Introduction
Daily rhythms, which are essential for normal
behavior and numerous physiological processes, including the
sleep-wake cycle, hormone secretion, core body temperature,
metabolism and cell cycle control are generated by the mammalian
circadian timing system (1). These
circadian clock rhythms are controlled at the molecular level
through interactions between positive and negative feedback loops,
which consist of various important clock regulators (2,3). One
molecular clock model, which has been well established, encompasses
a feedback system that involves the heterodimer transcriptional
factors Clock and Bmal1, two cryptochromes (Cry1 and Cry2) and
three period regulator genes (Per1, Per2 and Per3); among these,
Clock acts as the master controller gene and serves as the positive
regulator in the primary feedback loop (4).
Previous studies have provided evidence which
suggested that disruption of the circadian rhythm or genetic
defects in the circadian core genes were associated with cancer
biology (5,6). In addition, disturbances of circadian
gene expression levels have been reported in various types of human
malignancies (7–13). Soták et al (14) demonstrated that circadian genes may
have a role in colorectal carcinoma (CRC) development and
progression. The circadian rhythmicity of certain circadian genes,
including Per1, Per2 and Bmal1, was significantly reduced in CRC
tumor tissue compared with that of a healthy colon (14). CRC is the most common type of solid
tumor worldwide (15); however,
further studies are required in order to fully elucidate the
molecular mechanisms of CRC and to control this cancer more
effectively (16,17).
It was previously reported that the circadian gene
human Clock (hClock) was highly expressed in CRC tissues compared
with that of peritumoral tissues in CRC patients, and was found to
be strongly associated with late tumor-node-metastasis (TNM) stage
as well as positive lymph node metastasis (18). Similar studies have also
demonstrated the overexpression of the hClock gene in human CRC
tissues (19,20). Furthermore, Alhopuro et al
(21) reported that ~53% of
microsatellite instability CRCs had point mutations in the hClock
gene DNA sequence and Zhou et al (22) showed that genetic variants in the
hClock gene have a significant effect on the risk of mortality in
CRC patients. Overall, these results strongly suggested a critical
role for hClock in CRC progression. However, the function of hClock
in CRC remains to be fully elucidated. It has been estimated that
up to 10% of all genes within the mammalian genome are regulated by
circadian genes (23). Numerous
genes regulating the cell cycle, including Wee1, cyclin D1 and
c-Myc, have been reported to be controlled by the rhythmic activity
of core clock genes (24–26). Molecular clockworks regulate the
expression of these clock-controlled genes associated with the cell
cycle, apoptosis and other pathways in cells; therefore, the
aberration of core circadian genes, such as hClock, may result in
the dysregulation of these processes and lead to tumor development
(27,28).
The aim of the present study was to investigate the
molecular changes and function of hClock in human CRC progression
through hClock upregulation and knock-down experiments in
vitro and in vivo.
Materials and methods
Ethics statement
The present study was approved by the Institutional
Review Board of Huashan Hospital affiliated to Fudan University
(HIRB; Shanghai, China). All in vivo experiments were
performed strictly in accordance with the National Institutes of
Health Guide for the Care and Use of Laboratory Animals and were
approved by Animal Care and Use Committee of Shanghai Medical
College of Fudan University (Shanghai, China).
Cell culture
The two human CRC cell lines SW480 and SW620, as
well as the human embryonic kidney cell line 293T were cultured in
Dulbecco’s modified Eagle’s medium (DMEM; Gibco Life Technologies,
Carlsbad, CA, USA) with 10% (v/v) newborn calf serum (Amresco LLC,
Solon, OH, USA), 100 U/ml penicillin and 100 ng/ml streptomycin
(both Amresco LLC) at 37°C in 5% CO2. All cell lines
were obtained from the Cell Bank of the Chinese Academy of Sciences
(Shanghai, China).
Western blot analysis
Cells were homogenized in ice-cold radio
immunoprecipitation assay lysis buffer (Beijing Dingguo
Biotechnology Co., Ltd., Beijing, China). Following centrifugation
(12,000 × g, 10 min at 4°C), the supernatant was collected and the
protein concentration was determined using a bicinchoninic acid
protein assay kit (HyClone-Pierce, Logan, UT, USA). Equal amounts
of proteins (40 μg) were extracted and separated using 10%
SDS-PAGE gels, the proteins were then electrotransferred to a
polyvinylidene fluoride membrane (Pall-Gelman, Port Washington, NY,
USA). The membrane was blocked with 5% fat-free milk powder for 1
h, and then incubated with the primary antibodies (1:500 dilution)
and GAPDH monoclonal antibody (1:1,000 dilution) in blocking buffer
for 18 h at 4°C. The membranes were subsequently incubated with a
secondary antibody (1:4,000 dilution) for 1 h at room temperature.
After washing, immunodetection was performed with enhanced
chemiluminescence (ECL-PLUS/kit; Amersham, Piscataway, NJ, USA) and
exposed on an X-ray film. Subsequently, the image was processed and
analyzed by ImageJ (v2.1.4.7). GAPDH was used as the loading
control. The antibodies anti-hClock (cat. no. ab43106), rabbit
monoclonal anti-B cell lymphoma (Bcl-2)-associated X protein (Bax;
cat. no. ab32503) and mouse monoclonal anti-Bcl-2 homology 3
interacting domain death agonist (Bid; cat. no. ab114051) were
obtained from Abcam (Cambridge, UK), while rabbit monoclonal
anti-GAPDH (cat. no. 3683), anti-AKT (cat. no. 4685) and rabbit
monoclonal anti-phosphorylated (p-)AKT (cat. no. 3783) were
obtained from Cell Signaling Technology, Inc. (Danvers, MA, USA).
The secondary antibody was alkaline phosphatase-labeled anti-rabbit
immunoglobulin G (Beijing Dingguo Biotechnology Co., Ltd.).
Plasmid construction
The full-length complementary (c)DNA of hClock
(Genbank accession number, NM_004898) was amplified and inserted
into the pcDNA3-Flag vector (Invitrogen Life Technologies,
Carlsbad, CA, USA) in order to generate the pcDNA3-Flag-hClock
expression plasmid. The cDNA fragment of Flag-hClock was then
inserted into pGV186 retroviral vector (Shanghai GeneChem Co.,
Ltd., Shanghai, China) to generate pGV186-Flag-hClock. All the
constructs were confirmed by DNA sequencing, which was performed by
Shanghai GeneChem Co., Ltd.
RNA interference
For small interfering (si)RNA-mediated hClock
silencing, the following target siRNA sequences of hClock
(NM_004898) were used: hClock RNAi-1, 5′-ACGAGAACTTGGCATTGAA-3′;
hClock RNAi-2, 5′-CAAGATTCTGGGTCAGATA-3′; hClock RNAi-3,
5′-CACACATAGGCCATCTTAT-3′; and hClock RNAi-4,
5′-TTCAACTTTCTTCTGGAAA-3′.
The RNA duplexes were synthesized by Genepharma Co.,
Ltd (Shanghai, China). Then, a pGV113 lentiviral vector, which had
an independent open reading frame of red fluorescence protein, was
used to produce double-stranded siRNA. In order to construct the
hairpin siRNA expression cassette, cDNA nucleotides of hClock siRNA
were synthesized, annealed and inserted into pGV113 by Shanghai
GeneChem Co., Ltd. A total of four groups of pGV113-hClock small
hairpin (sh)RNA were constructed (Shanghai GeneChem Co., Ltd,
Shanghai, China). The targeting sequences for hClock were as
follows: hClock RNAi-1, 5′-CCGGCGACGAGA
ACTTGGCATTGAACTCGAGTTCAATGCCAAGTTCTCGTCGTTTTTG-3′; hClock RNAi-2,
5′-CCGGGCCAAGATTCTGGGTCAGATACTCGAGTATCTGACCCAGAATCTTGGCTTTTTG-3′;
hClock RNAi-3,
5′-CCGGCGCACACATAGGCCATCTTATCTCGAGATAAGATGGCCTATGTGTGCGTTTTTG-3′;
and hClock RNAi-4,
5′-CCGGCGTTCAACTTTCTTCTGGAAACTCGAGTTTCCAGAAGAAAGTTGAACGTTTTTG-3′.
Lentiviral vector pGV113 (Shanghai GeneChem Co., Ltd.) was used as
a negative control.
Retrovirus packing, transduction and
infection of target cells
Retroviruses carrying the hClock cDNA or hClock
siRNA were generated by co-transfection of recombinant pGV186 or
pGV113 plasmids, respectively, with pHelper1.0 and pHelper2.0
plasmids (Shanghai GeneChem Co., Ltd.) into 293T cells using
Lipofectamine 2000 (Invitrogen Life Technologies). These 293T cells
were then cultured in DMEM supplemented with 10% newborn calf serum
in a 37°C incubator containing 5% CO2. Forty-eight hours
post-transfection, the supernatant was collected by centrifugation
at 1,000 × g for 10 min, and the culture medium containing the
recombinant virus was harvested and purified using a a
0.45-μm filter (Shanghai GeneChem Co., Ltd.). Target cells
were seeded (5×105/well) into six-well plates and
incubated with recombinant virus supplemented with 5 μg/ml
polybrene (Shanghai GeneChem Co., Ltd.) for a spin infection
procedure. The SW480 cells were transduced with the retroviruses
containing hClock or control sequence plasmids, while the SW620
cells were transduced with the retroviruses containing hClock siRNA
or control sequence plasmids. Two weeks after infection, the stable
cells infected with the target retrovirus were selected.
Cell viability assay
Following 12 h of transfection, the MTT standard
method was used to determine the cell growth. Cells were seeded
into the 96-well plates at a density of 1,000 cells/well and
cultured at 37°C in a humidified atmosphere containing 5%
CO2. Following 96 h, the cells were incubated with 100
μg/well MTT solution (Sigma-Aldrich, St. Louis, MO, USA) for
4 h, the medium was replaced with 150 μl dimethyl sulfoxide
(DMSO; Sigma-Aldrich) and agitated for 10 min. Absorbance was
recorded at 490 nm using an automatic microwell plate reader (3350;
Bio-Rad Laboratories, Inc., Hercules, CA, USA). Cell viability was
calculated as percentage of that of the untreated cells.
Flow cytometric analysis
Cells were harvested through 0.25% trypsin (Amresco
LLC) digestion at 72 h post-seeding, and then washed with ice-cold
phosphate-buffered saline (Sigma-Aldrich). Cells were re-suspended
in 100 μl 1X binding buffer (Invitrogen Life Technologies)
at a concentration of 1×106 cells/ml and double stained
with Annexin V and propidium iodide (PI; Invitrogen Life
Technologies). Following 30 min of incubation at room temperature
in the dark, cells were added to 400 μl 1X binding buffer
and immediately analyzed by flow cytometry (FACS Calibur; BD
Biosciences, San Jose, CA, USA).
Tumor growth in xenograft models
A total of 20 female Balb/c nude mice (4–5 weeks
old, ~20 g) were obtained from Shanghai Experimental Animal Center
and maintained in a pathogen-free environment. The mice were
divided at random into two groups (n=10/group), housed two per cage
and provided with food and water ad libitum. All of the mice
were exposed to a 12 h light/12 h dark cycle. The two groups of
mice were subcutaneously injected with SW480-hClock or
SW480-control infected cells (1×106) in each right
flank. The tumors which developed were measured weekly using a
caliper, and the diameters were recorded. Tumor volume
(cm3) was calculated using the following formula: Length
× width2 × 0.5326. The animals were sacrificed by
cervical dislocation six weeks post-injection of cells. The tumors
were then resected and snap-frozen in liquid nitrogen for further
analysis. Tumor tissues were fixed in 10% formalin for at least 24
h, then embedded in paraffin wax and sectioned (4 μm) for
histopathological evaluation. The sections were stained with
hematoxylin and eosin using a standard protocol and analyzed by
light microscopy (DM IL LED; Leica, Wetzlar, Germany). All animal
procedures in the present study were conducted in accordance with
the guidelines of the Institutional Animal Care and Use Committee.
All mice were treated humanely throughout the experimental
period.
Statistical analysis
Values are presented as mean ± standard deviation,
unless stated otherwise. Statistical significance of the in
vitro and in vivo studies was analyzed using Student’s
t-test. All statistical analyses were conducted using SPSS
19.0 software (International Business Machines, Armonk, NY, USA).
All P-values were two-sided and P<0.05 was considered to
indicate a statistically significant difference between values.
Results
hClock promotes CRC cell proliferation in
vitro
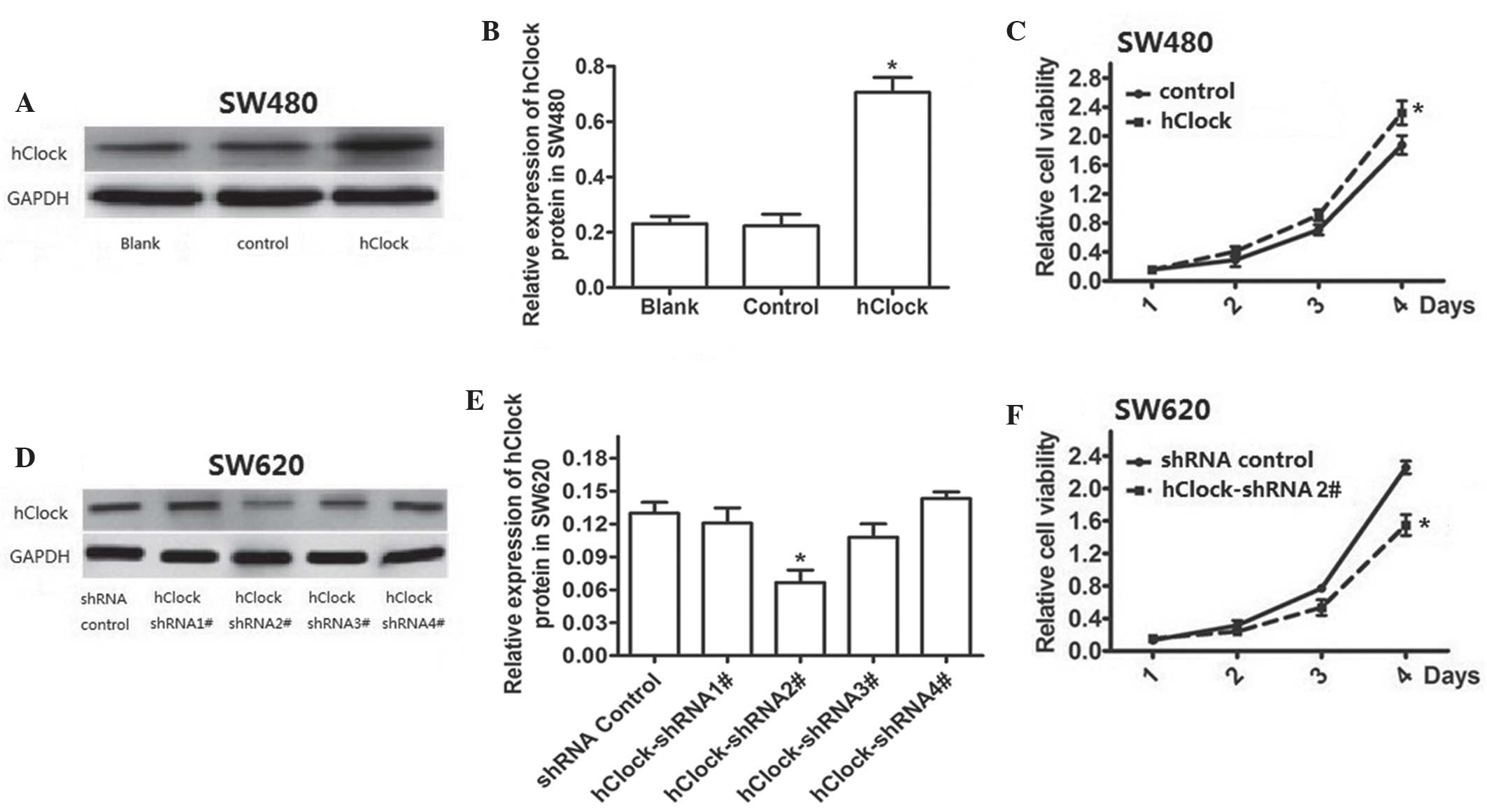
In order to investigate the role of hClock in CRC,
the effects of upregulation or knockdown of hClock expression on
the proliferation of CRC cells were examined. A lentivirus
expression system was used to overexpress exogenous hClock in SW480
cells, which were found to have a relatively low endogenous hClock
expression compared with that of the loading control; in addition,
hClock was confirmed to be significantly upregulated in transfected
cells compared with that of the control (P<0.05) (Fig. 1A and B). As shown in Fig. 1C, overexpression of hClock had a
significant effect on promoting cellular proliferation in SW480
cells by four days post-transfection (P<0.05). Endogenous hClock
expression in CRC SW620 cells was successfully knocked down using
lentivirus-mediated hClock shRNAs (Fig. 1D and E). The results showed that
knockdown of endogenous hClock expression resulted in the
significant inhibition of cell proliferation in SW620 cells
compared with that of the shRNA control group (P<0.05) (Fig. 1F).
hClock inhibits apoptosis of CRC
cells
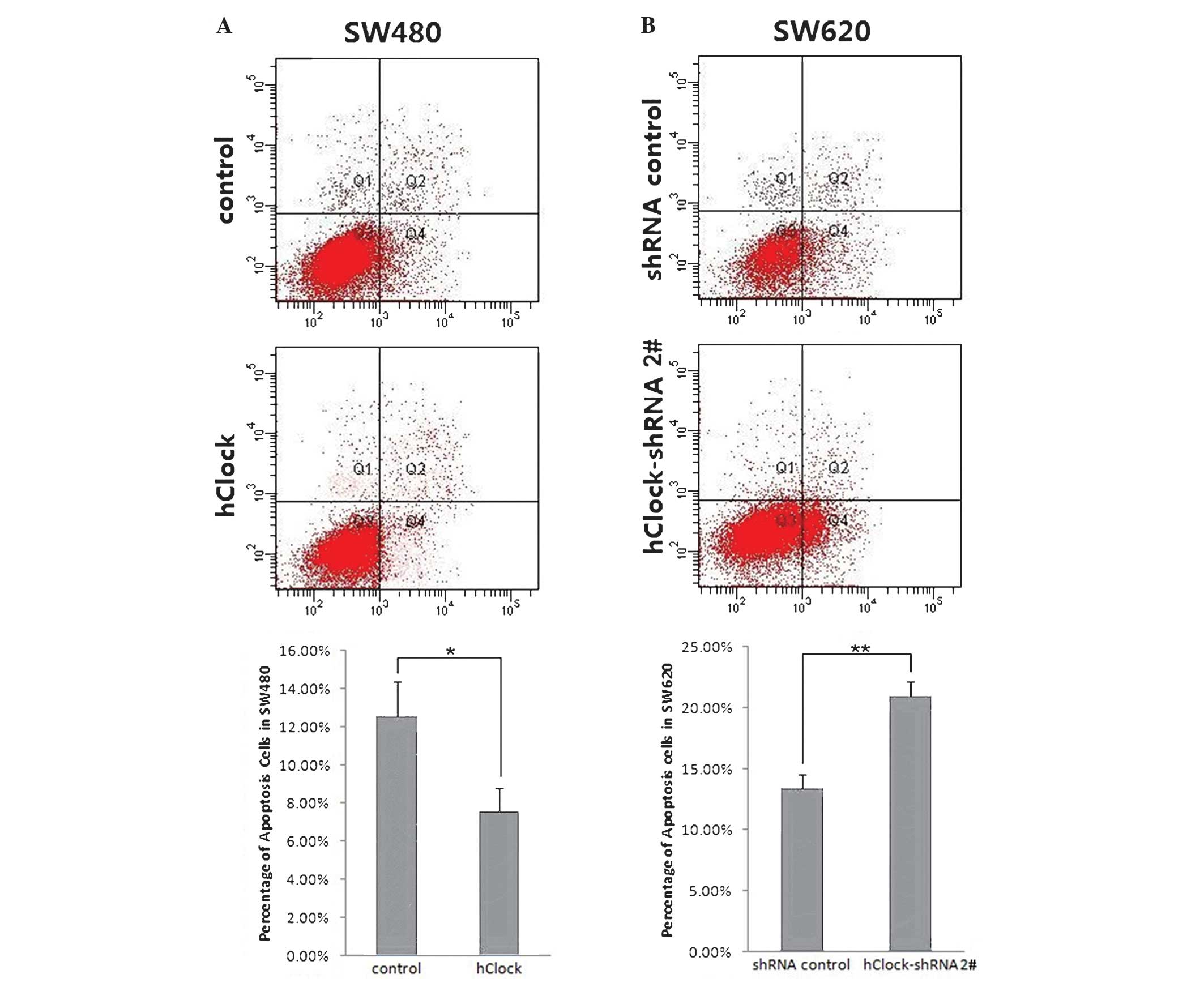
The effects of hClock upregulation or knockdown on
apoptosis were investigated in CRC cells using flow cytometric
analysis. The results showed that overexpression of hClock
significantly inhibited apoptosis in SW480 cells compared with that
of the control group (P<0.05) (Fig.
2A). By contrast, knockdown of hClock expression markedly
enhanced apoptosis in SW620 cells compared with that of the
shRNA-transfected control group (P<0.01) (Fig. 2B). These results indicated that
hClock inhibited CRC cell apoptosis and hClock knockdown may, at
least in part, have contributed to increased spontaneous apoptosis
in CRC cells in vitro.
Altered expression of hClock is
associated with changes in the expression of apoptosis-associated
proteins Bax, Bid and p-AKT
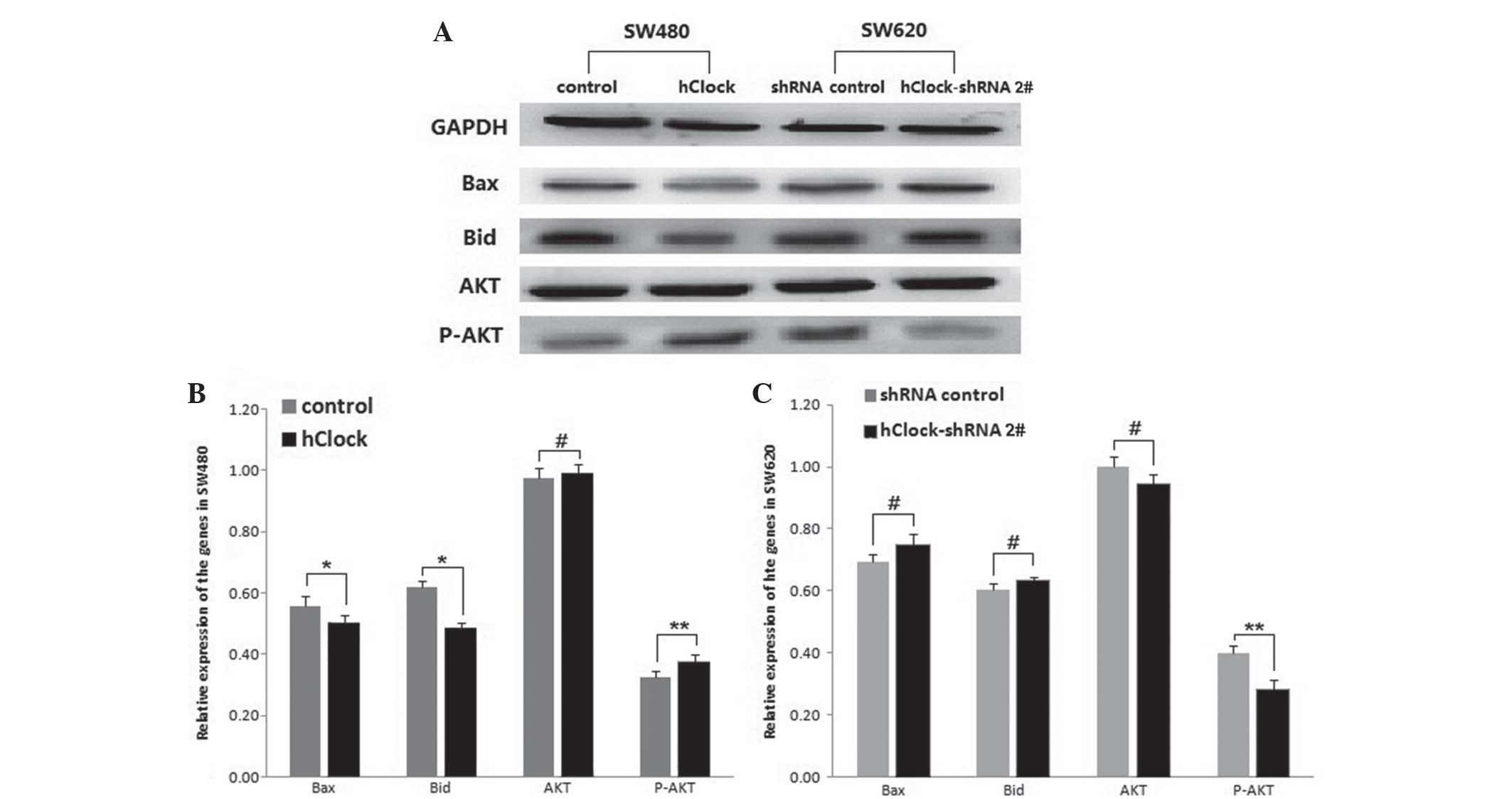
In order to further investigate the association
between hClock expression and CRC cell apoptosis, the expression of
certain apoptosis-associated proteins, including Bax, Bid, AKT and
p-AKT, in CRC cells with hClock overexpression or silencing. As
shown in Fig. 3A and B, expression
levels of Bax and Bid were significantly decreased in SW480 cells
overex-pressing hClock compared with those of the vector
only-treated SW480 cells (P<0.05), whereas upregulated hClock
resulted in a significant increase in p-AKT expression (P<0.01).
By contrast, silencing of endogenous hClock expression resulted in
a marked decrease in p-AKT expression compared with that of the
shRNA-transfected controls (P<0.01) (Fig. 3A and C), whereas Bax and Bid
expression levels were not significantly altered. Total AKT
expression levels were unaffected by changes in hClock expression
(P>0.05) (Fig. 3A–C).
Overexpression of hClock promotes growth
of CRC cells in vivo
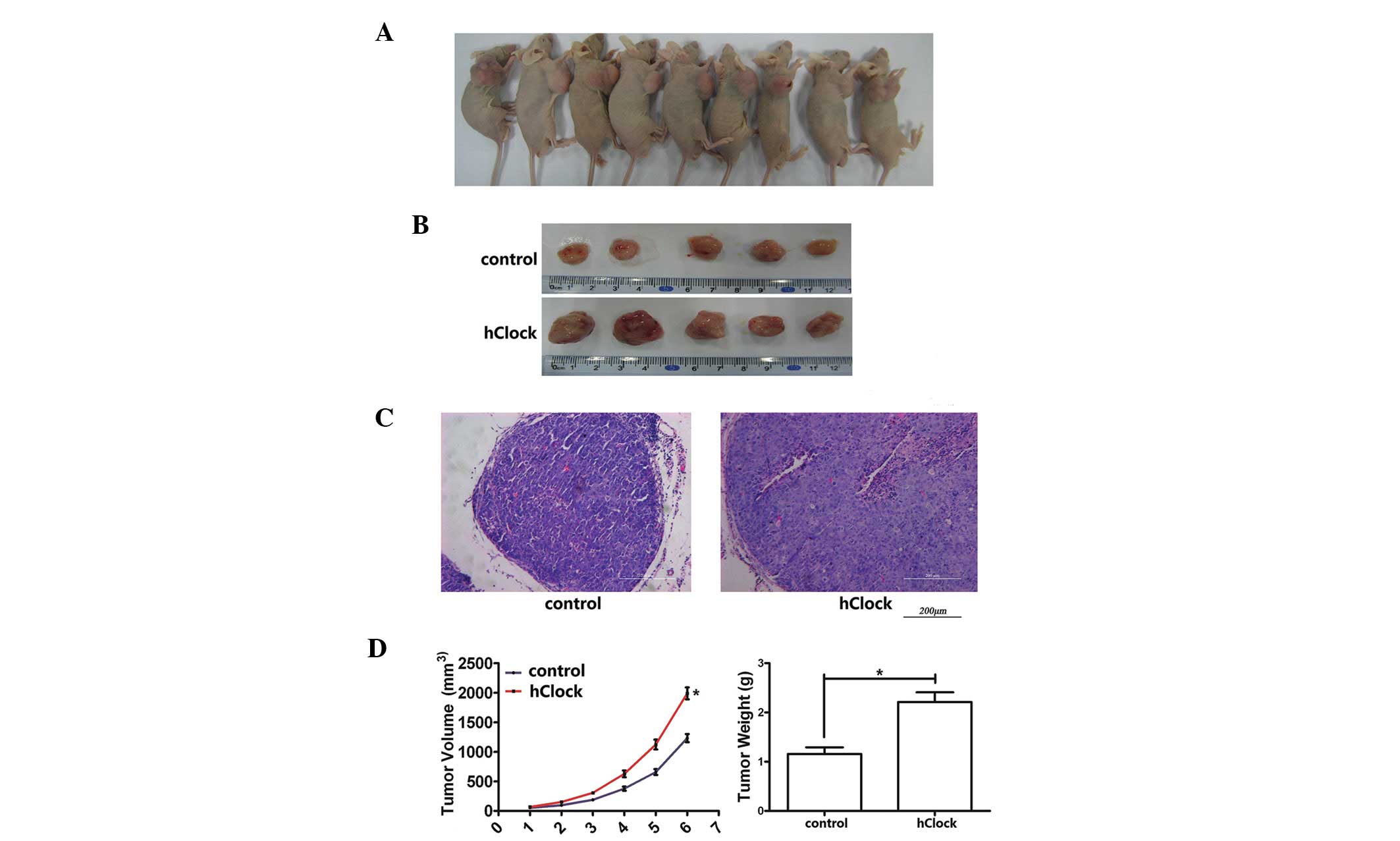
In order to examine whether hClock has an effect on
CRC cell growth in vivo, a xenograft model of CRC cells in
BALB/c nude mice was performed. SW480 CRC tumor cells, which were
specifically transduced with hClock or the control vector only,
were implanted subcutaneously into the right flank of the nude mice
(10 mice/group; 1×106 cells/injection). The volume of
the developed tumors was measured once a week for six weeks. All
the animals had developed a subcutaneous tumor at one week
post-injection of transfected-CRC cells. The mean tumor volume was
higher in the hClock over-expression group at two weeks
post-injection compared with that of the control group; in
addition, the difference in mean tumor volume increased between the
two groups from two to six weeks post-transplantation, with
significant differences observed at six weeks. Furthermore,
upregulated expression of hClock accounted for a 61.43% increase in
tumor volume and a 91.38% increase in tumor weight at day 42
post-injection compared with that of the control group (Fig. 4); the developed tumor volume and
weight were 1990.0±101.4 mm3 and 2.21±0.20 g,
respectively, in the hClock overexpression group, compared with
1232.7±69.2 mm3 and 1.16±0.14 g in the control group,
respectively (P<0.01).
Discussion
CRC is one of the most prevalent causes of
malignancy-associated mortality worldwide. Despite improvements in
surveillance and clinical treatment strategies for CRC, the
mortality remains high (15). It
is therefore important to identify the factors which are associated
with CRC tumor progression and the mechanisms by which they proceed
(29).
A previous study demonstrated that expression of the
circadian gene hClock was significantly increased in CRC patients
compared with that in adjacent normal tissue and was strongly
correlated with the late stages of TNM classification (18). It was also shown that genetic
variants in the hClock gene have a significant effect on the risk
of mortality in CRC patients (22). Overall, these studies indicated
that hClock overexpression may be a potential biomarker for CRC and
therefore, the role of this gene in CRC requires further
elucidation.
In the present study, in order to further
investigate the role of hClock in promoting CRC progression, hClock
was exogenously overexpressed in SW480 CRC cells, which have a
relatively low endogenous hClock expression, through transfection
of a lentivirus hClock expression system. Overexpression of hClock
was found to accelerate SW480 cell proliferation as well as inhibit
cell apoptosis. A xenograft model was then used to evaluate the
function of hClock in promoting CRC cell growth in vivo. The
results showed that tumor volume and weight were significantly
increased in mice in the hClock overexpression group compared with
those injected with control SW480 cells by six weeks
post-injection. This therefore suggested that hClock had an active
role in promoting CRC proliferation and development in vitro
and in vivo.
It remained to be elucidated whether downregulation
of hClock had an effect on CRC cell growth. Therefore, in the
present study, an shRNA plasmid targeting hClock was constructed
and stably transfected into SW620 CRC cells in order to investigate
the effect of hClock silencing on the growth of CRC cells. MTT
assays revealed that silencing hClock with shRNA significantly
reduced SW620 cell proliferation and enhanced apoptosis in
vitro. These data provided evidence for the function of hClock
as a key regulator of CRC cell growth, which therefore indicated
that hClock may be a promising target for CRC treatment.
Circadian genes are important for regulating certain
downstream clock-controlled genes (CCGs), including a variety of
tumor-associated genes (30). In
the present study, in order to further explore the underlying
mechanism of the positive role of hClock in CRC progerssion, the
expression levels of apoptosis-associated proteins were examined in
response to hClock dysregulation. The results showed that hClock
overexpression resulted in a significant decrease in Bax and Bid
expression as well as a significant increase in p-AKT expression.
By contrast, hClock silencing resulted in a significant reduction
in p-AKT expression; however, total AKT levels were unaffected by
up- or downregulation of hClock. Apoptosis, or programmed cell
death, is a normal cellular function which controls excessive
proliferation by eliminating ‘unnecessary’ cells; cancer cells are
known to develop certain mechanisms in order to avoid apoptosis and
prolong their survival (31). Bax
and Bid proteins are members of the Bcl-2 family and function as
promoters of cell apoptosis (32).
Following an appropriate stimulus, the Bax or Bid product primarily
enhances apoptotic cell death (33). By contrast, AKT functions in an
anti-apoptotic pathway and is activated by a dual regulatory
mechanism, which involves translocation to the plasma membrane as
well as phosphorylation at Thr308 and Ser473 (34). The mechanism by which activated AKT
(p-AKT) protects cells from death is multifactorial. Through
activating or inhibiting several downstream components of the cell
death machinery, including Bcl-2-associated death promoter,
caspase-9, fork-head homologue in rhabdomyosarcoma, nuclear
factor-κB and p53, via phosphorylation, p-AKT regulates numerous
cell activities, including cell proliferation, differentiation,
apoptosis and migration (31). The
results of the present study indicated that hClock promoted the
activity of anti-apoptosis gene p-AKT and inhibited the expression
of the pro-apoptotic genes Bax and Bid, thereby inhibiting the
apoptosis of CRC cells. By contrast, targeted inhibition of hClock
expression partially reduced the levels of p-AKT, which may be used
as a novel target of CRC therapy. Overall, these data suggested
that the p-AKT, Bax and Bid may work as CCGs and have important
roles in regulating cell apoptosis under the control of the
circadian gene hClock.
In conclusion, the results of the present study
demonstrated that overexpression of the circadian gene hClock had
an enhancing role in CRC progression and may inhibit apoptosis in
CRC cells in vitro and in vivo. In addition,
silencing hClock in CRC cells had an opposite effect. These results
indicated that hClock was functionally important in regulating the
progression of CRC and may serve as a novel target for CRC
therapy.
Acknowledgments
This work was supported by grants from the National
Natural Science Foundation of China (nos. 81070234 and
81000355).
References
|
1
|
Dibner C, Schibler U and Albrecht U: The
mammalian circadian timing system: organization and coordination of
central and peripheral clocks. Ann Rev Physiol. 72:517–549. 2010.
View Article : Google Scholar
|
|
2
|
Chen-Goodspeed M and Lee CC: Tumor
suppression and circadian function. J Biol Rhythms. 22:291–298.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Ko CH and Takahashi JS: Molecular
components of the mammalian circadian clock. Hum Mol Genet. 15(Spec
No.2): R271–R277. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Doi M, Hirayama J and Sassone-Corsi P:
Circadian regulator CLOCK is a histone acetltransferase. Cell.
125:497–508. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Rey G and Reddy AB: Connecting cellular
metabolism to circadian clocks. Trends Cell Biol. 23:234–241. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Savvidis C and Koutsilieris M: Circadian
rhythm disruption in cancer biology. Mol Med. 18:1249–1260. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Hoffman AE, Yi CH, Zheng T, et al: CLOCK
in breast tumorigenesis: genetic, epigenetic and transcriptional
profiling analyses. Cancer Res. 70:1459–1468. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Chen ST, Choo KB, Hou MF, et al:
Deregulated expression of the PER1, PER2 and PER3 genes in breast
cancers. Carcinogenesis. 26:1241–1246. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Lin YM, Chang JH, Yeh KT, et al:
Disturbance of circadian gene expression in hepatocellular
carcinoma. Mol Carcinog. 47:925–933. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Mazzoccoli G, Piepoli A, Carella M, et al:
Altered expression of the clock gene machinery in kidney cancer
patients. Biomed Pharmacother. 66:175–179. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Yeh KT, Yang MY, Liu TC, et al: Abnormal
expression of period 1 (PER1) in endometrial carcinoma. J Pathol.
206:111–120. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Chen Z, Liu P, Li C, et al: Deregulated
expression of the clock genes in gliomas. Technol Cancer Res Treat.
12:91–97. 2013.
|
|
13
|
Fu L, Pelicano H, Liu JS, et al: The
circadian gene period2 plays an important role in tumor suppression
and DNA damage response in vivo. Cell. 111:41–50. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Soták M, Polidarová L, Ergang P, et al: An
association between clock genes and clock-controlled cell cycle
genes in murine colorectal tumors. Int J Cancer. 132:1032–1041.
2013. View Article : Google Scholar
|
|
15
|
Jemal A, Bray F, Center MM, et al: Global
cancer statistics. CA Cancer J Clin. 61:69–90. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Fearon ER: Molecular genetics of
colorectal cancer. Annu Rev Pathol. 6:479–507. 2011. View Article : Google Scholar
|
|
17
|
Cancer Genome Altas Network: Comprehensive
molecular characterization of human colon and rectal cancer.
Nature. 487:330–337. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Wang L, Chen B, Wang Y, et al: hClock gene
expression in human colorectal carcinoma. Mol Med Rep. 8:2017–2022.
2013.
|
|
19
|
Oshima T, Takenoshita S, Akaike M, et al:
Expression of circadian genes correlates with liver metastasis and
outcomes in colorectal cancer. Oncol Rep. 25:1439–1446. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Karantanos T, Theodoropoulos G, Gazouli M,
et al: Expression of clock genes in patients with colorectal
cancer. Int J Biol Markers. 28:280–285. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Alhopuro P, Björklund M, Sammalkorpi H, et
al: Mutations in the circadian gene CLOCK in colorectal cancer. Mol
Cancer Res. 8:952–960. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Zhou F, He X, Lui H, et al: Functional
polymorphisms of circadian positive feedback regulation genes and
clinical outcome of Chinese patients with resected colorectal
cancer. Cancer. 118:937–946. 2012. View Article : Google Scholar
|
|
23
|
Storch KF, Lipan O, Leykin I, et al:
Extensive and divergent circadian gene expression in liver and
heart. Nature. 417:78–83. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Bechtold DA, Gibbs JE and Loudon AS:
Circadian dysfunction in disease. Trends Pharmacol Sci. 31:191–198.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Levi F and Schibler U: Circadian rhythms:
mechanisms and therapeutic implications. Annu Rev Pharmacol
Toxicol. 47:593–628. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Sahar S and Sassone-Corsi P: Circadian
clock and breast cancer: a molecular link. Cell Cycle. 6:1329–1331.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Spengler ML, Kuropatwinshki KK, Comas M,
et al: Core circadian protein CLOCK is a positive regulator of
NF-κB-mediated transcription. PNAS. 109:E2457–E2465. 2012.
View Article : Google Scholar
|
|
28
|
Fu L and Lee CC: The circadian clock:
pacemaker and tumour suppressor. Nat Rev Cancer. 3:350–361. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Ludwig JA and Weinstein JN: Biomarkers in
cancer staging, prognosis and treatment selection. Nat Rev Cancer.
5:845–856. 2005. View
Article : Google Scholar : PubMed/NCBI
|
|
30
|
Gery S and Koeffler HP: The role of
circadian regulation in cancer. Cold Spring Harb Symp Quant Biol.
72:459–464. 2007. View Article : Google Scholar
|
|
31
|
Vivanco I and Sawyers CL: The
phosphatidylinositol 3-Kinase AKT pathway in human cancer. Nat Rev
Cancer. 2:489–501. 2002. View
Article : Google Scholar : PubMed/NCBI
|
|
32
|
Adams JM and Cory S: Life-or-death
decisions by the Bcl-2 protein family. Trends Biochem Sci.
26:61–66. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Kiefer MC, Brauer MJ, Powers VC, et al:
Modulation of apoptosis by the widely distributed Bcl-2 homologue
Bak. Nature. 374:736–739. 1995. View
Article : Google Scholar : PubMed/NCBI
|
|
34
|
Bellacosa A, Chan TO, Ahmed NN, et al: Akt
activation by growth factors is a multiple-step process: the role
of the PH domain. Oncogene. 17:313–325. 1998. View Article : Google Scholar : PubMed/NCBI
|