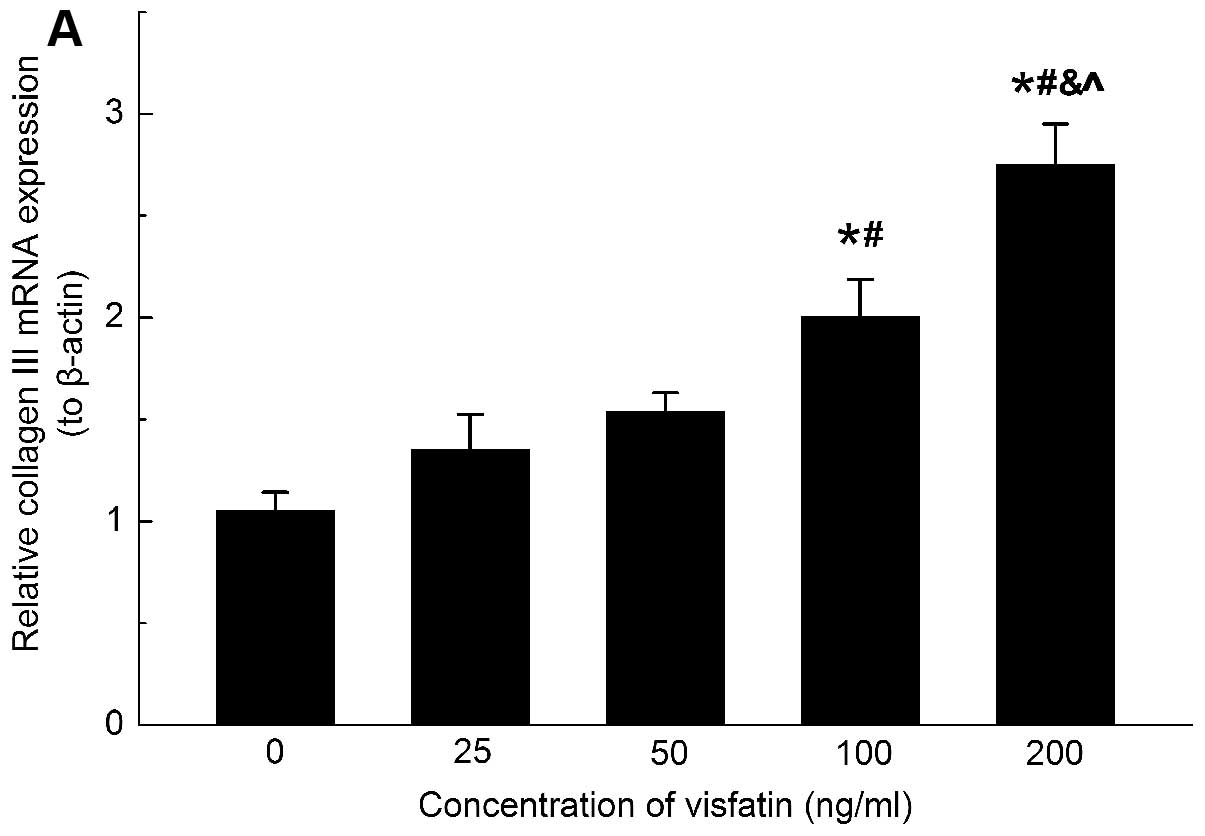
The mRNA expression levels of collagen types I and
III were increased upon visfatin stimulation (Figs. 3A and 4A). To further confirm the effects of
visfatin on the protein regulation of collagen types I and III, the
present study also examined the production of protein. The western
blot analysis revealed that visfatin induced a significant
elevation in the protein accumulation of collagen types I and III
in the HSC (Figs. 3B and 4B). These data indicated that visfatin
was involved in the upregulation of ECM secretion in the HSC.
Visfatin induces the expression of CTGF
in HSC
CTGF is regarded as the main downstream mediator of
the fibrogenic master cytokine TGF-β and has been implicated in the
pathogenesis of hepatic fibrosis (26). In HSC, CTGF is important in the
transition of HSC to myofibroblast-like cells and stimulates the
synthesis of ECM protein (23). In
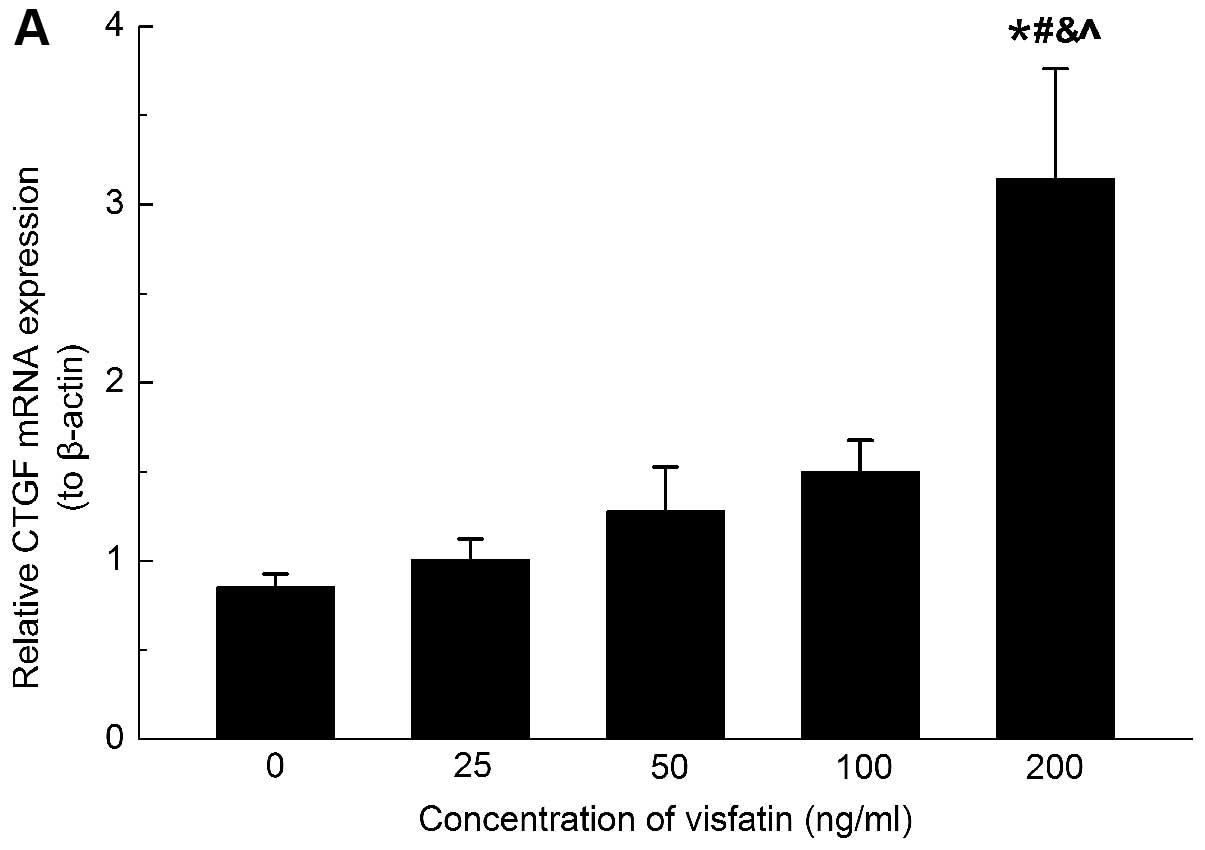
the present study, to determine whether CTGF was regulated in
response to visfatin, the gene and protein expression of CTGF was
analyzed in the HSC upon treatment with different concentrations of
visfatin. Only the concentration of 200 ng/ml visfatin increased
the mRNA expression of CTGF mRNA after 24 h of treatment (Fig. 5A), however, the protein expression
of CTGF was significantly induced at all experimental
concentrations of visfatin (Fig.
5B). These inconsistencies in mRNA and protein upregulation may
be due to post-transcriptional regulation.
Discussion
The present study identified several significant
findings: Visfatin upregulated the gene and protein production of
α-SMA, a marker of HSC activation; visfatin increased the mRNA and
protein expression of collagen types I and III in the HSC and the
production of CTGF was increased following exposure to visfatin.
These observations established the possibility that CTGF mediated
the visfatin-induced activation of HSC and it is therefore
suggested that visfatin may be involved in liver fibrogenesis.
The interaction between various adipokines,
including asleptin and adiponectin, is important in the process of
hepatic fibrogenesis (3). Leptin
can directly target HSC via activation of its receptor and all the
features of activated HSC are modulated by leptin in a
profibrogenic manner (4). Lower
levels of leptin in quiescent HSC are associated with increased
expression of adiponectin, an antifibrogenic factor (27). Adiponectin-knockout mice exhibited
more extensive fibrosis compared with wild-type animals following
chronic CCl4 intoxication (28). Visfatin is a recently identified
adipokine, which is produced and secreted mainly by visceral
adipose tissue. It can induce the proliferation of various cell
types, including human umbilical vein endothelial cells (29,30),
vascular smooth muscle cells (8)
and human osteoblasts (31).
However, the roles of visfatin in HSC activation, which may induce
HSC proliferation, remain to be elucidated. To the best of our
knowledge, the present study was the first to observe that visfatin
promoted the expression of α-SMA in HSC and provided direct
evidence that visfatin may be involved in the activation of
HSC.
When activated, HSC lose their retinoid shape, begin
to proliferate and the process of ECM synthesis begins to produce
fibrous scar tissue (32).
Collagen types I and III are the main components of the resulting
ECM. In the present study, collagen types I and III were induced
upon visfatin stimulation, indicating that visfatin may be involved
in processes of liver fibrogenesis.
Previous pilot studies have confirmed induction of
the mRNA and protein production of CTGF in cultured HSC in response
to visfatin (33). HSC are the
major cellular source of CTGF in the liver during hepatic
fibrogenesis (34). CTGF can be
regulated by TGF-β and is important in the overproduction of ECM in
activated HSC (35). The mRNA
levels of CTGF were markedly increased in cultured HSC and fibrotic
livers and the curcumin-induced inhibition of collagen type I gene
expression in activated HSC correlates with suppression in the gene
expression of CTGF (36). Knocking
down CTGF by small interfering RNA inhibited the expression of
carbon tetrachloride-induced collagen in HSC and in liver fibrosis
(37). Certain biological
functions of CTGF, including the induction of cell adhesion,
migration, proliferation and DNA synthesis, may explain the
visfatin-induced activation of HSC. CTGF itself can promote cell
adhesion and migration in a wide variety of cell types (38), induce fibroblast growth
factor-mediated cell proliferation and insulin-like growth
factor-promoted matrix synthesis (39) and enhance DNA synthesis in
chondrocytes and osteoblasts (40). CTGF-knockout mice exhibited damaged
chondrocyte proliferation and ECM composition (41). Fibroblasts obtained from
CTGF-deficient mice featured impaired induction of TGF-β-induced
adhesive signaling and were unable to respond to TGF-β by inducing
α-SMA and collagen type I (42).
Based on these previous studies, the present study hypothesized
that visfatin-induced expression of collagen in HSC may be mediated
by upregulating the expression of CTGF through TGF-β signaling.
In conclusion, the present study provided evidence
supporting the involvement of visfatin in HSC activation. However,
the underlying mechanism remains to be elucidated. Visfatin may
provide a novel potential therapeutic target for the treatment of
hepatic cirrhosis, particularly in patients with obesity and/or
diabetes.
Acknowledgements
This study was supported by the Chengdu City Science
and Technology Bureau of Sichuan Province, China (no.
13PPYB994SF-014), the Science and Technology Department of Sichuan
Province, China (nos. 2013FZ0085 and 2014CZ0002) and the National
Natural Science Foundation of China (no. 11072163).
References
|
1
|
Bertolani C and Marra F: The role of
adipokines in liver fibrosis. Pathophysiology. 15:91–101. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Wu D, Li H, Xiang G, et al: Adiponectin
and its receptors in chronic hepatitis B patients with steatosis in
china. Hepat Mon. 13:e60652013. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Tsochatzis EA, Papatheodoridis GV and
Archimandritis AJ: Adipokines in nonalcoholic steatohepatitis: from
pathogenesis to implications in diagnosis and therapy. Mediators
Inflamm. 2009:8316702009. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Marra F and Bertolani C: Adipokines in
liver diseases. Hepatology. 50:957–969. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Romacho T, Villalobos LA, Cercas E,
Carraro R, Sanchez-Ferrer CF and Peiró C: Visfatin as a novel
mediator released by inflamed human endothelial cells. PLoS One.
8:e782832013. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Van den Bergh R, Morin S, Sass HJ, et al:
Monocytes contribute to differential immune pressure on R5 versus
X4 HIV through the adipocytokine visfatin/NAMPT. PLoS One.
7:e350742012. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Yu XY, Qiao SB, Guan HS, Liu SW and Meng
XM: Effects of visfatin on proliferation and collagen synthesis in
rat cardiac fibroblasts. Horm Metab Res. 42:507–513. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Wang P, Xu TY, Guan YF, Su DF, Fan GR and
Miao CY: Perivascular adipose tissue-derived visfatin is a vascular
smooth muscle cell growth factor: role of nicotinamide
mononucleotide. Cardiovasc Res. 81:370–380. 2009. View Article : Google Scholar
|
|
9
|
Song HK, Lee MH, Kim BK, et al: Visfatin:
a new player in mesangial cell physiology and diabetic nephropathy.
Am J Physiol Renal Physiol. 295:F1485–F1494. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Liu XJ, Yang L, Mao YQ, et al: Effects of
the tyrosine protein kinase inhibitor genistein on the
proliferation, activation of cultured rat hepatic stellate cells.
World J Gastroenterol. 8:739–745. 2002.PubMed/NCBI
|
|
11
|
Friedman SL: Hepatic stellate cells:
protean, multifunctional, and enigmatic cells of the liver. Physiol
Rev. 88:125–172. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Nieto N: A systems biology approach for
understanding the collagen regulatory network in alcoholic liver
disease. Liver Int. 32:189–198. 2012. View Article : Google Scholar
|
|
13
|
Zhu Y, Men R, Wen M, Hu X, Liu X and Yang
L: Blockage of TRPM7 channel induces hepatic stellate cell death
through endoplasmic reticulum stress-mediated apoptosis. Life Sci.
94:37–44. 2014. View Article : Google Scholar
|
|
14
|
Liu XJ, Yang L, Wu HB, Qiang O, Huang MH
and Wang YP: Apoptosis of rat hepatic stellate cells induced by
anti-focal adhesion kinase antibody. World J Gastroenterol.
8:734–738. 2002.PubMed/NCBI
|
|
15
|
Kim JG, Kim EO, Jeong BR, et al: Visfatin
stimulates proliferation of MCF-7 human breast cancer cells. Mol
Cells. 30:341–345. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Patel ST, Mistry T, Brown JE, et al: A
novel role for the adipokine visfatin/pre-B cell colony-enhancing
factor 1 in prostate carcinogenesis. Peptides. 31:51–57. 2010.
View Article : Google Scholar
|
|
17
|
Höinghaus R, Hewicker-Trautwein M and
Mischke R: Immunocytochemical differentiation of neoplastic and
hyperplastic canine epithelial lesions in cytologic imprint
preparations. Vet J. 173:79–90. 2007. View Article : Google Scholar
|
|
18
|
Gilliver SC, Ruckshanthi JP, Atkinson SJ
and Ashcroft GS: Androgens influence expression of matrix proteins
and proteolytic factors during cutaneous wound healing. Lab Invest.
87:871–881. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Maeda E, Shelton JC, Bader DL and Lee DA:
Differential regulation of gene expression in isolated tendon
fascicles exposed to cyclic tensile strain in vitro. J Appl Physiol
(1985). 106:506–512. 2009. View Article : Google Scholar
|
|
20
|
Li L, Chen GP, Yang Y, Ye Y, Yao L and Hu
SJ: Chronic inhibition of farnesyl pyrophosphate synthase
attenuates cardiac hypertrophy and fibrosis in spontaneously
hypertensive rats. Biochem Pharmacol. 79:399–406. 2010. View Article : Google Scholar
|
|
21
|
Chen A and Zheng S: Curcumin inhibits
connective tissue growth factor gene expression in activated
hepatic stellate cells in vitro by blocking NF-kappaB and ERK
signalling. Br J Pharmacol. 153:557–567. 2008. View Article : Google Scholar
|
|
22
|
Dodig M, Ogunwale B, Dasarathy S, Li M,
Wang BC and McCullough AJ: Differences in regulation of type I
collagen synthesis in primary and passaged hepatic stellate cell
cultures: the role of alpha5beta1-integrin. Am J Physiol
Gastrointest Liver Physiol. 293:G154–G164. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Troeger JS, Mederacke I, Gwak GY, Dapito
DH, Mu X, Hsu CC, Pradere JP, et al: Deactivation of hepatic
stellate cells during liver fibrosis resolution in mice.
Gastroenterology. 143:1073–1083. e10222012. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Ponomarenko Y, Leo MA, Kroll W and Lieber
CS: Effects of alcohol consumption on eight circulating markers of
liver fibrosis. Alcohol and Alcoholism. 37:252–255. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Cheng F, Li Y, Feng L and Li S: Hepatic
stellate cell activation and hepatic fibrosis induced by
ischemia/reperfusion injury. Transplant Proc. 40:2167–2170. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Gressner OA, Lahme B, Demirci I, Gressner
AM and Weiskirchen R: Differential effects of TGF-beta on
connective tissue growth factor (CTGF/CCN2) expression in hepatic
stellate cells and hepatocytes. J Hepatol. 47:699–710. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Jiang JX, Mikami K, Shah VH and Torok NJ:
Leptin induces phagocytosis of apoptotic bodies by hepatic stellate
cells via a Rho guanosine triphosphatase-dependent mechanism.
Hepatology. 48:1497–1505. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Kamada Y, Tamura S, Kiso S, et al:
Enhanced carbon tetrachloride-induced liver fibrosis in mice
lacking adiponectin. Gastroenterology. 125:1796–1807. 2003.
View Article : Google Scholar
|
|
29
|
Adya R, Tan BK, Punn A, Chen J and Randeva
HS: Visfatin induces human endothelial VEGF and MMP-2/9 production
via MAPK and PI3K/Akt signalling pathways: novel insights into
visfatin-induced angiogenesis. Cardiovasc Res. 78:356–365. 2008.
View Article : Google Scholar
|
|
30
|
Kim SR, Bae SK, Choi KS, et al: Visfatin
promotes angiogenesis by activation of extracellular
signal-regulated kinase 1/2. Biochem Biophys Res Commun.
357:150–156. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Xie H, Tang SY, Luo XH, et al:
Insulin-like effects of visfatin on human osteoblasts. Calcif
Tissue Int. 80:201–210. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Brenner DA: Molecular pathogenesis of
liver fibrosis. Trans Am Clin Climatol Assoc. 120:361–368.
2009.PubMed/NCBI
|
|
33
|
Liu Y, Liu H, Meyer C, Li J, Nadalin S,
Konigsrainer A, Weng H, et al: Transforming growth factor-beta
(TGF-beta)-mediated connective tissue growth factor (CTGF)
expression in hepatic stellate cells requires Stat3 signaling
activation. J Biol Chem. 288:30708–30719. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Paradis V, Dargere D, Bonvoust F, Vidaud
M, Segarini P and Bedossa P: Effects and regulation of connective
tissue growth factor on hepatic stellate cells. Lab Invest.
82:767–773. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Leask A, Holmes A, Black CM and Abraham
DJ: Connective tissue growth factor gene regulation. Requirements
for its induction by transforming growth factor-beta 2 in
fibroblasts. J Biol Chem. 278:13008–13015. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Zheng S and Chen A: Curcumin suppresses
the expression of extracellular matrix genes in activated hepatic
stellate cells by inhibiting gene expression of connective tissue
growth factor. Am J Physiol Gastrointest Liver Physiol.
290:G883–G893. 2006. View Article : Google Scholar
|
|
37
|
Li G, Xie Q, Shi Y, et al: Inhibition of
connective tissue growth factor by siRNA prevents liver fibrosis in
rats. J Gene Med. 8:889–900. 2006. View
Article : Google Scholar : PubMed/NCBI
|
|
38
|
Crean JK, Finlay D, Murphy M, et al: The
role of p42/44 MAPK and protein kinase B in connective tissue
growth factor induced extracellular matrix protein production, cell
migration, and actin cytoskeletal rearrangement in human mesangial
cells. J Biol Chem. 277:44187–44194. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Gore-Hyer E, Pannu J, Smith EA,
Grotendorst G and Trojanowska M: Selective stimulation of collagen
synthesis in the presence of costimulatory insulin signaling by
connective tissue growth factor in scleroderma fibroblasts.
Arthritis Rheum. 48:798–806. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Kubota S and Takigawa M: Role of
CCN2/CTGF/Hcs24 in bone growth. Int Rev Cytol. 257:1–41. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Ivkovic S, Yoon BS, Popoff SN, et al:
Connective tissue growth factor coordinates chondrogenesis and
angiogenesis during skeletal development. Development.
130:2779–2791. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Shi-wen X, Stanton LA, Kennedy L, et al:
CCN2 is necessary for adhesive responses to transforming growth
factor-beta1 in embryonic fibroblasts. J Biol Chem.
281:10715–10726. 2006. View Article : Google Scholar : PubMed/NCBI
|