Introduction
Diabetic nephropathy (DN) is a serious and common
complication of diabetes type I and II, leading to end-stage renal
disease (ESRD) (1,2). Accumulating evidence suggested that
glomerular podocytes have a pivotal role in the pathogenesis of
diabetic kidney disease (3,4).
Podocytes are terminally differentiated cells residing on the outer
surface of the glomerular basement membrane (GBM) and have a key
role in maintaining the structure and function of the glomerular
filtration barrier (3). Previous
studies have demonstrated podocyte depletion and loss in the early
stages of DN (5,6). These stages of podocyte depletion are
accompanied by corresponding degrees of proteinuria (7,8).
Therefore, establishment of novel and innovative therapeutic
strategies targeted to block the shedding of podocytes, decrease
proteinuria and delay the progression of DN has become increasingly
urgent.
Integrins are heterodimeric transmembrane adhesion
receptors composed of α- and β-subunits. Binding of extracellular
matrix molecules or other ligands to the extracellular domain of
integrins delivers a variety of signals into the cell. Integrins
have key roles in a number of important biological processes,
including migration, survival, proliferation, gene expression and
receptor tyrosine kinase signaling (9,10).
Integrin α3β1 is the principal adhesion complex which is
responsible for the attachment of podocytes to the GBM. An in
vitro study has demonstrated that high-glucose conditions
decreased α3β1 integrin expression in rat and human podocytes
(11). Genetic ablation of β1
integrin in mice was shown to cause embryonic lethality shortly
after implantation (12). Mice
featuring knockout of podocyte-specific Itgb1, which encodes
β1 integrin, were shown to develop a similar phenotype to
podocyte-specific Itga3-knockout mice, with massive
proteinuria soon after birth, a laminated GBM with extensive
splitting, and foot process effacement followed by mortality within
1–5 weeks due to ESRD (13.14). Besides β1 integrins, αvβ3 integrin
is highly expressed in glumerular podocytes (15,16).
In mice and humans, activation of αvβ3 by the (soluble) urokinase
receptor (uPAR) was demonstrated to result in foot process
effacement, proteinuria and focal segmental glomerulosclerosis
(15,17). Inhibition of αvβ3 with an anti-β3
antibody or the small-molecule inhibitor cilengitide alleviated
proteinuria induced by urokinase receptor (15,17).
It has been suggested that aldosterone has an
important role in the pathogenesis of DN (18). Podocytes are one type of target
cells for the deleterious effects of aldosterone (19). Spironolactone (SPL), a
non-selective aldosterone receptor blocker, was reported to exert
beneficial effects not only by reducing proteinuria but delaying DN
progression (20,21). Similarly, a previous study
demonstrated that SPL prevents podocytic adhesion in
streptozotocin-induced diabetic rats, decreases urine albumin and
podocyte levels, and upregulates integrin α3 expression (22). However, whether spironolactone has
an effect on β1 and β3 integrin has remained elusive.
Based on the abovementioned findings, the present
study was designed to observe whether spironolactone has an impact
on integrin β1 and β3 expression, and podocyte motility under in
vitro diabetic conditions.
Materials and methods
Drugs and reagents
Spironolactone (S3378), d-glucose (G7021) and mannitol
(M9647) were purchased from Sigma-Aldrich (St. Louis, MO, USA).
Dulbecco's modified Eagle's medium (DMEM) was obtained from
Corning, Inc. (Corning, NY, USA). Fetal bovine serum (FBS),
recombinant interferon γ and rat tail collagen type I were
purchased from Invitrogen Life Technologies, Inc., (Carlsbad, CA,
USA), ProSpec Tany Technogene Ltd. (East Brunswick, NJ, USA) and BD
Biosciences (Franklin Lakes, NJ, USA), respectively. All primary
antibodies, including rabbit polyclonal antibody integrin β1 (cat.
no. sc-8978), rabbit polyclonal anti-integrin β3 (cat. no.
sc-14009), goat polyclonal anti-synaptopodin (N-14; cat. no.
sc-21536) and mouse monoclonal anti-GAPDH (cat. no. sc-365062),
were purchased from Santa Cruz Biotechnology, Inc. (Dallas, TX,
USA). Fluorescein isothiocyanate-conjugated (FITC)-donkey anti-goat
immunoglobulin (Ig)G (H+L; cat. no. sc-2024) and goat anti-rabbit
Alexa Fluor 555 (cat. no. sc-362272) were obtained from Santa Cruz
Biotechnology, Inc., and Cell Signaling Technology, Inc. (Beverly,
MA, USA), respectively. Dimethyl sulfoxide (DMSO) and TRIzol were
purchased from Sigma-Aldrich (St. Louis, MO, USA) and Invitrogen
Life Technologies, Inc, respectively. PrimeScript 1st Strand cDNA
Synthesis kit (cat. no. RR001A) and SYBR Premix Ex Taq Green kit
(cat. no. RR420A) were purchased from Takara Biotechnology Inc.
(Dalian, China). A radioimmunoprecipitation assay (RIPA) protein
extraction kit (cat. no. C1053) and bicinchoninic acid (BCA)
Protein Assay kit (cat. no. FD2001) were purchased from Puli Lai
company (Beijing, China) and Fabio Science company (Hangzhou,
China), respectively.
Cell culture and treatment
The conditionally immortalized mouse podocyte cell
line (MPC) was kindly provided by Dr. Jochen Reiser (Rush
University Medical Center, Chicago, IL, USA) and were cultured as
previously described (23).
Differentiated podocytes (passage 13–18) were serum-starved for 24
h and then treated with normal glucose (NG; 5.3 mM), high glucose
(HG; 20 mM) or with NG (5.3 mM) plus mannitol (14.7 mM; osmolality
control) for 48 h prior to the assays. For intervention
experiments, aldosterone receptor blocker SPL (Sigma-Aldrich) at
concentrations of 10−8 mol/l and 10−7 mol/l
was respectively added to cells treated with HG at a concentration
of 20 mM for 48 h.
Reverse transcription quantitative
polymerase chain reaction (RT-qPCR)
TRIzol reagent was used to extract the total RNA
from the cultured podocytes from the experimental groups according
to the manufacturer's instructions (Invitrogen Life Technologies,
Inc.). The complementary DNA was synthesized using 1,000 ng RNA in
20 µl using the PrimerScript™ RT Regent kit (cat. no.
RR001A; Takara Biotechnology Inc.) according to the manufacturer's
instructions. RT-PCR was performed with a Bio-Rad CFX96 Touch q-PCR
system (Bio-Rad Laboratories, Inc., Hercules, CA, USA) using Power
SYBR Green PCR Master Mix with the cDNA and primer pairs
(Invitrogen Life Technologies). The sequences of primer pairs used
for real-time PCR were as follows: Integrin β1 forward,
5′-GGCTGAAGATTACCCTAT-3′ and reverse, 5′-CATTCATCAAATCCGTTC-3′;
integrin β3 forward, 5′-GCCTTCGTGGACAAGCCTGTA-3′ and reverse,
5′-GGACAATGCCTGCCAGTCTTC-3′; GAPDH forward,
5′-TGTGTCCGTCGTGGATCTGA-3′ and reverse,
5′-TTGCTGTTGAAGTCGCAGGAG-3′. The PCR conditions were as follows: An
initial step of 2 min at 95°C, followed by 40 cycles of 5 sec at
95°C, 25 sec at 60°C, and 40 sec at 72°C. Every reaction was
amplified in triplicate and the fold change in the expression of
each gene was calculated using ΔΔCt method with GAPDH mRNA as an
internal control.
Immunofluorescence staining
After being subjected to various treatments,
podocytes were fixed with 4% paraformaldehyde at −20°C for 20 min
and then incubated with 5% BSA for 20 min at RT to block
non-specific binding. Samples were incubated with the primary
antibodies overnight at 4°C, followed by 1 h of incubation with
secondary antibodies goat anti-rabbit Alexa Fluor 555 (1:1,000) or
FITC-donkey anti-goat IgG (H+L) (1:250) at room temperature,
followed by counter-staining with DAPI (Roche Diagnostics, Basel,
Switzerland) for 5 min to visualize the nuclei. After being washed,
the slides were mounted with anti-fade mounting medium (Beyotime
Institute of Biotechnology, Haimen, China). Photomicrographs were
captured using confocal microscopy (Leica SP5-FCS; Leica
Microsystems, Oberkochen, Germany). The primary antibodies used in
the present study were as follows: Rabbit polyclonal anti-integrin
β1 (1:100), rabbit polyclonal anti-integrin β3 (1:100) and goat
polyclonal anti-synaptopodin (N-14) (1:100). All images were
analyzed by two investigators blinded to the identity of the
samples.
Western blot analysis
Differentiated podocytes, subjected to various
experimental conditions, were washed twice with cold
phosphate-buffered saline (PBS). The washed podocytes were drained
and scraped with RIPA lysis buffer using a cold plastic cell
scraper. Protein concentration was quantified using the BCA Protein
Assay Reagent kit. An aliquot of cell lysates containing 30
µg protein was separated by 8% SDS-PAGE and transferred onto
a polyvinylidene fluoride membrane (Millipore, Billerica, MA, USA).
Membranes were blocked with 5% fat-free milk for 1 h at room
temperature and then incubated overnight at 4°C with the following
primary antibodies: Rabbit polyclonal anti-integrin β1 (1:1,000),
rabbit polyclonal anti-integrin β3 (1:1,000) and mouse monoclonal
anti-GAPDH (1:2,000). After washing, HRP-conjugated goat
anti-rabbit IgG or goat anti-mouse IgG (Jackson Immuno Research,
West Grove, PA, USA, 1:5,000) was added and incubated 1 h at room
temperature. The immunoblots were washed three times with
Tris-buffered staline containing Tween 20 and immersed in ECL Plus
Western Blotting Detection Reagents (Beyotime Institute of
Biotechnology). Results were analyzed using Image J v1.47 software
(National Institutes of Health, Bethesda, MD, USA) and normalized
to the protein expression of GAPDH.
Wound healing assay
Cultured differentiated podocytes
(1×105/ml) were seeded overnight on vitronectin-coated
coverslips in six-well plates. Each coverslip was then scratched
with a sterile 200-µl pipette tip, washed with PBS and
placed into fresh medium. After 24 h, cells were fixed with cold
methanol, permeabilized with 0.5% Triton X-100 in PBS and cell
nuclei were stained with DAPI (Roche Diagnostics). Images were
captured by phase-contrast microscopy under a ×10 objective on a
Leica SP5-FCS microscope (Leica Microsystems) at 0 and 24 h after
scratching, and the numbers of cells that had migrated into the
same-sized square fields were counted. Results are presented
presented as the mean ± standard deviation (SD) of six independent
experiments.
Statistical analysis
Values are expressed as the mean ± SD. Statistical
analysis was performed by SPSS 17.0 (SPSS Inc, Chicago, IL,USA)
using analysis of variance followed by Bonferoni's multiple
comparisons test. P<0.05 was considered to indicate a
significant difference between values.
Results
HG decreases integrin β1 and increases
integrin β3 expression in cultured podocytes
To determine the effects of HG on integrin β1 and
integrin β3, podocytes were cultured in DMEM containing 5.3 mM
glucose (NG group), 5.3 mM glucose plus 14.7 mM mannitol (M group,
as an osmolality control) or 20 mM glucose (HG group) for 48 h. The
mRNA and protein expression of integrin β1 and integrin β3 were
determined using RT-qPCR and western blot analysis. As shown in
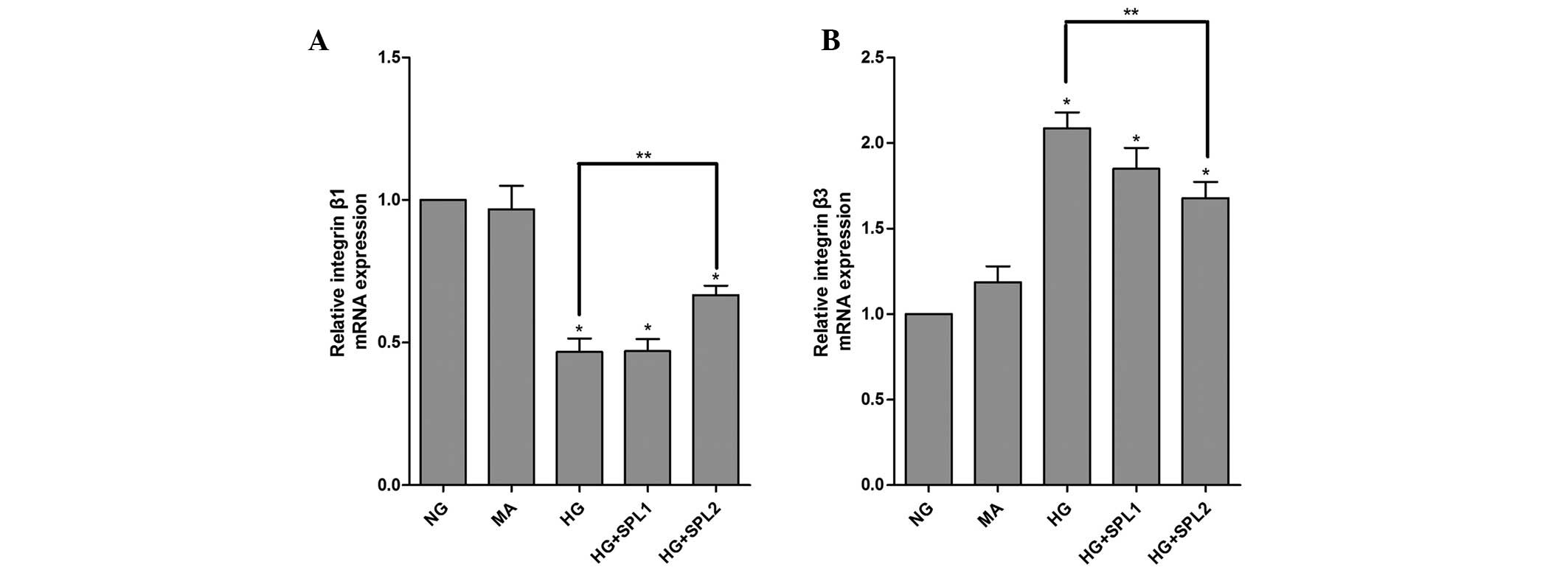
Fig. 1A, the mRNA expression of
integrin β1 was significantly decreased in the HG group compared to
that in the NG group (P<0.001). By contrast, the mRNA levels of
integrin β3 were markedly increased in podocytes incubated with HG
for 48 h (Fig. 1B).
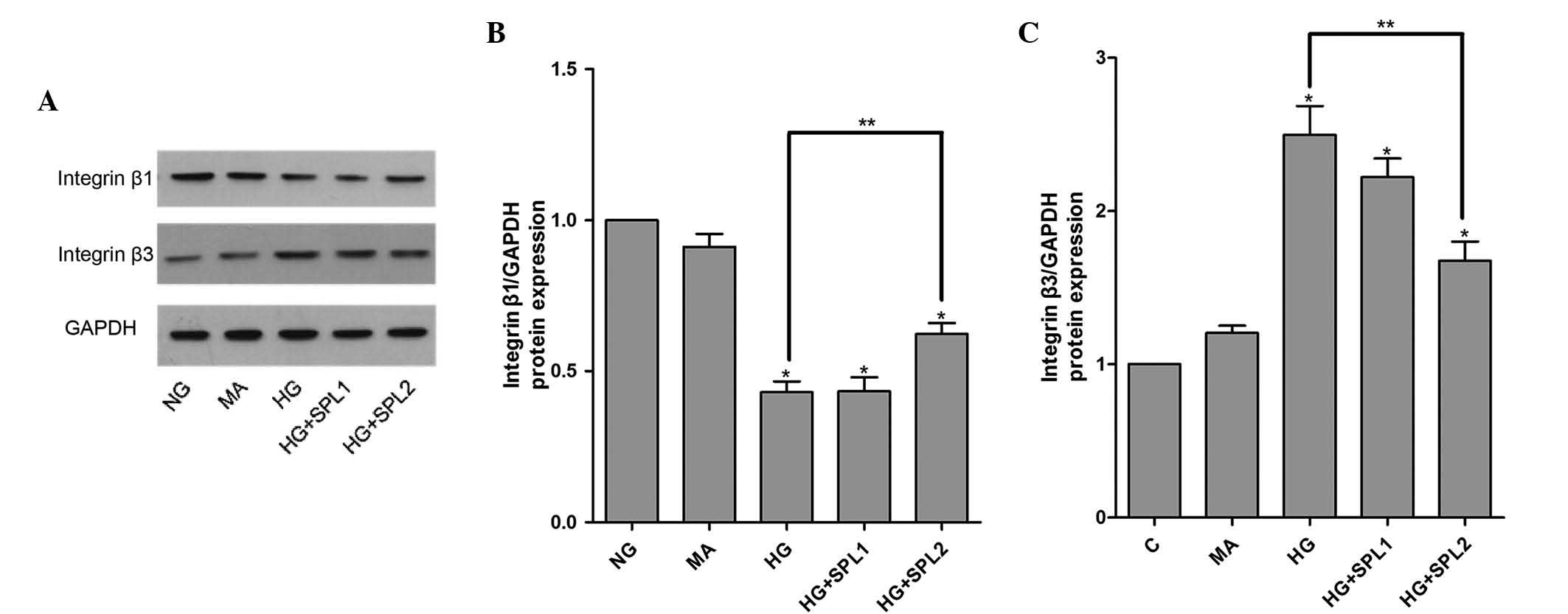
Immunofluorescent staining and western blot analysis also showed a
decreased protein expression of integrin β1 and an increased
protein expression of integrin β3 in the HG group (Figs. 2 and 3). As expected, mannitol had no effect on
the mRNA expression of integrin β1 and integrin β3, suggesting that
the decreased integrin β1 and β3 mRNA expression under HG did not
result from high osmolality.
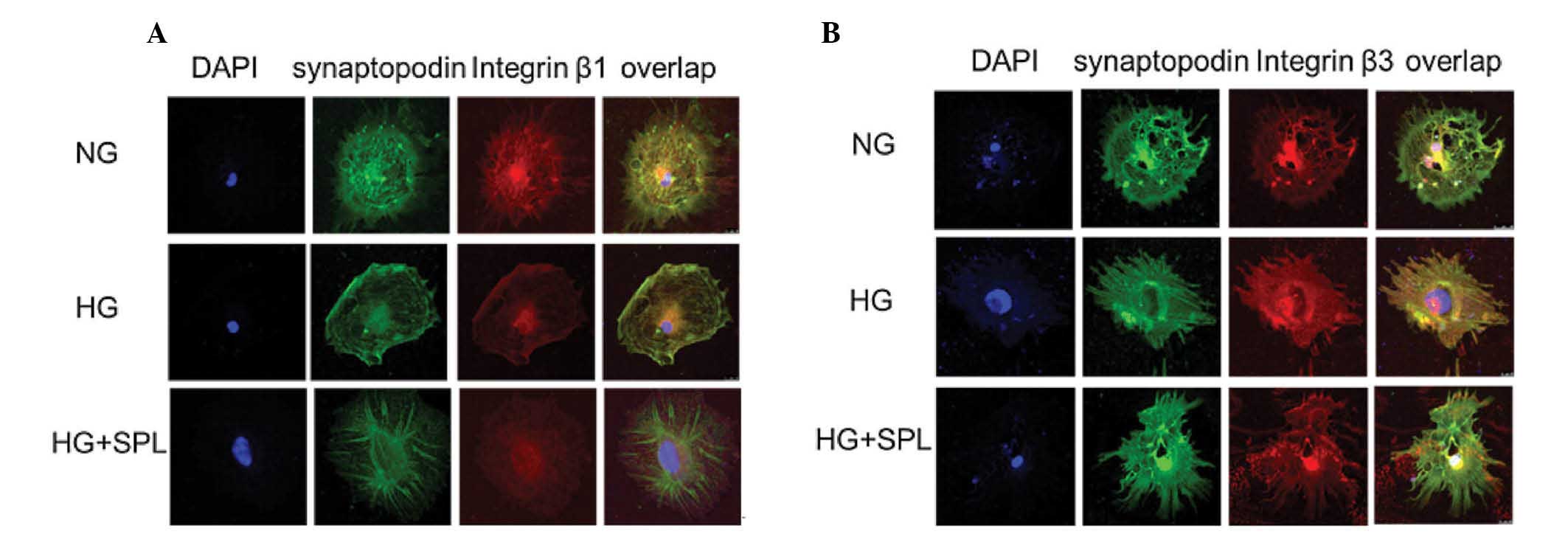 | Figure 2Effects of spironolactone on integrin
β1 and β3 expression were analyzed by immunofluorescent staining in
podocytes under HG conditions. (A) Double immunofluorescent
staining of integrin β1 (red), synaptopodin (green), DAPI-stained
nuclei (blue) and merged images in cultured podocytes treated with
NG, HG and HG plus SPL for 48 h, respectively. (B) Double
immunofluorescent staining of integrin β3 (red), synaptopodin
(green), DAPI-stained nuclei (blue) and merged images in cultured
podocytes treated with NG, HG and HG plus SPL for 48 h,
respectively. NG, normal glucose (5.3 Mm); HG, high glucose (20
mM); HG + SPL, high glucose (20 mM) + spironolactone
(10−7 mol/l). |
SPL normalizes integrin β1 and integrin
β3 expression in podocytes under HG
To evaluate the effects of SPL on the expression of
integrin β1 and integrin β3 in HG-cultured podocytes, cells
incubated under various conditions were analyzed using RT-qPCR,
immunoblotting and immunofluorescence. As shown in Fig. 1A, the HG-mediated decrease in
integrin β1 mRNA expression in podocytes was significantly
attenuated by SPL (10−7 mol/l) (P<0.01). Similarly,
immunofluorescence and western blot analysis demonstrated that
decreases in integrin β1 protein expression in HG-cultured
podocytes were restored by SPL (10−7 mol/l) (Fig. 2A, and 3A and B). Furthermore, the significant
HG-induced increase in the expression of integrin β3 was reduced by
treatment with SPL (10−7 mol/l) at the mRNA level
(Fig. 1B) and at the protein level
(Figs. 2B, and 3A and C). However, a low dose of SPL
(10−8 mol/l) did not affect integrin β1 and integrin β3
expression in podocytes cultured under HG (P>0.05) (Figs. 1 and 3).
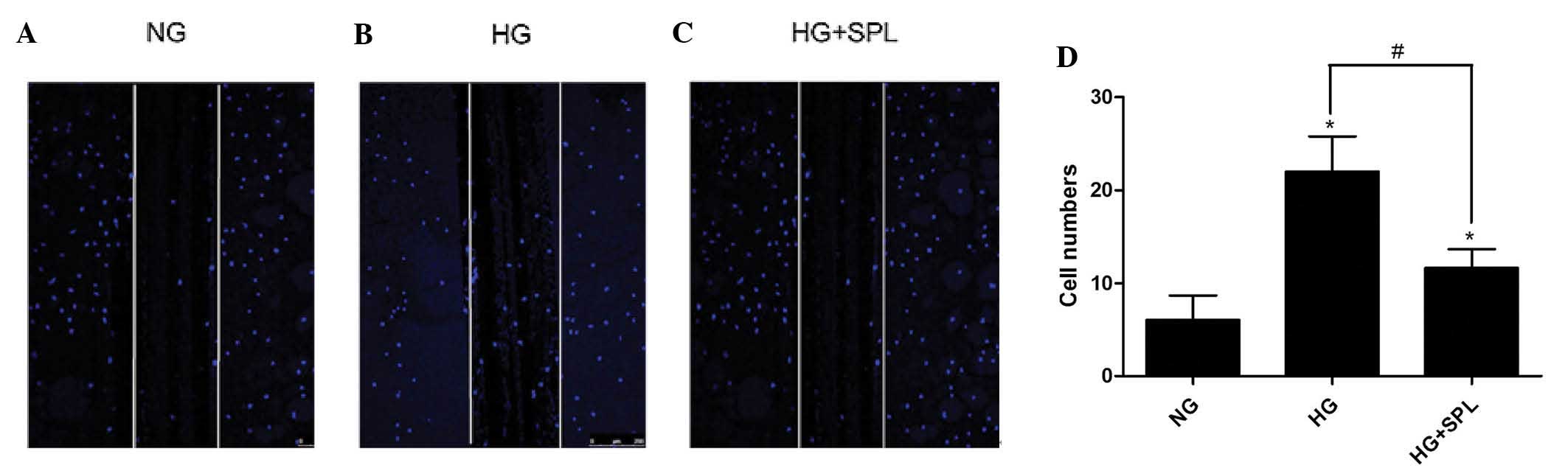
SPL inhibits podocyte motility induced by
HG
Podocyte motility is regarded as a surrogate
indicator for proteinuria and effacement of podocyte foot processes
in vivo (15,17,24).
Therefore, the present study next explored whether SPL has a role
in inhibiting cell motility of podocytes in vitro. The
effects of SPL on the spatial motility of podocytes was analyzed
using a scrape-wound assay (Fig.
4). As compared with the control (6.0±3.0), HG treatment
significantly promoted podocyte wound closure (22.0±4.0; P<0.05)
(Fig. 4A, B and D). By contrast,
treatment with SPL reduced HG-induced podocyte motility (12.0±2.0;
P<0.05) (Fig. 4C and D). These
results demonstrated that SPL inhibits podocyte motility induced by
HG.
Discussion
The incidence of diabetes has been increasing on a
yearly basis; in addition, DN has gradually become a severe
complication in patients with diabetes (1,2).
Experimental and clinical studies have shown that a decrease in the
number of podocytes due to shedding or apoptosis leads to
proteinuria in DN (5,6,25).
Apart from the administration of angiotensin-converting enzyme
inhibitors, angiotensin receptor blockers and other drugs, no other
treatments for effectively lowering urinary protein and delaying
disease progression are currently available. As a novel approach
for the development of therapies for the treatment of DN, the
prevention or inhibition of podocyte shedding or apoptosis is an
obvious and promising therapeutic target.
Under normal circumstances, podocytes are anchored
to the GBM through the α3β1-integrin complex that is present in the
sole of the foot processes. However, under pathological conditions,
resembled by a rat model of diabetes or podocytes cultured under
HG, podocyte shedding or podocye motility were increased,
accompanied by changes in the levels of integrin, which was
observed at early stages of DN (5,6,26).
Therefore, blocking this process by increasing the levels of
integrin β1 is expected to enable podocytes to closely attach to
the GBM. In addition, a recent study by our group showed that uPAR
expression was increased in HG-cultured podocytes and DN patients
(27). Previous studies have
demonstrated that uPAR and integrin β3 co-localize in podocytes and
form a lipid-dependent complex with integrin β3, thereby causing a
structural configurational change of integrin β3, resulting in its
activation with increased affinity for ligand binding (5,6).
In vivo gene delivery of a structurally fixed and
constitutively active integrin β3 was shown to be sufficient to
induce proteinuria in mice; conversely, the inhibition of uPAR
expression and integrin β3 activation improved the structure of the
podocyte foot process and had an anti-proteinuric effect (24). Therefore, the present study
addressed the question whether decreased expression of integrin β3
by drug intervention under diabetic conditions can inhibit podocyte
motility.
The results of the present study demonstrated that
HG decreased integrin β1 expression and increased integrin β3
expression, accompanied by increased podocyte motility. In
HG-cultured podocytes treated with various concentrations of SPL, a
non-selective aldosterone receptor antagonist, changes in integrin
β1 and integrin β3 expressions were partially normalized by high
concentrations of SPL (10−7 mol/l). HG-induced decreases
in integrin β1 expression increases in integrin β3 expression and
increases in podocyte motility were significantly attenuated by
SPL. However, a low concentration of SPL (10−8 mol/l)
had no marked effects on HG-induced changes in integrin expression
and podocyte motility. Whether the attenuating effects of SPL on
the HG-induced effects on podocytes are based on respective
interactions with the two integrin β sub-units or on mutual
reactions between integrin β1 and integrin β3 themselves remains
elusive. A cancer-associated study reported that integrin α3β1
inhibits integrin αvβ3 expression (28). Hence, it was hypothesized in the
present study that, under physiological conditions, integrin β1
expression in podocytes may inhibit the expression of integrin β3.
However, under pathological conditions, the role of inhibited
integrin β3 is expected to be attenuated due to decreased levels of
integrin β1. In the present study, decreases in integrin β1 and
increases in integrin β3 expression were attenuated by
co-incubation with SPL in HG-cultured podocytes.
Recent clinical and experimental studies have
demonstrated that aldosterone has pathogenetic roles in podocyte
injury and DN (18,19,29).
Aldosterone is a potent inducer of proteinuria. Siragy and Xue
(30) demonstrated that diabetes
increased local aldosterone production in the kidney, which
contributed to the development of renal inflammation, matrix
formation and albuminuria. Another previous study also showed that
the local aldosterone system is activated and is involved in
podocyte apoptosis under diabetic conditions (31). These results suggested that
blockade of the aldosterone system may represent a novel
therapeutic strategy to prevent proteinuria and podocyte injury
under hyperglycemic conditions. In fact, experimental as well as
clinical studies have demonstrated renoprotective effects of SPL in
DN (20,21,32–37).
However, the mechanisms by which SPL attenuates proteinuria and
podocyte injury in DN remain to be fully elucidated. The results of
the present study revealed that under HG conditions, the expression
of integrin β1 was increased and integrin β3 expression was
decreased by SPL, accompanied with the inhibition of podocyte
motility. This effect may be one of the possible mechanisms of the
protective effects of SPL on podocyte injury.
In conclusion, the present study showed that HG
conditions decreased integrin β1 and increased integrin β3
expression in podocytes, accompanied with enhanced podocyte
motility. Treatment with SPL markedly inhibited podocyte motility
and partly restored integrin β1 and integrin β3 expression in this
cell model. These results indicated that the effects of SPL on
podocyte motility may, proceed via restoring integrin β1 and β3
expression, which may be one of the underlying mechanisms of its
protective effects against podocyte injury under HG conditions.
Acknowledgments
This study was supported by the National Natural
Science Foundation (nos. 81170683, 81270784, S2013040015908 and
81470930) and National Clinical Key Specialty Construction
Projects.
References
|
1
|
Molitch ME, DeFronzo RA, Franz MJ, Keane
WF, Mogensen CE, Parving HH and Steffes MW; American Diabetes
Association: Nephropathy in diabetes. Diabetes Care. 27(Suppl 1):
S79–S83. 2004. View Article : Google Scholar
|
|
2
|
Ritz E, Rychlik I, Locatelli F and Halimi
S: End-stage renal failure in type 2 diabetes: A medical
catastrophe of worldwide dimensions. Am J Kidney Dis. 34:795–808.
1999. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Wolf G, Chen S and Ziyadeh FN: From the
periphery of the glomerular capillary wall toward the center of
disease: Podocyte injury comes of age in diabetic nephropathy.
Diabetes. 54:1626–1634. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Li JJ, Kwak SJ, Jung DS, Kim JJ, Yoo TH,
Ryu DR, Han SH, Choi HY, Lee JE, Moon SJ, et al: Podocyte biology
in diabetic nephropathy. Kidney Int Suppl. S36–S42. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Pagtalunan ME, Miller PL, Jumping-Eagle S,
Nelson RG, Myers BD, Rennke HG, Coplon NS, Sun L and Meyer TW:
Podocyte loss and progressive glomerular injury in type II
diabetes. J Clin Invest. 99:342–348. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
White KE, Bilous RW, Marshall SM, El Nahas
M, Remuzzi G, Piras G, De Cosmo S and Viberti G: Podocyte number in
normotensive type 1 diabetic patients with albuminuria. Diabetes.
51:3083–3089. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
D'Agati VD: Podocyte injury in focal
segmental glomerulo-sclerosis: Lessons from animal models (a play
in five acts). Kidney Int. 73:399–406. 2008. View Article : Google Scholar
|
|
8
|
Wharram BL, Goyal M, Wiggins JE, Sanden
SK, Hussain S, Filipiak WE, Saunders TL, Dysko RC, Kohno K, Hozman
LB and Wiggins RC: Podocyte depletion causes glomerulosclerosis:
Diphtheria toxin-induced podocyte depletion in rats expressing
human diphtheria toxin receptor transgene. J Am Soc Nephrol.
16:2941–2952. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Wu X, Wang J, Jiang H, Hu Q, Chen J, Zhang
J, Zhu R, Liu W and Li B: Wnt3a activates β1-integrin and regulates
migration and adhesion of vascular smooth muscle cells. Mol Med
Rep. 9:1159–1164. 2014.PubMed/NCBI
|
|
10
|
Zaidel-Bar R, Itzkovitz S, Ma'ayan A,
Iyengar R and Geiger B: Functional atlas of the integrin adhesome.
Nat Cell Biol. 9:858–867. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Kitsiou PV, Tzinia AK, Stetler-Stevenson
WG, Michael AF, Fan WW, Zhou B and Tsilibary EC: Glucose-induced
changes in integrins and matrix-related functions in cultured human
glomerular epithelial cells. Am J Physiol Renal Physiol.
284:F671–F679. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Fassler R and Meyer M: Consequences of
lack of beta 1 integrin gene expression in mice. Genes Dev.
9:1896–1908. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Kanasaki K, Kanda Y, Palmsten K, Tanjore
H, Lee SB, Lebleu VS, Gattone VH Jr and Kalluri R: Integrin
beta1-mediated matrix assembly and signaling are critical for the
normal development and function of the kidney glomerulus. Dev Biol.
313:584–593. 2008. View Article : Google Scholar
|
|
14
|
Pozzi A, Jarad G, Moeckel GW, Coffa S,
Zhang X, Gewin L, Eremina V, Hudson BG, Borza B, Harris RC, et al:
Beta1 integrin expression by podocytes is required to maintain
glomerular structural integrity. De Biol. 316:288–301. 2008.
|
|
15
|
Wei C, Möller CC, Altintas MM, Li J,
Schwarz K, Zacchigna S, Xie L, Henger A, Schmid H, Rastaldi MP, et
al: Modification of kidney barrier function by the urokinase
receptor. Nat Med. 14:55–63. 2008. View
Article : Google Scholar
|
|
16
|
Sachs N and Sonnenberg A: Cell-matrix
adhesion of podocytes in physiology and disease. Nat Rev Nephrol.
9:200–210. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Wei C, El Hindi S, Li J, Fornoni A, Goes
N, Sageshima J, Maiguel D, Karumanchi SA, Yap HK, Saleem M, et al:
Circulating urokinase receptor as a cause of focal segmental
glomerulosclerosis. Nat Med. 17:952–960. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Cha DR, Kang YS, Han SY, Jee YH, Han KH,
Kim HK, Han JY and Kim YS: Role of aldosterone in diabetic
nephropathy. Nephrology (Carlton). 10(Suppl): S37–S39. 2005.
View Article : Google Scholar
|
|
19
|
Shibata S, Nagase M, Yoshida S, Kawachi H
and Fujita T: Podocyte as the target for aldosterone: Roles of
oxidative stress and Sgk1. Hypertension. 49:355–364. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Schjoedt KJ, Rossing K, Juhl TR, Boomsma
F, Tarnow L, Rossing P and Parving HH: Beneficial impact of
spironolactone on nephrotic range albuminuria in diabetic
nephropathy. Kidney Int. 70:536–542. 2006.PubMed/NCBI
|
|
21
|
Aguilar C and Rodríguez-Delfín L: Effects
of spironolactone administration on the podocytes loss and
progression of experimental diabetic nephropathy. Rev Peru Med Exp
Salud Publica. 29:490–497. 2012.In Spanish. View Article : Google Scholar
|
|
22
|
Lin S, Li D, Jia J, Zheng Z, Jia Z and
Shang W: Spironolactone ameliorates podocytic adhesive capacity via
restoring integrin alpha 3 expression in streptozotocin-induced
diabetic rats. J Renin Angiotensin Aldosterone Syst. 11:149–157.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Mundel P, Reiser J, Zúñiga Mejía Borja A,
Pavenstadt H, Davidson GR, Kriz W and Zeller R: Rearrangements of
the cytoskeleton and cell contacts induce process formation during
differentiation of conditionally immortalized mouse podocyte cell
lines. Exp Cell Res. 236:248–258. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Zhang B, Xie S, Shi W and Yang Y:
Amiloride off-target effect inhibits podocyte urokinase receptor
expression and reduces proteinuria. Nephrol Dial Transplant.
27:1746–1755. 2012. View Article : Google Scholar
|
|
25
|
Weil EJ, Lemley KV, Yee B, Lovato T,
Richardson M, Myers BD and Nelson RG: Podocyte detachment in type 2
diabetic nephropathy. Am J Nephrol. 33(Suppl 1): 21–24. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Kitsiou PV, Tzinia AK, Stetler-Stevenson
WG, Michael AF, Fan WW, Zhou B and Tsilibary EC: Glucose-induced
changes in integrins and matrix-related functions in cultured human
glomerular epithelial cells. Am J Physiol Renal Physiol.
284:F671–F679. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Zhang L, Li R, Shi W, Liang X, Liu S, Ye
Z, Yu C, Chen Y, Zhang B, Wang W, et al: NFAT2 inhibitor
ameliorates diabetic nephropathy and podocyte injury in db/db mice.
Br J Pharmacol. 170:426–439. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Borza CM, Pozzi A, Borza DB, Pedchenko V,
Hellmark T, Hudson BG and Zent R: Integrin alpha3beta1, a novel
receptor for alpha3(IV) noncollagenous domain and a trans-dominant
inhibitor forintegrin alphavbeta3. J Biol Chem. 281:20932–20939.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Nagase M and Fujita T: Aldosterone and
glomerular podocyte injury. Clin Exp Nephrol. 12:233–242. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Siragy HM and Xue C: Local renal
aldosterone production induces inflammation and matrix formation in
kidneys of diabetic rats. Exp Physiol. 93:817–824. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Lee SH, Yoo TH, Nam BY, Kim DK, Li JJ,
Jung DS, Kwak SJ, Ryu DR, Han SH, Lee JE, et al: Activation of
local aldosterone system within podocytes is involved in apoptosis
under diabetic conditions. Am J Physiol Renal Physiol.
297:F1381–F1390. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Han SY, Kim CH, Kim HS, Jee YH, Song HK,
Lee MH, Han KH, Kim HK, Kang YS, Han JY, et al: Spironolactone
prevents diabetic nephropathy through an anti-inflammatory
mechanism in type 2 diabetic rats. J Am Soc Nephrol. 17:1362–1372.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Toyonaga J, Tsuruya K, Ikeda H, Noguchi H,
Yotsueda H, Fujisaki K, Hirakawa M, Taniguchi M, Masutani K and
Iida M: Spironolactone inhibits hyperglycemia-induced podocyte
injury by attenuating ROS production. Nephrol Dial Transplant.
26:2475–2484. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Taira M, Toba H, Murakami M, Iga I,
Serizawa R, Murata S, Kobara M and Nakata T: Spironolactone
exhibits direct renoprotective effects and inhibits renal
renin-angiotensin-aldosterone system in diabetic rats. Eur J
Pharmacol. 589:264–271. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Han KH, Kang YS, Han SY, Jee YH, Lee MH,
Han JY, Kim HK, Kim YS and Cha DR: Spironolactone ameliorates renal
injury and connective tissue growth factor expression in type II
diabetic rats. Kidney Int. 70:111–120. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Nielsen SE, Persson F, Frandsen E, Sugaya
T, Hess G, Zdunek D, Shjoedt KJ, Parving HH and Rossing P:
Spironolactone diminishes urinary albumin excretion in patients
with type 1 diabetes and microalbuminuria: A randomized
placebo-controlled crossover study. Diabet Med. 29:e184–e190. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Rossing K, Schjoedt KJ, Smidt UM, Boomsma
F and Parving HH: Beneficial effects of adding spironolactone to
recommended anti-hypertensive treatment in diabetic nephropathy: A
randomized, double-masked, cross-over study. Diabetes Care.
28:2106–2112. 2005. View Article : Google Scholar : PubMed/NCBI
|