Introduction
Alzheimer's disease (AD) is a progressive
neurodegenerative disorder of the central nervous system, which is
characterized by a progressive loss of memory, deficits in
cognitive function and dementia (1,2). AD
is neuropathologically characterized by extracellular senile
plaques that contain β-amyloid (Aβ) precursor protein (APP), and
intracellular neurofibrillary tangles (NFTs), which occur as a
result of hyperphosphorylation of the tau protein (3,4). The
currently available treatment methods suppress a number of the
symptoms of AD, however they are unable to halt of reverse the
progression of this disease (5).
Numerous neuroprotective drugs have been discovered
in animal studies (2,4). However, no drug has been shown to
significantly slow or stop the progression of AD. This may be due
to the multiple pathways involved in the pathogenesis of AD,
including mitochondrial dysfunction, short-term and long-term
oxidative stress, energy crises, excitotoxicity, neuroinflammation
and protein aggregation (6).
Furthermore, the available neuroprotective agents often only target
one aspect of the disease process. Therefore, identifying effective
drug targets that are associated with the pathogenesis of AD is
critical for the development of novel AD therapeutic
strategies.
Ginsenoside Rg1 is a steroidal saponin that is
highly abundant in ginseng, in which ginsenoside Rg1 is one of the
most important components (7).
Recent studies have identified neuroprotective effects of Rg1
(8,9). However, the neuroprotective effects
of Rg1 on an AD animal model, and the underlying mechanisms, have
yet to be elucidated. Therefore, the aim of the present study was
to examine the effects of ginsenoside Rg1 on an animal model of AD,
and to determine the mechanisms underlying any such effects.
Materials and methods
Animals
Female Wistar rats (~3 months old; Animal Center of
Fuzhou Medical University, Fuzhou, China), were used in the present
study. The rats were housed in a temperature-controlled
environment, with a 12 h light/dark cycle and ad libitum
access to standard feed and water, which was limited to 4 h per
day. All animal experimentation was conducted in accordance with
the Canadian Council on Animal Care guidelines for the Care and Use
of Laboratory Animals, and was approved by Fujian Medical
University (Fuzhou, China). The present study was approved by the
Ethics Committee of Fuzhou General Hospital and Clinical Medical
College of Fujian Medical University.
Establishment of an AD model
A double transgenic APP/PS1 rat model (AD rats) was
established, which co-expressed mutations associated with AD that
are known to lead to accelerated plaque formation and increased Aβ
production. The model was generated by introducing the APP Swedish,
Florida and London mutations, and the presenilin 1 (PS1) M146L and
L286V mutations via Quikchange site-directed mutagenesis kit
(Agilent Technologies, Inc., Santa Clara, CA, USA). All of the
experimental procedures were conducted according to the
manufacturer's instructions. Transgenic assays were performed
according to the methods described by Devi et al (10). Normal control and APP/PS1 rats were
maintained in stainless-steel cages with food and water
available.
Treatment with Rg1
AD rats were fed with 0.5% Rg1-enriched food (Langze
Medicine Company, Fujian, China) for two months, prior to further
analyses. All of the rats were treated in accordance with the
guiding principles for the Care and Use of Animals of Fujian
Medical University (Fujian, China).
Preparation of brain tissue lysates
After two months, tissue lysates were prepared from
the brains of the rats. The rats were anesthetized with sodium
pentobarbital (30 mg/kg; Sigma-Aldrich, St. Louis, MO, USA) and
sacrificed, prior to isolation of brain tissue. Briefly, the brain
tissue was carefully isolated and incubated with 0.25% trypsin-EDTA
(Sigma-Aldrich) at 37°C for 1 h. The incubated tissue was then
lysed in 1 ml sample buffer (125 mM tris-HCl, pH 6.8; 2% SDS; 5%
glycerol; 0.003% bromophenol blue; and 1% 2-ME; Sigma-Aldrich). The
final protein concentrations of the brain tissue lysates were
determined using a Bicinchoninic Acid assay (Pierce Biotechnology,
Inc., Rockford, IL, USA), according to the manufacturer's
instructions. The tissue lysates were stored at −80°C until
required.
mRNA extraction and reverse
transcription-polymerase chain reaction (RT-PCR)
RNA was extracted from the brain tissue using
RNAsimple Total RNA kit (TIANGEN Biotech Co., Ltd., Beijing,
China). cDNA was reverse transcribed from 2 mg total mRNA using
SuperScript™ III First-Strand Synthesis system (Invitrogen Life
Technologies, Carlsbad, CA, USA), according to the manufacturer's
instructions. The specific primers for the endoplasmic reticulum
(ER) stress-associated genes, glucose-regulated protein 94 (Grp94),
tumor necrosis factor receptor-associated factor 2 (TRAF2),
X-box-binding protein 1 (XBP1) and eukaryotic initiating factor
(eIF)2α, were synthesized by Sangon Biotech Co., Ltd. (Shanghai,
China) according to Table I
(11,12). Quantitative RT-PCR was conducted
using SYBR Premix ExTaq II (Takara Biotechonology, Co., Ltd.,
Dalian, China), and detected with the ABI 7500 Real-Time PCR system
(Applied Biosystems Life Technologies, Foster City, CA, USA). The
reaction products were amplified by PCR in a volume of 50
µl, containing 1 µl forward primer, 1 µl
reverse primer, 1 µl Taq polymerase, 21 µl cDNA made
up to 50 µl with water, under the following conditions: 30
cycles at 94°C for 30 sec, 54°C for 30 sec and 72°C for 40 sec.
Following PCR, the products were electropho-resed on 1.5% agarose
gel, and the gel images were digitally captured using a
charge-coupled device camera (Bio-Rad Laboratories, Inc., Hercules,
CA, USA), prior to analysis with the National Institutes of Health
(NIH) Imager beta version 2 (NIH, Bethesda, MA, USA). The relative
transcriptional value of each gene in the semi-quantitative RT-PCR
experiments are presented as a ratio of the signal value, relative
to that of β-actin.
 | Table ISequences of the primers used for
semi-quantitative reverse transcription-polymerase chain reaction
analysis, to determine the mRNA expression of endoplasmic reticulum
stress-associated genes. |
Table I
Sequences of the primers used for
semi-quantitative reverse transcription-polymerase chain reaction
analysis, to determine the mRNA expression of endoplasmic reticulum
stress-associated genes.
| Gene | Primers | Sequence |
|---|
| TRAF2 | Sense |
5′-AAAGGGTCAGGAAGCCGTAG-3′ |
| Anti-sense |
5′-CCGCACATAGGAATTCTTGG-3′ |
| Grp94 | Sense |
5′-GGCCAGTTTGGTGTCGGTTT-3′ |
| Anti-sense |
5′-CTGGCCCCGTCCTAGAGTGTT-3′ |
| XBP1 | Sense |
5′-CCTTGTAGTTGAGAACCAGG-3′ |
| Anti-sense |
5′-GGGGCTTGGTATATATGTGG-3′ |
| eIF2α | Sense |
5′-GCGAATTCATGCCGGGGCTAAGTTGTAG-3′ |
| Anti-sense |
5′-CGCTCGAGTTAATCTTCAGCTTTGGCTT-3′ |
Western blot analysis
The brain tissue lysates were centrifuged at 10,000
× g for 30 min. Equal quantities of protein (50 µg) from
every group were separated by SDS-PAGE, using 15% gradient
tris/glycine gels, and then transferred to polyvinylidene
difluoride membranes (EMD Millipore, Billerica, MA, USA). The
membranes were blocked in 5% non-fat milk for 1 h, and then
incubated with the following primary antibodies: Mouse monoclonal
anti-caspase-3 (1:2,000; cat. no. sc-7148, Santa Cruz
Biotechnology, Inc., Dallas, TX, USA), mouse monoclonal anti-Grp78
(1:3,000; cat. no. sc-1050, Santa Cruz Biotechnology, Inc.), rabbit
polyclonal anti-B-cell lymphoma 2 (Bcl-2)(1:3,000; cat. no. sc-492,
Santa Cruz Biotechnology, Inc.), mouse monoclonal anti-Bcl-2
associated X protein (Bax) (1:2,000; cat. no. sc-23959, Santa Cruz
Biotechnology, Inc.), mouse monoclonal anti-inositol-requiring
enzyme 1 (IRE1) (1:2,000; cat. no. sc-390960, Santa Cruz
Biotechnology, Inc.), mouse monoclonal anti-activating
transcription factor 6 (ATF6) (1:1,000; cat. no. sc-166659, Santa
Cruz Biotechnology, Inc.), rabbit polyclonal anti-protein kinase
RNA-like ER kinase (PERK) (1:2,000; cat. no. ab-156919, Abcam,
Cambridge, UK), mouse monoclonal anti-cAMP response element-binding
transcription factor homologous protein (CHOP) (1:2,000; cat. no.
sc-7351; Santa Cruz Biotechnology, Inc.), mouse monoclonal
anti-caspase-12 (1:1,000, cat. no. Str-AAP-122C; Stressgen
Biotechnologies Corporation, San Diego, USA) and mouse monoclonal
anti-β-actin (1:3,000; cat. no. sc-8432; Santa Cruz Biotechnology,
Inc.) at 4°C overnight. The membranes were then washed and
incubated with horseradish peroxidase (HRP)-conjugated goat
anti-rabbit and rabbit anti-mouse secondary antibodies (1:4,000;
Santa Cruz Biotechnology, Inc.) for 2 h at room temperature.
Immunoreactive bands were visualized using the SuperSignal West
Pico Chemiluminescent substrate (Pierce Biotechnology, Inc.) and a
ChemiDoc XRS system with Quantity One software (Bio-Rad
Laboratories, Inc.).
Terminal
deoxynucleotidyl-transferase-mediated dUTP nick end labeling
(TUNEL) assay
Potential DNA fragments were examined using the
TUNEL Apoptosis Detection kit (EMD Millipore, Temecula, CA, USA).
Briefly, the cells were fixed with 4% paraformaldehyde in 0.1 M
NaH2PO4 (pH 7.4), and endogenous peroxidase
was inactivated using 3% H2O2. The cells were
incubated with the solution containing biotin-dUTP and terminal
deoxynucleotidyly transferase for 60 min. After end-horseradish
peroxidase, stained with diaminobenzidine, and counterstained with
ethyl green to detect biotin-labeled nuclei. Apoptotic bodies were
stained brown. The number of positively-stained cell nuclei was
counted under a light microscope (CX31; Olympas, Tokyo, Japan) in
three fields, by at least three independent observers.
Immunohistochemistry
The indirect streptavidin-biotin-peroxidase (SBP)
complex method (SBP kits; Wuhan Boster Biological Technology,
Wuhan, China) was used to visualize 4-µm cryostat serial
sections of the brain tissue. The cryostat serial sections were cut
using a Microm HM 500 M Cryostat (Menzel-Gläser, Brunswick,
Germany) and were placed on Superfrost Plus Gold glass slides
(Menzel-Gläser). The sections were incubated with rabbit polyclonal
antibodies against rat filamentous NFTs (1:1,000; cat. no.
ab136407, Abcam) and Aβ (1:2,000, cat. no. ab-120851, Abcam).
Antigen-antibody binding was determined using a HRP-labeled polymer
conjugated to goat anti-rabbit secondary antibodies (1:1,000, cat.
no. ab-6721; Abcam), using the dextranpolymer technique (DAKO,
Tokyo, Japan). The immunohistochemical images were observed and
captured using a photomicroscope (CX31; Olympus, Tokyo, Japan).
Statistical analysis
Statistical analyses were performed using SPSS
version 20.0 (IBM SPSS, Armonk, NY, USA). Student's two-tailed
t-test was used to compare protein and mRNA expression in the brain
tissue between the AD model and normal control rats. All results
are expressed as the mean ± standard deviation. In order to
determine the significance of differences between groups, an
analysis of variance was conducted. P<0.05 was considered to
indicate a statistically significant difference.
Results
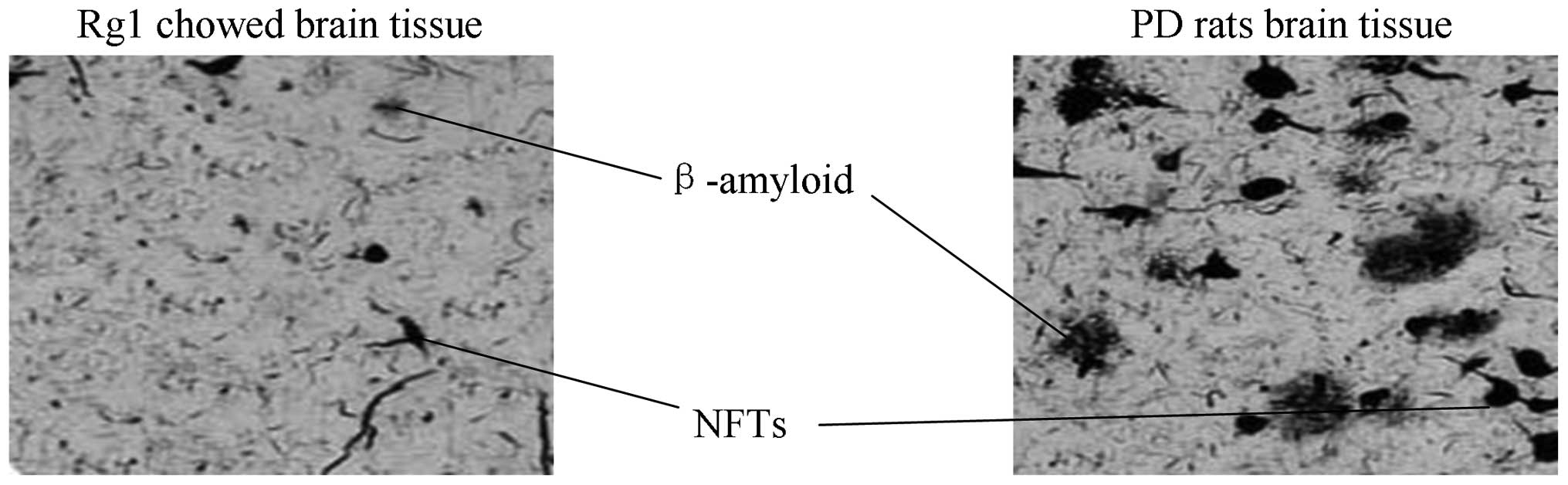
NFTs and Aβ plaques are decreased in
Rg1-treated AD rats
The present study determined the effects of Rg1
treatment on the formation of Aβ plaques and NFTs. NFTs were
significantly decreased in the Rg1-treated rats, compared with that
in the AD rats (Fig. 1). The
number of Aβ plaques were also significantly decreased in the Rg1
rats, compared with that in the AD rats (Fig. 1).
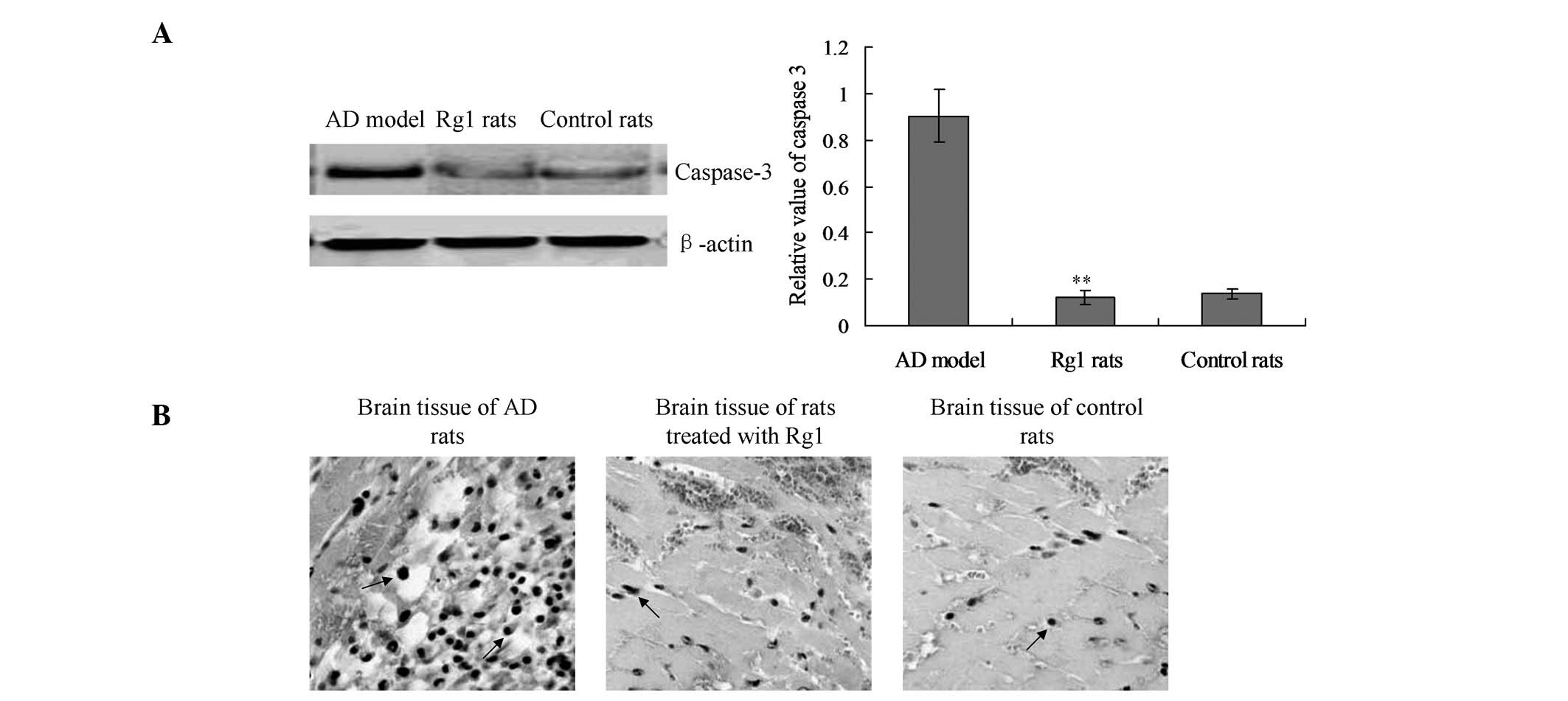
Rg1 inhibits cell apoptosis in AD
rats
Cell apoptosis of the rat brain tissue was detected
by a western blot analysis of caspase-3 and TUNEL analysis. The
protein levels of activated caspase-3 were significantly enhanced
in the AD rats, as compared with the control rats (Fig. 2A). Notably, the activation of
caspase-3 was significantly decreased in the Rg1-treated rats, as
compared with the AD rats (Fig.
2A; P<0.01). Furthermore, the levels of caspase-3 in the
Rg1-treated rats were similar to the levels observed in the control
rats.
The TUNEL assay also demonstrated that treatment
with Rg1 may affect apoptosis in AD rats. The number of
TUNEL-positive cells was significantly decreased in the Rg1-treated
rats, compared with that in the AD rats (Fig. 2B). These results suggest that Rg1
treatment may inhibit the apoptosis of cells in the brain tissue of
AD rats.
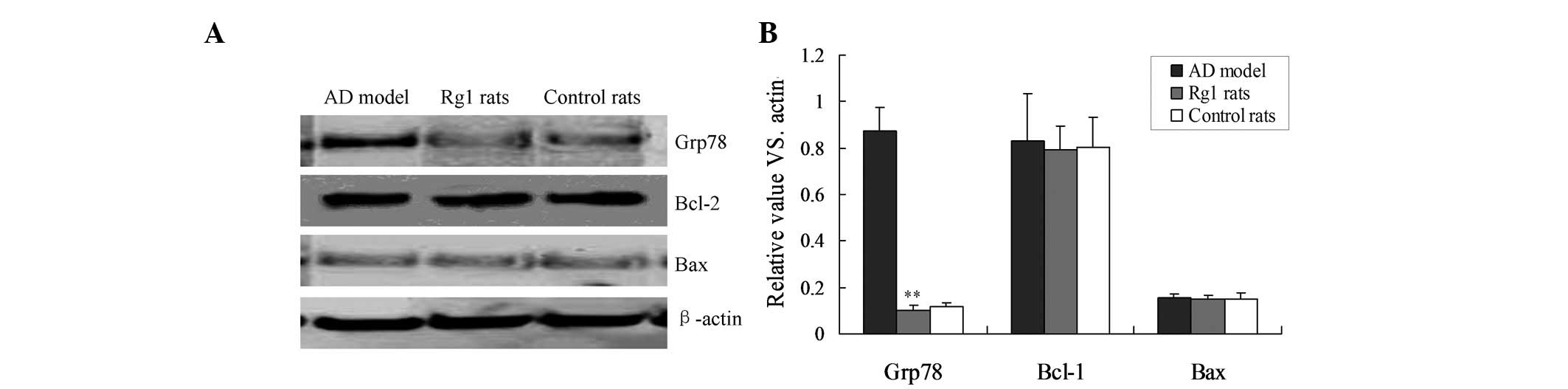
Rg1 inhibits apoptosis by blocking ER
stress
Grp78 has been identified as a marker of ER stress,
and Bcl-2 and Bax are markers of the mitochondrial apoptotic
pathway (13,14). Therefore the expression of the
Grp78, Bcl-2 and Bax proteins were detected by western blot
analysis. The protein expression levels of Grp78 were significantly
lower in the Rg1-treated rats, compared with that in the AD rats
(Fig. 3; P<0.01). However, no
significant differences were detected in the protein expression
levels of the mitochondria-associated apoptosis markers, Bcl-2 and
Bax, between the three groups (Fig.
3). These results indicate that ER stress, but not the
mitochondrial pathway, is involved in Rg1-mediated neuroprotective
function.
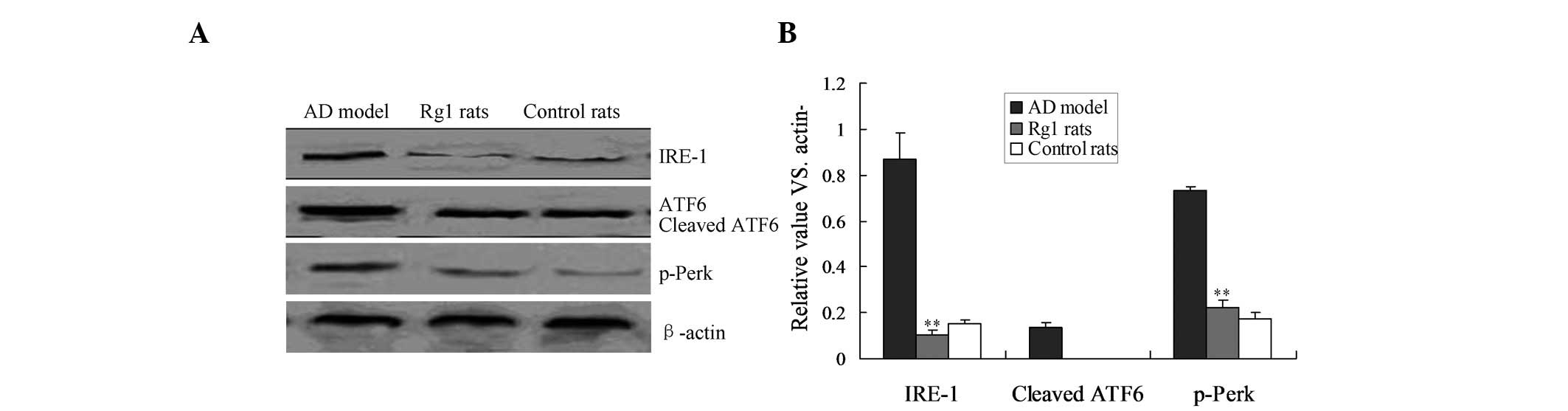
IRE-1 UPR pathway is involved in the
inhibition of Rg1-mediated apoptosis
According to the results of the present study, Rg1
inhibits apoptosis by blocking molecules involved in ER stress.
Therefore, the UPR factors involved in ER stress, including PERK,
IRE-1 and ATF6, were analyzed by western blot analysis. Treatment
with Rg1 reduced the expression of IRE-1 in the AD rats, and IRE-1
expression in the Rg1-treated rats was significantly decreased,
compared with that in the AD rats (Fig. 4; P<0.01). However, p-PERK and
cleaved ATF6 protein expression levels were not altered in any of
the three groups (Fig. 4).
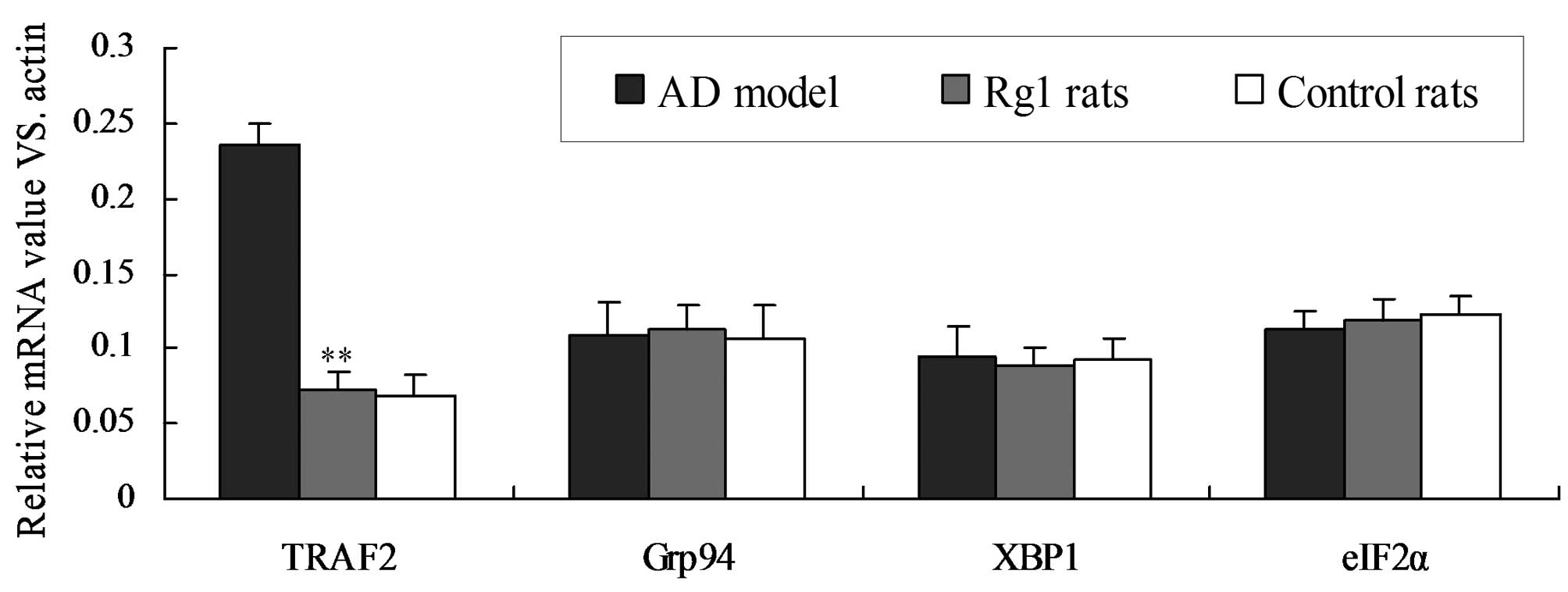
Rg1 inhibits ER stress by inhibiting the
transcription of ER stress-associated genes
In order to further determine the effects of Rg1 on
ER stress, the transcriptional levels of numerous genes associated
with ER stress, namely. TRAF2, Grp94, XBP1 and eIF2α, were analyzed
by semi-quantitative RT-PCR (Fig.
5). The signal intensity of each PCR product was determined
using a computer-assisted scanner and the relative expression value
was calculated by equilibrating the signal intensity to that of
β-actin. The mRNA expression of TRAF2 was significantly decreased
in the Rg1-treated rats, compared with that in the AD rats
(Fig. 5; P<0.01). However, the
expression levels of Grp94, XBP1 and eIF2α was similar between the
different groups. These results suggest that ER stress is inhibited
in AD rats following treatment with Rg1.
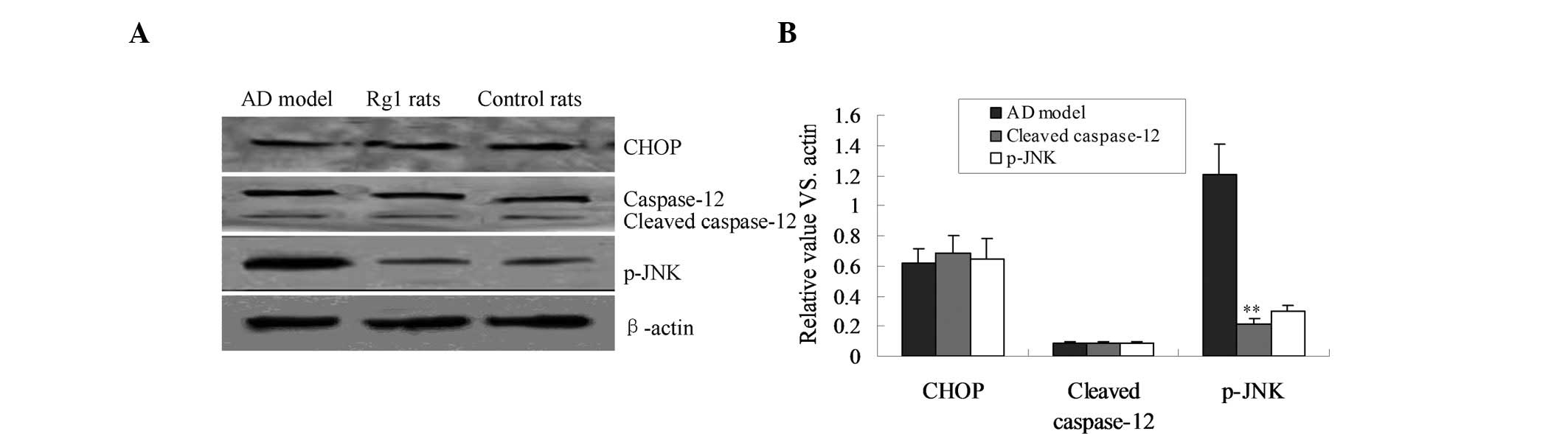
JNK apoptotic pathway is involved in the
neuroprotective effects of Rg1
In order to investigate the possible apoptotic
pathways involved in the neuroprotective effects of Rg1, the JNK,
CHOP and caspase-12 apoptotic pathways were evaluated by western
blot analysis. The protein expression of p-JNK was significantly
increased in the AD rats, compared with that in the control rats
(Fig. 6). However, when the AD
rats were treated with Rg1 the p-JNK expression was shown to be
significantly reduced, compared with that in the AD rats, and these
expression levels were similar to those detected in the control
rats (Fig. 6; P<0.01).
Furthermore, treatment with Rg1 had no effect on the expression of
CHOP, and the cleavage of caspase-12.
Discussion
Aβ and intracellular NFTs are the primary pathogenic
agents involved in the development of AD (3,15).
Therefore, the formation of Aβ and NFTs is an important target in
the treatment of AD. Furthermore, the progression of NFTs and Aβ
accumulation is associated with the severity of the neuronal
cytopathology of AD (16,17). The present study aimed to explore
the neuroprotective functions of Rg1, and to determine its
underlying mechanisms, in an AD rat model. Previous studies have
demonstrated that Ginsenoside Rg1 may attenuate the generation of
Aβ and improve cognitive impairment, in a rat AD model (17,18).
However, the specific neuroprotective mechanisms of Rg1 have yet to
be elucidated. Therefore, the present study aimed to investigate
the possible mechanisms by which Rg1 inhibits the accumulation of
NFTs and Aβ.
The present study demonstrated that the accumulation
of NFTs and Aβ was significantly decreased in AD rats, following
treatment with Rg1. Dai et al (19) previously indicated that Aβ 1–42 may
induce neurotoxicity, by initiating the apoptosis of rat primary
hippocampal neurons. Therefore, the present study investigated the
level of cell apoptosis in the brain tissue of AD rats. Caspase-3
levels and TUNEL-positive cells were significantly reduced in
Rg1-treated rats, compared with those in AD rats, thus indicating
that apoptosis was inhibited in the brain tissue of AD rats,
following treatment with Rg1. Therefore, it is hypoth-esized that
the accumulation of NFTs and Aβ may be caused by apoptosis of brain
tissue. This result is in accordance with findings from previous
studies (20,21).
Three primary apoptotic pathways have previously
been identified: Mitochondrial, ER stress and death receptor
pathways (22). It is well-known
that Bcl-2 and Bax are key biomarkers for the mitochondria-mediated
apoptotic pathway (11), whereas
Grp78 is the critical biomarker for the ER stress-mediated
apoptotic pathway (23).
Therefore, the present study detected the expression levels of
Bcl-2, Bax and Grp78 in the rat AD model and Rg1-treated rats.
There was no significant difference between the expression levels
of Bcl-2 and Bax in the AD model and Rg1-treated rats. However, the
Grp78 expression levels were significantly reduced in the brain
tissue of the Rg1-treated rats, compared with those in the AD rats
(P<0.01). These results indicate that the ER stress-associated
apoptotic pathway is involved in the neuroprotective function of
Rg1.
In order to investigate the underlying mechanism of
Rg1 in neuroprotection, the expression of the ER stress UPR
pathway-associated proteins, p-PERK, IRE-1 and cleaved ATF6, was
measured. The protein expression of IRE-1 and p-PERK was increased
in the AD model rats, while it was significantly decreased in the
Rg1-treated rats (P<0.01). This suggests that the PERK and IRE-1
pathway may be involved in the Rg1-mediated inhibition of the ER
stress apoptotic pathway. Understanding the mechanism underlying
Rg1-mediated apoptosis inhibition may help to further illuminate
the role of Rg1 in delaying AD progression. A previous study showed
that activation of IRE-1 in turn activates TRAF2 and XBP1 (24), and that PERK activation leads to
phosphorylation of eIF-2α (25).
The expression of TRAF2, XPB1 and PERK are all associated with the
subsequent initiation of apoptosis. The results of the present
study provide evidence that indicates the emergence of ER stress in
the AD model. Notably, treatment with Rg1 could inhibit the mRNA
transcription of TRAF2 in the AD rat model. Therefore it may be
hypothesized that treatment with Rg1 may inhibit the formation of
NFTs and Aβ in AD.
An important ER stress-mediated apoptotic pathway is
the JNK pathway, which is mediated by IRE-1 (26). The IRE-1 cytoplasmic domain
interacts with the adaptor protein, TRAF2, which subsequently
phosphorylates and activates JNK. In the present study IRE-1 and
TRAF2 expression was shown to be altered in Rg1-treated rats.
Furthermore, the ER stress-associated proteins, cleaved caspase-12
and CHOP, were also measured in order to identify the specific
apoptotic proteins involved in the development of AD. Xu et
al (27) previously reported
that cleaved caspase-12 activates caspase-3 and triggers apoptosis,
and that CHOP and p-JNK directly induce ER stress-associated
apoptosis. The present study demonstrated that there was no
significant changes to the expression of CHOP and cleaved
caspase-12 in any of the three of groups (P>0.05). Notably, when
treated with Rg1 the p-JNK expression in AD rats was significantly
decreased, compared with that in the AD model rats (P<0.05).
These results suggest that treatment with Rg1 may inhibit p-JNK
pathway-induced apoptosis, and indirectly inhibit the accumulation
of NFTs and Aβ. Thus, Rg1 may block p-JNK-induced ER stress, and
slow the progression of AD.
In conclusion, in the present study Rg1 was shown to
act as an important factor that inhibits the accumulation of NFTs
and Aβ, through blocking the ER stress-mediated apoptotic pathway.
Blocking of this pathway was triggered by the IRE-1 and TRAF2
pathway, via inhibition of the expression of p-JNK.
References
|
1
|
Xing S, Shen D, Chen C, Wang J, Liu T and
Yu Z: Regulation of neuronal toxicity of β-amyloid oligomers by
surface ATP synthase. Mol Med Rep. 8:1689–1694. 2013.PubMed/NCBI
|
|
2
|
He Y, Zhao H and Su G: Ginsenoside Rg1
decreases neurofibrillary tangles accumulation in retina by
regulating activities of neprilysin and PKA in retinal cells of AD
rats model. J Mol Neurosci. 52:101–106. 2014. View Article : Google Scholar
|
|
3
|
Galimberti D and Scarpini E: Progress in
Alzheimer's disease. J Neurol. 259:201–211. 2012. View Article : Google Scholar
|
|
4
|
Sima X, Xu J, Li J, Zhong W and You C:
Expression of β-amyloid precursor protein in refractory epilepsy.
Mol Med Rep. 9:1242–1248. 2014.PubMed/NCBI
|
|
5
|
Sethi KD: Clinical aspects of Parkinson
disease. Curr Opin Neurol. 15:457–460. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Goes AT, Souza LC, Filho CB, Del Fabbro L,
De Gomes MG, Boeira SP and Jesse CR: Neuroprotective effects of
swimming training in a mouse model of Parkinson's disease induced
by 6-hydroxydopamine. Neuroscience. 256:61–71. 2014. View Article : Google Scholar
|
|
7
|
Wu J, Pan Z, Cheng M, Shen Y, Yu H, Wang Q
and Lou Y: Ginsenoside Rg1 facilitates neural differentiation of
mouse embryonic stem cells via GR-dependent signaling pathway.
Neurochem Int. 62:92–102. 2013. View Article : Google Scholar
|
|
8
|
Zhuang P, Zhang Y and Pang T:
Proliferation effect of neural stem cell of ginsenoside Rg1 in
vitro. Zhongguo Zhong Yao Za Zhi. 34:443–446. 2009.In Chinese.
PubMed/NCBI
|
|
9
|
Lu MC, Lai TY, Hwang JM, Chen HT, Chang
SH, Tsai FJ, Wang HL, Lin CC, Kuo WW and Huang CY: Proliferation-
and migration-enhancing effects of ginseng and ginsenoside Rg1
through IGF-1 and FGF-2 signaling pathways on RSC96 Schwann cells.
Cell Biochem Funct. 27:186–192. 2009. View
Article : Google Scholar : PubMed/NCBI
|
|
10
|
Devi L and Ohno M: Genetic reductions of
beta-site amyloid precursor protein-cleaving enzyme 1 and
amyloid-beta ameliorate impairment of conditioned taste aversion
memory in 5XFAD Alzheimer's disease model rats. Eur J Neurosci.
31:110–118. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Xu K, Wang X, Shi Q, Chen C, Tian C, Li
XL, Zhou RM, Chu YL and Dong XP: Human prion protein mutants with
deleted and inserted octarepeats undergo different pathways to
trigger cell apoptosis. J Mol Neurosci. 43:225–234. 2011.
View Article : Google Scholar
|
|
12
|
Lee SK and Kim YS: Phosphorylation of
eIF2α attenuates statin-induced apoptosis by inhibiting the
stabilization and translocation of p53 to the mitochondria. Int J
Oncol. 42:810–816. 2013.PubMed/NCBI
|
|
13
|
Hegde RS, Mastrianni JA, Scott MR, DeFea
KA, Tremblay P, Torchia M, DeArmond SJ, Prusiner SB and Lingappa
VR: A transmembrane form of the prion protein in neurodegenerative
disease. Science. 279:827–834. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Li Z, Shen J, Chen Y, Pan J, Zeng H, Fang
H, Ye Z, Zeng C, Zhang R and Cai D: Mitochondrial genome
sequenceing of chondrocytes in osteoarthritis by human mitochondria
RT2 Profiler™ PCR array. Mol Med Rep. 6:39–44. 2012.PubMed/NCBI
|
|
15
|
Zhao H, Chang R, Che H, Wang J, Yang L,
Fang W, Xia Y, Li N, Ma Q and Wang X: Hyperphosphorylation of tau
protein by calpain regulation in retina of Alzheimer's disease
transgenic mouse. Neurosci Lett. 551:12–16. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Lai MK, Chen CP, Hope T and Esiri MM:
Hippocampal neuro-fibillary tangle changes and aggressive behaviour
in dementia. Neuroreport. 21:1111–1115. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Kril JJ, Patel S, Harding AJ and Halliday
GM: Neuron loss from the hippocampus of Alzheimer's disease exceeds
extracellular neurofibrillary tangle formation. Acta Neuropathol.
103:370–376. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Chen LM, Lin ZY, Zhu YG, Lin N, Zhang J,
Pan XD and Chen XC: Ginsenoside Rg1 attenuates β-amyloid generation
via suppressing PPARγ-regulated BACE1 activity in N2a-APP695 cells.
Eur J Pharmacol. 675:15–21. 2012. View Article : Google Scholar
|
|
19
|
Dai X, Chang P, Zhu Q, Liu W, Sun Y, Zhu S
and Jiang Z: Chitosan oligosaccharides protect rat primary
hippocampal neurons from oligomeric β-amyloid 1-42-induced
neurotoxicity. Neurosci Lett. 554:64–69. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Chen Z and Zhong C: Decoding Alzheimer's
disease from perturbed cerebral glucose metabolism: implications
for diagnostic and therapeutic strategies. Prog Neurobiol.
108:21–43. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Salminen A, Kaarniranta K, Kauppinen A,
Ojala J, Haapasalo A, Soininen H and Hiltunen M: Impaired autophagy
and APP processing in Alzheimer's disease: The potential role of
Beclin 1 interactome. Prog Neurobiol. 106–107:33–54. 2013.
View Article : Google Scholar
|
|
22
|
Gillies LA and Kuwana T: Apoptosis
regulation at the mitochondrial outer membrane. J Cell Biochem.
115:632–640. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Wang X, Shi Q, Xu K, Gao C, Chen C, Li XL,
Wang GR, Tian C, Han J and Dong XP: Familial CJD associated PrP
mutants within transmembrane region induced Ctm-PrP retention in ER
and triggered apoptosis by ER stress in SH-SY5Y cells. PLoS One.
6:e146022011. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Fan Q, Hu Y, Pang H, Sun J, Wang Z and Li
JM: Melittin protein inhibits the proliferation of MG63 cells by
activating inositol-requiring protein-1α and X-box binding protein
1-mediated apoptosis. Mol Med Rep. 9:1365–1370. 2014.PubMed/NCBI
|
|
25
|
Kimball SR and Jefferson LS: Induction of
REDD1 gene expression in the liver in response to endoplasmic
reticulum stress is mediated through a PERK, eIF2α phosphorylation,
ATF4-dependent cascade. Biochem Biophys Res Commun. 427:485–489.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Yenki P, Khodagholi F and Shaerzadeh F:
Inhibition of phosphorylation of JNK suppresses Aβ-induced ER
stress and upregulates prosurvival mitochondrial proteins in rat
hippocampus. J Mol Neurosci. 49:262–269. 2013. View Article : Google Scholar
|
|
27
|
Xu K and Zhu XP: Endoplasmic reticulum
stress and prion diseases. Rev Neurosci. 23:79–84. 2012. View Article : Google Scholar : PubMed/NCBI
|