Introduction
Diabetes has become a global health epidemic. In
2012, the economic burden of diagnosed diabetes was estimated to be
in excess of $245 billion in the United States alone, representing
a >40% increase in cost over 2007 estimates, with most of this
burden attributed to the treatment and management of diabetes
(1,2). Diabetes is commonly associated with
lipid metabolism disorders and abnormal serum lipid levels, which
accelerate the progression of diabetes and lead to atherosclerosis
and cardiovascular diseases (3,4), the
primary causes of death in patients with diabetes. The liver has a
pivotal role in regulating lipid metabolism, accounting for the
production and degradation of fatty acid (FA), cholesterol,
glycolipids, ketone bodies, phospholipids, steroids and
triacylglycerols (5). Fatty liver
disease is highly prevalent in patients with type 2 diabetes
mellitus (6). Increased
circulating levels of free FAs (FFAs) lead to increased delivery of
FFAs to the liver, which subsequently drive the synthesis of excess
triglycerides (TG) in the liver; the accumulation of excess liver
fat is worsened by impaired hepatic FA oxidation in patients with
type 2 diabetes (5). Ameliorating
hepatic lipid metabolism disorders may be an effective way of
improving whole-body lipid metabolism, decelerating the progression
of diabetes complications and improving the prognosis of patients
with diabetes.
AMP-activated protein kinase (AMPK), a
heterotrimeric serine-threonine kinase, is an important cellular
energy sensor in the majority of tissues (7). A previous study demonstrated that
dysfunction of hepatic AMPK in diabetes represents a key mechanism
for hepatic lipid accumulation and hyperlipidemia associated with
diabetes (8). AMPK phosphorylation
in the liver results in the stimulation of FA oxidation and
inhibition of lipogenesis (9,10);
active AMPK phosphorylates and inactivates certain rate-limiting
enzymes in the liver that are associated with lipolysis, such as
acetyl-CoA carboxylase (ACC). ACC catalyzes malonyl CoA synthesis,
which is a major building block for de novo FA synthesis and
also functions as an allosteric inhibitor of carnitine palmitoyl
transferase 1 (CPT1) (10).
Unphosphorylated ACC inhibits CPT1, which is the rate-limiting
enzyme responsible for the transfer of long-chain fatty acyl CoA to
the mitochondria for β-oxidation (11). Therefore, AMPK activation is an
important therapeutic target in hepatic lipid metabolism disorders
and hyperlipidemia, specifically in diabetes (12).
Procyanidins are a complex family of polyphenol
polymers that are present in a wide variety of natural products,
including grape wines, fruits and vegetables. Grape seed
proanthocyanidin extracts (GSPEs) have been demonstrated to exhibit
a variety of potent pharmacological activities, including functions
against oxidative stress, inflammation and atherosclerosis
(13,14). Grape seed procyanidin B2 (GSPB2) is
one of the major components of GSPEs and possesses similar
pharmacological activities. Furthermore, in diabetes models, GSPEs
have lipid-lowering (15) and
hepatocyte protective effects (16), and lead to the activation of AMPK;
GSPE treatment has been demonstrated to ameliorate mitochondrial
dysfunction and inhibit oxidative stress or apoptosis in mesangial
cells treated with high-dose glucosamine (17) or in diabetic nephropathy (18), via AMPK-dependent signaling.
Considering that inactivation of hepatic AMPK is a key event in the
pathogenesis of hyperlipidemia in diabetes (12), GSPB2 may be a useful agent to
ameliorate liver lipid metabolism disorders and to improve
hyperlipidemia in diabetes. To the best of our knowledge, this
hypothesis has not been previously investigated.
The present study aimed to evaluate the effect of
GSPB2 on liver lipid metabolism in the db/db diabetic mouse model
and the potential underlying mechanism. The db/db mice are a
well-established animal model for the investigation of diabetic
complications. We hypothesize that GSPB2 may ameliorate liver lipid
metabolic disorders in db/db mice via the activation of AMPK and
downstream pathways.
Materials and methods
Animals and treatments
Male C57BLKS/J db/db and db/m mice (n=24; 7 weeks
old; average weight, 32.1 g) were purchased from the Model Animal
Research Center of Nanjing University (Nanjing, China). They were
housed in standard animal cages and received laboratory pellet chow
and tap water ad libitum in a constant environment (room
temperature, 20–22°C; humidity, 40–60%) with a 12-h light/dark
cycle. All experimental procedures were approved by the Animal
Ethics Committee of Shandong University (Jinan, China). Mice were
adapted for one week prior to initiation of the study. Age-matched
db/m mice were used as a control group (Control, n=8). GSPB2
(>90% pure) was purchased from Tianjin Jianfeng Natural Produce
R&D Co., Ltd (Tianjin, China). Db/db mice were randomly divided
into two groups for treatment (n=8 each): Vehicle (DM group; normal
saline solution) and GSPB2 (DMT group; 30 mg/kg body weight per day
in normal saline solution orally for 10 weeks). Each group was
observed between weeks 8 and 18 of age without any other
intervention. Animals were weighed each week. At the end of the
intervention, all mice were fasted overnight and sacrificed.
Fasting blood was collected and the liver tissue was dissected. The
sera and tissues were stored at −80°C.
Measurement of body weight, fasting
blood glucose (FBG) and serum lipids
Animals were weighed every week. Fasting blood was
collected prior to sacrifice and centrifuged at 7,700 × g for 10
min at 4°C to measure FBG, TG and total cholesterol (TC) using an
automatic biochemistry and analysis instrument (ADVIA-1650
autoanalyzer; Bayer AG, Leverkusen, Germany). Serum FFA levels were
determined with a FFA Detection kit using the acylCoA
synthetase-acylCoA oxidase (ACS-ACOD) method, according to the
manufacturer's protocol (Wako Pure Chemical Industries, Ltd.,
Osaka, Japan).
Hepatic lipid analysis
The stored liver samples (100 mg) were lysed and
homogenized in 2 ml of a solution containing 150 mmol/l NaCl, 0.1%
Triton X-100 and 10 mmol/l Tris, using a polytron homogenizer (cat.
no. NS-310E; Microtec Co., Ltd., Chiba, Japan) for 1 min at room
temperature. Liver TG level was analyzed with a Tissue triglyceride
assay kit using the glycerol phosphate oxidase-peroxidase method
(Applygen Technologies, Beijing, China) and normalized to protein
levels measured using a bicinchoninic acid (BCA) Protein Assay kit
(Beyotime Institute of Biotechnology, Haimen, China). Liver
homogenate FFA level was determined with a FFA Detection kit using
the ACS-ACOD method (Wako Pure Chemical Industries, Ltd.) and
normalized to protein levels, as previously described (19).
Hepatic pathological examination
The excised parts of livers were immediately fixed
in 4% paraformaldehyde at room temperature for 12 h and embedded in
paraffin. After solidification, 5-µm sections were cut from blocks.
After hematoxylin and eosin (H&E) staining, sections were
examined by light microscopy at a magnification of ×100.
Western blot analysis
Western blot analysis was performed on samples of
livers obtained from the three groups of mice. Mice livers were
homogenized in radioimmunoprecipitation lysis buffer (Beyotime
Institute of Biotechnology), sonicated for 20 sec at 4°C and
normalized with the BCA Protein Assay kit (Beyotime Institute of
Biotechnology). Equal amounts of protein (50 mg) were separated by
10% SDS-PAGE, transferred to polyvinylidene difluoride membranes,
incubated with blocking buffer (5% non-fat dry milk and 0.05%
Tween-20 in TBS) for 1 h at room temperature and probed with
antibodies for phosphorylated AMPK (1:1,000 dilution; cat. no.
#5256; Cell Signaling Technology, Inc., Danvers, MA, USA), AMPK
(1:1,000 dilution; cat. no. #2532; Cell Signaling Technology,
Inc.), phosphorylated ACC (1:1,000 dilution; cat. no. #3661; Cell
Signaling Technology, Inc.), CPT1 (1:1,000 dilution; cat. no.
ab128568; Abcam, Cambridge, UK), 4-hydroxynonenal (4-HNE, 1:1,000
dilution; cat. no. ab46545; Abcam) and GAPDH (1:1,000 dilution;
cat. no. #2118; Cell Signaling Technology, Inc.) overnight at 4°C.
Membranes were subsequently incubated with horseradish
peroxidase-conjugated goat anti-rabbit or anti-mouse (1:4,000
dilution; cat. nos. ZB-2301 and ZB-2305; ZSGB-BIO; OriGene
Technologies, Inc., Beijing, China) as secondary antibodies for 2 h
at room temperature and visualized by chemiluminescence
immunoblotting detection (Amersham Imager 600; GE Healthcare
Bio-Sciences, Pittsburgh, PA, USA). The intensity of each protein
band was quantified by densitometry using ImageJ software (version
1.48; National Institutes of Health, Bethesda, MD, USA).
Isolation of liver mitochondria
Liver tissue (0.5–0.8 g; n=4 per group) was excised
immediately after mice were sacrificed and immersed in ice-cold
mitochondrial isolation buffer [MIB; 210 mM mannitol, 70 mM
sucrose, 10 mM HEPES, 1 mM EDTA; final pH, 7.2; with 0.5% FA-free
bovine serum albumin (BSA)]. Tissue was minced and homogenized with
additional MIB and 0.5% BSA by using a Potter Elvehjem homogenizer
and loose-fitting Teflon pestle. Mitochondrial isolation involved
differential centrifugation at 4°C, as previously described
(20). The mitochondrial pellet
was resuspended in MIB without BSA and centrifuged for an
additional 10 min at 9,600 × g at 4°C for further mitochondrial
purification. The final mitochondrial pellet was resuspended in MIB
and the protein concentration was determined by the Biuret method
(20).
Mitochondrial respiratory
capacity
Oxygen consumption was measured by high-resolution
respirometry (Oroboros Instruments, Innsbruck, Austria). A standard
substrate/inhibitor titration protocol was used as described
previously (21) for functional
analysis of mitochondrial respiratory-chain complexes after adding
isolated mitochondria (0.15 mg) to respiration medium [110 mM
mannitol, 0.5 mM EGTA, 3 mM MgCl2, 20 mM taurine 10 mM
KH2PO4, 60 mM K lactobionate, 0.3 mM DTT and
0.1% BSA (FA-free), adjusted to pH 7.1; 37°C] (22). Briefly, after stabilization (3–5
min), real-time oxygen concentration and flux data were collected
sequentially. Complex I (CI)-dependent mitochondrial respiration
was induced by adding glutamate (10 mM), malate (5 mM) and ADP (1
mM). Complex II (CII)-dependent respiration was induced by adding
rotenone (0.5 µM) to selectively inhibit CI, followed by succinate
(10 mM), which is a CII substrate. Antimycin A (5 µM) was then
added to inhibit CIII, followed by the addition of TMPD (0.5 mM)
and ascorbate (2 mM) as artificial electron donors for CIV. To
ensure that the respiratory capacity of CIV was not limited by
cytochrome c depletion, respiration was measured after the addition
of cytochrome c (10 µM).
Statistical analysis
Statistical analysis was performed using SPSS 16.0
(SPSS Inc., Chicago, IL, USA) and GraphPad Prism 4 (Graphpad
Software Inc., La Jolla, CA, USA). Data were analyzed by one-way
analysis of variance followed by a least significant difference
post hoc test. Results are presented as the mean ± standard error
of the mean, unless otherwise stated. P<0.05 was considered to
indicate a statistically significant difference.
Results
GSPB2 decreases the body weight and
serum lipid levels in diabetic mice
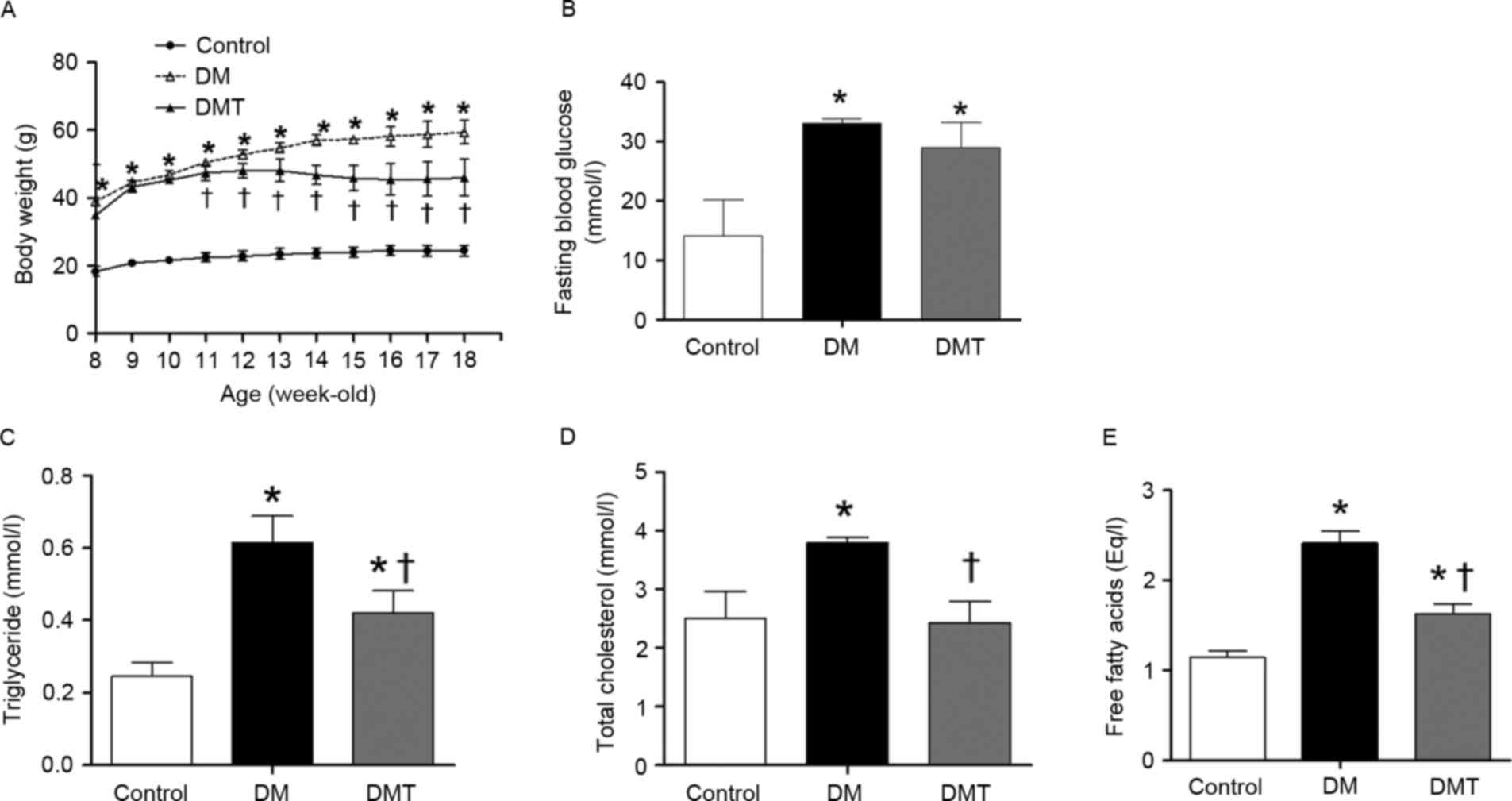
DM and DMT mice had substantially more body weight
compared with age-matched control mice at the beginning of the
study (8 weeks of age; P<0.05; Fig.
1A). Four weeks later however (starting at 11 weeks of age and
throughout the study duration), GSPB2 treatment significantly
decreased the body weight of DMT group compared with the DM group
(P<0.05; Fig. 1A). FBG was
significantly higher in DM and DMT mice compared with control mice
(P<0.05; Fig. 1B), and GSPB2
treatment did not significantly affect FBG levels compared with the
DM group (P>0.05; Fig. 1B).
Compared with control mice, the DM group exhibited higher serum
levels of TG (P<0.05; Fig. 1C),
TC (P<0.05; Fig. 1D) and FFA
(P<0.05; Fig. 1E). GSPB2
treatment significantly decreased the TG, TC and FFA levels
compared with the DM group (all P<0.05), however, serum TG and
FFA levels remained higher compared with control mice in the DMT
group (Fig. 1C-E).
GSPB2 decreases lipid droplet
accumulation and hepatic lipid levels
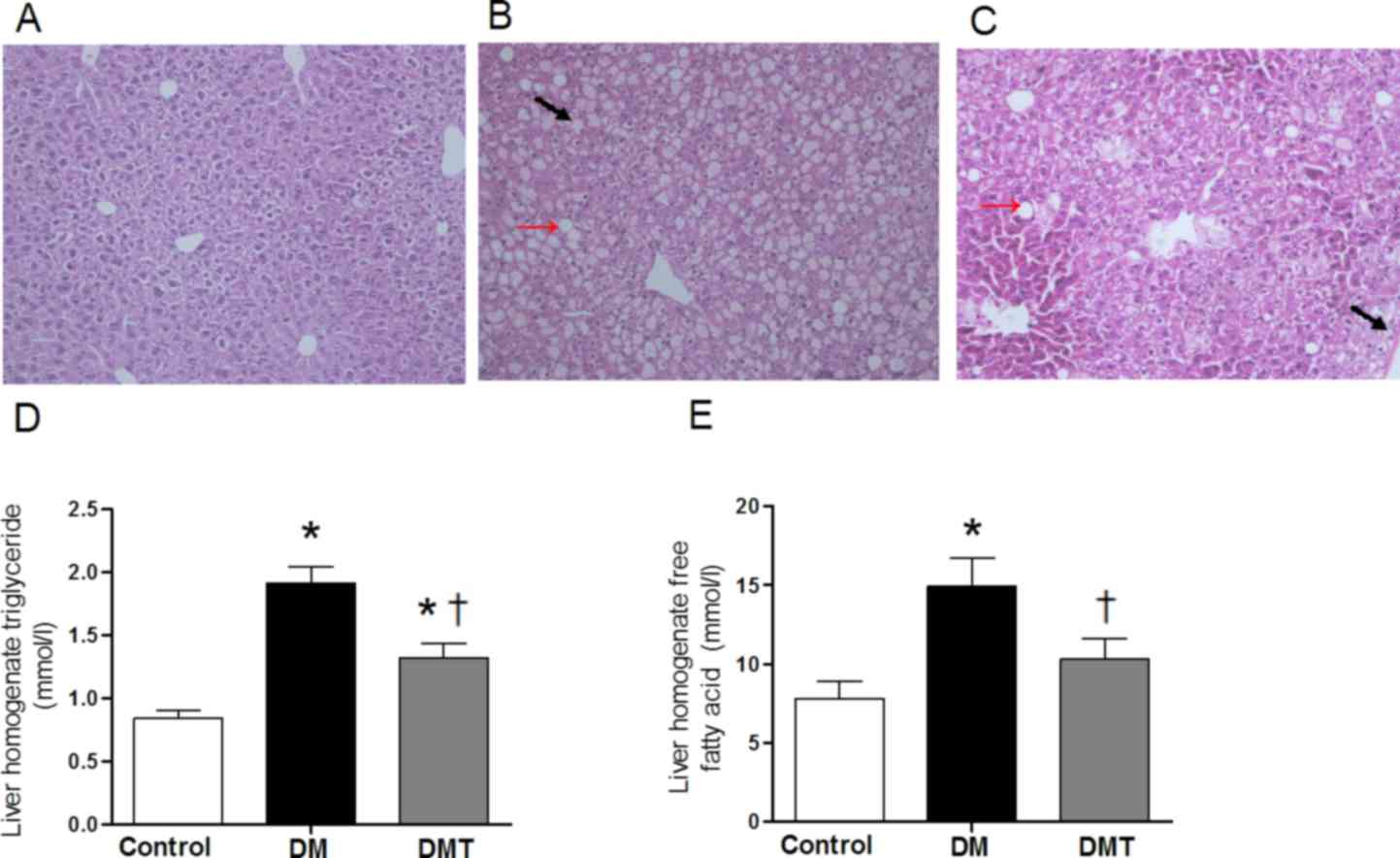
H&E-stained paraffin sections revealed normal
hepatic architecture with clear hepatic lobule, radial liver cell
cord and clear hepatic sinusoid in livers of control mice (Fig. 2A). By contrast, livers of the DM
group exhibited pathological symptoms: Accumulation of lipid
droplets in the cytoplasm and ballooning degeneration (Fig. 2B). After mice were fed GSPB2 for 10
weeks, this hepatocellular damage was ameliorated (Fig. 2C). Changes in the liver homogenate
lipid levels appeared to be similar to serum lipid levels. The DM
group exhibited significantly higher liver homogenate TG and FFA
levels compared with controls (both P<0.05; Fig. 2D and E), which were significantly
decreased by GSPB2 treatment (both P<0.05; Fig. 2D and E).
GSPB2 restores liver AMPK and ACC
phosphorylation levels, increases CPT1 levels and ameliorates lipid
peroxidation damage
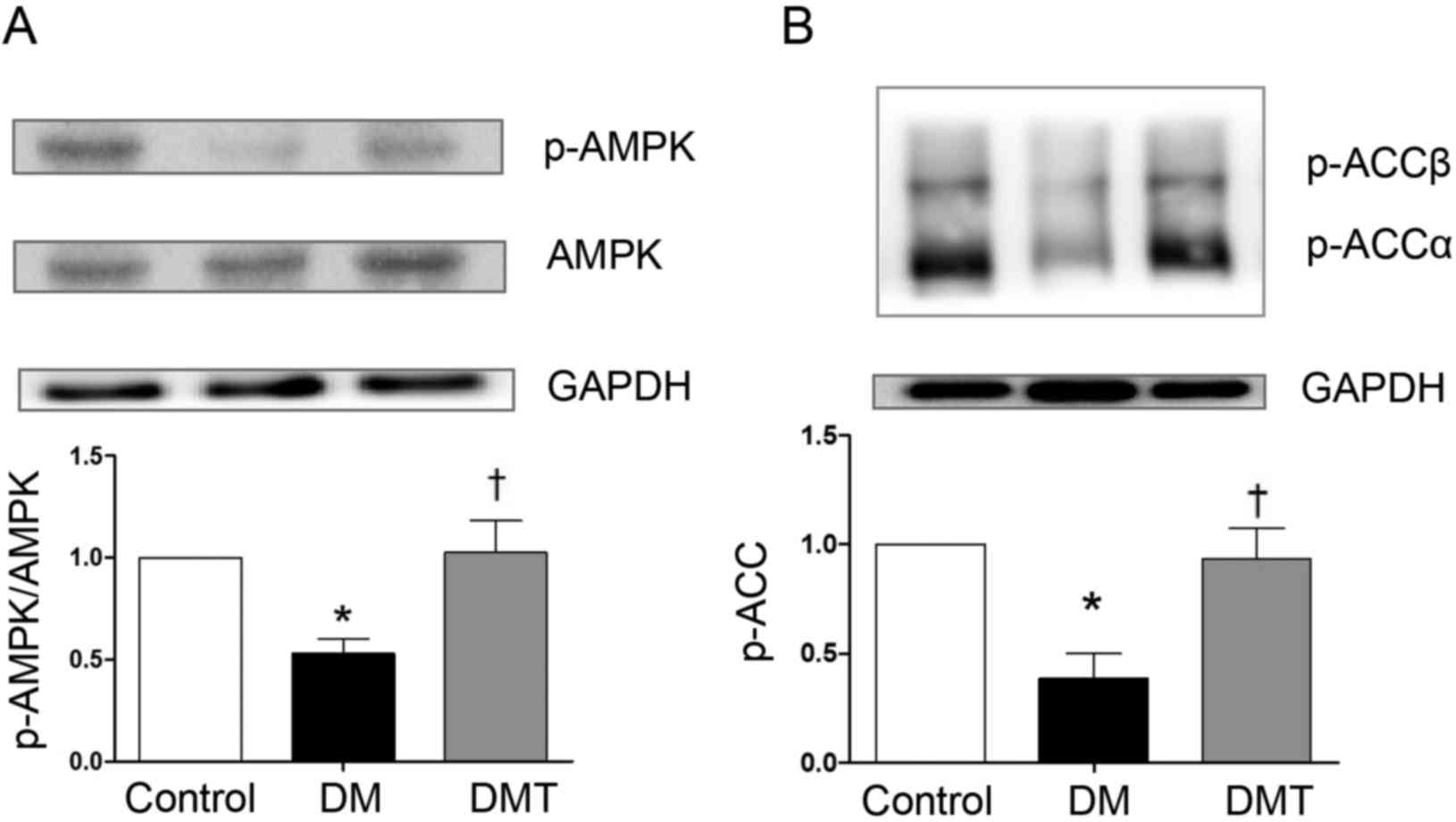
The phosphorylation levels of AMPK and ACC were
lower in the DM group compared with the control group (both
P<0.05; Fig. 3). Following
GSPB2 treatment, the phosphorylation of AMPK and ACC was
significantly increased compared with the DM group (both P<0.05;
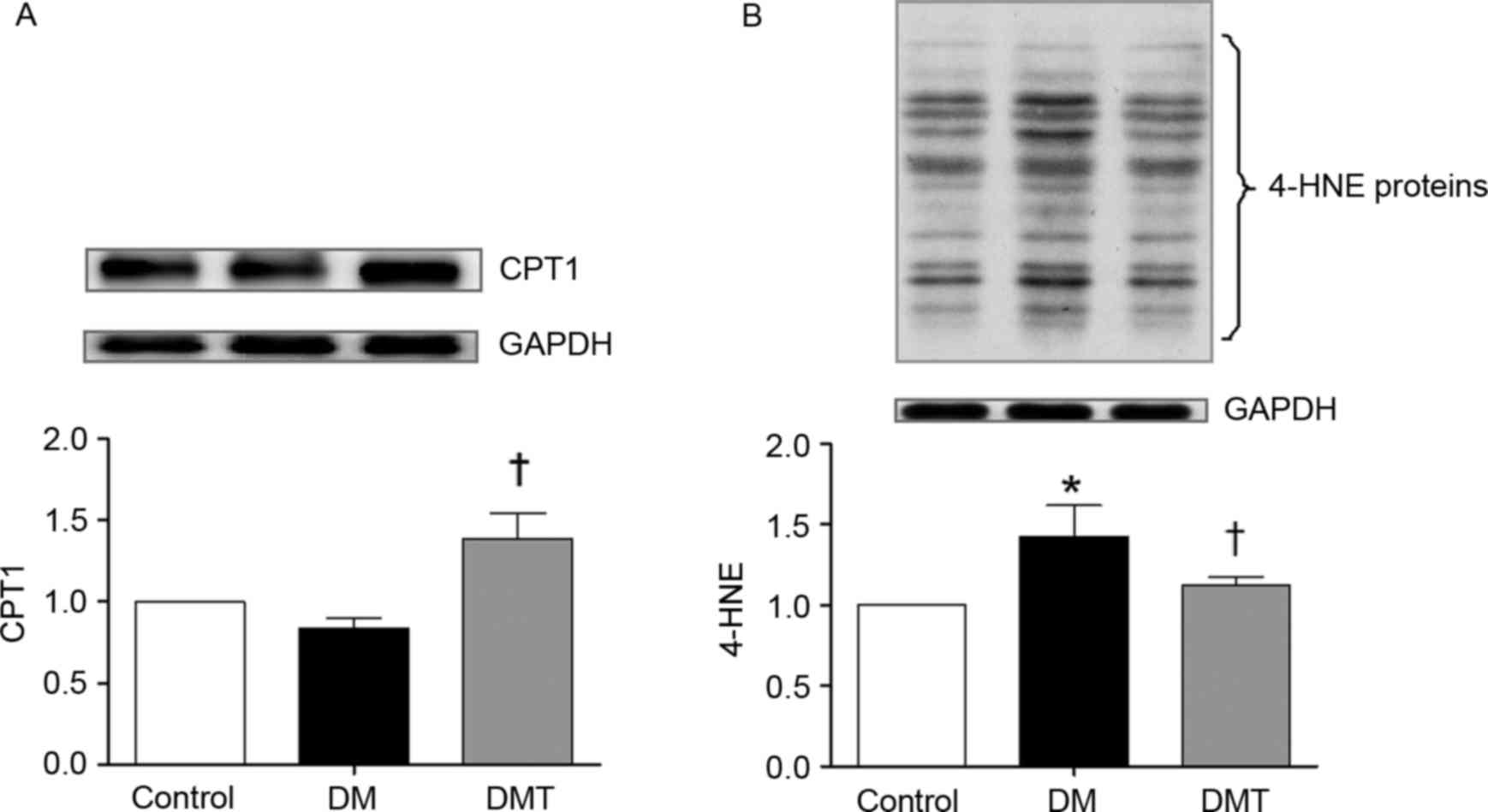
Fig. 3). Furthermore, CPT1 protein
levels were also significantly higher in the DMT group compared
with the DM group (P<0.05; Fig.
4A). 4-HNE protein levels, which are markers of cellular lipid
peroxidation damage, were significantly increased in the DM group
compared with control (P<0.05), but were significantly reduced
following GSPB2 treatment (P<0.05; Fig. 4B).
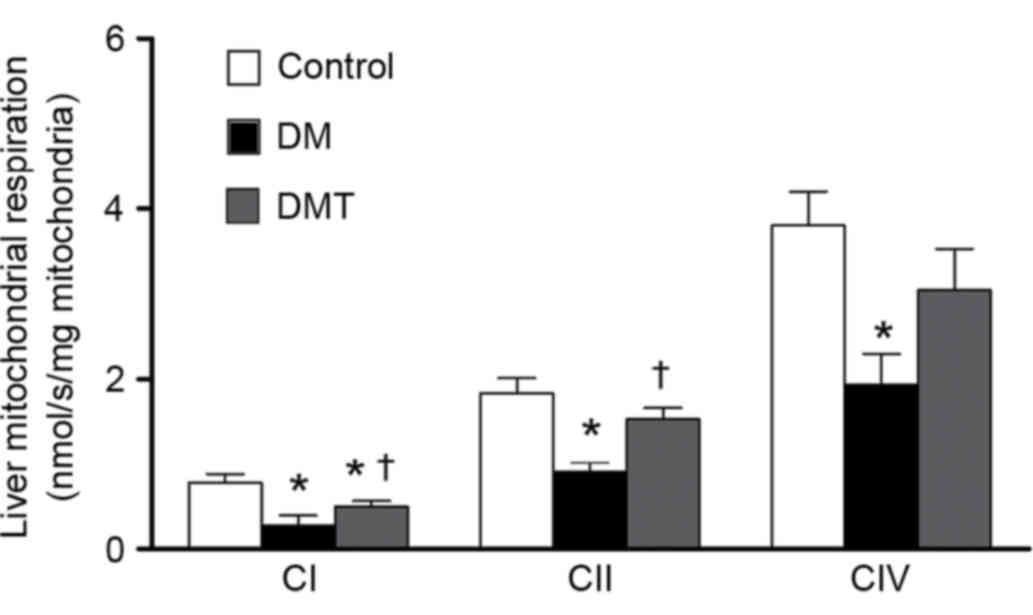
GSPB2 restores damaged liver
mitochondrial respiratory capacity
Liver samples from DM mice exhibited a significant
decrease in the respiration function of all mitochondrial complexes
(CI, CII and CIV), compared with controls (all P<0.05; Fig. 5). However, GSPB2 treatment
significantly restored the respiratory capacities of CI and CII in
the DMT group compared with the DM group (both P<0.05; Fig. 5). The respiratory capacity of CIV
was also increased by GSPB2 treatment in the DMT group compared
with the DM group, however, this increase was not significant
(P>0.05; Fig. 5).
Discussion
GSPB2 is a dimeric form of GSPE, an extract compound
from grape seeds with established lipid-lowering,
anti-atherosclerosis and hepatocyte protective properties (15,16,23,24).
Previous studies from our group have demonstrated the protective
effects of GSPB2 on diabetic complications, including diabetic
nephropathy and diabetic arterial damage (23,24).
As diabetes is frequently complicated by hepatic lipid metabolism
disorders and high lipid levels, in the present study, we
hypothesized that GSPB2 treatment may alleviate hepatic lipid
metabolism disorders and lower serum lipid levels in diabetes.
Long-term oral treatment of diabetic mice with GSPB2 significantly
decreased whole body weight and serum lipid levels, as well as
hepatic lipid droplet accumulation and hepatic TG and FFA levels.
GSPB2 restored liver AMPK and ACC phosphorylation levels, increased
CPT1 protein levels and minimized the subsequent damage from lipid
peroxidation. Furthermore, GSPB2 treatment significantly
ameliorated hepatic mitochondrial dysfunction and restored
mitochondrial CI- and CII-dependent respiratory capacity. The
results of the present study indicate that long-term oral treatment
with GSPB2 may benefit hepatic lipid metabolism disorders,
potentially by increasing hepatic FFA β-oxidation in mitochondria
and decreasing hepatic lipid synthesis via AMPK-ACC-mediated lipid
metabolism.
Although GSPB2 did not significantly reduce FBG, it
normalized serum and hepatic elevated lipid levels and reduced body
weight gain in db/db mice. These results indicate that GSPB2
treatment predominantly affects lipid, but not glucose, metabolism,
suggesting that its main benefit in diabetes may occur through a
potential protective effect against atherosclerosis and
cardiovascular diseases (24,25).
The liver is the major site for the storage and release of
carbohydrates, and the synthesis of FAs. In this diabetic model,
liver lipid accumulation and lipid metabolic disorders were
alleviated by GSPB2 treatment, therefore, GSPB2 may be an effective
agent to improve lipid metabolism, decelerate the progression of
diabetes complications and improve prognosis in diabetes.
In order to uncover the underlying mechanisms of
GSPB2 on liver lipid metabolism, the current study investigated the
potential involvement of AMPK. AMPK has a key role in regulating
lipid metabolism through multiple signaling pathways, including
directly catalyzing its downstream substrates and the transcription
of multiple genes (26). AMPK is
activated >200-fold by phosphorylation at Thr172 (7). Similar to the results of a previous
study (27), the present study
demonstrated that phosphorylated AMPK levels were lower in DM mice
compared with controls. Several other studies have demonstrated
that GSPE treatment ameliorated mitochondrial dysfunction and
inhibited oxidative stress or apoptosis via AMPK-dependent
signaling (17,18). Similarly, in the present study,
GSPB2 treatment significantly restored the hepatic phosphorylation
of AMPK. The activation of AMPK may lead to the stimulation of FA
oxidation and inhibition of lipogenesis (9), reduced hepatic lipid accumulation
and, in turn, attenuated hyperlipidemia and atherosclerosis in DM
(12).
In the present study, from the H&E-stained
paraffin sections and liver homogenate lipid measurements, lipid
synthesis and accumulation in the liver were demonstrated to be
reduced in diabetes, possibly by inactivation of ACC by AMPK. The
phosphorylation of ACC was lower in DM mice compared with controls,
however, ACC phosphorylation was restored by GSPB2 treatment. ACC
is a key downstream target of AMPK and catalyzes the production of
malonyl-CoA from acetyl-CoA, which is a key intermediate of FA
synthesis and oxidation (10).
Increased AMPK activity phosphorylates and inactivates ACCα (at
Ser79) and ACCβ (at Ser221) in the liver, which results in reduced
malonyl-CoA production (28) and
may lead to reduced hepatic FA synthesis (9).
CPT1 is the rate-limiting enzyme that transfers the
long-chain fatty acyl CoA to mitochondria for β-oxidation (11). It has been implicated as
contributing to elevated FFA levels, fat accumulation and decreased
ability to oxidize FAs in diabetes. The increased levels of
malonyl-CoA caused by ACC activation inhibit CPT1, leading to a
subsequent decrease in the transport of long-chain FAs into
mitochondria and decreased FA oxidation (29,30).
The shunting of long-chain FAs away from mitochondria leads to an
increase in FFA levels and the accumulation of fat (29,30).
In the current study, GSPB2 significantly restored the decreased
CPT1 level in DM and increased FA movement as fuel into
mitochondria for β-oxidation, which would lead to increased oxygen
consumption to generate ATP. The mitochondrial CI- and
CCII-dependent respiratory capacity was significantly reduced in DM
mice compared with controls, however, this was significantly
restored by GSPB2 treatment, which may be due to increased fuel
supply for mitochondrial ATP production or ameliorated lipid
peroxidation.
GSPB2 significantly decreased hepatic FFA levels and
minimized subsequent lipid peroxidation damage, as measured by
4-HNE proteins levels. 4-HNE is a measure of lipid peroxidation
from reactive oxygen species (ROS). Elevated FFA levels increase
ROS production in diabetes and obesity (31), which exacerbates mitochondrial
dysfunction. ROS directly damages mitochondrial proteins and
promotes the formation of the mitochondrial permeability transition
pore, increases the release of cytochrome c and enhances apoptosis
(32). Injured mitochondria cannot
effectively reduce oxygen or transfer electrons, which results in
high levels of ROS that further damage tissues, trigger apoptosis
and lead to a vicious cycle of ROS production and mitochondrial
dysfunction (33).
Grape seed extract is generally well tolerated when
taken orally. It has been used safely for up to 8 weeks in clinical
trials (34). Administration of
the grape seed extract to male and female Sprague Dawley rats in
the feed at levels of 0.5, 1.0, or 2.0% for 90 days did not induce
any significant toxicological effects (35). In the present study and using GSPB2
at a dose of 30 mg/kg body weight per day orally for 10 weeks, no
obvious toxicity to the mice was observed. However, certain side
effects of grape seed extract have been reported, including a dry,
itchy scalp, dizziness, headache, high blood pressure, hives,
indigestion and nausea (34).
Consumption of grape seed extract polyphenols may inhibit non-heme
iron absorption and may lead to iron depletion in populations with
marginal iron stores, and also may interact with certain
pharmaceutical agents and enhance their biologic effects (36). A higher dose of grape seed extract
may lead to increased toxicity to animals, and this requires
further investigation. In the current study, the effects of GSPB2
on hepatic lipid metabolism disorders in diabetic mice were
investigated. GSPB2 decreased hepatic lipid droplet accumulation
and hepatic lipid levels. These protective effects appear to be
mediated by increased hepatic FFA β-oxidation in mitochondria and
decreased hepatic lipid synthesis by stimulating an AMPK-ACC lipid
metabolic pathway. In order to fully investigate the potential
molecular mechanism, experiments with either antagonists of AMPK or
an AMPK knock-out mouse strain would be required in future studies.
Overall, GSPB2 may represent a novel therapeutic agent to improve
whole-body lipid metabolism, decelerate the progression of diabetes
complications and improve the prognosis of patients with
diabetes.
Acknowledgements
This work was supported by grants from the National
Natural Science Foundation of China (grant no. 81501786), the Major
Projects of National Science and Technology of China (grant no.
2012ZX09303016-003) and the Horizontal Project of Shandong
University (grant no. 11671614). The sponsor of the funding had no
active role in the design, methods, data collections, analysis, or
preparation of this manuscript or in the decision to submit the
manuscript for publication. We thank Ms. Laura Smales
(BioMedEditing, Toronto, ON, Canada) for language editing of this
manuscript.
References
|
1
|
American Diabetes Association, . Economic
costs of diabetes in the U.S. in 2012. Diabetes Care. 36:1033–1046.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Ginter E and Simko V: Type 2 diabetes
mellitus, pandemic in 21st century. Adv Exp Med Biol. 771:42–50.
2012.PubMed/NCBI
|
|
3
|
Goldberg IJ: Clinical review 124: Diabetic
dyslipidemia: Causes and consequences. J Clin Endocrinol Metab.
86:965–971. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Yan L, Xu MT, Yuan L, Chen B, Xu ZR, Guo
QH, Li Q, Duan Y, Fu J Huang, Wang YJ, et al: Prevalence of
dyslipidemia and its control in type 2 diabetes: A multicenter
study in endocrinology clinics of China. J Clin Lipidol.
10:150–160. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Bhatt HB and Smith RJ: Fatty liver disease
in diabetes mellitus. Hepatobiliary Surg Nutr. 4:101–108.
2015.PubMed/NCBI
|
|
6
|
Stefan N and Häring HU: The metabolically
benign and malignant fatty liver. Diabetes. 60:2011–2017. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Hardie DG: AMP-activated protein kinase:
An energy sensor that regulates all aspects of cell function. Genes
Dev. 25:1895–1908. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Hou X, Xu S, Maitland-Toolan KA, Sato K,
Jiang B, Ido Y, Lan F, Walsh K, Wierzbicki M, Verbeuren TJ, et al:
SIRT1 regulates hepatocyte lipid metabolism through activating
AMP-activated protein kinase. J Biol Chem. 283:20015–20026. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Viollet B, Foretz M, Guigas B, Horman S,
Dentin R, Bertrand L, Hue L and Andreelli F: Activation of
AMP-activated protein kinase in the liver: A new strategy for the
management of metabolic hepatic disorders. J Physiol. 574:41–53.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Wong AK, Howie J, Petrie JR and Lang CC:
AMP-activated protein kinase pathway: A potential therapeutic
target in cardiometabolic disease. Clin Sci (Lond). 116:607–620.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Ruderman NB, Saha AK and Kraegen EW:
Minireview: Malonyl CoA, AMP-activated protein kinase and
adiposity. Endocrinology. 144:5166–5171. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Zang M, Xu S, Maitland-Toolan KA, Zuccollo
A, Hou X, Jiang B, Wierzbicki M, Verbeuren TJ and Cohen RA:
Polyphenols stimulate AMP-activated protein kinase, lower lipids
and inhibit accelerated atherosclerosis in diabetic LDL
receptor-deficient mice. Diabetes. 55:2180–2191. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Terra X, Montagut G, Bustos M, Llopiz N,
Ardèvol A, Bladé C, Fernández-Larrea J, Pujadas G, Salvadó J, Arola
L and Blay M: Grape-seed procyanidins prevent low-grade
inflammation by modulating cytokine expression in rats fed a
high-fat diet. J Nutr Biochem. 20:210–218. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Sharma SD and Katiyar SK: Dietary grape
seed proanthocyanidins inhibit UVB-induced cyclooxygenase-2
expression and other inflammatory mediators in UVB-exposed skin and
skin tumors of SKH-1 hairless mice. Pharm Res. 27:1092–1102. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Pajuelo D, Díaz S, Quesada H,
Fernández-Iglesias A, Mulero M, Arola-Arnal A, Salvadó MJ, Bladé C
and Arola L: Acute administration of grape seed proanthocyanidin
extract modulates energetic metabolism in skeletal muscle and BAT
mitochondria. J Agric Food Chem. 59:4279–4287. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Mansouri E, Khorsandi L and Abedi HA:
Antioxidant effects of proanthocyanidin from grape seed on hepatic
tissue injury in diabetic rats. Iran J Basic Med Sci. 17:460–464.
2014.PubMed/NCBI
|
|
17
|
Bao L, Cai X, Zhang Z and Li Y: Grape seed
procyanidin B2 ameliorates mitochondrial dysfunction and inhibits
apoptosis via the AMP-activated protein kinase-silent mating type
information regulation 2 homologue 1-PPARγ co-activator-1α axis in
rat mesangial cells under high-dose glucosamine. Br J Nutr.
113:35–44. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Bao L, Cai X, Dai X, Ding Y, Jiang Y and
Li Y, Zhang Z and Li Y: Grape seed proanthocyanidin extracts
ameliorate podocyte injury by activating peroxisome
proliferator-activated receptor-γ coactivator 1α in low-dose
streptozotocin-and high-carbohydrate/high-fat diet-induced diabetic
rats. Food Funct. 5:1872–1880. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Han S, Jiao J, Zhang W, Xu J, Wan Z, Zhang
W, Gao X and Qin L: Dietary fiber prevents obesity-related liver
lipotoxicity by modulating sterol-regulatory element binding
protein pathway in C57BL/6J mice fed a high-fat/cholesterol diet.
Sci Rep. 5:152562015. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Palmeira CM and Moreno AJ: Mitochondrial
bioenergetics: Methods and protocols. In: High-resolution
respirometryOXPHOS protocols for human cells and permeabilized
fibers from small biopsies of human muscle. Pesta D and Gnaiger E:
Humana Press; New York, NY: pp. 25–58. 2012
|
|
21
|
Kuznetsov AV, Veksler V, Gellerich FN,
Saks V, Margreiter R and Kunz WS: Analysis of mitochondrial
function in situ in permeabilized muscle fibers, tissues and cells.
Nat Protoc. 3:965–976. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Karamercan MA, Weiss SL, Villarroel JP,
Guan Y, Werlin E, Figueredo R, Becker LB and Sims C: Can peripheral
blood mononuclear cells be used as a proxy for mitochondrial
dysfunction in vital organs during hemorrhagic shock and
resuscitation? Shock. 40:476–484. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Zhang Z, Li BY, Li XL, Cheng M, Yu F, Lu
WD, Cai Q, Wang JF, Zhou RH, Gao HQ and Shen L: Proteomic analysis
of kidney and protective effects of grape seed procyanidin B2 in
db/db mice indicate MFG-E8 as a key molecule in the development of
diabetic nephropathy. Biochim Biophys Acta. 1832:805–816. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Yu F, Li BY, Li XL, Cai Q, Zhang Z, Cheng
M, Yin M, Wang JF, Zhang JH, Lu WD, et al: Proteomic analysis of
aorta and protective effects of grape seed procyanidin B2 in db/db
mice reveal a critical role of milk fat globule epidermal growth
factor-8 in diabetic arterial damage. PLoS One. 7:e525412012.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Luan SS, Yu F, Li BY, Qin RJ, Li XL, Cai
Q, Yin WB, Cheng M and Gao HQ: Quantitative proteomics study of
protective effects of grape seed procyanidin B2 on diabetic
cardiomyopathy in db/db mice. Biosci Biotechnol Biochem.
78:1577–1583. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Hardie DG: The AMP-activated protein
kinase pathway-new players upstream and downstream. J Cell Sci.
117:5479–5487. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Viollet B, Lantier L, Devin-Leclerc J,
Hebrard S, Amouyal C, Mounier R, Foretz M and Andreelli F:
Targeting the AMPK pathway for the treatment of Type 2 diabetes.
Front Biosci (Landmark Ed). 14:3380–3400. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Kim MK, Kim SH, Yu HS, Park HG, Kang UG,
Ahn YM and Kim YS: The effect of clozapine on the AMPK-ACC-CPT1
pathway in the rat frontal cortex. Int J Neuropsychopharmacol.
15:907–917. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Rasmussen BB, Holmbäck UC, Volpi E,
Morio-Liondore B, Paddon-Jones D and Wolfe RR: Malonyl coenzyme A
and the regulation of functional carnitine palmitoyltransferase-1
activity and fat oxidation in human skeletal muscle. J Clin Invest.
110:1687–1693. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
McGarry JD, Mills SE, Long CS and Foster
DW: Observations on the affinity for carnitine and malonyl-CoA
sensitivity, of carnitine palmitoyltransferase I in animal and
human tissues. Demonstration of the presence of malonyl-CoA in
non-hepatic tissues of the rat. Biochem J. 214:21–28. 1983.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Inoguchi T, Li P, Umeda F, Yu HY, Kakimoto
M, Imamura M, Aoki T, Etoh T, Hashimoto T, Naruse M, et al: High
glucose level and free fatty acid stimulate reactive oxygen species
production through protein kinase C-dependent activation of NAD(P)H
oxidase in cultured vascular cells. Diabetes. 49:1939–1945. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Skripchenko A, Myrup A,
Thompson-Montgomery D, Awatefe H, Moroff G and Wagner SJ: Periods
without agitation diminish platelet mitochondrial function during
storage. Transfusion. 50:390–399. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Cuzzocrea S, Riley DP, Caputi AP and
Salvemini D: Antioxidant therapy: A new pharmacological approach in
shock, inflammation, and ischemia/reperfusion injury. Pharmacol
Rev. 53:135–159. 2001.PubMed/NCBI
|
|
34
|
Sovak M: Grape Extract, Resveratrol and
Its Analogs: A Review. J Med Food. 4:93–105. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Wren AF, Cleary M, Frantz C, Melton S and
Norris L: 90-day oral toxicity study of a grape seed extract
(IH636) in rats. J Agric Food Chem. 50:2180–2192. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Mennen LI, Walker R, Bennetau-Pelissero C
and Scalbert A: Risks and safety of polyphenol consumption. Am J
Clin Nutr. 81:(1 Suppl). S326–S329. 2005.
|