Introduction
Exosomes are small (50–100 nm) membrane vesicles of
endocytic origin that are released into the extracellular
environment upon fusion of multivesicular bodies (100 nm–1
μm) with the plasma membrane (1,2).
Various cells have been shown to release exosomes, including
reticulocytes (3), as well as
dendritic (4), B- (5) and T- (6), mast (7), epithelial (8) and tumor cells (9). In addition, exosomes have been
isolated and characterized from various biological fluids, such as
urine (10), serum (11) and human breast milk (12). The biological functions of exosomes
depend mainly on their surface proteins and the cell types from
which they are derived. Although the exosome function remains
poorly understood, previous studies have suggested that exosomes
may be associated with a wide range of biological processes, such
as intercellular communication (13) and the development of immune
tolerance (14). Exosomes have
been shown to target specific recipient cells, exchanging proteins
and lipids between the cells and delivering messenger (m)RNAs and
microRNAs (7). Therefore, the
exosome-mediated transfer of mRNAs and microRNAs may be a potential
mechanism for the exchange of genetic material between cells.
Multiorgan tumorigenesis has low incidence; however,
it is associated with poor clinical prognosis. Previous studies
have hypothesized that environmental or chemical factors contribute
towards multiorgan tumorigenesis (15–17).
However, the precise underlying mechanisms of the communication of
multiorgan tumors with host stromal cells and tumor cells remain to
be elucidated.
The present study evaluated the intercellular
communication through exosomes in multiorgan tumorigenesis, using
LoVo human colon cancer and U-87 MG human glioblastoma cells. A
U-87-green fluorescent protein (GFP) cell line was initially
established, which expressed GFP in U-87 MG cells. U-87-GFP
cell-derived exosomes were hypothesized to transfer GFP mRNAs,
which would then be translated into proteins in the recipient LoVo
cells. The present study aimed to reveal the function of exosomes
in intercellular communication during multiorgan tumorigenesis. The
cellular uptake pathway of U-87-GFP cell-derived exosomes into LoVo
cells was also investigated in order to reveal how exosomes are
associated with intercellular communication between cancer cells,
which may have potential therapeutic applications.
Materials and methods
Cell lines
The LoVo human colon cancer and U-87 MG
ATCC® HTB-14™ human glioma cell lines were obtained from
the American Type Culture Collection (Manassas, VA, USA). U-87-GFP
cells expressing GFP were generated by stable infection of U-87 MG
cells with a lentiviral vector expressing GFP (pHAGE-CMV-MCS-IZs
green vector; provided by Professor Chun Lu, Nanjing Medical
University, Nanjing, China). The LoVo cells were grown in RPMI-1640
medium supplemented with 10% fetal bovine serum, 100 U/ml
penicillin and 100 μg/ml streptomycin (all Life
Technologies, Grand Island, NY, USA). The U-87-GFP cells were grown
in Dulbecco’s modified Eagle’s medium supplemented with 10% fetal
bovine serum, 100 U/ml penicillin and 100 μg/ml streptomycin
(Life Technologies). All the cells were maintained at 37°C in a
humidified atmosphere containing 5% CO2.
Isolation of exosomes
The U-87-GFP cells were cultured in
microvesicle-free medium generated by ultracentrifugation in an
SW41 rotor (Beckman Instruments, Fullerton, CA, USA) at 100,000 × g
for 1 h and the culture medium (2×107 cells) was
collected following a 48 h incubation. The U-87-GFP cell-derived
exosomes were purified by a series of differential centrifugations,
as previously described (18).
Briefly, the collected culture supernatants were centrifuged for 10
min at 300 × g in order to eliminate cell contamination.
Subsequently, the supernatants were further centrifuged at 1,200 ×
g for 20 min and then at 10,000 × g for 30 min. The supernatant
from the final spin was ultracentrifuged (Beckman SW-41; Beckman
Coulter, Inc., Brea, CA, USA) at 100,000 × g for 2 h in order to
pellet the exosomes. Next, the exosome pellet was resuspended in
200 μl phosphate-buffered saline (PBS) and filtered through
a 0.22 μm filter (EMD Millipore, Bedford, MA, USA). The
protein concentration of exosomes was assessed using a Quick Start™
Bradford Protein assay (Bio-Rad Laboratories, Inc., Hercules, CA,
USA).
Characteristics of exosomes
In order to determine the exosome morphology, the
U-87-GFP cell-derived exosome dispersion (10 μl) was loaded
onto a Formvar/carbon coated grid (Agar Scientific, Stansted, UK)
for 1 min, negatively stained with 10 μl neutral 1% aqueous
phosphotungstic acid (Sigma-Aldrich, St. Louis, MO, USA) for 1 min
and visualized using an electron microscope (XL-30 ESEM FEG; FEI,
Hillsboro, OR, USA). The mean particle size (number-weighted size
distribution) and ζ-potential of the U-87-GFP cell-derived exosomes
were measured by dynamic light scattering using a Zetasizer 3000
(Malvern Instruments, Ltd., Malvern, UK). Briefly, 10 μl of
the exosomes was dispersed into 1 ml deionized water. The resultant
dispersion was analyzed in triplicate and each replicate was
analyzed eleven times in order to yield the average particle size.
Similarly, the ζ-potential of each dispersion sample was
investigated in triplicate and each replicate was analyzed eleven
times to obtain the average ζ-potential.
Quantitative polymerase chain reaction
(qPCR) and reverse transcription-PCR (RT-PCR)
The cells or exosomes were harvested and total RNA
was extracted using the TRIzol® reagent (Invitrogen Life
Technologies, Carlsbad, CA, USA). In total, 2 μg total RNA
was isolated, according to the manufacturer’s instructions, and
cDNA was synthesized using a conventional method
(SuperScript® III; Life Technologies). qPCR was
performed using SYBR® Green PCR Master mix (Applied
Biosystems Life Technologies, Foster City, CA, USA) on an ABI 7300
Prism real-time PCR (Applied Biosystems Life Technologies)
instrument. The following primers were used for qPCR amplification
in the present study: GFP sense, 5′-CCA CTG CCA TTC TCC GAA GA-3′,
and antisense, 5′-TGC TGG ATG AAG TGC CTG TC-3′; and β-actin sense,
5′-AGG GAA ATC GTG CGT GAC-3′, and antisense, 5′-CGC TCA TTG CCG
ATA GTG-3′. The cycle threshold (Ct) values were measured and the
2−ΔCt method (where ΔCt = target gene Ct - β-actin Ct)
was used for quantification. The primer sequences used for RT-PCR
amplification in the present study were as follows: GFP sense,
5′-AAG TTC ATC TGC ACC ACCG-3′, and anti-sense, 5′-TCC TTG AAG AAG
ATG GTG CG-3′; and β-actin sense, 5-TGA CGG GGT CAC CCA CAC TGT GCC
CAT CTA-3′, and antisense, 5′-CTA GAA GCA TTT GCG GTG GAC GAT GGA
GGG-3′. All primers were synthesized by GenePharma Co., Ltd
(Shanghai, China). In total, 30 cycles of PCR were performed and
the annealing temperature was set at 56°C. The PCR cycle consisted
of an initial step of 95°C for 3 min, followed by 30 cycles of 95°C
for 15 sec and 56°C for 30 sec, followed by an elongation step at
72°C for 7 min. The PCR products were confirmed by agarose gel
electrophoresis, followed by ethidium bromide staining
(Sigma-Aldrich).
Transfer experiments
The LoVo cells (5×104 cells) were seeded
on 24-well plates prior to incubation. The exosomes (20
μg/ml) were added to the LoVo cells at a ratio of 8:1
between the donor and recipient cells, according to a previously
described method (7). Next, the
cells were analyzed by fluorescence microscopy (CKX41-URFLT50;
Olympus Corporation, Tokyo, Japan), following 0, 24 and 48 h of
incubation.
Cellular uptake of exosomes
U-87-GFP cell-derived exosomes were initially
labeled with the PKH26 fluorescent dye (Sigma-Aldrich), according
to the manufacturer’s instruction. Cellular uptake of U-87-GFP
cell-derived exosomes by LoVo cells was conducted by incubating the
PKH26-labeled exosomes with the LoVo cells for 12 h, in the
presence of 5 μM cytochalasin D, 500 nM bafilomycin A1 or 50
μm indomethacin (Sigma-Aldrich). Following the 12-h
incubation, the LoVo cells were washed three times with PBS and
detached by trypsinization (Life Technologies). The cells were then
centrifuged at 300 × g for 5 min and resuspended in 0.4% (w/v)
trypan blue (Sigma-Aldrich) in Hank’s balanced salt solution (Life
Technologies) to quench the extracellular fluorescence. The treated
samples were washed and analyzed using a BD FACSCanto™ system (BD
Biosciences, Franklin Lakes, NJ, USA) with 5,000–10,000 cells
measured for each sample.
Statistical analyses
Statistical analyses were performed by Student’s
t-test using SPSS version 16.0 software (SPSS Inc., Chicago, IL,
USA). P<0.05 was considered to indicate a statistically
significant difference.
Results
Isolation and identification of tumor
cell-derived exosomes
Previous studies have hypothesized that
environmental or chemical factors may contribute to multiorgan
tumorigenesis (17,19). However, the precise mechanisms
underlying the communication of multiorgan tumors with host cells
and tumor cells remain to be elucidated.
In the present study, exosomes derived from tumor
cells were hypothesized to promote multiorgan tumorigenesis. In
order to assess this hypothesis, the most aggressive tumor models
were selected, including U-87 MG human glioblastoma and LoVo colon
cancer cell lines. Initially, the ability of these tumor cells to
produce exosomes was investigated. Next, the U-87-GFP human glioma
cell line, showing a stable expression of GFP, was generated.
U-87-GFP cell-derived exosomes were isolated from these cells
through a series of differential microfiltration and
ultracentrifugation steps, modified from a previous study (20). The exosomes were collected and
characterized using electron microscopy and dynamic light
scattering techniques. The exosomes were constitutively derived
from tumor cells (with a density ranging between 0.75 and 1.10
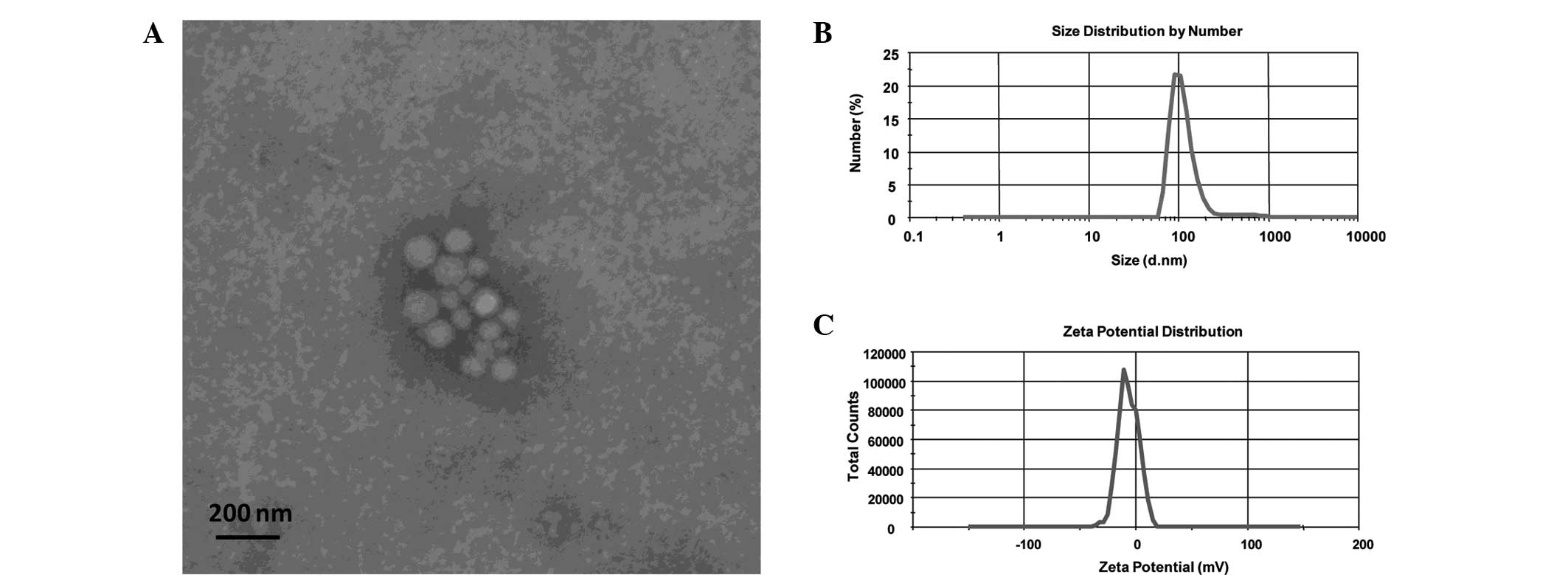
g/ml). Electron micrographs revealed that the exosomes appeared to
be round and well-delimited vesicles, with a size of ~100 nm
(Fig. 1A), which was similar to
previously described exosomes (7,20–22).
Following redispersion of the U-87-GFP cell-derived exosomes in
deionized water, the average size and ζ-potential were found to be
140 nm and −6.23 mV, respectively (Fig. 1B and C). The exosome size observed
using dynamic light scattering was comparable with the results
obtained using electron microscopy.
Intercellular transfer of GFP mRNAs by
tumor cell-derived exosomes
The ability of tumor cell-derived exosomes to
deliver mRNAs into tumors from different organs/sites was
investigated as a potential pathway of intercellular communication
during multiorgan tumorigenesis. To determine whether the exosomes
were capable of transferring exogenous GFP mRNAs, LoVo cells were
incubated with exosomes derived from U-87-GFP cells, which showed a
stable expression of GFP. Following incubation with the exosomes
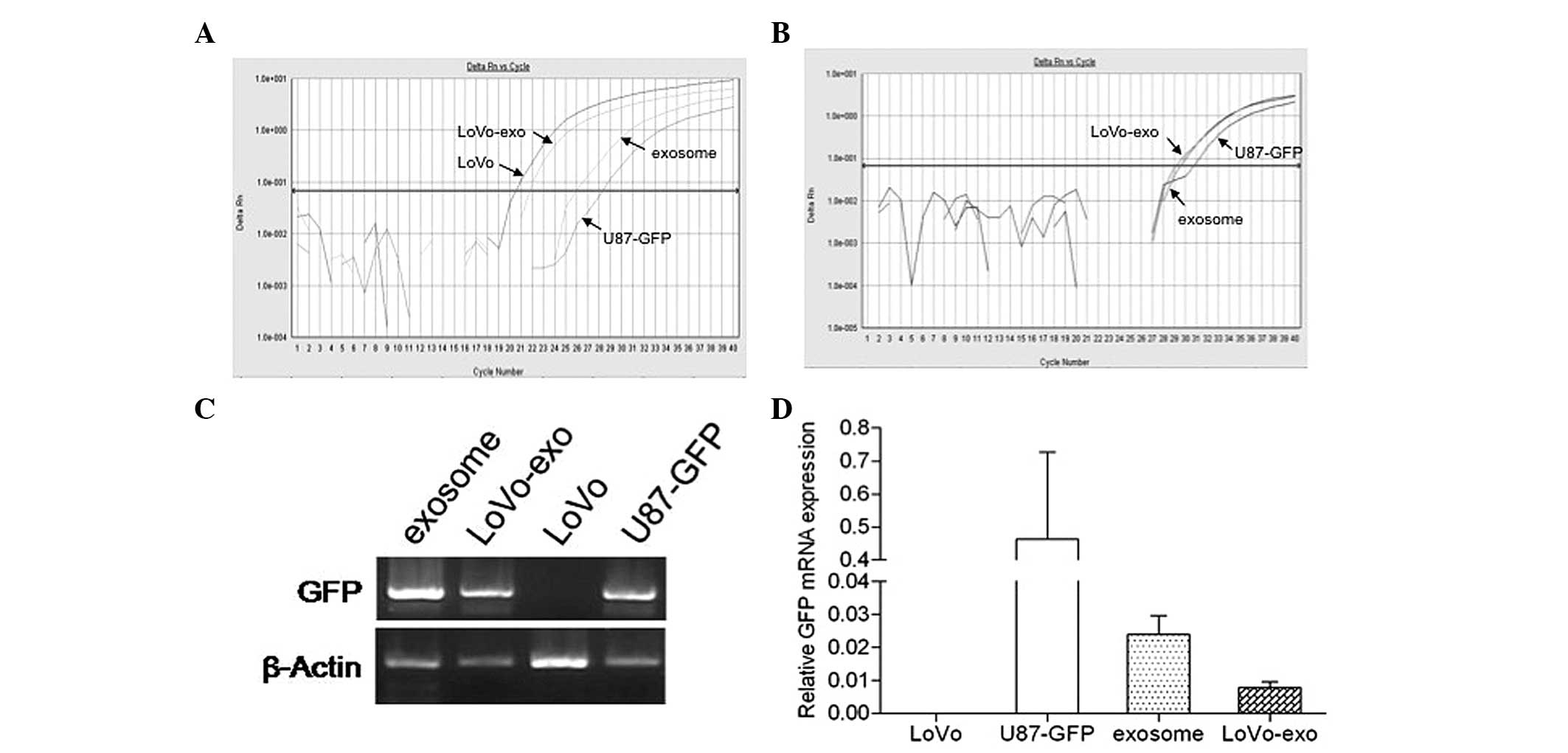
for 24 h, GFP mRNA expression was detected using qPCR and was
further verified by agarose gel electrophoresis. Amplification of
the housekeeping gene β-actin mRNA expression was detected in the
LoVo and U-87-GFP cells, U-87-GFP cell-derived exosomes and LoVo
cells incubated with U-87-GFP cell-derived exosomes after 21, 28.2,
25.9 and 22.0 cycles, respectively (Fig. 2A). Notably, no amplification of GFP
mRNA expression was detected in the LoVo cells. By contrast,
amplification of GFP mRNA was observed in the U-87-GFP cells,
U-87-GFP cell-derived exosomes and LoVo cells incubated with
U-87-GFP cell-derived exosomes after 31.8, 29.5 and 29.1 cycles,
respectively (Fig. 2B). The mRNA
expression levels of GFP were highest in the U-87-GFP cells,
followed by the U-87-GFP cell-derived exosomes and LoVo cells
incubated with U-87-GFP cell-derived exosomes (Fig. 2C). The results obtained from the
agarose gel electrophoresis further supported the observation that
GFP mRNA was detected in LoVo cells following incubation with
U-87-GFP cell-derived exosomes. These data indicated that GFP mRNA
was transferred from the U-87-GFP cells to the LoVo cells by
U-87-GFP cell-derived exosomes.
GFP mRNAs transferred by exosomes are
translated into functional proteins
To determine whether the mRNA transferred by the
U-87-GFP cell-derived exosomes was functional, GFP mRNA delivered
by exosomes were investigated to establish whether they can be
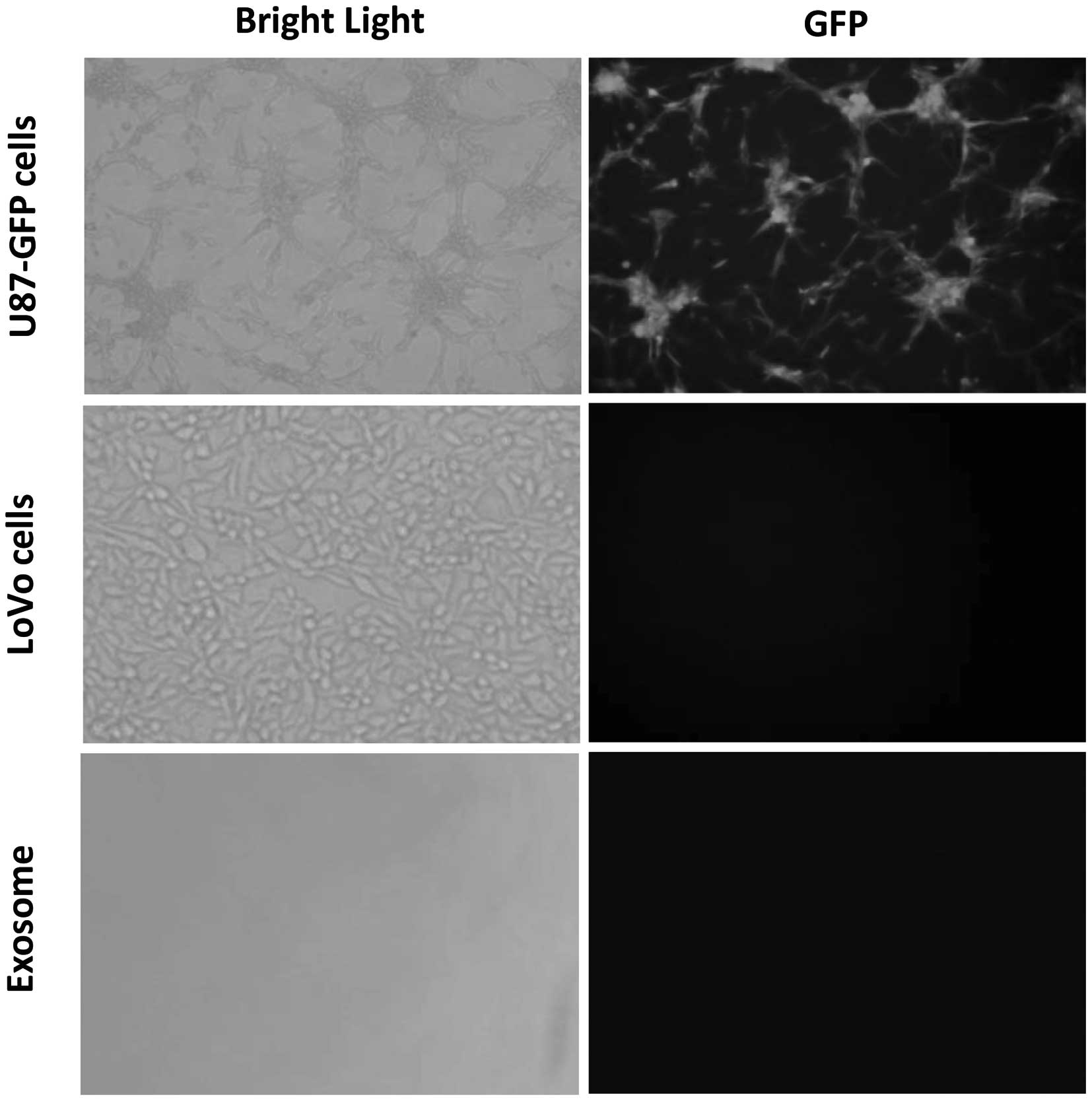
translated into proteins in the LoVo cells. The exosomes were
analyzed by immunofluorescence microscopy (Fig. 3). GFP was detected in the U-87-GFP
cells; however, it was not detected in U87-GFP cell-derived
exosomes or LoVo cells indicating that they did not contain GFP.
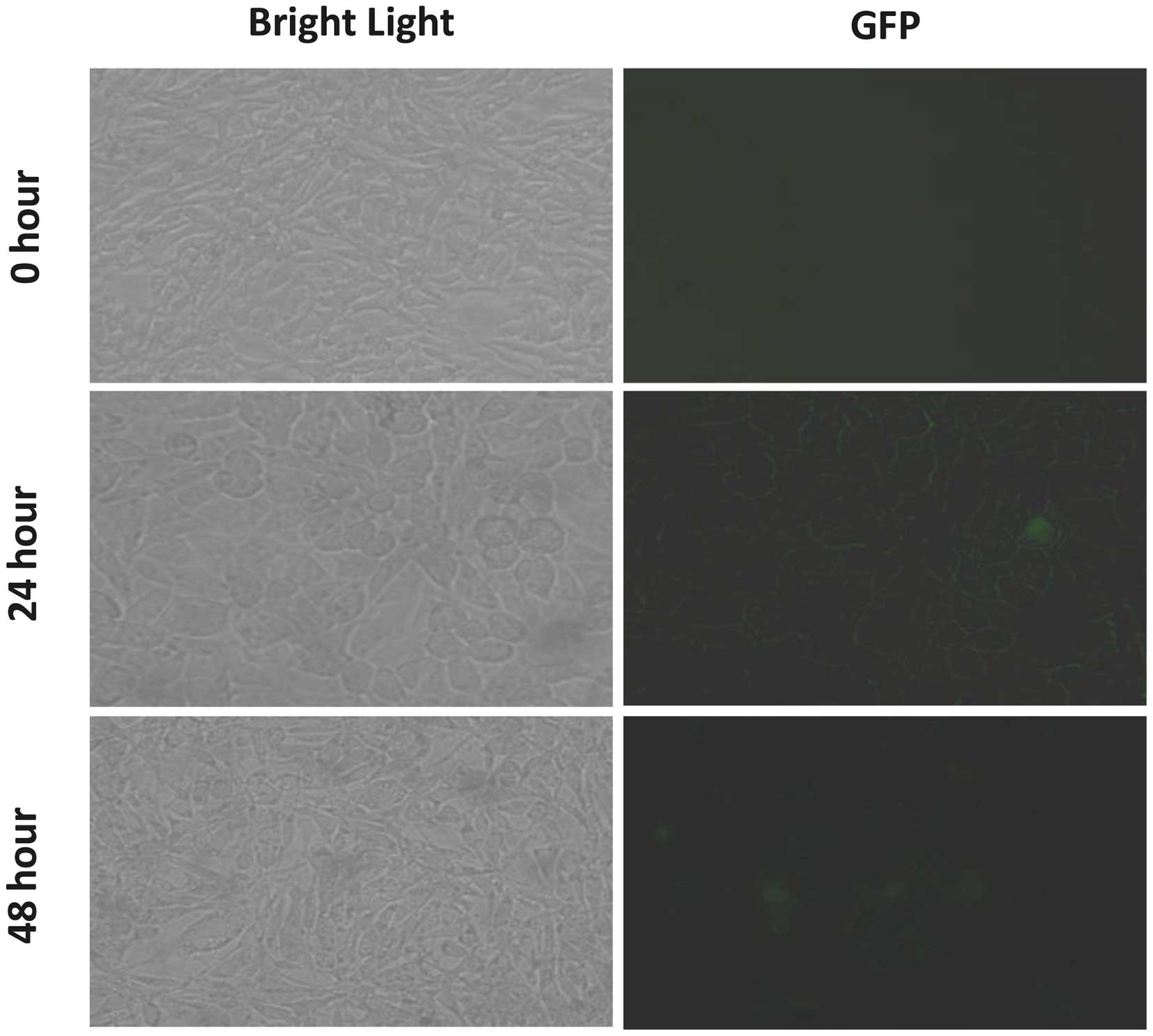
Subsequently, LoVo cells were incubated with the exosomes derived
from the U-87-GFP cells, and these cells were visualized using
immunofluorescence microscopy at the following time points: 0, 24
and 48 h. A marked increase in GFP fluorescence was detected in the
cytoplasm and near the plasma membrane at 24 h (Fig. 4). Thus, the GFP mRNA delivered by
the exosomes into the LoVo cells was successfully translated into
GFP.
Cellular uptake mechanism of
exosomes
To clearly identify the role of specific endocytic
pathways involved in U-87-GFP cell-derived exosome cellular
internalization, LoVo cells were treated with known endocytic
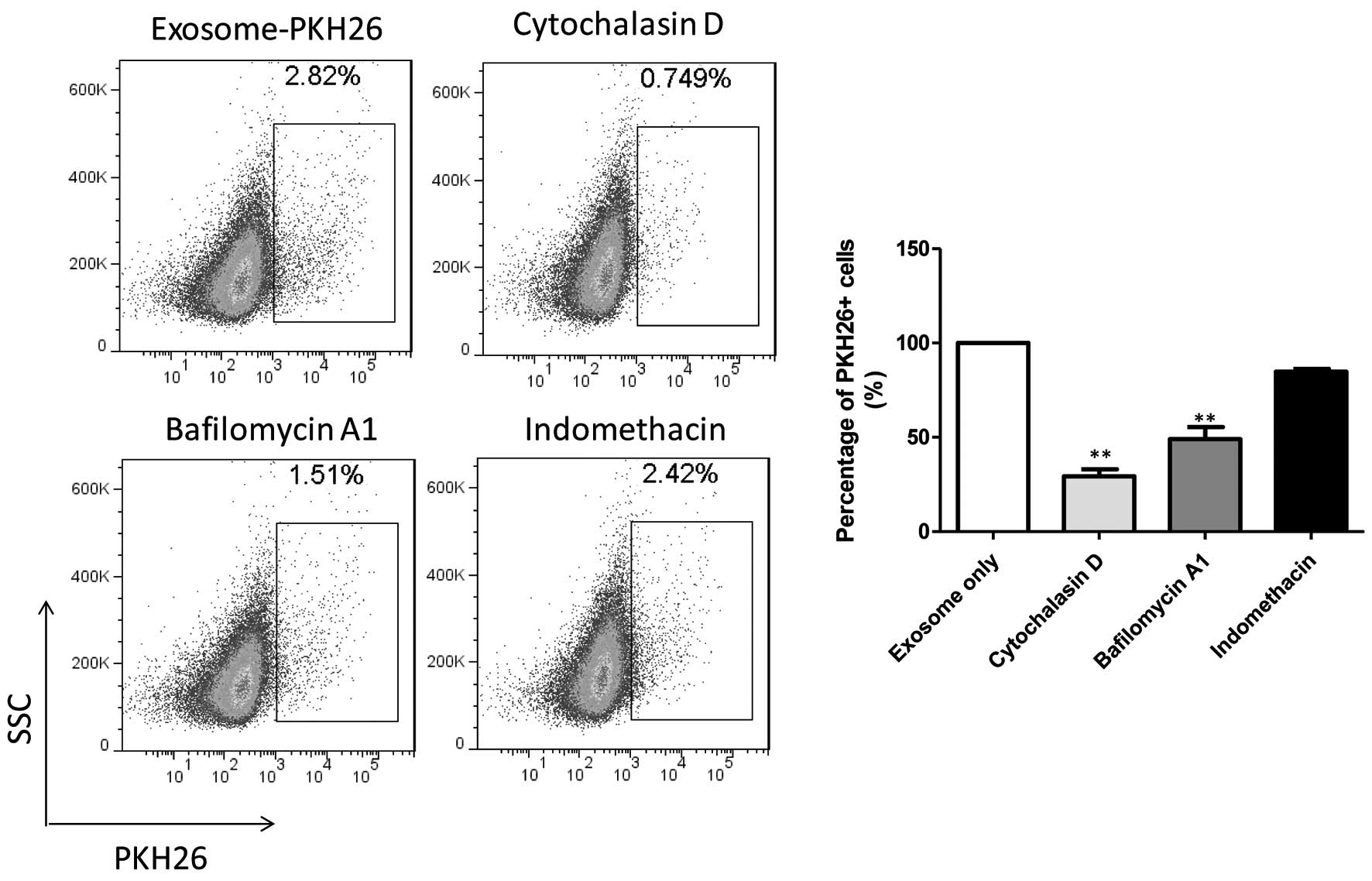
inhibitors (Fig. 5).
Internalization of U-87-GFP cell-derived exosomes was significantly
decreased in the presence of cytochalasin D (29.25%), an inhibitor
of actin polymerization. Clathrin- and caveolae-mediated pathways
have been previously shown to require actin polymerization,
suggesting that the cellular uptake of U-87-GFP cell-derived
exosomes is predominantly achieved through these pathways (23). Furthermore, internalization of the
U-87-GFP cell-derived exosomes into LoVo cells was markedly reduced
in the presence of bafilomycin A1, a vacuolar proton ATPase
inhibitor (49.11%). However, no inhibitory effect was observed on
the cellular uptake of U-87-GFP cell-derived exosomes by LoVo cells
in the presence of indomethacin, a caveolae-mediated endocytic
inhibitor that blocks the cyclooxygenase pathway (23,24).
These results indicated that exosomes derived from U-87-GFP cells
were internalized into LoVo cells predominantly through
clathrin-mediated endocytosis.
Discussion
The present study identified a pathway of mRNA
transport between different tumor cells by tumor cell-derived
exosomes. The tumor cell-derived exosomes were shown to be capable
of transferring mRNAs to other types of tumor cells through
clathrin-mediated endocytosis. Furthermore, GFP mRNA delivered by
exosomes may be translated into protein in the recipient cells.
These results indicate that tumor cell-derived exosomes may be
capable of delivering mRNAs to recipient cells, which may be a
potential pathway of intercellular communication during multiorgan
tumorigenesis.
Previous studies have hypothesized that tumor
cell-derived microvesicles contain mRNAs that may be transferred
(21); however, little is
currently known about the precise mechanisms underlying the
transfer of RNAs between different tumor types by tumor
cell-derived exosomes. A previous study indicated that mast cells
secrete exosomes that contain mRNAs and microRNAs (7). Based on these previous findings, the
present study hypothesized that exosomal RNAs may be transported
between tumors from different organs/sites by tumor cell-derived
exosomes. The results of the present study demonstrated that
exosomes were spontaneously and constitutively derived from tumor
cells, which was consistent with the observations of previous
studies (7,20,21,25).
In addition, these tumor cell-derived exosomes were found capable
of transferring GFP mRNA between tumor cells, which may be
translated into functional proteins in the recipient cells.
Recipient cells were found to uptake exosomes through
clathrin-mediated endocytosis. The exosome uptake pathway may have
a therapeutic application in multiorgan tumorigenesis.
A previous study has demonstrated that embryonic
stem cell microvesicles may be engineered to transfer
exogenously-expressed mRNAs and microRNAs (26). In addition, the authors
demonstrated that GFP was readily detected in embryonic stem cell
microvesicles, where it was less abundant than in embryonic stem
cells. By contrast, in the present study, GFP was not detected in
the U-87-GFP cell-derived exosomes, indicating that protein
transfer by exosomes occurs in a cell type-dependent manner. Simons
et al (27) initially
proposed the term microvesicles or microparticles for vesicles of
heterogeneous size and shape (100 nm–1 μm) and the term
exosomes for vesicles with a size of 40–100 nm that carry the
typical exosomal marker proteins. The constituents of microvesicles
or exosomes require further investigation, in order to elucidate
whether they contain the same constituents. Notably, the results of
the present study demonstrated that translation of the mRNA
delivered by exosomes was initiated at 24 h and decreased at 48 h.
Therefore, certain mRNAs that are transferred into recipient cells
may fall into decay. Future studies should be conducted to examine
the mechanisms underlying the regulation of exosome-transferred
mRNAs by recipient cells. Based on the present findings,
tumor-derived exosomes were hypothesized to deliver genetic
information to multiorgan tumors during intercellular
communication.
The tumor microenvironment is known to be involved
in cancer development and progression. Increasing evidence has
indicated that microvesicles/exosomes may be important constituents
of the tumor microenvironment, indicating a pivotal role of
microvesicles/exosomes in tumor invasion and metastasis,
angiogenesis and immune system suppression (9,28,29).
Furthermore, previous studies have identified
microvesicles/exosomes as major mediators of intercellular
communication (25,30). Attention is currently focused on
the transfer of nucleic acids by microvesicles/exosomes in
intercellular communication. For instance, embryonic stem
cell-derived microvesicles have been shown to reprogram
hematopoietic progenitors by horizontal transfer of pluripotent and
early hematopoietic stem cell mRNA (31). In addition, numerous studies have
indicated that microvesicles derived from tumor cells may be
involved in the transfer of mRNAs and proteins to
antigen-presenting cells (21), as
well as oncogenetic receptors among tumor cells (20). Further evidence regarding nucleic
acid transfer was provided by a study identifying that mast
cell-derived exosomes contain mRNAs and microRNAs, which can be
delivered to other cells; these RNAs were termed as ‘exosomal
shuttle RNAs’ (7). In later
studies, this novel mechanism of nucleic acid transfer has been
observed in tumor cells. Skog et al (18) suggested that glioblastoma
microvesicles promote tumor growth through the transportation of
RNAs and proteins. Furthermore, Hunter et al (32) identified 33 significantly expressed
microRNAs in plasma microvesicles and predicted that these
microRNAs may be important factors in the regulation of immune
responses and hematopoiesis. In addition, Taylor et al
(33) demonstrated that exosomal
microRNA profiles from ovarian cancer patients were significantly
distinct from the profiles observed in benign diseases. Based on
these previous findings, the present study postulated that nucleic
acids transferred by tumor cell-derived exosomes, particularly
mRNAs and microRNAs, may enable malignant cells to influence the
surrounding nonmalignant cells and microenvironment, assisting the
tumor in invasion, metastasis and angiogenesis, as well as immune
suppression. Furthermore, tumor cells from various organs or sites
may communicate by transferring genetic information through tumor
cell-derived exosomes, thereby promoting tumor progression during
multi-organ tumorigenesis. Inhibition of the uptake pathways of
tumor exosomes may have potential therapeutic applications in
multiorgan tumorigenesis.
In conclusion, the present study demonstrated that
U-87-GFP cell-derived exosomes were capable of transferring mRNAs
to LoVo cells, through clathrin-mediated endocytic uptake. The mRNA
delivered by the exosomes may then be translated into functional
proteins in the recipient LoVo cells. These results indicate that
tumor cell-derived exosomes may represent vesicular carriers that
regulate gene expression, which may provide a pathway of
intercellular communication between tumor cells in the tumor
microenvironment during multiorgan tumorigenesis.
References
|
1
|
Zitvogel L, Regnault A, Lozier A, et al:
Eradication of established murine tumors using a novel cell-free
vaccine: dendritic cell derived exosomes. Nat Med. 4:594–600. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Théry C, Zitvogel L and Amigorena S:
Exosomes: composition, biogenesis and function. Nat Rev Immunol.
2:569–579. 2002.PubMed/NCBI
|
|
3
|
Johnstone RM, Bianchini A and Teng K:
Reticulocyte maturation and exosome release: transferrin receptor
containing exosomes shows multiple plasma membrane functions.
Blood. 74:1844–1851. 1989.PubMed/NCBI
|
|
4
|
Théry C, Boussac M, Véron P, et al:
Proteomic analysis of dendritic cell-derived exosomes: a secreted
subcellular compartment distinct from apoptotic vesicles. J
Immunol. 166:7309–7318. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Clayton A, Turkes A, Dewitt S, Steadman R,
Mason MD and Hallett MB: Adhesion and signaling by B cell-derived
exosomes: the role of integrins. FASEB J. 18:977–979.
2004.PubMed/NCBI
|
|
6
|
Blanchard N, Lankar D, Faure F, et al: TCR
activation of human T cells induces the production of exosomes
bearing the TCR/CD3/zeta complex. J Immunol. 168:3235–3241. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Valadi H, Ekström K, Bossios A, Sjöstrand
M, Lee JJ and Lötvall JO: Exosome-mediated transfer of mRNAs and
microRNAs is a novel mechanism of genetic exchange between cells.
Nat Cell Biol. 9:654–659. 2007. View
Article : Google Scholar : PubMed/NCBI
|
|
8
|
Van Niel G, Mallegol J, Bevilacqua C, et
al: Intestinal epithelial exosomes carry MHC class II/peptides able
to inform the immune system in mice. Gut. 52:1690–1697. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Wolfers J, Lozier A, Raposo G, et al:
Tumor-derived exosomes are a source of shared tumor rejection
antigens for CTL cross-priming. Nat Med. 7:297–303. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Nilsson J, Skog J, Nordstrand A, et al:
Prostate cancer-derived urine exosomes: a novel approach to
biomarkers for prostate cancer. Br J Cancer. 100:1603–1607. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Caby MP, Lankar D, Vincendeau-Scherrer C,
Raposo G and Bonnerot C: Exosomal-like vesicles are present in
human blood plasma. Int Immunol. 17:879–887. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Admyre C, Johansson SM, Qazi KR, et al:
Exosomes with immune modulatory features are present in human
breast milk. J Immunol. 179:1969–1978. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
André F, Chaput N, Schartz NE, et al:
Exosomes as potent cell-free peptide-based vaccine. I. Dendritic
cell-derived exosomes transfer functional MHC class I/peptide
complexes to dendritic cells. J Immunol. 172:2126–2136. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Karlsson M, Lundin S, Dahlgren U, Kahu H,
Pettersson I and Telemo E: ‘Tolerosomes’ are produced by intestinal
epithelial cells. Eur J Immunol. 31:2892–2900. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Stoner G, Casto B, Ralston S, Roebuck B,
Pereira C and Bailey G: Development of a multi-organ rat model for
evaluating chemopreventive agents: efficacy of indole-3-carbinol.
Carcinogenesis. 23:265–272. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Pazzaglia S: Ptc1 heterozygous knockout
mice as a model of multi-organ tumorigenesis. Cancer Lett.
234:124–134. 2006. View Article : Google Scholar
|
|
17
|
Yamagishi M, Natsume M, Osakabe N, et al:
Chemoprevention of lung carcinogenesis by cacao liquor
proanthocyanidins in a male rat multi-organ carcinogenesis model.
Cancer Lett. 191:49–57. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Skog J, Würdinger T, van Rijn S, et al:
Glioblastoma microvesicles transport RNA and proteins that promote
tumour growth and provide diagnostic biomarkers. Nat Cell Biol.
10:1470–1476. 2008. View
Article : Google Scholar : PubMed/NCBI
|
|
19
|
Fero ML, Rivkin M, Tasch M, et al: A
Syndrome of multiorgan hyperplasia with features of gigantism,
tumorigenesis, and female sterility in p27(Kip1)-deficient mice.
Cell. 85:733–744. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Al-Nedawi K, Meehan B, Micallef J, et al:
Intercellular transfer of the oncogenic receptor EGFRvIII by
microvesicles derived from tumour cells. Nat Cell Biol. 10:619–624.
2008. View
Article : Google Scholar : PubMed/NCBI
|
|
21
|
Baj-Krzyworzeka M, Szatanek R, Weglarczyk
K, et al: Tumour-derived microvesicles carry several surface
determinants and mRNA of tumour cells and transfer some of these
determinants to monocytes. Cancer Immunol Immunother. 55:808–818.
2006. View Article : Google Scholar
|
|
22
|
Martinez MC, Larbret F, Zobairi F, et al:
Transfer of differentiation signal by membrane microvesicles
harboring hedgehog morphogens. Blood. 108:3012–3020. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Gratton SEA, Ropp PA, Pohlhaus PD, et al:
The effect of particle design on cellular internalization pathways.
Proc Natl Acad Sci USA. 105:11613–11618. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Weggen S, Eriksen JL, Das P, et al: A
subset of NSAIDs lower amyloidogenic Abeta42 independently of
cyclooxygenase activity. Nature. 414:212–216. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Ratajczak J, Wysoczynski M, Hayek F,
Janowska-Wieczorek A and Ratajczak MZ: Membrane-derived
microvesicles: important and underappreciated mediators of
cell-to-cell communication. Leukemia. 20:1487–1495. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Yuan A, Farber EL, Rapoport AL, et al:
Transfer of microRNAs by embryonic stem cell microvesicles. PLoS
One. 4:e47222009. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Simons M and Raposo G: Exosomes -
vesicular carriers for intercellular communication. Curr Opin Cell
Biol. 21:575–581. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Janowska-Wieczorek A, Wysoczynski M,
Kijowski J, et al: Microvesicles derived from activated platelets
induce metastasis and angiogenesis in lung cancer. Int J Cancer.
113:752–760. 2005. View Article : Google Scholar
|
|
29
|
Liu C, Yu S, Zinn K, et al: Murine mammary
carcinoma exosomes promote tumor growth by suppression of NK cell
function. J Immunol. 176:1375–1385. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Schorey JS and Bhatnagar S: Exosome
function: from tumor immunology to pathogen biology. Traffic.
9:871–881. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Ratajczak J, Miekus K, Kucia M, et al:
Embryonic stem cell-derived microvesicles reprogram hematopoietic
progenitors: evidence for horizontal transfer of mRNA and protein
delivery. Leukemia. 20:847–856. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Hunter MP, Ismail N, Zhang X, et al:
Detection of microRNA expression in human peripheral blood
microvesicles. PLoS One. 3:e36942008. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Taylor DD and Gercel-Taylor C: MicroRNA
signatures of tumor-derived exosomes as diagnostic biomarkers of
ovarian cancer. Gynecol Oncol. 110:13–21. 2008. View Article : Google Scholar : PubMed/NCBI
|