Introduction
Hepatocellular carcinoma (HCC), which accounts for
between 85 and 90% of cases of primary liver cancer, is the fifth
most common cancer worldwide and is currently the third leading
cause of cancer-associated mortality (1). The prognosis of patients with HCC
continues to be poor, despite improving treatment approaches, with
a 5-year overall survival less than 30% (2). The curative rate of early-stage HCC
has been improved by the use of liver resection or transplantation.
However, the curative efficacy of these therapies is limited due to
the fact that the majority of patients with HCC are diagnosed at a
late stage with severe intrahepatic and extra-hepatic metastases
(3). Novel diagnostic markers,
prognostic indicators and effective therapeutic targets are
required, thus it is important to investigate the underlying
mechanisms of the pathogenesis of HCC.
Rhotekin 2 (RTKN2) is a member of the rhotekin
protein family. The two rhotekin proteins, RTKN and RTKN2, have an
N-terminal HR1 domain (Rho-GTPase binding domain) and pleckstrin
homology domain (4). The similar
protein architecture of rhotekin proteins indicated that they may
share functional characteristics (4). The HR1 domain of murine RTKN has been
demonstrated to bind to Ras homolog gene family member A (Rho A)
and to inhibit the GTPase activity of Rho A (5). Rho proteins are crucial in cell
cytoskeletal organization, cell growth, cell differentiation and
transformation. Dysregulation of the Rho signal transduction
pathway has been implicated in numerous types of cancer (6). The inhibitory role of RTKN2 on Rho A
has previously indicated an important function for rhotekin
proteins in cancer (5).
Previous studies have demonstrated an anti-apoptotic
role for rhotekin (7,8). Liu et al (8) reported the overexpression of RTKN in
the majority of gastric cancer patients and demonstrated an
association between RTKN expression and metastatic progression.
Stable RTKN-expressing gastric cells have been reported to be
resistant to apoptosis induced by sodium butyrate and serum
deprivation (7). A previous study
reported that overexpression of RTKN resulted in the activation of
nuclear factor (NF)-κB and an induction of a number of
NF-κB-regulated anti-apoptotic genes (7). Conversely, suppressing expression of
RTKN by short interfering RNA (siRNA) greatly sensitized cells to
apoptosis (7). Similarly, several
studies have been conducted concerning the anti-apoptotic effects
of RTKN2 (9–12). RTKN2 was originally identified in a
promyelocytic cell line resistant to 25-OHC-induced apoptosis
(12). RTKN2 has been reported to
serve a role in the process of intrinsic apoptosis of HEK cells via
activating NF-κB signaling (10).
On the contrary, suppression of RTKN2 in primary human CD4+
lymphocytes reduced viability and increased sensitivity to 25-OHC
(10). Collectively, the previous
studies demonstrated the anti-apoptotic role of RTKN2. However,
little is known about the expression pattern and the role of RTKN2
in cancer, particularly in HCC.
In the present study, the role of RTKN2 in HCC and
the associated mechanisms were investigated. Firstly, we found that
the mRNA level of RTKN2 was significantly increased in HCC tissues
compared to non-tumorous tissues. Then we investigated the role of
RTKN2 in multiple cellular progress including cell proliferation,
cell cycle, apoptosis and invasion.
Materials and methods
Patients and tissue samples
Tumor tissues and paired noncancerous tissues were
collected from 30 patients with HCC admitted to the Department of
Radiology, Jingzhou Central Hospital (Jingzhou, China) between 2010
and 2012. Ethical approval for the study was provided by the
Independent Ethics Committee of Jingzhou Central Hospital. Informed
and written consent was obtained from all patients or their
advisers according to the ethics committee guidelines.
Cell lines
HEK293T, HCC, HepG2, HuH7, BEL-7404, SMMC-7721,
MHCC-97L and MHCC-97H cells were obtained from the Cell Bank of
Shanghai Biology Institute, Chinese Academy of Science (Shanghai,
China). All culture media were supplemented with 10% fetal bovine
serum, 100 mg/ml penicillin G and 50 µg/ml streptomycin
(Life Technologies; Thermo Fisher Scientific, Inc., Waltham, MA,
USA). HEK293T, HepG2, HuH7, BEL-7404, MHCC-97L and MHCC-97H cells
were cultured in Dulbecco's modified Eagle's medium (DMEM; Life
Technologies; Thermo Fisher Scientific, Inc.) and the SMMC-7721
cells were cultured in Roswell Park Memorial Institute 1640 medium
(Life Technologies; Thermo Fisher Scientific, Inc.). All cells were
maintained at 37°C in 5% CO2.
Vector construction
pLKO.1, psPAX2 and pMD2.G were purchased from
Addgene, Inc. (Cambridge, MA, USA). Three small hairpin RNAs
(shRNAs; Generay Biotech Co., Ltd., Shanghai, China) targeting
human RTKN2 mRNA were cloned into a lentiviral vector (PLKO.1). A
non-specific scramble shRNA sequence (CCT AAG GTT AAG TCG CCC TCG)
was used as negative control. The constructs were then transfected
into HEK293T cells with lentiviral packaging vectors (psPAX2 and
pMD2.G) using Lipofectamine 2000 (Invitrogen; Thermo Fisher
Scientific, Inc.) according to the manufacturer's instructions.
Viruses were collected 48 h subsequent to transfection and used to
infect HepG2 and BEL-7404 cells. After 48 h, the cells were
processed for reverse transcription-quantitative polymerase chain
reaction (RT-qPCR) and western blotting. The shRNA target sequences
for RTKN2 were listed in Table
I.
 | Table IshRNA target sequences for RTKN2. |
Table I
shRNA target sequences for RTKN2.
| shRNA (targeting
position) | Sequence |
|---|
| RTKN2-RNAi-1
(385–407) |
GAGCACTCAGAAAGATCAAGTCT |
| RTKN2-RNAi-2
(1418–1440) |
AAAGGTTCTATGTGCCTAAATCT |
| RTKN2-RNAi-3
(1967–1989) | CAGGGAAAGAACAATAG
AAGTCT |
RNA extraction and RT-qPCR
Total RNA was isolated using TRIzol reagent
(Invitrogen; Thermo Fisher Scientific, Inc.) according to the
manufacturer's instructions. cDNA was synthesized using 1 µg
total RNA, 0.5 mM dNTPs, 2.5 µM oligo-dT primer (for mature
mRNAs), 1 U/µl RiboLock RNase Inhibitor and 10 U/µl
M-MuLV Reverse Transcriptase (all from Thermo Fisher Scientific,
Inc.). The cDNA synthesis was performed for 1 h at 37°C and the
reaction was stopped by 10 min incubation at 70°C.
RT-qPCR was performed in an Applied Biosystems 7300
Fast Real-Time PCR System (Applied Biosystems; Thermo Fisher
Scientific, Inc.) using the Absolute Blue qPCR SYBR Green Low ROX
mix (Thermo Fisher Scientific, Inc.). For each reaction 5 µl
cDNA template, 1 µl primer pairs (10 µM) and 10
µl of the qPCR SYBR mixture to a final reaction volume of 20
µl were used. All reactions were conducted using the
following cycling parameters: 95°C for 10 min, followed by 40
cycles of 95°C for 15 sec and 60°C for 45 sec, and a final
extension step at 72°C for 5 min. Relative expression levels at the
tested experimental conditions were calculated within each
independent experiment using the using the ΔΔCq method (13). All data represent the average of
three replicates. The quality of the PCR product was monitored
using post-PCR melt curve analysis. The sequences of the primer
pairs used are as follows: RTKN2 (NM_001282941.1), F
5′-ACAGTTCGCGTTGGAGATGGAG-3′ and R 5′-GTCGAGCATTGCACACCATGAG-3′;
glyceraldehyde 3-phosphate dehydrogenase (GAPDH) (NM_001256799.1),
F 5′-CACCCACTCCTCCACCTTTG-3′ and R 5′-CCACCACCCTGTTGCTGTAG-3′.
Western blotting
Treated and untreated HepG2 and BEL-7404 cells were
washed twice with phosphate-buffered saline (PBS) and lysed in
ice-cold radioimmunoprecipitation assay buffer (JRDUN Biotechnology
Co., Ltd., Shanghai, China) with freshly added 0.01% protease
inhibitor cocktail (Sigma-Aldrich, St. Louis, MO, USA) and
incubated on ice for 30 min. Subsequent to centrifugation,
supernatants were collected and stored at −80°C until required for
use. For western blot analysis, the cell lysates were run on a
10–15% sodium dodecyl sulfate-polyacrylimide gel electrophoresis
gel and transferred electrophoretically to a nitrocellulose
membrane (EMD Millipore, Billerica, MA, USA). The blots were
blocked with 5% skimmed milk with gentle shaking, followed by
incubation with primary antibodies. Blots were then incubated with
the horseradish peroxidase-conjugated goat anti-mouse (cat. no.
A0216; dilution 1:1,000) or goat anti-rabbit (cat. no. A0208;
dilution 1:1,000) secondary antibodies (Beyotime Institute of
Biotechnology, Shanghai, China) then were visualized using enhanced
chemiluminescence (ECL; EMD Millipore). Mouse monoclonal anti-PTKN2
(cat. no. ab118069; dilution 1:200), rabbit monoclonal
anti-proliferating cell nuclear antigen (PCNA; cat. no. ab92552;
1:5,000) and mouse monoclonal anti-cyclin-dependent kinase 1 (CDK1;
cat. no. ab18; 1:1,000) antibodies were purchased from Abcam
(Cambridge, MA, USA). The rabbit monoclonal anti-GAPDH antibody
(cat. no. 5174; 1:1,500) was from Cell Signaling Technology, Inc.
(Danvers, MA, USA).
Determination of cell proliferation
Cell proliferation was measured using the
Cell-Counting Kit-8 (CCK-8) Assay kit (Dojindo Molecular
Technologies, Kumamoto, Japan) according to the manufacturer's
instructions. In brief, the treated and untreated HepG2 and
BEL-7404 cells were seeded onto 96-well plates. At the indicated
time points, CCK-8 solution (10 µl in 100 µl DMEM)
was added to each well and incubated for 1 h. Optical density
values at a wavelength 450 nm were measured using a microplate
reader (model 550; Bio-Rad Laboratories, Inc., Hercules, CA, USA).
All experiments were run in triplicate and repeated a minimum of
three times.
Cell cycle distribution assay
The cell cycle was analyzed as previously described
(14) with slight modifications.
Briefly, cells cultured on 60-mm diameter dish were infected with
the indicated virus. After 48 h, cells were harvested by
trypsinization (Beijing Solarbio Science & Technology Co.,
Ltd., Beijing, China), washed with ice-cold PBS, and fixed with
ice-cold 70% ethanol for a minimum of 2 h at −20°C. The fixed cells
were washed with PBS, incubated with ribonuclease A (Sigma-Aldrich)
and propidium iodide (PI; 0.05 mg/ml, Sigma-Aldrich) at room
temperature in the dark for 30 min. DNA content was then analyzed
using a FACScan flow cytometer (BD Biosciences, San Jose, CA, USA).
The percentage of cells in the G0/G1, S and
G2/M phases were determined with FlowJo software version
7.6 (Tree Star, Inc., Ashland, OR, USA). Experiments were performed
in triplicate and 3×104 cells were analyzed per
sample.
Cell apoptosis assay
The percentage of cells actively undergoing
apoptosis was determined by double staining with annexin
V-fluorescein isothiocyanate (FITC; BD Biosciences) and PI
(Sigma-Aldrich). Cells were cultured on 60-mm diameter dishes and
were infected with the indicated viruses. After 48 h, cells were
harvested and then double-labeled with annexin V-FITC and PI as
described by the manufacturer. Cells were analyzed using a FACScan
flow cytometer.
Boyden chamber invasion assay
Invasion chambers (BD Biosciences) were prepared by
coating the membranes (8 mm pores) of 24-well inserts with 10 mg/ml
Matrigel (BD Biosciences) leaving them to set for 1 h at 37°C.
Indicated cells were serum starved for 24 h, then a total of
5×104 cells were added to the upper chamber and
suspended in 200 µl DMEM, with 500 µl serum-free DMEM
added into the lower chamber. Subsequent to 48 h of incubation, the
cells on the upper surface of the filter were completely removed
using a cotton bud. The remaining invaded cells were washed with
PBS, fixed in 4% paraformaldehyde (Beijing Solarbio Science &
Technology Co., Ltd.) and then were stained with 0.2% crystal
violet (Beijing Solarbio Science & Technology Co., Ltd.). The
invading cells were observed under the microscope (ECLIPSE Ti-S;
Nikon Corporation, Tokyo, Japan) at a magnification of ×200 in 10
different fields from each filter. Cells were counted in the
central field of triplicate membranes. Experiments were repeated
three times.
Statistical analysis
All data are presented as the mean ± standard
deviation. Statistical significance was determined by Student's
two-tailed t-test. P<0.05 was considered to indicate a
statistically significant difference.
Results
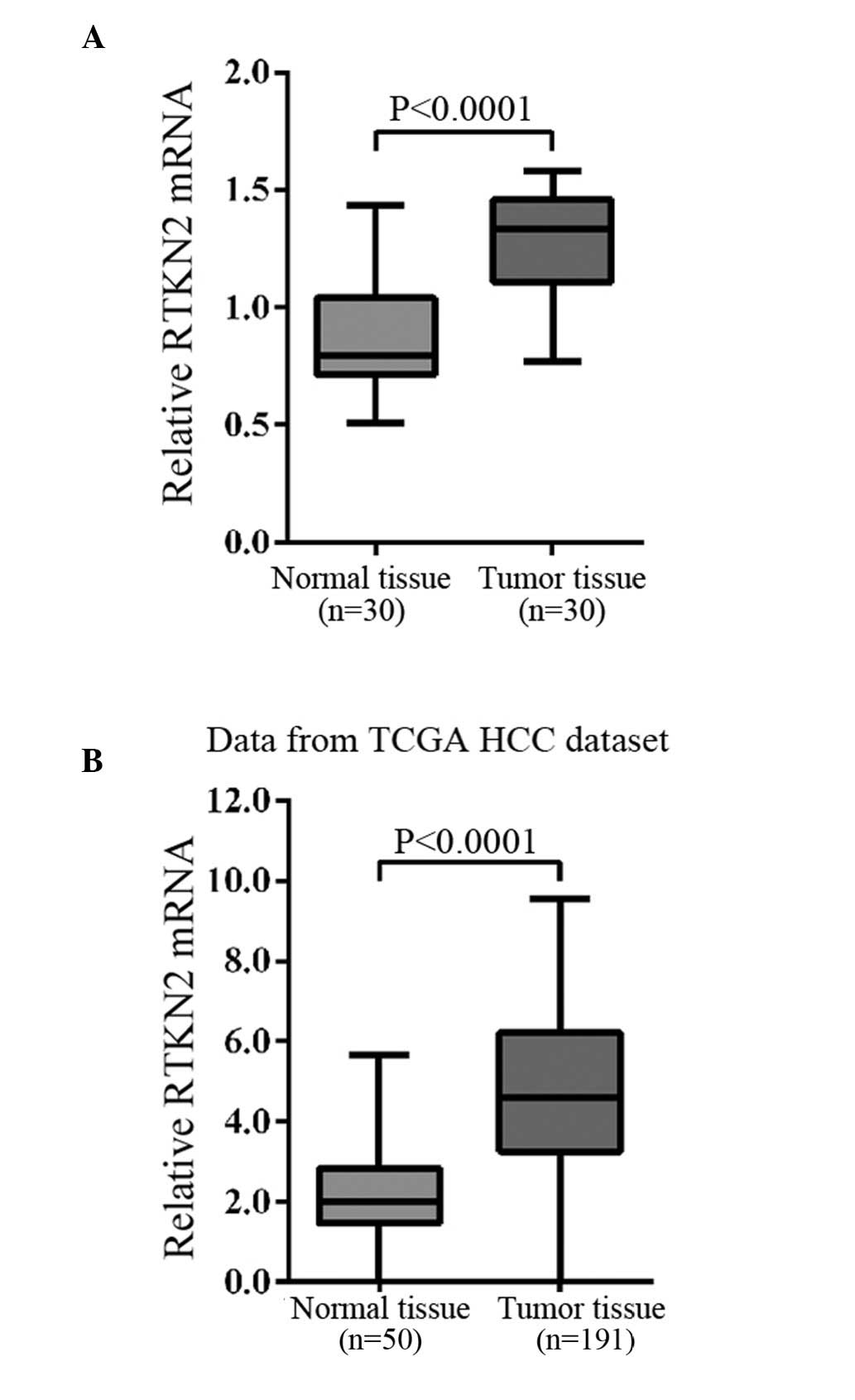
Upregulated RTKN2 expression was observed
in HCC
The mRNA expression levels of RTKN2 were compared
between HCC tissues and the adjacent normal tissues using RT-qPCR.
Overexpression of RTKN2 was identified in 83% (25/30) of the
measured HCC tissues. Statistical analysis using Student's t-test
demonstrated that the RTKN2 mRNA expression levels were
significantly overexpressed in HCC tissues when compared with
normal tissues (Fig. 1A;
P<0.0001).
In addition, the RNA-sequencing data of the HCC
cohort of The Cancer Genome Atlas project (15) was analyzed and it was identified
that RTKN2 expression significantly increased in HCC tumor tissues
compared with the adjacent tissues of patients (Fig. 1B).
RTKN2 was downregulated by RNA
interference (RNAi) in HCC cells
The expression levels of RTKN2 were evaluated using
RT-qPCR and western blotting in the following six HCC cell lines:
HepG2, HuH7, BEL-7404, SMMC-7721, MHCC-97L and MHCC-97H. Two cell
lines, HepG2 and BEL-7404, exhibited higher RTKN2 mRNA and protein
expression, while the remaining four cell lines, HuH7, SMMC-7721,
MHCC-97 L and MHCC-97H, exhibited reduced mRNA and protein
expression (Fig. 2A).
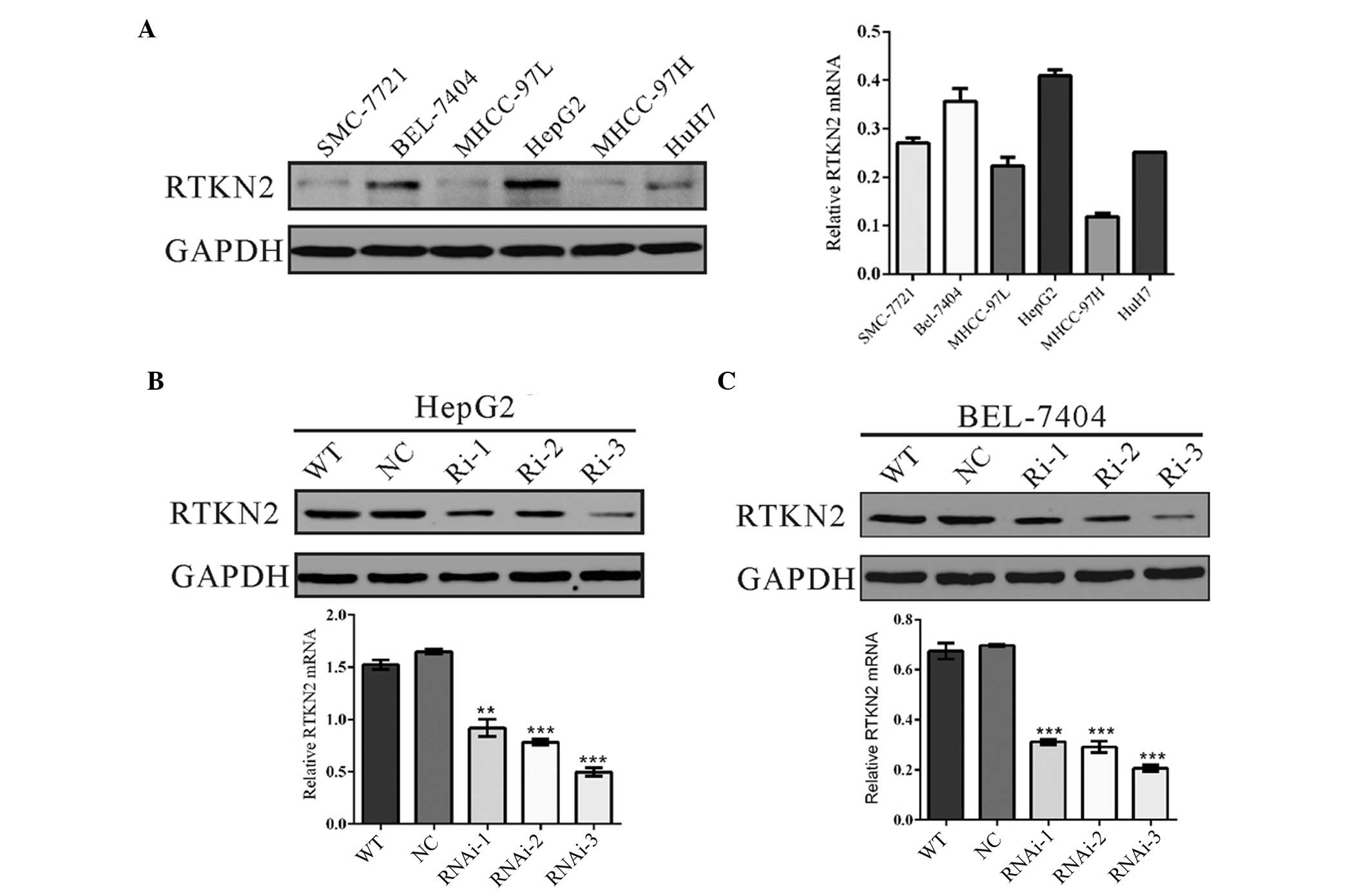 | Figure 2RTKN2 was suppressed by RNAi in HCC
cells. (A) RTKN2 expression level in six HCC cell lines was
analyzed by western blotting (left panel) and RT-qPCR (right
panel). Data were based on a minimum of three independent
experiments. Expression of RTKN2 in (B) HepG2 and (C) BEL-7404
cells was analyzed by western blotting (upper panel) and RT-qPCR
(lower panel). **P<0.01 and ***P<0.001,
vs. the NC. RTKN2, Rhotekin 2; HCC, hepatocellular carcinoma;
RT-qPCR, reverse transcription-quantitative polymerase chain
reaction; GAPDH, glyceraldehyde 3-phosphate dehydrogenase; WT, wild
type cells; NC, scrambled shRNA virus-infected cells; Ri-1, Ri-2
and Ri-3, RTKN2-shRNA-1, -2 and -3 virus-infected cells. |
To investigate the functions of RTKN2 on HCC, the
HepG2 and MHCC-97H cell lines were selected for further study due
to the observed overexpression of RTKN2. Three shRNAs targeting
RTKN2 were all able to efficiently suppress endogenous RTKN2 in
HepG2 (Fig. 2B) and BEL-7404 cells
(Fig. 2C). The RTKN2-Ri-3 shRNA
virus was identified to be the most efficient shRNA, thus was
stably infected into HepG2 and BEL-7404 cells for further
functional analysis.
Suppressing RTKN2 expression inhibited
cell proliferation
To examine the effects of RTKN2 downregulation on
the proliferation of HCC cells, the proliferation of RTKN2
knockdown cells was assessed using the CCK-8 assay. As presented in
Fig. 3A and B, cell growth was
markedly impaired in RTKN2-RNAi virus-infected cells compared with
wild type cells (WT) and scramble shRNA virus-infected cells (NC).
The data indicated a role for RTKN2 in the promotion of HCC cell
proliferation.
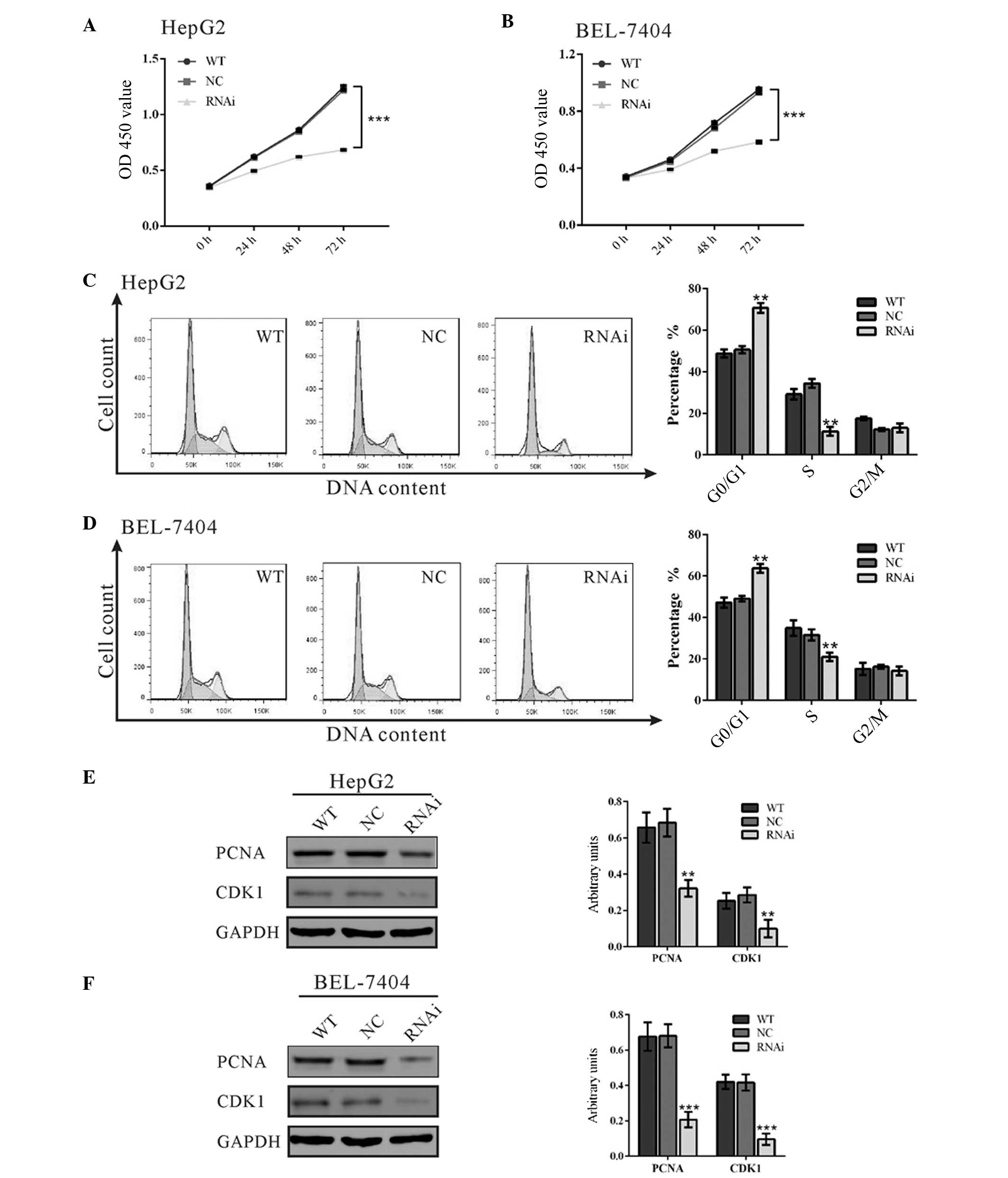 | Figure 3RTKN2 promote cell proliferation by
accelerating G1/S phase transition in HCC cells. (A and
B) Cell proliferation was detected at 0, 24, 48 and 72 h in HepG2
and BEL-7404 cells. (C and D) The cell cycle profile was analyzed
using flow cytometry; (E and F) expression of cell cycle-associated
proteins, PCNA and CD1 were evaluated by western blotting. Data
were based on a minimum of 3 independent experiments, and presented
as the mean ± standard deviation (**P<0.01,
***P<0.001). RTKN2, Rhotekin 2; HCC, hepatocellular
carcinoma; OD, optical density; WT, wild type cells; NC, scrambled
shRNA virus-infected cells; RNAi, RTKN2-shRNA virus-infected
cells. |
Knockdown of RTKN2 repressed
G1/S cell cycle transition
The possible effect of RTKN2 knockdown on cell cycle
progression was then investigated. PI staining and flow cytometry
analysis (Fig. 3C) indicated that
the population of RTKN2-RNAi HepG2 cells in
G0/G1 phase was significantly increased by
39.6% (P<0.05), and the population of S phase cells was markedly
reduced, by 68.0%, compared with cells infected with scramble shRNA
virus-infected cells (NC). Similar results were obtained in
BEL-7404 cells (Fig. 3D). These
results suggested that RTKN2 promoted G1/S cell cycle
transition in HCC cells.
The expression levels of cell cycle-associated
proteins, PCNA and CDK1 were evaluated by western blotting
(Fig. 3E and F). Suppression of
RTKN2 expression significantly reduced PCNA expression (16), a marker of cell proliferation, by
53.0 and 69.6% in HepG2 and BEL-7404 cells, respectively, compared
with corresponding NC cells. The protein level of CDK1, which
induces mitosis during cell cycle progression (17), was also reduced in RTKN2 knockdown
cells. These results suggested that suppression of RTKN2 expression
inhibited cell cycle transition in HCC cells.
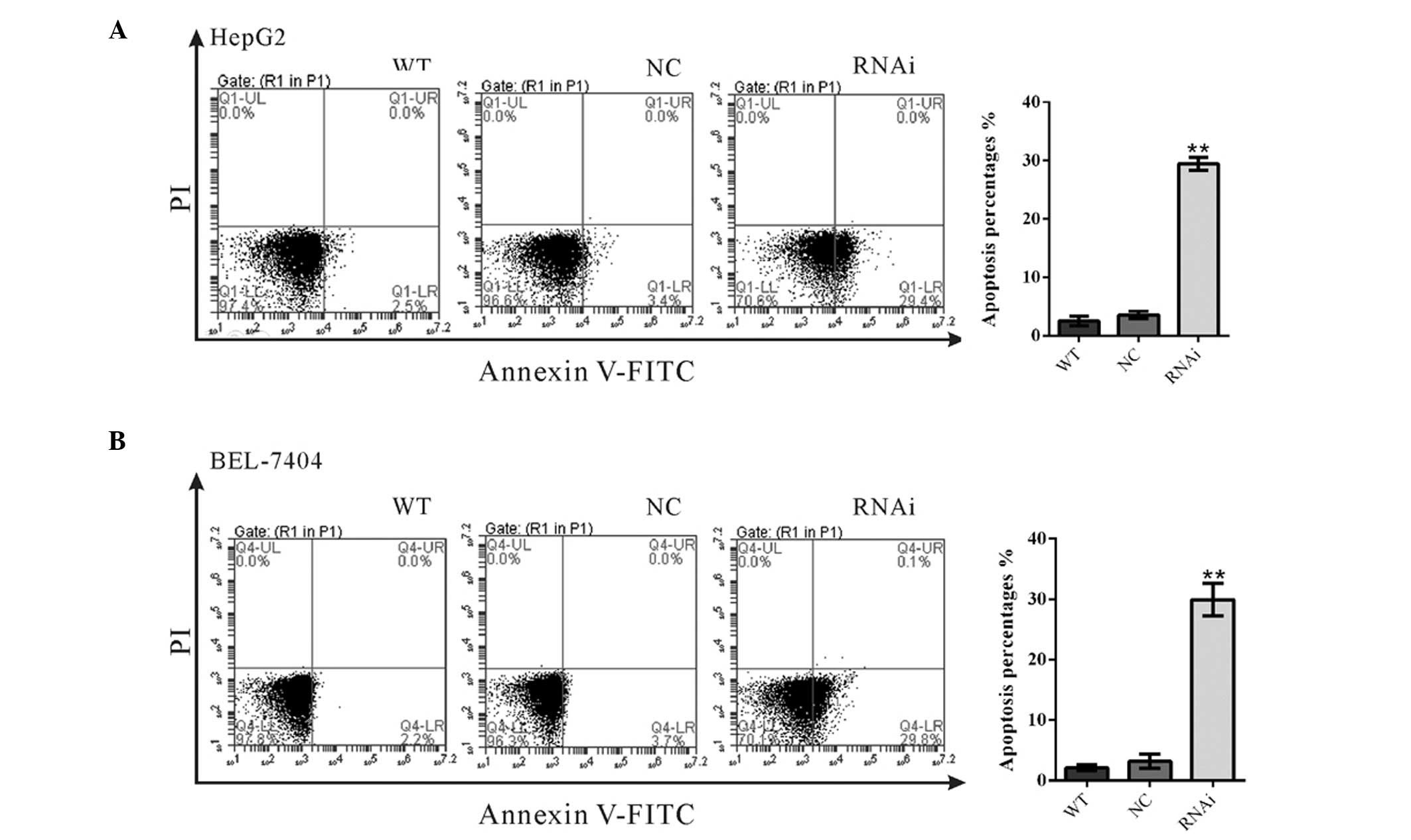
RTKN2 knockdown induced cell
apoptosis
To substantiate the role of RTKN2 on cell apoptosis,
annexin V/PI staining was performed (Fig. 4). The ratio of HepG2 cells
undergoing apoptosis was significantly increased to 29.4±0.66% in
RTKN2-RNAi cells, compared with WT (2.5±0.46%) and NC cells
(3.53±0.35%) (Fig. 4A). Similar
results were observed in BEL-7404 cells (Fig. 4B). These data suggested RTKN2 may
serve an anti-apoptotic role in HCC cells.
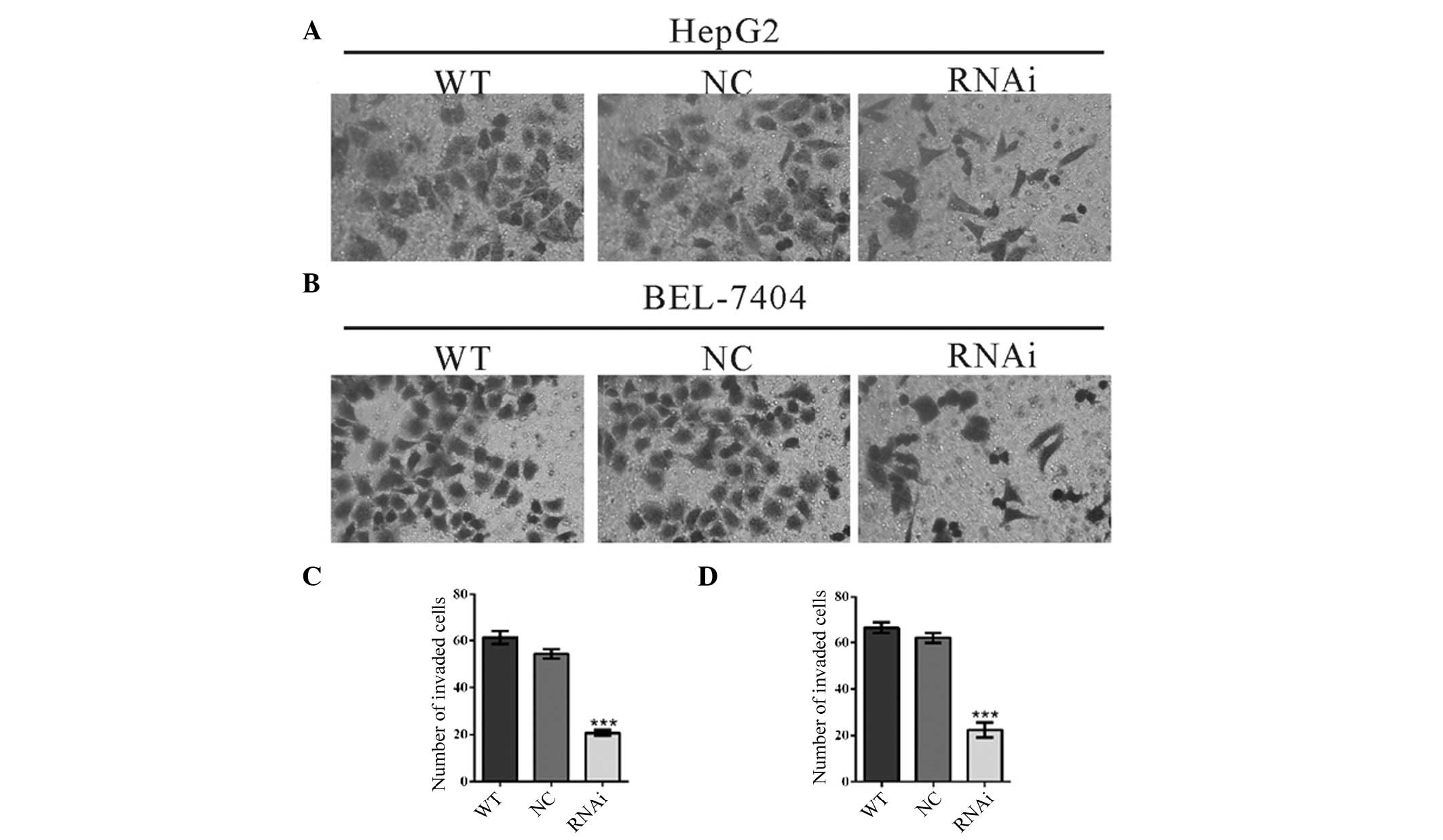
Downregulation of RTKN2 by siRNA
inhibited the invasion of HCC cells
To investigate whether RTKN2 affected the invasive
ability of HCC cells, a Matrigel-coated membrane chamber invasion
assay was conducted (Fig. 5). By
contrast to control cells (WT and NC), a significant reduction in
the invasive ability was observed in RTKN2 knockdown cells. The
number of invaded RTKN2-RNAi cells was less than 40% of that of NC
cells (HepG2 cells: WT, 61±3; NC, 54±2; RTKN2-RNAi, 21±1. BEL-7404
cells: WT, 66±2; NC, 62±2; RTKN2-RNAi, 22±3). These data suggested
that RTKN2 promoted HCC cell invasion.
Discussion
RTKN2 is a newly identified Rho-GTPase effector
protein. Deregulation of Rho GTPase pathways has been reported to
serve an important role in tumorigenesis and cancer metastasis of
HCC cells (18,19). However, the functional implication
of RTKN2 in HCC remains to be fully defined. In the present study,
it was identified that RTKN2 was overexpressed in HCC tissues
(Fig. 1). The effects of RTKN2 on
proliferation, cell cycle, apoptosis and invasion were then
investigated by suppressing its expression in HepG2 and BEL-7404
cells, which have higher levels of endogenous RTKN2 expression. It
was observed that knockdown of RTKN2 significantly reduced cell
growth (Fig. 3A and B).
Furthermore, cell cycle analysis (Fig.
3C and D) indicated that suppression of RTKN2 expression in HCC
cells inhibited cell cycle progression. The expression of the cell
cycle regulators, CDK1 and PCNA, was additionally demonstrated
(Fig. 3E and F). CDK1 forms a
heterodimer with cyclin B, thus inducing mitosis during cell cycle
progression (17), while PCNA
expression has been reported to be a marker of cell proliferation
(16). The data of the current
study indicated that the expression levels of CDK1 and PCNA were
downregulated in RTKN2 knockdown cells, which was consistent with
the results of cell proliferation and cell cycle analysis.
Previous studies have demonstrated the
anti-apoptotic role of RTKN2 (9–12).
Consistent with these observations, the data of the current study
demonstrated that knockdown of RTKN2 significantly stimulated the
apoptosis of HCC cells (Fig. 4).
Invasion is an essential process towards the metastasis of cancer
(20). In the current study,
reduction of the RTKN2 expression in HCC cells reduced their
invasive ability (Fig. 5).
Taken together, the current study indicated that
RTKN2 was overexpressed in HCC tissues. In addition, it was
demonstrated for the first time, to the best of our knowledge, that
RTKN2 served an important role in tumorigenesis and metastasis of
HCC. Whether RTKN2 can be used as a diagnostic marker or
therapeutic target for HCC remains to be further investigated.
References
|
1
|
Jemal A, Bray F, Center MM, Ferlay J, Ward
E and Forman D: Global cancer statistics. CA Cancer J Clin.
61:69–90. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Bosch FX, Ribes J, Díaz M and Cléries R:
Primary liver cancer: Worldwide incidence and trends.
Gastroenterology. 127(5 Suppl 1): S5–S16. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Finn RS: Development of molecularly
targeted therapies in hepatocellular carcinoma: Where do we go now?
Clin Cancer Res. 16:390–397. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Collier FM, Gregorio-King CC, Gough TJ,
Talbot CD, Walder K and Kirkland MA: Identification and
characterization of a lymphocytic Rho-GTPase effector: Rhotekin-2.
Biochem Biophys Res Commun. 324:1360–1369. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Reid T, Furuyashiki T, Ishizaki T,
Watanabe G, Watanabe N, Fujisawa K, Morii N, Madaule P and Narumiya
S: Rhotekin, a new putative target for Rho bearing homology to a
serine/threonine kinase, PKN and rhophilin in the rho-binding
domain. J Biol Chem. 271:13556–13560. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Karlsson R, Pedersen ED, Wang Z and
Brakebusch C: Rho GTPase function in tumorigenesis. Biochim Biophys
Acta. 1796:91–98. 2009.PubMed/NCBI
|
|
7
|
Liu CA, Wang MJ, Chi CW, Wu CW and Chen
JY: Rho/Rhotekin-mediated NF-kappaB activation confers resistance
to apoptosis. Oncogene. 23:8731–8742. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Liu CA, Wang MJ, Chi CW, Wu CW and Chen
JY: Overexpression of rho effector rhotekin confers increased
survival in gastric adenocarcinoma. J Biomed Sci. 11:661–670. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Collier FM, Baker AJ, Walder K, Stupka N,
Martin SD and Kirkland MA: A Rho-GTPase effector, rhotekin-2
(RTKN2) is associated with BMP8b and IL-16 cytokine expression anti
increased sensitivity to apoptosis in lymphocytes. Blood amer soc
hematology 1900 M street. NW suite 200, Washington, DC 20036 USA:
pp. 679A–680A. 2007
|
|
10
|
Collier FM, Loving A, Baker AJ, McLeod J,
Walder K and Kirkland MA: RTKN2 induces NF-KappaB dependent
resistance to intrinsic apoptosis in HEK cells and regulates BCL-2
genes in human CD4(+) lymphocytes. J Cell Death. 2:9–23.
2009.PubMed/NCBI
|
|
11
|
Li W, Wu YF, Xu RH, Lu H, Hu C and Qian H:
miR-1246 releases RTKN2-dependent resistance to UVB-induced
apoptosis in HaCaT cells. Mol Cell Biochem. 394:299–306. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Gregorio-King CC, Gough T, Van Der Meer
GJ, Hosking JB, Waugh CM, McLeod JL, Collier FM and Kirkland MA:
Mechanisms of resistance to the cytotoxic effects of oxysterols in
human leukemic cells. J Steroid Biochem Mol Biol. 88:311–320. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Saha SK, Roy S and Khuda-Bukhsh AR:
Ultra-highly diluted plant extracts of Hydrastis canadensis and
Marsdenia condurango induce epigenetic modifications and alter gene
expression profiles in HeLa cells in vitro. J Integr Med.
13:400–411. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Papavasiliou FN and Schatz DG:
Cell-cycle-regulated DNA double-strand breaks in somatic
hypermutation of immunoglobulin genes. Nature. 408:216–221. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Ye G, Qin Y, Lu X, Xu X, Xu S, Wu C, Wang
X, Wang S and Pan D: The association of renin-angiotensin system
genes with the progression of hepatocellular carcinoma. Biochem
Biophys Res Commun. 459:18–23. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Kubben FJ, Peeters-Haesevoets A, Engels
LG, Baeten CG, Schutte B, Arends JW, Stockbrügger RW and Blijham
GH: Proliferating cell nuclear antigen (PCNA): A new marker to
study human colonic cell proliferation. Gut. 35:530–535. 1994.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Ferrell JE Jr, Wu M, Gerhart JC and Martin
GS: Cell cycle tyrosine phosphorylation of p34cdc2 and a
microtubule-associated protein kinase homolog in Xenopus oocytes
and eggs. Mol Cell Biol. 11:1965–1971. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Wong CC, Wong CM, Au SL and Ng IO:
RhoGTPases and Rho-effectors in hepatocellular carcinoma
metastasis: ROCK N'Rho move it. Liver Int. 30:642–656. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Grise F, Bidaud A and Moreau V: Rho
GTPases in hepatocellular carcinoma. Biochim Biophys Acta.
1795:137–151. 2009.PubMed/NCBI
|
|
20
|
Leber MF and Efferth T: Molecular
principles of cancer invasion and metastasis (review). Int J Oncol.
34:881–895. 2009.PubMed/NCBI
|