Introduction
Bone marrow stem cells (BMSCs) are currently under
active investigation due to their therapeutic potential for bone
tissue engineering and cell replacement therapy (1). BMSCs were initially described in
1966, when they were isolated and cultured from bone marrow rat
cartilage cells (2). A previous
study showed that BMSCs migrate toward damaged bone tissue, thereby
demonstrating the therapeutic potential of BMSCs for tissue injury
repair and cell replacement therapy (3). In addition, BMSCs have been reported
to survive in high hypoxic/ischemic inflammatory environments
(4). The present study aimed to
investigate the mechanism underlying BMSC migration, as well as the
role of BMSCs in tissue repair-associated protein regulation. This
was achieved by modulating BMSC motility and viability through
genetic engineering (5,6).
Peroxisome proliferator activated receptor γ (PPARγ)
is a subtype of PPAR, which mediates BMSC differentiation into
adipocytes (7). PPARγ is a
ligand-activated nuclear transcription factor, which is involved in
cellular differentiation, growth, and apoptosis (8). PPARγ mediates the differentiation of
BMSCs into adipocytes, and an increase in the protein expression
levels of PPARγ results in a significant increase in adipocyte
differentiation, concomitant with reduced levels of osteogenic
differentiation, in the bone marrow (9). Therefore, PPARγ may prove useful in
the treatment and prevention of osteoporosis, a disease in which
deregulated BMSC differentiation, dedifferentiation, and
transdifferentiation leads to a decrease in the number of
osteoblasts, and an increase in the number of adipocytes (10).
Micro (mi)RNAs are small (19–22 nucleotides)
endogenous non-coding RNA molecules, which suppress the expression
of target genes (11). miRNAs have
an important role in numerous biological processes, including
cellular proliferation, differentiation, and apoptosis. Previous
studies have shown that the upregulation of miR-20a expression
leads to an increase in the protein expression levels of the
adipogenic PPARγ marker gene in rats, as compared with the controls
(12,13).
Naringin is a dihydrotestosterone flavonoid
compound, which markedly inhibits bone loss, improves bone density,
and enhances biomechanical anti-compression performance (14). Naringin has been shown to promote
the proliferation and differentiation of MC3T3-E1 osteoblast
precursor cells in vitro; however, it does not promote bone
mineralization (15). High
concentrations of naringin increase the synthesis and activity
levels of alkaline phosphatase (ALP), promote osteoblast
differentiation, and stimulate bone formation (16). A previous study demonstrated that
naringin increases the activity levels of osteocalcin (OC) and ALP,
and promotes the proliferation and differentiation of MC3T3-E1
osteoblast precursor cells, without affecting bone mineralization
(16).
The present study aimed to investigate whether
naringin was able to enhance BMSC differentiation and inhibit
adipocyte formation by modulating osteogenesis via the upregulation
of miR-20a expression, and the downregulation of PPARγ
expression.
Materials and methods
Drugs and reagents
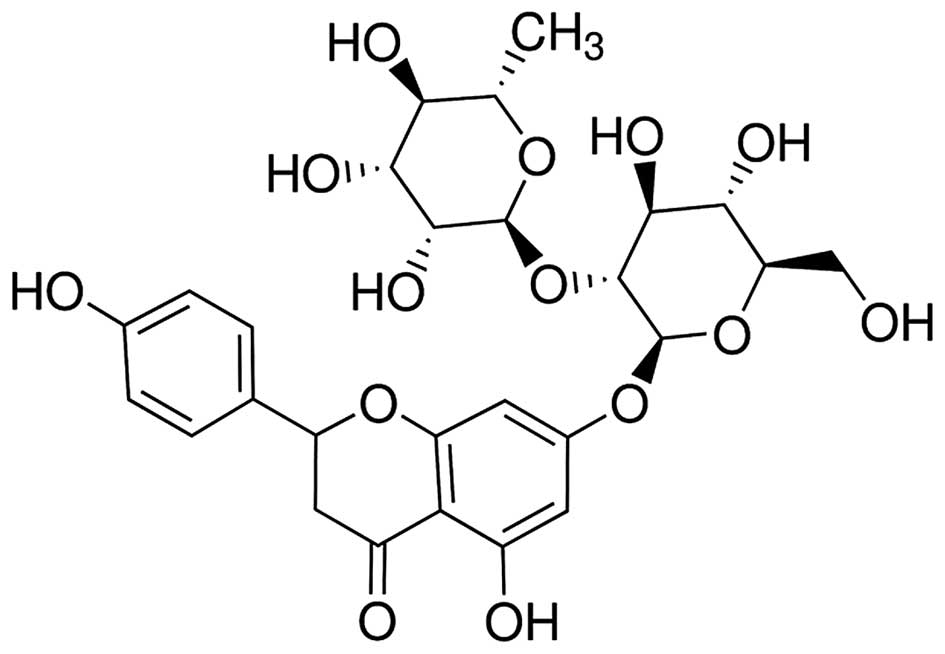
Naringin (Fig. 1)
at a purity >95% (Sigma-Aldrich, St Louis, MO, USA) was
dissolved in physiological saline, according to the manufacturer's
instructions. Dulbecco's modified Eagle's medium (DMEM) and fetal
bovine serum (FBS) were purchased from Gibco Life Technologies
(Carlsbad, CA, USA), and Invitrogen Life Technologies (Carlsbad,
CA, USA), respectively. MTT assay and Lipofectamine®
2000 were purchased from Sigma-Aldrich.
Animals and BMSC isolation
Male and female New Zealand white rabbits (4–8 weeks
old) weighing 2.0±0.5 kg were provided by the Experimental Animal
Center of Dalian Medical University (Dalian, China). The rabbits
were maintained in individual cages at 21–23°C, under a 12 h
light-dark cycle (8:00; 20:00) and humidity level of 55–65%. The
study was approved by the ethics committee of The First Affiliated
Hospital of Dalian Medical University (Dalian, China). The rabbits
were anesthetized by intravenous injection of 30 mg/kg
pentobarbital (Invitrogen Life Technologies). A total of 2–3 ml
bone marrow was subsequently collected from the lateral tibial
tubercle of the rabbits using a puncture needle. The bone marrow
cell suspension was then mixed with an equal volume of Percoll
solution (Duke Scientific, Palo Alto, CA, USA), forming a clear
interface. Density gradient centrifugation was performed at 1,200 ×
g for 20 min, in order to isolate the BMSCs. The supernatant was
discarded and the mononuclear BMSCs were extracted. The BMSCs were
washed with phosphate-buffered saline (PBS) and further centrifuged
at 1,000 × g for 10 min. The supernatant was discarded and 10 ml
DMEM supplemented with 15–20% FBS was added to the precipitate,
prior to incubation at 37°C in a humidified atmosphere containing
5% CO2. The cells were cultured until they reached 80%
confluence and were then used for subsequent experimentation.
Cell proliferation assay
The levels of BMSC proliferation were determined
using an MTT assay. BMSCs were seeded at a density of
2.0×104 cells/well into 96-well plates at 37°C in a
humidified atmosphere containing 5% CO2. The BMSCs were
then treated with either 0.01, 0.1, 1, 10 and 100 µM
naringin (17) for 48 h, or with
medium containing 0, 0.1, 1, and 10 µM naringin for 3, 7, 14
and 21 days. For the MTT assay, 10 µl MTT was added to each
well prior to incubation at 37°C in a humidified atmosphere
containing 5% CO2 for 4 h. The medium was discarded, and
150 µl dimethyl sulfoxide (Beyotime Institute of
Biotechnology, Nanjing, China) was added to the solution prior to
further incubation at room temperature with gentle agitation for 10
min. The optical density of the solution was read at 570 nm using a
microplate reader (Sunrise, Tecan Trading AG, Männedorf,
Switzerland).
Quantification of the protein expression
levels of PPARγ by western blot analysis
BMSCs were treated with various concentrations of
naringin (0.1, 1, and 10 µM) for 48 h. The protein
expression levels of PPARγ were then determined by western
blotting. The protein concentrations were determined using a
Novagen® Bicinchoninic Acid Protein Assay kit (Novagen,
Merck Millipore, Darmstadt, Germany). Protein was extracted from
cells using Cell Lysis Buffer (Cell Signaling Technology, Inc.,
Danvers, MA, USA). Equal quantities of protein was separated by 10%
SDS-polyacrylamide gel and blotted onto polyvinylidene fluoride
membranes (Millipore). The primary antibodies used for the western
blot analysis were as follows: Anti-PPARγ (cat. no. 2435S; 1:500;
Cell Signaling Technology, Inc.) and anti-β-actin (cat. no.
sc-69879; 1:2,000; Santa Cruz Biotechnology, Inc., Dallas, TX, USA)
at 4°C overnight. Subsequently, membranes were washed and incubated
with a horseradish peroxidase-conjugated anti-rabbit IgG secondary
antibody (cat. no. C520011; 1:20,000, Sangon Biotech Co., Ltd.,
Shanghai, China). The blots were visualized using an enhanced
chemiluminescence assay kit (GE Healthcare Life Sciences, Little
Chalfont, UK), and analyzed using VersaDoc Gel Imaging system
(Bio-Rad Laboratories, Inc., Hercules, CA, USA).
Quantification of the mRNA expression
levels of OC, ALP, collagen type I (Col I), and miR-20a by reverse
transcription-quantitative polymerase chain reaction
In order to determine the mRNA expression levels of
OC, ALP, and Col I, the BMSCs were treated with 1 µM
naringin for 3, 7, 14 and 21 days or with medium containing 0, 0.1,
1, and 10 µM naringin for 48 h. Total RNA was extracted from
the BMSCs using TRIzol® reagent (Invitrogen Life
Technologies). A total of 1 µg total RNA was subsequently
reverse transcribed into cDNA using an RTq-PCR mixture (Takara
Biotechnology Co., Ltd., Dalian, China). A 5 ng cDNA template was
used to carry out the reaction. Various primers (Sangon Biotech
Co., Ltd.) were used in order to amplify OC, ALP, and Col I
(Table I). In order to determine
the expression levels of miR-20a, the BMSCs were treated with 0.1,
1, and 10 µM naringin for 48 h. Total RNA containing miR-20a
was extracted from the BMSCs using a miRNeasy RNA Isolation kit
(Bogoo Biomart, Shanghai, China). The expression levels of miR-20a
were subsequently analyzed by RT-qPCR qPCR (7500 Real-Time PCR
system; Applied Biosystems, Foster City, CA, USA) according to the
manufacturer's instructions. PCR cycling conditions were as
follows: 35 cycles at 95°C for 30 sec, 60°C for 45 sec and 72°C for
50 sec. The sequence of the miR-20a primer is shown in Table I. cDNA was reverse transcribed from
100 ng total miRNA using a QuantiTect Reverse Transcription kit
(Bogoo Biomart). mRNA expression levels were determined using the
2−ΔΔCt method of relative quantification.
 | Table IPrimer design for reverse
transcription-quantitative polymerase chain reaction. |
Table I
Primer design for reverse
transcription-quantitative polymerase chain reaction.
| Gene | Primers | Annealing temperature
(cycle) |
|---|
| Osteocalcin | F:
5′-CATGAGAGCCCTCACA-3′ | 56°C (32) |
| R:
5′-AGAGCGACACCCTAGAC-3′ | |
| Alkaline
phosphatase | F:
5′-TCAGAAGCTCAACACCAACG-3′ | 55°C (32) |
| R:
5′-GTCAGGGACCTGGGCATT-3′ | |
| Collagen type I | F:
5′-TGACCTCAAGATGTGCCACT-3′ | 57°C (30) |
| R:
5′-GGGAGTTTCCATGAAGCCAC-3′ | |
| microRNA-20a |
F:5′-AAGAATTTAAATAAAAAAAAAGAACA-3′ | 57°C (32) |
| R:
5′-CACGGGCTGAGGAAAATA-3′ | |
| β-actin |
F:5′-GCTCTCCAGAACATCACTCCTGCC-3′ | 57°C (30) |
|
R:5′-CGTTGTCATACCAGGAAATGAGCTT-3′ | |
BMSC transfection with miR-20a and
anti-miR-20a
miR-20a precursor and anti-miR-20a (Ambion Life
Technologies, Carlsbad, CA, USA) were synthesized by Sangon Biotech
Co., Ltd. miR-20 a precursor:
5′-GTAGTAGCACTAAAGTGCTTATAGTGCAAGTAGTGTTTAGTTATCTACTGCATTATGAGCACTTAAAGTACTGC-3′;
and
3′-GCAGTACTTTAAGTGCTCATAATGCAGTAGATAACTAAACACTACCTGCACTATAAGCACTTTAGTGCTA-5′;
anti-miR-2 0a, 5′-CTAAACACTACCTGCACTATAAGCACTTTAGTGCTAC-3′, and
5′-GAAATGTACTGCGCGTGGAGACGTTTTGGCCACTGAC-3′. The BMSCs were seeded
into 6-well plates at 1.0–2.0×106 cells/well for 48 h in
order to perform the transfection. The cells were transfected using
Lipofectamine® 2000 with 100 ng miR-20a precursor,
anti-miR-20a, or a negative precursor (pcDNA3.1+). After
24 h, the BMSCs were treated with 0.1, 1, and 10 µM naringin
for 48 h. In all experiments untreated BMSCs were used as
controls.
Statistical analysis
All statistical analyses were performed using SPSS
17.0 (SPSS, Inc., Chicago, IL, USA). The data are presented as the
mean ± standard deviation. A χ2 and exact probability
test were performed in order to compare two groups. P<0.05 was
considered to indicate a statistically significant difference.
Results
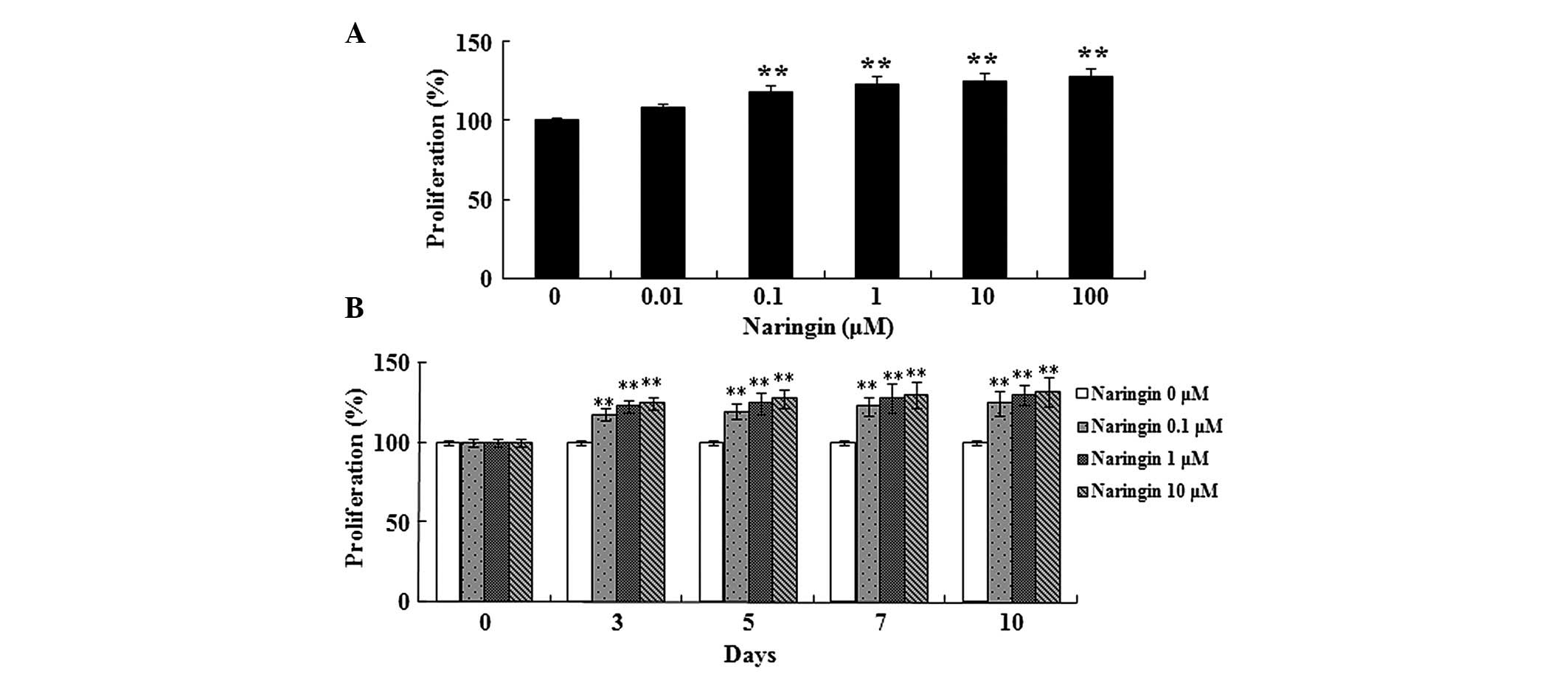
Effects of naringin on the proliferation
of BMSCs
As shown in Fig. 2,
naringin dose-dependently enhanced the proliferation of BMSCs.
Following treatment with 0.1, 1, 10 and 100 µM naringin for
48 h, BMSC proliferation was significantly enhanced (Fig. 2A). In addition, treatment with 0.1,
1, and 10 µM naringin for 3–10 days significantly enhanced
BMSC proliferation (Fig. 2B).
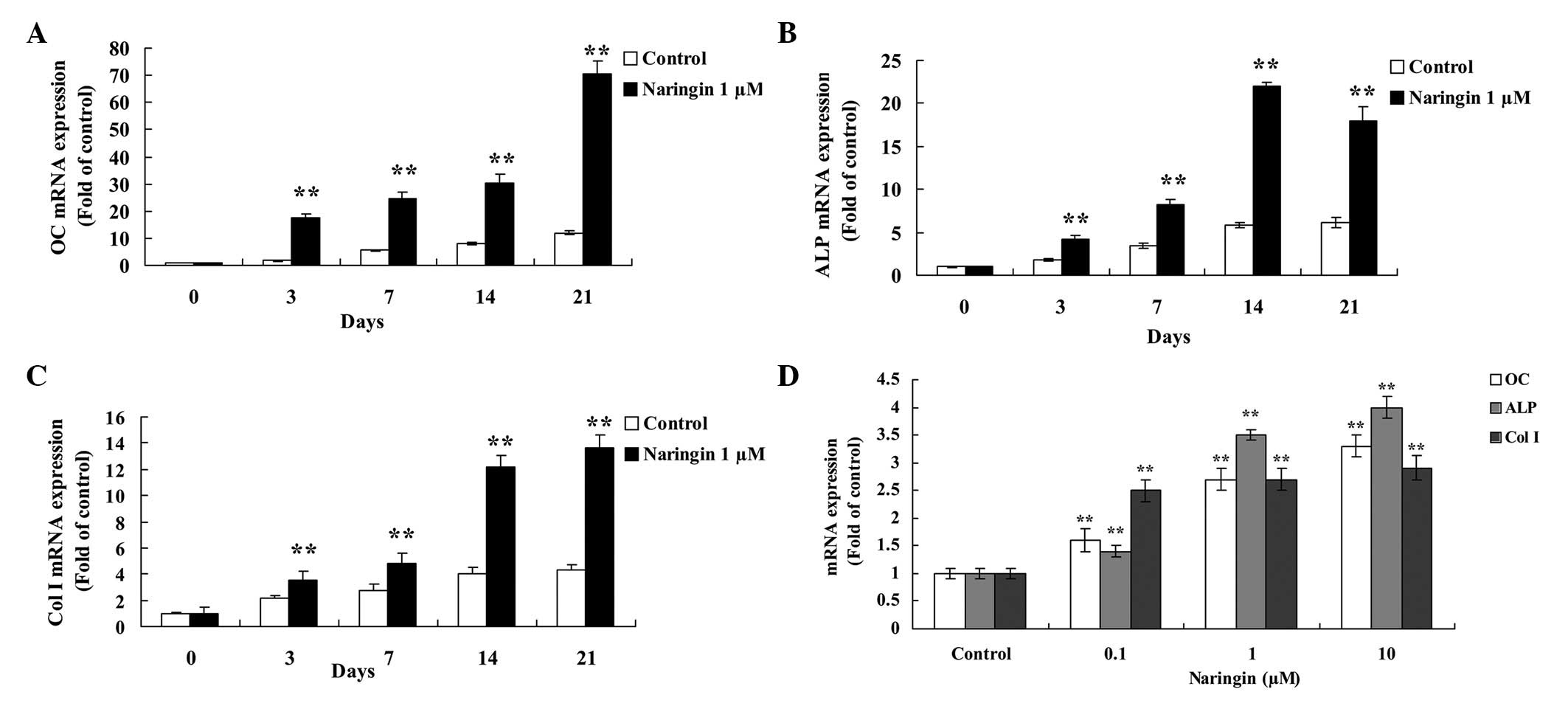
mRNA expression levels of OC, ALP, and
Col I in BMSCs
BMSCs were treated with 1 µM naringin for 3,
7, 14 and 21 days. There was a significant increase in the mRNA
expression levels of OC, ALP, and Col I in the naringin-treated
BMSCs, as compared with the control group (Fig. 3A–3C). Following treatment with 1
µM naringin for 21 days, the mRNA expression levels of OC
were the highest, with expression levels 5.8-fold greater, as
compared with that of the control cells (Fig. 3A). The mRNA expression levels of
ALP were increased by 2.3, 2.4, 3.9 and 2.9-fold following 3, 7,
14, and 21 days, respectively, as compared with the control group
(Fig. 3B). The mRNA expression
levels of Col I significantly increased between 14–21 days during
osteogenic differentiation, as compared with the control group
(Fig. 3C). In addition, BMSC
treatment with 0.1, 1, and 10 µM naringin for 48 h
significantly increased the mRNA expression levels of OC, ALP, and
Col I (Fig. 3D).
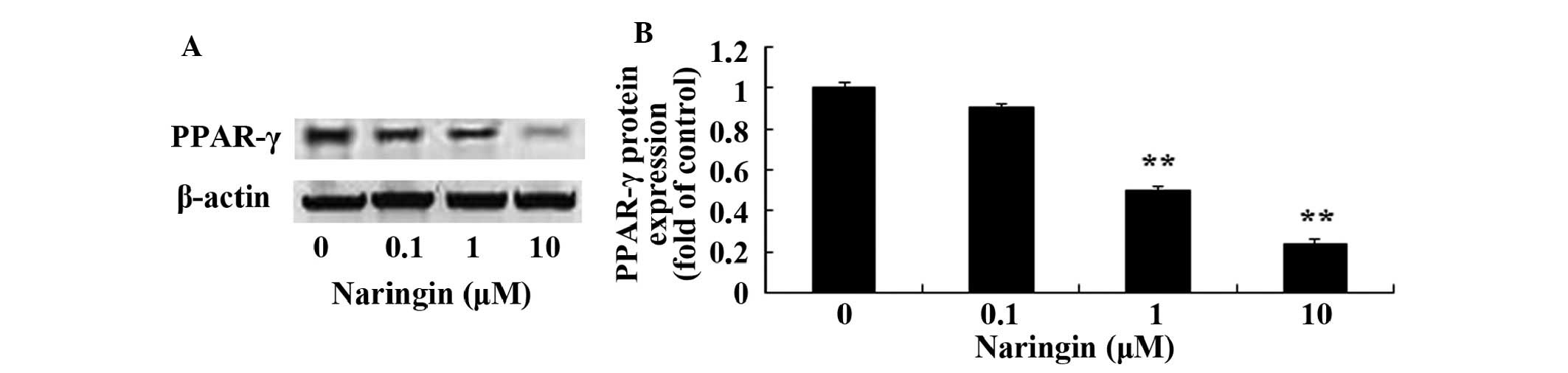
Protein expression levels of PPARγ in
BMSCs
Following treatment with 0, 0.1, 1, and 10 µM
naringin for 48 h, the protein expression levels of PPARγ were
analyzed by western blotting (Fig.
4A). Treatment with 1 and 10 µM naringin significantly
decreased the protein expression levels of PPARγ, as compared with
the control group (Fig. 4B).
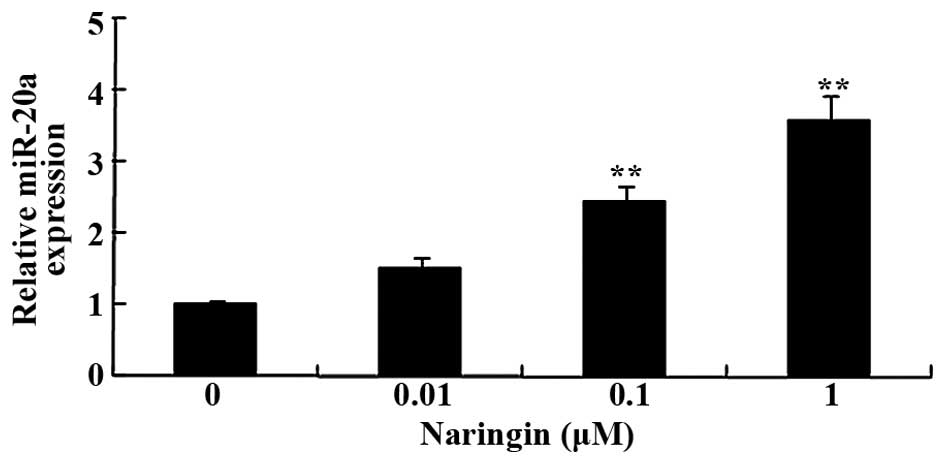
RT-qPCR analysis of miR-20a expression in
BMSCs
Following treatment with 0, 0.1, 1, and 10 µM
naringin for 48 h, the expression levels of miR-20a in the BMSCs
were analyzed by RT-qPCR. The expression levels of miR-20a were
significantly increased in the BMSCs treated with naringin (1 and
10 µM), as compared with the control group (Fig. 5).
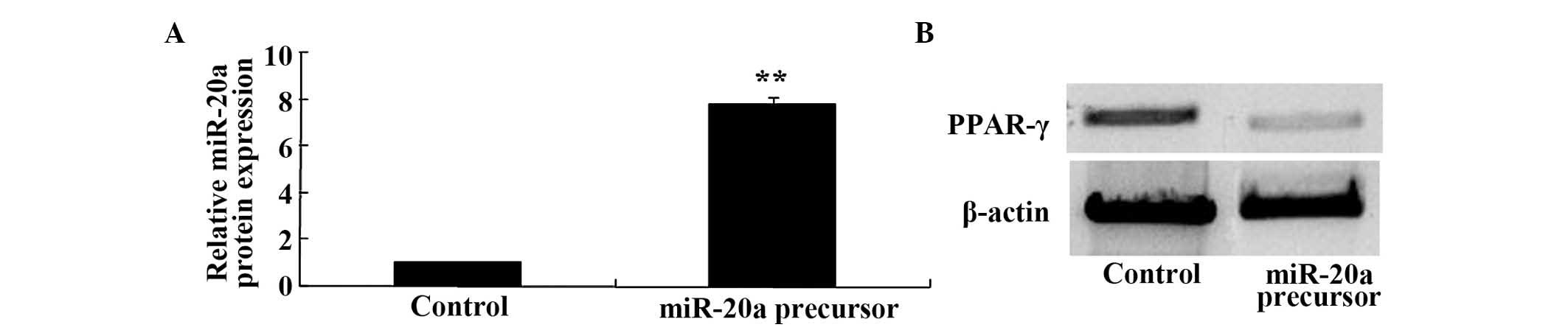
Overexpression of miR-20a suppresses the
expression levels of PPARγ in BMSCs
In order to determine whether miR-20a regulates the
protein expression of PPARγ in BMSCs, the protein expression levels
of PPARγ were analyzed in BMSCs post-transfection with a miR-20a
precursor. The effects of the miR-20a precursor on the expression
levels of miR-20a expression were analyzed by RT-qPCR. BMSC
transfection with a miR-20a precursor significantly increased the
expression levels of miR-20a in BMSCs (Fig. 6A). Conversely, the protein
expression levels of PPARγ were significantly decreased following
transfection with miR-20a (Fig.
6B).
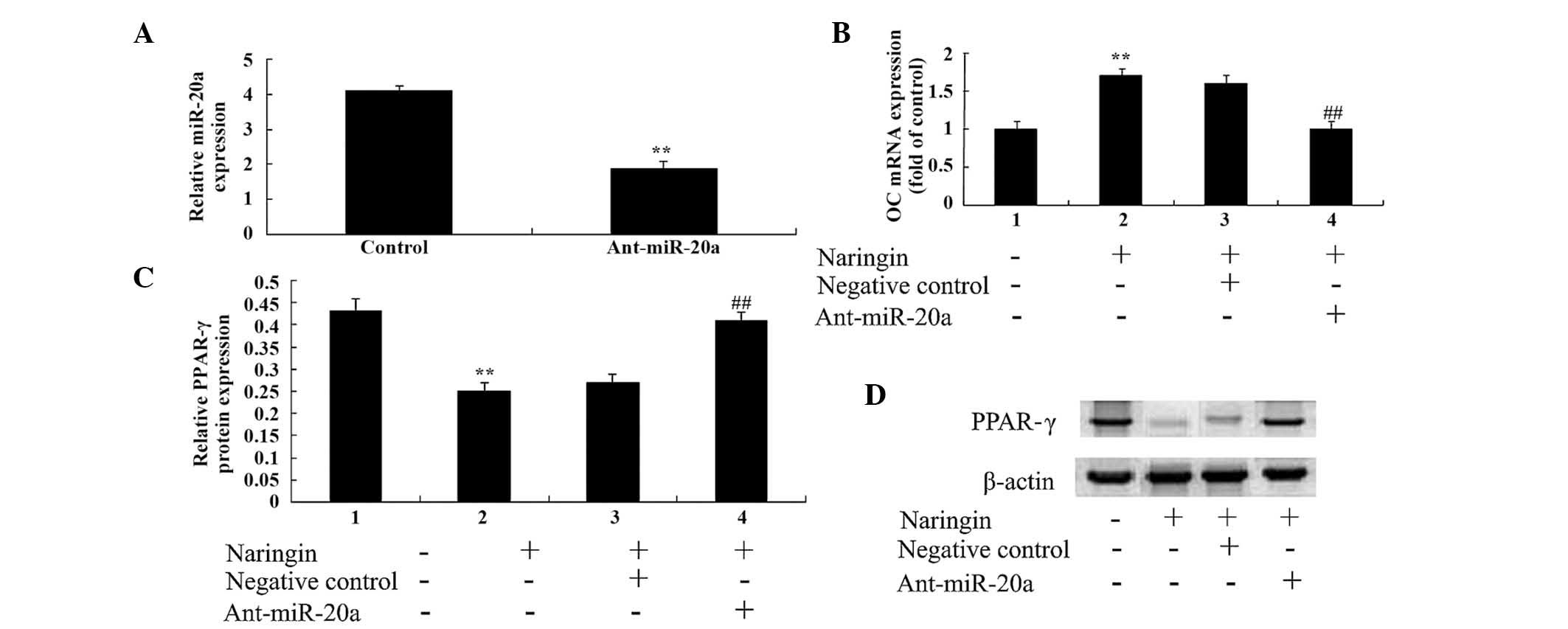
Anti-miR-20a reverses the effects of
naringin in BMSCs
Anti-miR-20a antibody was used to investigate the
effects of miR-20a on the naringin-induced inhibition of BMSCs.
BMSC transfection with anti-miR-20a antibody significantly reduced
the expression levels of miR-20a in BMSCs (Fig. 7A). In addition, BMSC transfection
with anti-miR-20a antibody also significantly reduced the effects
of 1 µM naringin on the mRNA expression levels of OC in
BMSCs (Fig. 7B), and significantly
increased the protein expression levels of PPARγ (Fig. 7C) at 48 h. These results suggest
that the anti-miR20a may neutralize the inhibitory effects of
naringin through the downregulation of miR-20a expression, and the
upregulation of PPARγ expression in BMSCs (Fig. 7D).
Discussion
BMSCs are adult stem cells located in the bone
marrow, which have high self-renewal and differentiation potentials
(18). Although BMSCs have been
isolated and cultured from numerous tissue types, such as the
liver, embryonic blood, umbilical cord blood, and amniotic fluid,
BMSCs are predominantly obtained from bone marrow tissues (19,20).
In the bone marrow, BMSCs account for 0.001%–0.1% of the total
cells, and exhibit high differentiation potential. Indeed, BMSCs
are able to differentiate into adipocytes, bone cells, cartilage
cells, and myoblasts (21). BMSCs
have been isolated from the bone marrow in numerous species,
including mice, rats, rabbits, and humans. The results of the
present study demonstrated that naringin was able to
dose-dependently enhance the proliferation of BMSCs, which prompted
the further investigation of the therapeutic effects of naringin on
BMSCs.
Osteoblasts determine not only the rate of bone
formation, they also adjust the activity levels of osteoclasts,
thereby determining the rate of bone resorption (22). Bone formation includes three
stages: Osteoblast proliferation, osteoblast maturation, and
mineralization of the extracellular matrix. Osteoblast
proliferation is the bone formation step that predominantly
determines the final amount of formed bone. ALP is an enzyme
secreted during the differentiation of osteoblasts (16). The levels of ALP are correlated
with the synthesis of Col I and with the formation of bone matrix;
ALP therefore serves as a marker for the early and mid-stages of
osteoblast differentiation (23).
In addition, the activity levels of ALP are correlated with the
activity levels of osteoblasts (24). Previous studies demonstrated that
mid to high concentrations of naringin are able to increase the
synthesis and activity levels of ALP, thereby demonstrating that
ALP promotes osteoblast differentiation and bone formation
(25,26). Naringin has previously been shown
to significantly increase the amount of bone morphogenetic
protein-2 in bone cells, and promote osteoblast differentiation
into osteogenic cells (27). The
results of the present study demonstrated that naringin is able to
significantly increase the mRNA expression levels of OC, ALP, and
Col I. The expression levels of OC following treatment with 1
µM naringin for 21 days were the highest. In addition,
treatment with 1 µM naringin significantly increased the
expression levels of ALP after 3 days. Furthermore, the mRNA
expression levels of Col I significantly increased following
treatment with 1 µM naringin between 14–21 days.
PPARγ is the primary regulator of adipocyte
differentiation, which has an important role in the regulation of
BMSC differentiation (28). A
recent study demonstrated that PPARγ has an important role in the
gene therapy of osteoporosis (29). The results of the present study
determined that treatment with 1 and 10 µM naringin for 48 h
decreased the protein expression levels of PPARγ in the BMSCs.
Furthermore, treatment with 1 and 10 µM naringin for 48 h
significantly increased the expression levels of miR-20a in the
BMSCs. The present study also investigated whether miR-20a
regulated the protein expression levels of PPARγ following
transfection of BMSCs with a miR-20a precursor or anti-miR-20a. The
results indicated that miR-20a regulated the protein expression
levels of PPARγ in BMSCs.
In conclusion, the results of the present study
revealed that naringin is able to promote BMSC differentiation into
osteoblasts, via the upregulation of miR-20a, and the
down-regulation of PPARγ. Thus suggesting that naringin may be a
potential novel drug that may promote BMSC differentiation into
osteoblasts, in the treatment of osteoporosis.
References
|
1
|
Wu G, Cui Y, Ma L, Pan X, Wang X and Zhang
B: Repairing cartilage defects with bone marrow mesenchymal stem
cells induced by CDMP and TGF-β1. Cell Tissue Bank. 15:51–57. 2014.
View Article : Google Scholar
|
|
2
|
Gong P, Wang Y, Zhang J and Wang Z: Bone
marrow mesenchymal stem cells in hepatocellular carcinoma. Front
Biosci (Landmark Ed). 18:811–819. 2013. View Article : Google Scholar
|
|
3
|
Sotiropoulou PA, Perez SA, Salagianni M,
Baxevanis CN and Papamichail M: Characterization of the optimal
culture conditions for clinical scale production of human
mesenchymal stem cells. Stem Cells. 24:462–471. 2006. View Article : Google Scholar
|
|
4
|
Hu X, Wei L, Taylor TM, et al: Hypoxic
preconditioning enhances bone marrow mesenchymal stem cell
migration via Kv2.1 channel and FAK activation. Am J Physiol Cell
Physiol. 301:C362–C372. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Pelled G, G T, Aslan H, Gazit Z and Gazit
D: Mesenchymal stem cells for bone gene therapy and tissue
engineering. Curr Pharm Des. 8:1917–1928. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Damoulis PD, Drakos DE, Gagari E and
Kaplan DL: Osteogenic differentiation of human mesenchymal bone
marrow cells in silk scaffolds is regulated by nitric oxide. Ann NY
Acad Sci. 1117:367–376. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Floyd ZE and Stephens JM: Controlling a
master switch of adipocyte development and insulin sensitivity:
Covalent modifications of PPARγ. Biochim Biophys Acta.
1822:1090–1095. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Krishnan A, Nair SA and Pillai MR: Biology
of PPAR gamma in cancer: A critical review on existing lacunae.
Curr Mol Med. 7:532–540. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Takada I, Suzawa M, Matsumoto K and Kato
S: Suppression of PPAR transactivation switches cell fate of bone
marrow stem cells from adipocytes into osteoblasts. Ann NY Acad
Sci. 1116:182–195. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Takada I and Kato S: Molecular mechanism
of switching adipocyte / osteoblast differentiation through
regulation of PPAR-gamma function. Clin Calcium. 18:656–661.
2008.In Japanese. PubMed/NCBI
|
|
11
|
Salmanidis M, Pillman K, Goodall G and
Bracken C: Direct transcriptional regulation by nuclear microRNAs.
Int J Biochem Cell Biol. 54:304–311. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Yu F, Cui Y, Zhou X, Zhang X and Han J:
Osteogenic differentiation of human ligament fibroblasts induced by
conditioned medium of osteoclast-like cells. Biosci Trends.
5:46–51. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Zhang JF, Fu WM, He ML, Xie WD, Lv Q, Wan
G, Li G, Wang H, Lu G, Hu X, et al: MiRNA-20a promotes osteogenic
differentiation of human mesenchymal stem cells by co-regulating
BMP signaling. RNA Biol. 8:829–838. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Li N, Jiang Y, Wooley PH, Xu Z and Yang
SY: Naringin promotes osteoblast differentiation and effectively
reverses ovariectomy-associated osteoporosis. J Orthop Sci.
18:478–485. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Chen LL, Lei LH, Ding PH, Tang Q and Wu
YM: Osteogenic effect of Drynariae rhizoma extracts and Naringin on
MC3T3-E1 cells and an induced rat alveolar bone resorption model.
Arch Oral Biol. 56:1655–1662. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Surampalli G, K Nanjwade B and Patil PA:
Corroboration of naringin effects on the intestinal absorption and
pharmacokinetic behavior of candesartan cilexetil solid dispersions
using in-situ rat models. Drug Dev Ind Pharm. June 11–2014.Epub
ahead of print. PubMed/NCBI
|
|
17
|
Kim HJ, Song JY, Park HJ, Park HK, Yun DH
and Chung JH: Naringin protects against rotenone-induced apoptosis
in human neuroblastoma SH-SY5Y Cells. Korean J Physiol Pharmacol.
13:281–285. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Heo JS, Choi SM, Kim HO, Kim EH, You J,
Park T, Kim E and Kim HS: Neural transdifferentiation of human bone
marrow mesenchymal stem cells on hydrophobic polymer-modified
surface and therapeutic effects in an animal model of ischemic
stroke. Neuroscience. 238:305–318. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Ge Z, Goh JC and Lee EH: The effects of
bone marrow-derived mesenchymal stem cells and fascia wrap
application to anterior cruciate ligament tissue engineering. Cell
Transplant. 14:763–773. 2005. View Article : Google Scholar
|
|
20
|
Lee OK, Kuo TK, Chen WM, Lee KD, Hsieh SL
and Chen TH: Isolation of multipotent mesenchymal stem cells from
umbilical cord blood. Blood. 103:1669–1675. 2004. View Article : Google Scholar
|
|
21
|
Mukhopadhyay A: Perspective on liver
regeneration by bone marrow-derived stem cells-a scientific
realization or a paradox. Cytotherapy. 15:881–892. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Shao J, Zhang Y, Yang T, Qi J, Zhang L and
Deng L: HIF-1alpha disturbs osteoblasts and osteoclasts coupling in
bone remodeling by up-regulating OPG expression. In Vitro Cell Dev
Biol Anim. April 10–2015.Epub ahead of print. View Article : Google Scholar
|
|
23
|
Aubin JE: Advances in the osteoblast
lineage. Biochem Cell Biol. 76:899–910. 1998. View Article : Google Scholar
|
|
24
|
Suzuki E, Ochiai-Shino H, Aoki H, et al:
Akt activation is required for TGF-beta1-induced osteoblast
differentiation of MC3T3-E1 pre-osteoblasts. PLoS One.
9:e1125662014. View Article : Google Scholar
|
|
25
|
Kumar VS, Rajmane AR, Adil M, Kandhare AD,
Ghosh P and Bodhankar SL: Naringin ameliorates acetic acid induced
colitis through modulation of endogenous oxido-nitrosative balance
and DNA damage in rats. J Biomed Res. 28:132–145. 2014.PubMed/NCBI
|
|
26
|
Li L, Zeng Z and Cai G: Comparison of
neoeriocitrin and naringin on proliferation and osteogenic
differentiation in MC3T3-E1. Phytomedicine. 18:985–989. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Wu JB, Fong YC, Tsai HY, Chen YF, Tsuzuki
M and Tang CH: Naringin-induced bone morphogenetic protein-2
expression via PI3K, Akt, c-Fos/c-Jun and AP-1 pathway in
osteoblasts. Eur J Pharmacol. 588:333–341. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Sun J, Wang Y, Li Y and Zhao G:
Downregulation of PPARγ by miR-548d-5p suppresses the adipogenic
differentiation of human bone marrow mesenchymal stem cells and
enhances their osteogenic potential. J Transl Med. 12:1682014.
View Article : Google Scholar
|
|
29
|
Harsløf T, Tofteng CL, Husted LB, Nyegaard
M, Børglum A, Carstens M, Stenkjær L, Brixen K, Eiken P, Jensen JE,
et al: Polymorphisms of the peroxisome proliferator-activated
receptor gamma (PPARγ) gene are associated with osteoporosis.
Osteoporos Int. 22:2655–2666. 2011. View Article : Google Scholar
|