Introduction
Chordoma is a rare tumor of the bone, known to arise
from notochord remnants and is the most common primary malignant
tumor of the sacrum and the mobile spine. The clinical management
of chordoma is extremely challenging. Surgical resection is the
primary curative strategy for chordoma, however, it is often not
possible to perform adequately due to the anatomical location of
the tumor (1–3). At present, there are no effective
drug treatments for patients with chordoma, although radiation has
been used as an adjuvant if complete resection is not possible, as
well as when non-contaminated surgical margins are achieved
(4,5).
A microRNA (miRNA) is a type of short, non-coding
RNA that suppresses the expression of protein coding genes by
partial complementary binding, particularly to the 3′-untranslated
regions (3′-UTRs) of messenger RNAs (mRNAs). Alterations in the
expression of miRNAs are associated with the initiation,
progression and metastasis of human cancer and it is hypothesized
that miRNAs function as tumor suppressors and oncogenes in cancer
development (6,7). Previous studies have revealed that
disturbed expression of microRNA (miR)-1 and miR-31 may be involved
in chordoma pathogenesis, however, the mechanisms remain to be
elucidated (8–10).
RNA editing is a widespread post-transcriptional
process contributing to cellular transcriptome diversity in
eukaryotes. The most well-characterized type of RNA editing
identified in mammals converts cytosine to uracil and adenosine to
inosine (A-to-I). In humans, the most frequent type of editing is
the conversion of A-to-I, which is catalyzed by the double-stranded
RNA specific adenosine deaminase acting on RNA (ADAR) family of
proteins. Accumulating evidence has indicated that disturbed RNA
editing may have a role in the pathogenesis of numerous types of
cancers, including prostate (11),
lung (12), liver (13) and malignant gliomas (14,15).
Several studies have demonstrated that altered miRNA precursor
editing is able to modulate miRNA processing and expression and is
associated with cancer pathogenesis (15,16).
In the present study, in order to investigate the
association between miRNA editing and chordoma, the sequences of
miRNA precursors were compared with those of their coding regions
in chordoma tissues. In addition, the expression levels of ADAR1
and ADAR1 were determined. Therefore, the present study aimed to
reveal a novel association between A-to-I RNA editing and
chordoma.
Materials and methods
Samples
Skull-base chordoma tissues and nucleus pulposus
tissues were obtained from the Department of Spinal Surgery, The
Second Xiangya Hospital of Central South University (Changsha,
China) with approval from the ethics committee of The Second
Xiangya Hospital of Central South University. Histopathological
confirmation and grading was performed by pathologists at The
Second Xiangya Hospital of Central South University.
Total RNA isolation
Total RNA from eight chordoma tissues and eight
nucleus pulposus tissues was isolated with TRIzol reagent according
to the manufacturer’s instructions (Invitrogen Life Technologies,
Carlsbad, CA, USA).
Cell culture
Human embryonic kidney (HEK)293T cells (China
Infrastructure of Cell Line Resources, Beijing, China) were
cultured in Dulbecco’s modified Eagle’s medium containing 10% fetal
bovine serum, 100 IU/ml penicillin and 10 mg/ml streptomycin, which
were all purchased from HyClone Laboratories, Inc. (Logan, UT,
USA). All cells were maintained at 37°C under an atmosphere of 5%
CO2.
miRNA reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
RT-qPCR analysis was used to determine the relative
expression level of candidate miRNAs. The expression level of
miRNAs was detected by TaqMan miRNA RT-qPCR. Single-stranded cDNA
was synthesized using a TaqMan MicroRNA Reverse Transcription kit
(Applied Biosystems, Foster City, CA, USA) and then amplified using
TaqMan Universal PCR Master mix (Applied Biosystems) together with
miRNA-specific TaqMan MGB probes (Applied Biosystems). miR-RNU48,
one of the most highly abundant and stably expressed miRs in human
tissues, was used as an endogenous control. Each sample in each
group was measured in triplicate and the experiment was repeated at
least three times for the detection of miRNAs.
DNA collection and genotyping
DNA from tissue samples was isolated using a TIANamp
Genomic DNA kit (Tiangen Biotech (Beijing) Co., Ltd., Beijing,
China). DNA specimens were amplified using standard PCR protocols.
A total of 322 bp of the pri-miR-125a coding region and 315 bp of
the pri-miR-10a coding region were obtained. The PCR products were
sequenced in the forward direction using the ABI 3730xl sequencing
platform (Applied Biosystems). The sequencing results were analyzed
using DNAMAN 7.0 software (Lynnon Corporation, Quebec, Canada) and
Chromas Lite 2.22 software (Technelysium Pty Ltd., Tewantin, QLD,
Australia).
3′-UTR luciferase reporter assays
To generate a 3′-UTR luciferase reporter, the
full length 3′-UTR from ERBB2 and homeobox A1 (HOXA1) were
cloned downstream of the firefly luciferase gene into the
pGL3-control vector (Promega Corporation, Madison, WI, USA). The
miR-375 mimic and miR-375 inhibitor were synthesized by GenePharma
Co., Ltd. (Shanghai, China). The Renilla
luciferase-expressing plasmid pRL-TK was co-transfected for data
normalization. For the luciferase reporter assays, HEK293T cells
were seeded in 48 well plates. Luciferase reporter vectors were
co-transfected with an miRNA mimic or miRNA inhibitor using
lipofectamine 2000 (Invitrogen Life Technologies). After two days,
cells were harvested and assayed with the dual-luciferase assay
(Promega Corporation). Each treatment was performed in triplicate
in three independent experiments. The results are expressed as
relative luciferase activity (Firefly LUC/Renilla LUC).
Western blotting
Protein extracts were boiled in
SDS/β-mercaptoethanol sample buffer (Sigma-Aldrich, St. Louis, MO,
USA) and 20 μg samples were loaded into wells of 8%
polyacrylamide gels. The proteins were separated by electrophoresis
and the proteins in the gels were blotted onto polyvinylidene
difluoride membranes (Amersham Pharmacia Biotech, St. Albans, UK)
by electrophoretic transfer. The membrane was incubated with goat
anti-ADAR1 polyclonal antibody (1:1,000; cat. no. sc-19077; Santa
Cruz Biotechnology, Inc., Santa Cruz, CA, USA), mouse anti-GAPDH
monoclonal antibody (1:5,000; cat. no. sc-365062; Santa Cruz
Biotechnology, Inc.) for 1 h at 37°C. The specific protein antibody
complex was detected using horseradish peroxidase-conjugated
polyclonal rabbit anti-goat (1:5,000; sc-2768) or rabbit anti-mouse
(1:5,000; cat. no. sc-2005) IgG (Santa Cruz Biotechnology, Inc.).
Detection of the chemiluminescence reaction was performed using the
ECL kit (Pierce Biotechnology, Inc., Appleton, WI, USA). The GAPDH
signal was used as a loading control.
Results
Differential expression of miRNAs in
chordoma samples
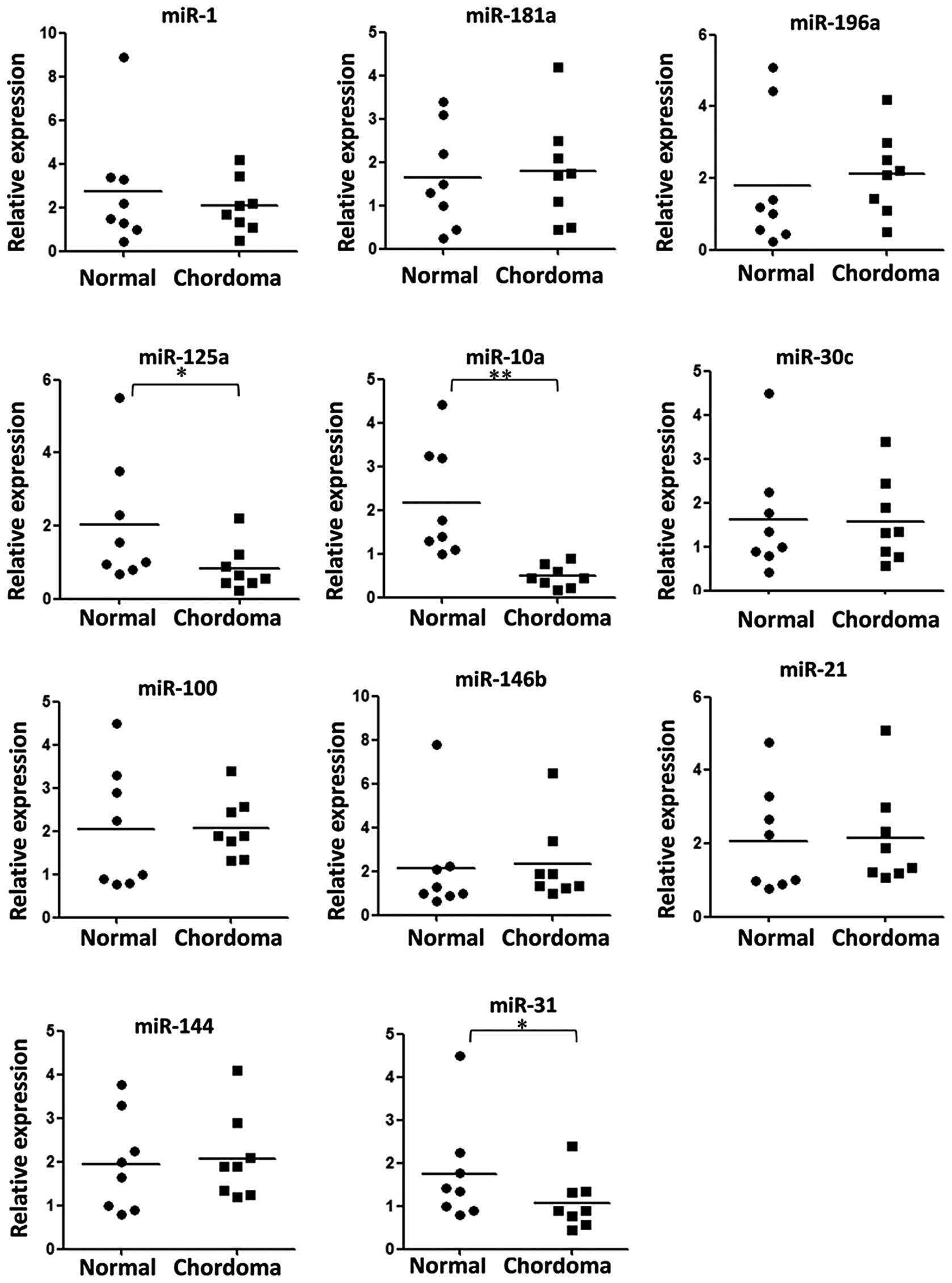
To determine whether there are differences in the
miRNA expression between chordoma and nucleus pulposus tissues, the
expression levels of 11 miRNAs that are associated with cancer
initiation were analyzed using RT-qPCR. The present results
demonstrated that the expression of miR-10a, miR-31 and miR-125a
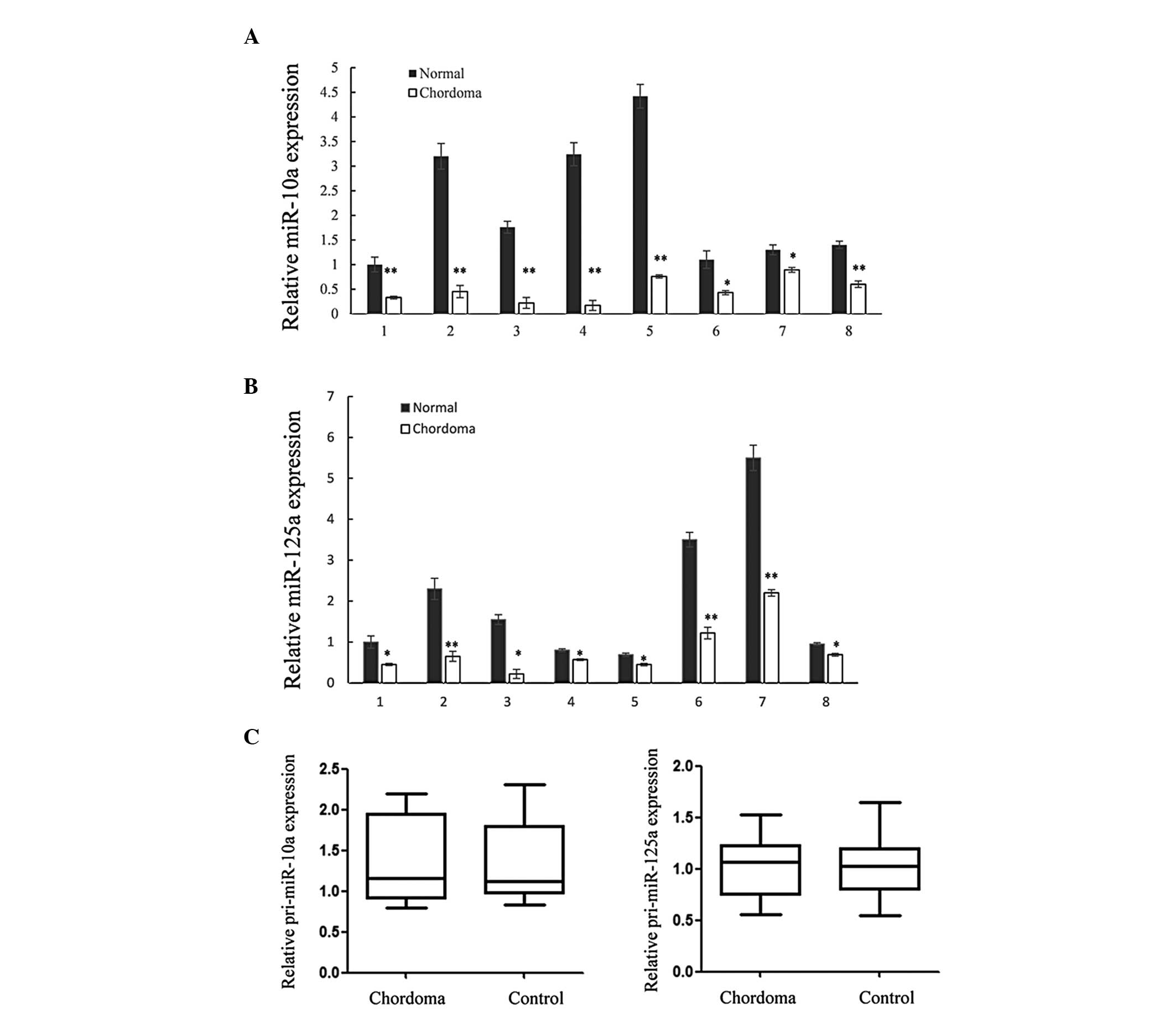
was significantly downregulated in chordoma tissues (Fig. 1). Furthermore, only the expression
of miR-10a and miR-125a was downregulated in all the sample pairs
(Fig. 2). To elucidate whether the
altered miRNA expression was caused at the transcriptional level or
post-transcriptional level, the expression of pri-miR-10a and
pri-miR-125a was also analyzed. As shown in Fig. 2C, the expression levels of
pri-miR-125a and pri-miR-10a were not altered in chordoma tissues,
suggesting that an altered post-transcriptional processing step was
performed.
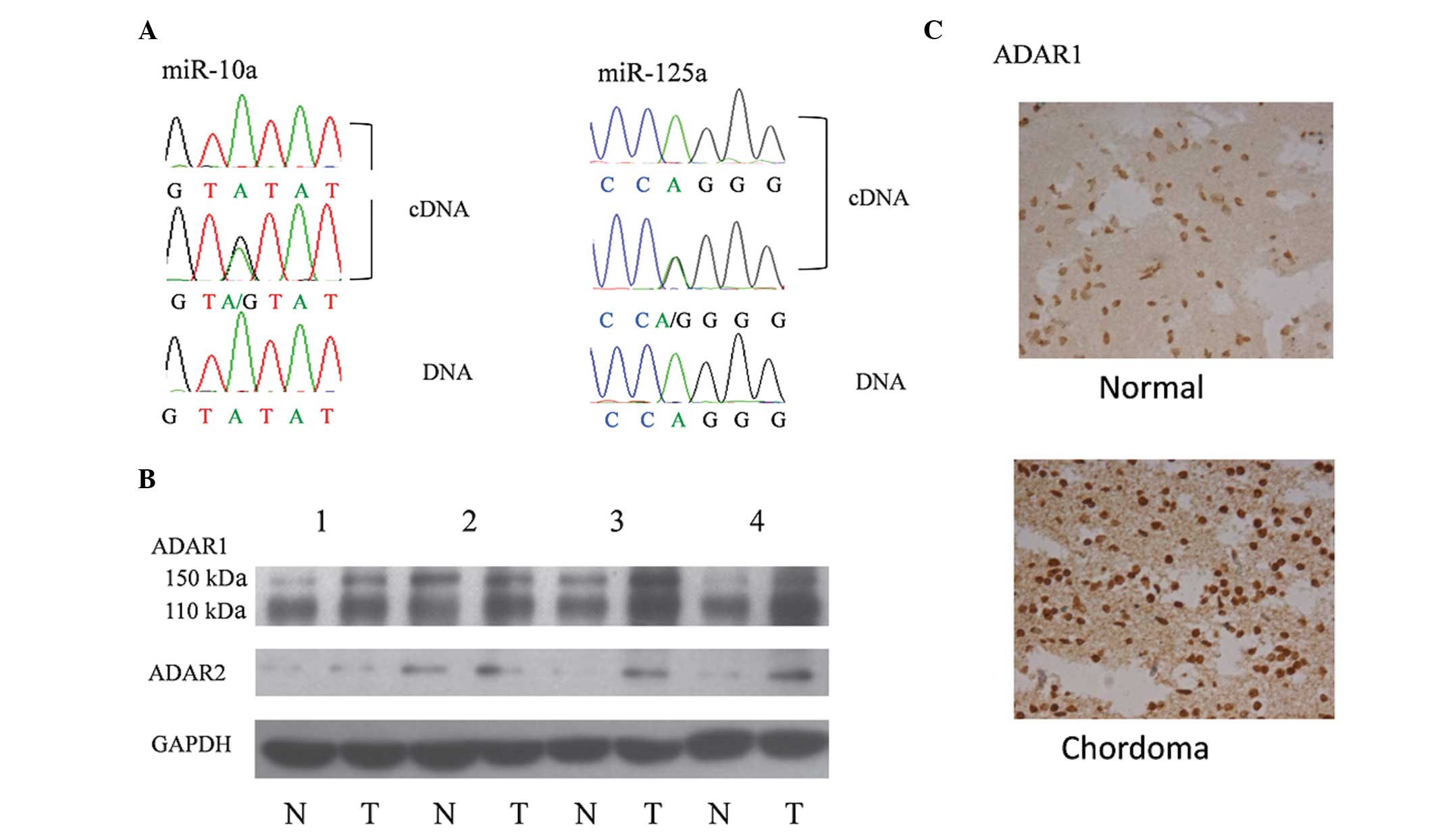
Adenosine to guanine (A-to-G) nucleotide
variations identified in sequencing results
Accumulating evidence has indicated that nucleotide
mutation may disturb miRNA processing and lead to altered miRNA
expression (17,18). Several studies have also reported
that RNA editing is common in mammals and altered RNA editing may
be associated with cancer initiation and processing (15,16).
To understand the reason for the reduction in miR-10a and miR-125a
expression, the DNA and cDNA sequences of these two miRNA
precursors were compared. Notably, an A-to-G variation in the
miR-10a and miR-125a cDNA sequence obtained from chordoma tissues
was identified, which was not observed in genomic DNA and cDNA from
the nucleus pulposus tissues (Fig.
3A). These results suggested that A-to-I RNA editing and
overexpression of ADARs may be associated with chordoma.
ADAR1 and ADAR2 are overexpressed in
chordoma tissues
To confirm the above hypothesis, the expression of
ADAR1 and ADAR2 was analyzed in chordoma and control tissues. The
results of western blot analysis and immunohistochemistry indicated
that the expression of ADAR1 was overexpressed in almost all the
chordoma tissues with upregulated ADAR2 in a number of the cancer
samples (Fig. 3B and C).
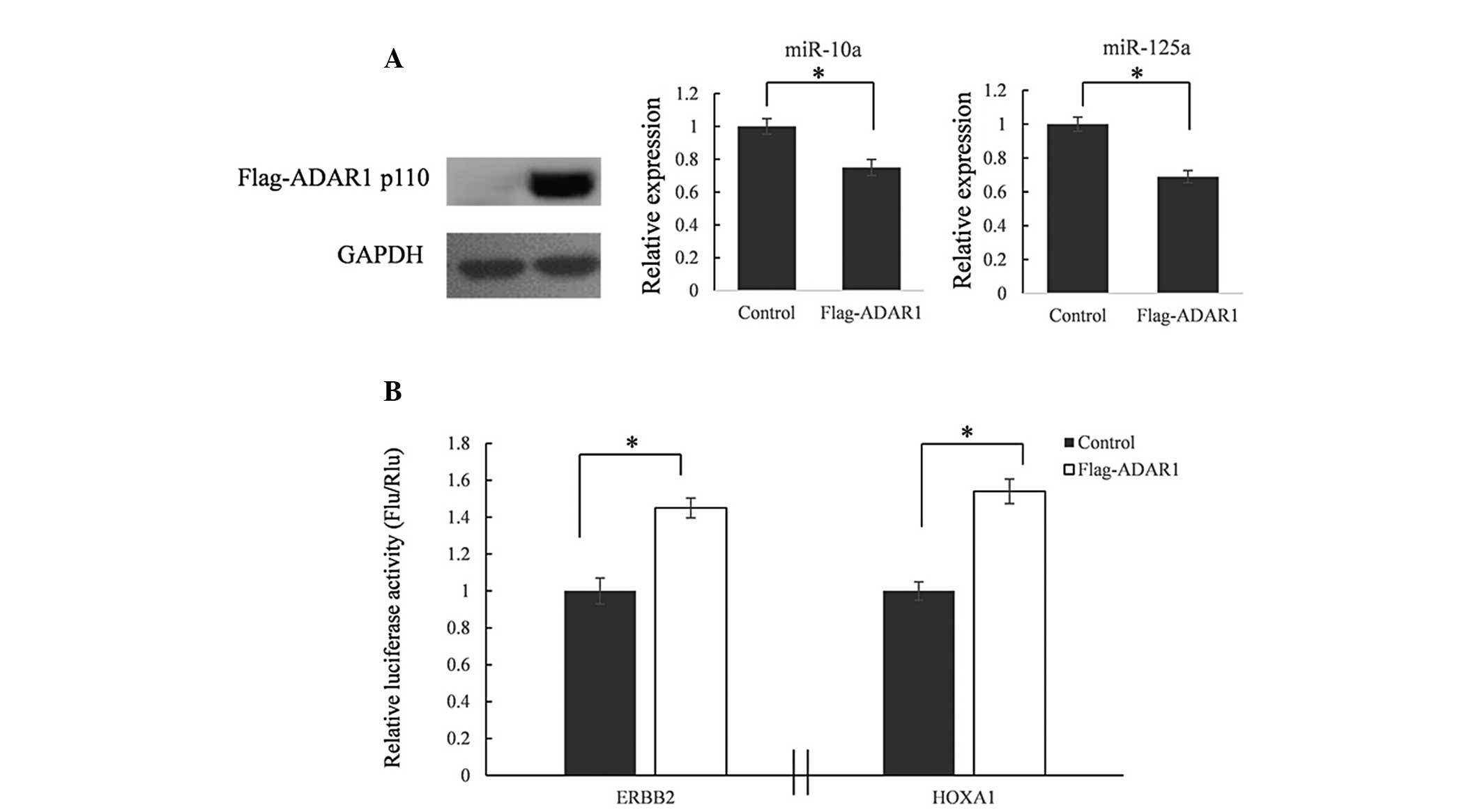
Overexpression of ADAR1 reduces miR-10a
and miR-125a expression and upregulates expression of their target
genes
To confirm the association between overexpressed
ADARs and downregulated miRNA expression, a FLAG-ADAR1 (110 kDa
isoform) expression vector was constructed. In HEK293T cells,
miR-10a and miR-125a were downregulated following overexpression of
ADAR1 using FLAG-ADAR1 expression vector transfection (Fig. 4A).
miRNAs function as important regulators in the
initiation, progression and metastasis of multiple types of cancer
through suppressing gene expression (19–22).
In order to determine the biological function of downregulated
miRNAs, ERBB2 and HOXA1 3′-UTR reporter vectors were
constructed, which were confirmed to be target genes of miR-10a or
miR-125a. 3′-UTR reporter vectors and FLAG-ADAR1 expression
vectors were co-transfected into HEK293T cells. After 48 h, total
RNAs were extracted and miR-10a and miR-125a were detected using
RT-qPCR. As shown in Fig. 4B, the
relative luciferase activities were significantly increased in the
ADAR1 overexpression groups.
Discussion
Chordoma, a rare type of bone tumor originating from
notochord remnants, is a slow growing malignant form of cancer.
Since it is relatively rare, few molecular studies have been
performed, however, further studies are required in order to enable
the development of novel strategies to treat chordoma. Previous
studies have demonstrated that disturbed expression of miR-1 and
miR-31 may be involved in chordoma pathogenesis, however, the
underlying mechanisms remain to be elucidated (8–10).
In the present study, the expression of 11 miRNAs were analyzed
using RT-qPCR and miR-125a, miR-10a and miR-31 were found to be
significantly down-regulated. By comparing the sequences of cDNA
and genomic DNA, it was identified that A-to-I RNA editing was
present in the pre-miR-10a and pri-miR-125a. In addition, the
expression of ADAR1 and ADAR2 was upregulated in chordoma tissues,
which is collateral evidence for RNA editing.
In the present study, the expression levels of
several miRNAs were detected, which have been reported tobe
dysregulated in chordoma, including miR-181a, miR-31, miR-1 and
miR-146b (8,9). Among them, only the reduction of
miR-31 expression was in accordance with the results reported in a
previous study (9). This
difference may be caused by differences in ancestries and the small
sample size.
It has been reported that miR-125a and miR-10a act
as tumor suppressors in numerous types of cancer, including gastric
cancer (23), thyroid cancer
(24), lung cancer (25,26)
and breast cancer (27,28). In the present study, ERBB2 (a
target gene of miR-125a) and HOXA1 (a target gene of miR-10a) were
selected to represent the biological function of ADAR1
overexpression. It was revealed that ADAR1 overexpression
upregulated luciferase activity by reducing the expression of
miR-125a and miR-10a, suggesting that ADAR1 is associated with
cancer pathogenesis by reducing antitumor miRNAs.
In conclusion, to the best of our knowledge, the
present study is the first to demonstrate disturbed ADAR1
expression in chordoma tissues. Overexpression of ADAR1 is
associated with chordoma pathogenesis by reducing tumor suppressor
miRNA expression, including miR-125a and miR-10a expression.
Acknowledgments
This study was supported by an AOSpine China
Research Grant (grant no. AOSCN(R)2013-06).
References
|
1
|
Bydon M, Papadimitriou K, Witham T, et al:
Novel therapeutic targets in chordoma. Expert Opin Ther Targets.
16:1139–1143. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Walcott BP, Nahed BV, Mohyeldin A, Coumans
JV, Kahle KT and Ferreira MJ: Chordoma: current concepts,
management and future directions. Lancet Oncol. 13:e69–e76. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
DeLaney TF, Duan Z and Hornicek FJ:
Proteomic profiling of chordoma. J Surg Oncol. 102:7192010.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Diaz RJ and Cusimano MD: The biological
basis for modern treatment of chordoma. J Neurooncol. 104:411–422.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Yang C, Schwab JH, Schoenfeld AJ, et al: A
novel target for treatment of chordoma: signal transducers and
activators of transcription 3. Mol Cancer Ther. 8:2597–2605. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Wu WK, Lee CW, Cho CH, et al: MicroRNA
dysregulation in gastric cancer: a new player enters the game.
Oncogene. 29:5761–5771. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Nicoloso MS, Spizzo R, Shimizu M, Rossi S
and Calin GA: MicroRNAs - the micro steering wheel of tumour
metastases. Nat Rev Cancer. 9:293–302. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Duan Z, Shen J, Yang X, et al: Prognostic
significance of miRNA-1 (miR-1) expression in patients with
chordoma. J Orthop Res. 32:695–701. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Bayrak OF, Gulluoglu S, Aydemir E, et al:
MicroRNA expression profiling reveals the potential function of
microRNA-31 in chordomas. J Neurooncol. 115:143–151. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Duan Z, Choy E, Nielsen GP, et al:
Differential expression of microRNA (miRNA) in chordoma reveals a
role for miRNA-1 in Met expression. J Orthop Res. 28:746–752.
2010.
|
|
11
|
Mo F, Wyatt AW, Sun Y, et al: Systematic
identification and characterization of RNA editing in prostate
tumors. PloS One. 9:e1014312014. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Valles I, Pajares MJ, Segura V, et al:
Identification of novel deregulated RNA metabolism-related genes in
non-small cell lung cancer. PloS One. 7:e420862012. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Chan TH, Lin CH, Qi L, et al: A disrupted
RNA editing balance mediated by ADARs (Adenosine DeAminases that
act on RNA) in human hepatocellular carcinoma. Gut. 63:832–843.
2014. View Article : Google Scholar :
|
|
14
|
Maas S, Patt S, Schrey M and Rich A:
Underediting of glutamate receptor GluR-B mRNA in malignant
gliomas. Proc Natl Acad Sci USA. 98:14687–14692. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Choudhury Y, Tay FC, Lam DH, et al:
Attenuated adenosine-to-inosine editing of microRNA-376a* promotes
invasiveness of glioblastoma cells. J Clin Invest. 122:4059–4076.
2012. View
Article : Google Scholar : PubMed/NCBI
|
|
16
|
Liu WH, Chen CH, Yeh KH, et al:
ADAR2-mediated editing of miR-214 and miR-122 precursor and
antisense RNA transcripts in liver cancers. PloS One. 8:e819222013.
View Article : Google Scholar
|
|
17
|
Yang W, Chendrimada TP, Wang Q, et al:
Modulation of microRNA processing and expression through RNA
editing by ADAR deaminases. Nat Struct Mol Biol. 13:13–21. 2006.
View Article : Google Scholar
|
|
18
|
Kawahara Y, Zinshteyn B, Sethupathy P,
Iizasa H, Hatzigeorgiou AG and Nishikura K: Redirection of
silencing targets by adenosine-to-inosine editing of miRNAs.
Science. 315:1137–1140. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Berindan-Neagoe I and Calin GA: Molecular
Pathways: microRNAs, cancer cells, and microenvironment. Clin
Cancer Res. 20:6247–6253. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Zhang Y, Schiff D, Park D and Abounader R:
MicroRNA-608 and microRNA-34a regulate chordoma malignancy by
targeting EGFR, Bcl-xL and MET. PloS One. 9:e915462014. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Fortunato O, Boeri M, Moro M, et al:
Mir-660 is downregulated in lung cancer patients and its
replacement inhibits lung tumorigenesis by targeting MDM2-p53
interaction. Cell Death Dis. 5:e15642014. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Finlay-Schultz J, Cittelly DM, Hendricks
P, et al: Progesterone downregulation of miR-141 contributes to
expansion of stem-like breast cancer cells through maintenance of
progesterone receptor and Stat5a. Oncogene. Sep 22–2014.Epub ahead
of print. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Jia H, Zhang Z, Zou D, et al: MicroRNA-10a
is down-regulated by DNA methylation and functions as a tumor
suppressor in gastric cancer cells. PLoS one. 9:e880572014.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Hudson J, Duncavage E, Tamburrino A, et
al: Overexpression of miR-10a and miR-375 and downregulation of
YAP1 in medullary thyroid carcinoma. Exp Mol Pathol. 95:62–67.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Markou A, Sourvinou I, Vorkas PA, Yousef
GM and Lianidou E: Clinical evaluation of microRNA expression
profiling in non small cell lung cancer. Lung Cancer. 81:388–396.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Jiang L, Huang Q, Chang J, Wang E and Qiu
X: MicroRNA HSA-miR-125a-5p induces apoptosis by activating p53 in
lung cancer cells. Exp Lung Res. 37:387–398. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Guo X, Wu Y and Hartley RS: MicroRNA-125a
represses cell growth by targeting HuR in breast cancer. RNA Biol.
6:575–583. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Hoppe R, Achinger-Kawecka J, Winter S, et
al: Increased expression of miR-126 and miR-10a predict prolonged
relapse-free time of primary oestrogen receptor-positive breast
cancer following tamoxifen treatment. Eur J Cancer. 49:3598–3608.
2013. View Article : Google Scholar : PubMed/NCBI
|