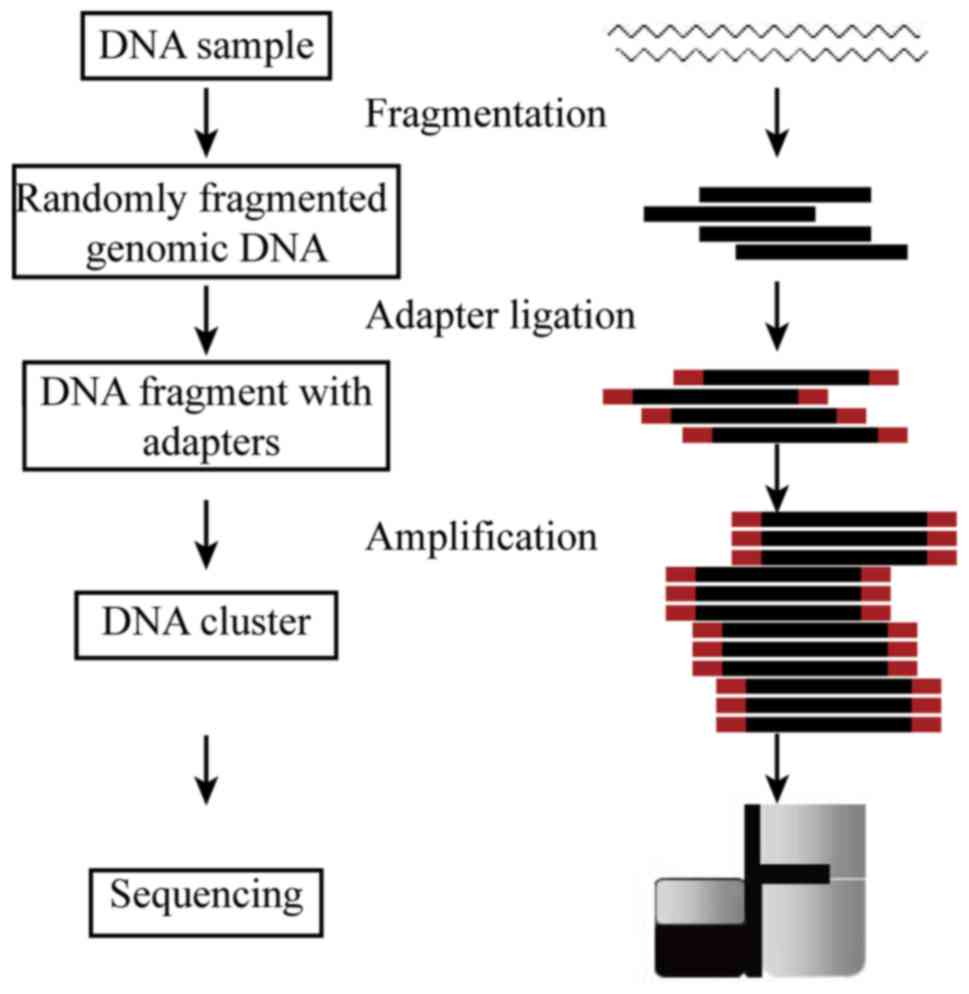
Introduction
Eesophageal squamous cell carcinoma (ESCC) is one of
the most common malignant diseases world wide, particularly in
China, where it is the fourth most common cause of
cancer-associated mortality (1).
Unlike cancers that have been extensively studied, such as breast
and colon cancers, the outcome of ESCC remains unchanged during the
last several decades, with a 5-year survival rate ranging from
15–25% (2). However, in general,
understanding of the genomic abnormalities in this disease is
limited to reports from small cohorts (3–5).
Advances in next-generation sequencing (NGS) technology are
facilitating the identification of novel disease-associated genes
and promising to transform the routine clinical diagnosis of
inherited disease. Since the initial reports in 2009 (6,7), NGS
has aided the discovery of >50 novel disease genes in research
settings. Thus, a there is a compelling requirement to extensively
identify genomic abnormalities underlying ESCC by NGS, including
single-nucleotide polymorphisms (SNPs), insertions/deletions
(INDELs), structure variations (SVs) and other information
variations (8,9), to elucidate its molecular basis, and
to aid the development of effective targeted therapies.
In-depth studies of mitochondria identified
mitochondrial DNA (mtDNA) at the basal membrane of mitochondria,
which is a type of special and unique genetic material, as it is
located outside the nuclei (10).
Though mtDNA has independent genetic function, the majority of
proteins (including the outer mitochondrial membrane and the matrix
proteins) are still encoded by nuclear genome (nDNA), among which
~1,500 proteins have important roles in maintaining mitochondrial
functions (11). The present study
performed high-throughput sequencing on two ESCC cell lines to
determine the common features of genomic variation in ESCC cells,
identify ESCC-associated abnormally expressed nDNA and mtDNA, and
determine their interactions that have important roles in the
occurrence and development of ESCC, thus providing direction for
ESCC basic research and an important experimental basis for the
clinical treatment of ESCC gene targeting therapies.
Materials and methods
Cell recovery and culture
The Ec9706 cell line was provided by the National
Key Laboratory of Molecular Oncology, Chinese Academy of Medical
Sciences (Beijing, China) and the Eca109 cell line was provided by
the laboratory, School of Pharmacy, Zhengzhou University
(Zhengzhou, China). The Ec9706 and Eca109 cells cryopreserved in
liquid-nitrogen were quickly removed and placed in a water bath at
37°C for l-2 min thawing. The cell suspension was then diluted with
an appropriate amount of RPMI-1640 medium containing 10% fetal
bovine serum (both from Beijing Solarbio Science and Technology
Co., Ltd., Beijing, China). Following centrifugation at 173 × g at
room temperature for 5 min, the supernatant was removed, and medium
was added; this process was repeated twice. The cells were diluted
in an appropriate amount of culture medium containing 10% fetal
bovine serum, the cells were seeded into culture flasks for the
culture at 37°C and 5% CO2. The culture medium was
replaced the next day, and subsequently the cells were routinely
cultured.
DNA extraction
Genomic DNAs was extracted according to the
instructions of the DNA extraction kit (Shanghai Genmed
Pharmaceutical Technology Co., Ltd., Shanghai, China).
The concentration and purity of the genomic DNA
extracted from each sample were determined three times using
ultra-trace spectrophotometry (Multiskan MK3; Thermo Fisher
Scientific, Inc., Waltham, MA, USA) with pure water as the control,
and the average of the three measurements was used for the
calculation. For pure genomic DNA, the ratio of absorbance at 260
nm/absorbance at 280 nm should be close to 1.8 (>1.9, indicating
RNA contamination; <1.6, indicating protein or phenol
pollution).
High-throughput sequencing
The high-throughput sequencing process was performed
by Genewiz, Inc. (Beijing, China). Workflow of the experiment is
presented in Fig. 1. Genomic DNA
was tested and randomly broken into manageable fragments by
ultrasound to facilitate the construction of an insert library. In
human genome re-sequencing, paired-end libraries with a span size
of 400–500 bp were usually adopted. The template DNA fragments of
the constructed libraries were hybridized to the surface of flow
cells and amplified to form clusters, and then sequenced with the
Illumina HiSeq X sequencing system (Illumina, Inc., San Diego, CA,
USA). Currently, read length of 150 bp paired-end sequencing
strategy is used in high-throughput whole genome sequencing.
Detected SNPs and INDELs were annotated against a
collection of comprehensive functional annotation databases,
including 1000 Genomes Project, Cosmic, GWAS, PolyPhen-2, VISIFT,
NHLBI, ClinVar, NCI60, YH genome, gene/protein structure, germline
variations (dbSNP; https://www.ncbi.nlm.nih.gov/projects/SNP/), and
functional elements (transcription binding sites, microRNA targets,
conserved elements). Structural variations were detected and
annotated by the chromosomal locations. Copy number variations
(CNV) were also calculated based on the read depth and summarized
in detail. WGC024118D and WGC024119D were the sequencing IDs of
Ec9706 and Eca109 cells, respectively.
Data analysis
The sample passed sample quality control and yielded
enough high quality sequencing data with an average 117.222G raw
data after Illumina pass filtering (PF). The passing filter was a
default standard processing of Illumina HiSeq sequencers, to remove
any reads that do not meet the overall quality as measured by the
Illumina chastity filter. The chastity of a base call was
calculated as the ratio of the brightest intensity divided by the
sum of the brightest and second brightest intensities. Clusters
passed filter if no more than one base call in the first 25 cycles
had a chastity of <0.6.
A comprehensive annotation package, including 10
different databases, including 1000 Genomes Project (http://www.internationalgenome.org), dbSNP
(https://www.ncbi.nlm.nih.gov/projects/SNP/), Cosmic
(http://cancer.sanger.ac.uk/cosmic),
GWAS (https://www.ebi.ac.uk/gwas/), PolyPhen-2
(http://genetics.bwh.harvard.edu/pph2/), VISIFT
(http://sift.jcvi.org/), NCI60 (http://genome-www.stanford.edu/nci60/index.shtml),
ClinVar (https://www.ncbi.nlm.nih.gov/clinvar/), YHgenome
(http://www.yhdatabase.com/), and
National Heart, Lung, and Blood Institute (https://www.nhlbi.nih.gov), was applied to annotate
the bioinformatics analysis of identified SNPs, INDELs, CNVs and
SVs from the sequencing data.
Results
Summary of bioinformatics
analysis
The PF sequencing reads were mapped to the human
genome version 19 (hg19) using Burrows-Wheeler Aligner (12). On average, 98.99% of whole genome
regions were sequenced and a 39.35-fold mappable-reads-coverage was
achieved in this sample set (Table
I).
 | Table I.Data coverage analysis. |
Table I.
Data coverage analysis.
| Quality control
statistics sample | WGC024118D | WGC024119D |
|---|
| Paired-end read
length | 150*2 | 150*2 |
| Total effective data
yield (Gb) | 116.434 | 118.01 |
| Total reads number
(M) | 776.23 | 786.73 |
| Reads mapping
rate | 99.04% | 98.70% |
| Properly paired
mapping reads rate | 96.97% | 96.81% |
| No-mismatch mapping
reads rate | 61.04% | 60.11% |
| Mismatch alignment
bases rate | 0.77% | 0.74% |
| Mean coverage
sequencing depth | 39.1 | 39.6 |
| Reference genome
coverage | 98.99% | 98.99% |
| Reference genome
coverage ≥4X | 98.53% | 98.53% |
| Reference genome
coverage ≥10X | 97.21% | 97.28% |
| Reference genome
coverage ≥20X | 90.00% | 90.49% |
| Polymerase chain
reaction duplication rate | 8.62% | 8.64% |
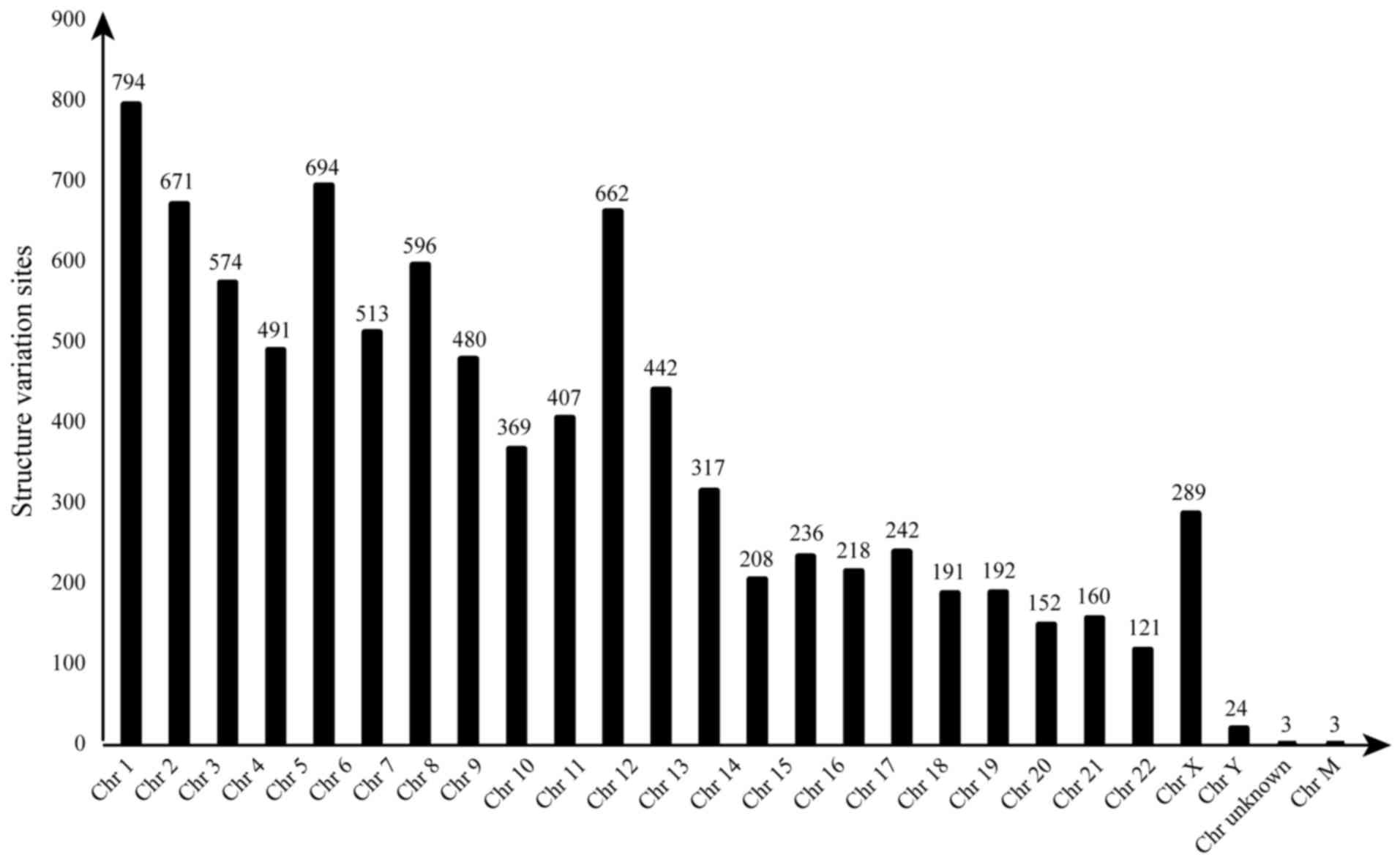
SV detection and annotation
The categories of SV include insertions,
duplications, deletions, inversions, recurring mobile elements and
other rearrangements, typically defined as those covering 50 or
more base pairs (Fig. 2). A total
of 187 structural variants were identified, including seven
inter-chromosomal translocations, three intra-chromosomal
translocations, zero inversion, 177 large insertions or deletions.
Although many SVs were detected, the majority of SVs may be common
SVs present in normal samples, or false detections because of the
limitations of NGS (Fig. 3).
CNV and annotation
As presented in Fig.
4, >40% genes exhibited gain or loss CNVs, and the main form
of CNV in WGC024118Ds was loss, and in WGC024119D it was gain.
Almost half genes on chromosome 9 (Chr 9) and 11 in WGC024118D and
WGC024119Ds had gain or loss, and nearly half genes on Chr 10 in
these two samples had gain or loss.
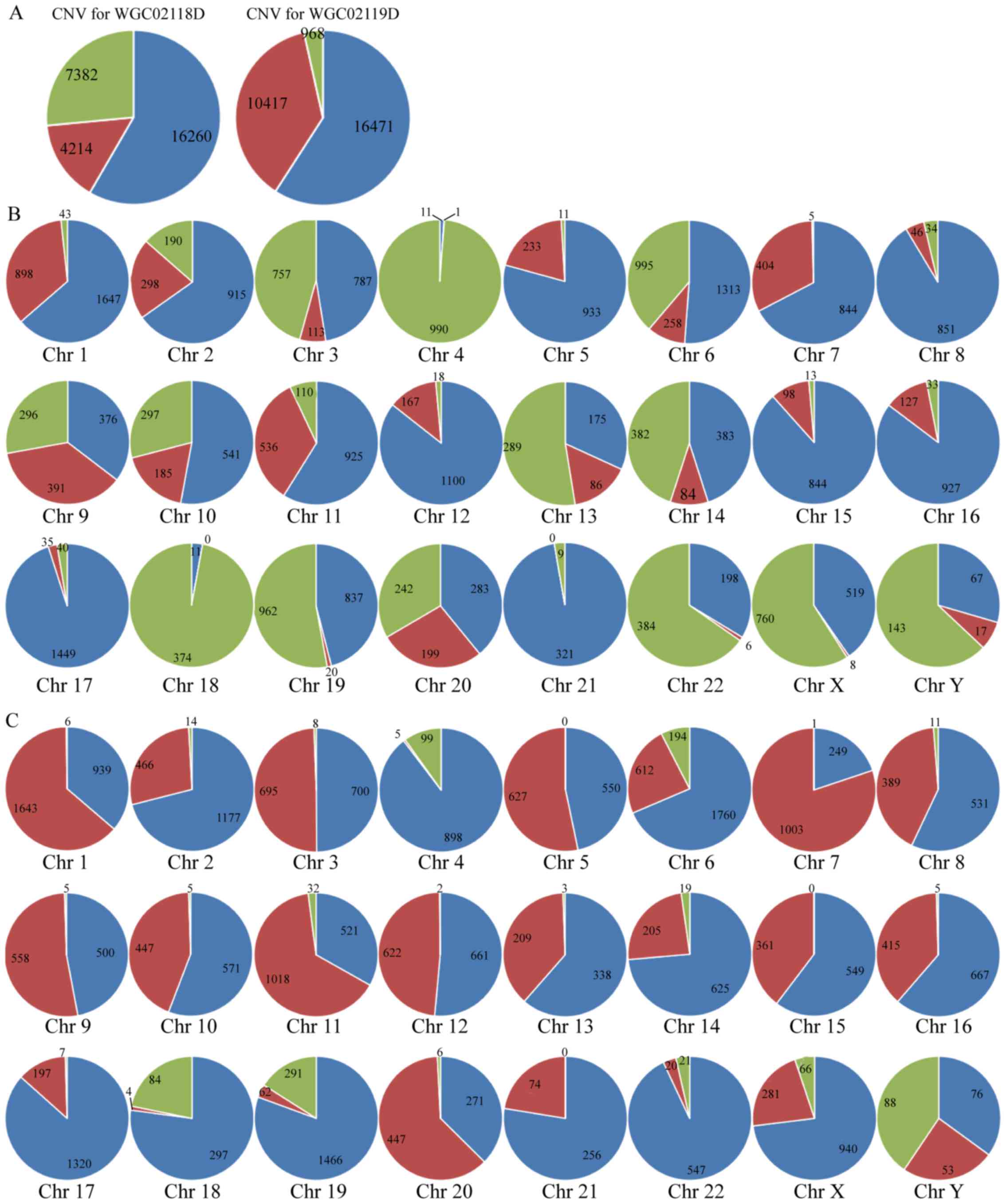 | Figure 4.CNVs in the Ec9706 (WGC024118D) and
Eca109 (WGC024119D) genome. (A) CNVs in the whole genome; the CNV
change trend in the whole genome is that loci with gain or loss
account for >40% of the entire sequence sites, the main form of
CNV in WGC024118D is loss, and for WGC024119D is gain. Red
indicates loss; green indicates gain; and blue indicates sequence
matching the reference hg19. (B) CNVs in WGC024118D and (C)
WGC024119D. Almost all of the loci on Chr 4, 17, and 18 in
WGC024118D have gain or loss; more than half loci on Chr 3, 9, 11,
13, 14, 19 and 20 in WGC024118D, and Chr 1, 5, 7, 9, 11, 20 and Y
in WGC024119D have gain or loss; almost half of loci on Chr 6, 10,
22, X and Y in WGC024118D and Chr 3, 8, 10, 12, 13, 15, and 16 in
WGC024119D have gain or loss; few loci on Chr 8, 12, 15, 16, and 17
in WGC024118D and Chr 4 and 22 in WGC024119D have gain or loss,
maintaining nearly normal sequence. Red indicates gain; green
indicates loss; and blue indicates normal sequence. Chr,
chromosome. |
Functional annotations of short
INDEL
Loci with INDEL in these two ESCC cell lines
predominantly occurred in the upstream, downstream, exon and intron
as well as among genes, among which inter-gene INDEL accounted for
the majority (Table II).
| Table II.Summary of INDEL detection
results. |
Table II.
Summary of INDEL detection
results.
| Feature | WGC024118D | WGC024119D |
|---|
| High-confidence INDEL
no. | 740783 | 766563 |
| Deletion | 392984 | 408253 |
| Insertion | 347799 | 358310 |
| Heterozygotes | 386393 | 409200 |
| Homozygotes | 327829 | 330279 |
| dbSNP | 455370 (61.5%) | 465321 (60.7%) |
| 1000 Genomes
Project | 237781 (32.1%) | 239886 (31.3%) |
| NA | 17882 | 18327 |
| 3′UTR | 6195 | 6354 |
| 5′UTR | 810 | 865 |
| 3′ UTR5, UTR3 | 6 | 3 |
| Downstream | 5402 | 5600 |
| Exonic | 615 | 645 |
| Exonic,
splicing | 3 | 2 |
| Intergenic | 403796 | 419365 |
| Intronic | 270105 | 277821 |
| ncRNA 3′UTR | 127 | 128 |
| ncRNA 5′UTR | 23 | 26 |
| ncRNA 5′UTR, ncRNA
3′UTR | 1 | 1 |
| ncRNA exonic | 1241 | 1299 |
| ncRNA intronic | 29459 | 30801 |
| ncRNA splicing | 13 | 14 |
| Splicing | 91 | 95 |
| Upstream | 4851 | 5049 |
| Upstream,
downstream | 163 | 168 |
| Exon frameshift
deletion | 99 | 108 |
| Exon frameshift
insertion | 87 | 89 |
| Exon nonframeshift
deletion | 189 | 194 |
| Exon nonframeshift
insertion | 148 | 158 |
| Exon stopgain
SNV | 7 | 8 |
| Exon stoploss
SNV | 1 | 2 |
| Exon unknown | 87 | 88 |
As demonstrated in Table III, the bases with short insert
or deletion ranged between 1–20 nt, and the mitochondrial proteins
mutated include mitochondrial electron respiratory chain-related
proteins, such as NADH:Ubiquinone oxidoreductase subunit S5
(NDUFS5), cytochrome P450 family 27 subfamily A member 1 (CYP27A1),
fatty acid metabolism-associated proteins, such as
3-hydroxyisobutyryl-CoA hydrolase (HIBCH), acyl-CoA dehydrogenase
family member 9 (ACAD9), ATP energy generation-associated proteins,
such as ATP synthase mitochondrial F1 complex assembly factor 1
(ATPAF1), and these proteins are all nDNA-encoding
intra-mitochondrial-transporting proteins.
| Table III.Genetic screening of nuclear DNA with
INDEL. |
Table III.
Genetic screening of nuclear DNA with
INDEL.
| Chr | Start | End | Reference
sequence | Sequence
alteration | QUALITY | Alteration ratio
(%) | Variation type | Gene location | Gene | Name
(description) |
|---|
| Chr 1 | 39500839 | 39500839 | C | – | 841.73 | 100 | Deletion | Downstream | NDUFS5 | NADH:Ubiquinone
oxidoreductase subunit S5, (nuclear gene encoding mitochondrial
protein, transcript variant 2) |
| Chr 1 | 45804431 | 45804431 | – | T | 819.73 | 100 | Insertion | Intronic | MUTYH | mutY DNA
glycosylase, (nuclear gene encoding mitochondrial protein,
transcript variant α3) |
| Chr 2 | 191091321 | 191091334 | CAAAAAAAAAAAAA | – | 1600.73 | 100 | Deletion | Intronic | HIBCH |
3-hydroxyisobutyryl-CoA hydrolase (nuclear
gene encoding mitochondrial protein, transcript variant 2) |
| Chr 2 | 219652424 | 219652424 | – | CCTCTTACCTG | 3035.73 | 100 | Insertion | Intronic | CYP27A1 | Cytochrome P450
family 27 subfamily A member 1 (nuclear gene encoding mitochondrial
protein) |
| Chr 3 | 179339343 | 179339343 | – | GGTCTCGG | 1811.73 | 100 | Insertion | Intronic | NDUFB5 | NADH:Ubiquinone
oxidoreductase subunit B5 (nuclear gene encoding mitochondrial
protein, transcript variant 1) |
| Chr 3 | 128614563 | 128614563 | – | CTC | 2388.73 | 100 | Insertion | Intronic | ACAD9 | acyl-CoA
dehydrogenase family member 9 (nuclear gene encoding mitochondrial
protein, transcript variant 1) |
| Chr 4 | 106312189 | 106312189 | – | C | 806.73 | 100 | Insertion | Intronic | PPA2 | Pyrophosphatase
(inorganic) 2 (nuclear gene encoding mitochondrial protein,
transcript variant 1) |
| Chr 4 | 89197868 | 89197875 | GACTGTCC | – | 1078.74 | 100 | Deletion | Intronic | PPM1K | Protein
phosphatase, Mg2+/Mn2+ dependent, 1K (nuclear
gene encoding mitochondrial protein) |
| Chr 1 | 29538224 | 29538225 | CT | – | 1835.73 | 100 | Deletion | Intronic | MECR | Mitochondrial
trans-2-enoyl-CoA reductase (nuclear gene encoding
mitochondrial protein) |
| Chr 1 | 47107042 | 47107042 | – | G | 1521.73 | 100 | Insertion | Intronic | ATPAF1 | ATP synthase
mitochondrial F1 complex assembly factor 1 (nuclear gene encoding
mitochondrial protein) |
Functional annotations of SNP
As presented in Table
IV, the loci with SNP features in these two ESCC cell lines are
located in the upstream, downstream, exon and intron, and within
genes. The majority of loci with SNP features were located in genes
and in exons.
| Table IV.Summary of SNP detection results. |
Table IV.
Summary of SNP detection results.
| Feature | WGC024118D | WGC024119D |
|---|
| High-confidence SNP
no. | 3851527 | 3861523 |
| Heterozygotes | 1938738 | 1944372 |
| Homozygotes | 1911032 | 1915229 |
| dbSNP | 3685859
(95.7%) | 3694088
(95.7%) |
| 1000 Genomes
Project | 3520593
(91.4%) | 3520791
(91.2%) |
| 3′UTR | 25886 | 25946 |
| 5′UTR | 5578 | 5570 |
| 5′UTR, 3′UTR | 10 | 12 |
| Downstream | 23646 | 23744 |
| Exonic | 23196 | 23198 |
| Exonic,
splicing | 6 | 7 |
| Intergenic | 2255882 | 2264134 |
| Intronic | 1327100 | 1327665 |
| ncRNA 3′UTR | 615 | 618 |
| ncRNA 5′UTR | 97 | 105 |
| ncRNA exonic | 9941 | 10103 |
| ncRNA intronic | 156029 | 156787 |
| ncRNA splicing | 52 | 55 |
| Splicing | 71 | 68 |
| Upstream | 22691 | 22772 |
| Upstream,
downstream | 727 | 739 |
| Exon nonsynonymous
SNV | 10720 | 10774 |
| Exon stopgain
SNV | 98 | 92 |
| Exon stoploss
SNV | 15 | 13 |
| Exon synonymous
SNV | 11962 | 11915 |
| Exon unknown | 407 | 411 |
Additionally, the mitochondrial protein genes with
SNP features were mostly located in exons (Table IV), including nDNA-encoding
intra-mitochondrial-transporting proteins, such as mitochondrial
trans-2-enoyl-CoA reductase (MECR), fatty acid
metabolism-associated proteins, such as
3-hydroxymethyl-3-methylglutaryl-CoA lyase (HMGCL), HIBCH, and
MECR, ATP energy generation-associated proteins, such as ATPAF1,
and succinate dehydrogenase complex. All these proteins are
nDNA-encoded intra-mitochondrial-transporting proteins.
Discussion
Oncogenes and tumor suppressor genes have important
roles in the occurrence and development of tumors, Changes to mtDNA
in tumor tissue has received increasing research interest. It has
been previously indicated that various solid tumors and
hematological malignancies have different mtDNA mutations,
deletions, and microsatellite instabilities at different sites, and
these may be associated with the occurrence of a variety of tumors
(13). In recent years, more
researchers hypothesize that the interactions between mtDNA and
nDNA are involved in the tumorigenic process (14). Compared with nDNA, mtDNA has higher
mutation rate due to the poor proofreading ability of
mtDNA-replicating DNA polymerase and the environment with high
concentrations of reactive oxygen species (ROS). Furthermore, mtDNA
has hologynic features, in which mutations accumulate continuously
along the matriarchal line, and the accumulation of mutations
increases the risk of tumorigenesis (9). In addition, mtDNA has more
polymorphisms (15), appearing as
significant sequence differences among different races and in
different regions. The function of mtDNA is known to be involved in
tumorigenesis in head and neck cancer (16), bladder cancer (10), breast cancer (17) and lung cancer (11). Hu et al (18) also identified gastric cancer and
esophageal cancer-associated genes using whole-genome sequencing,
including p53, Janus kinase 3, BRCA2, fibroblast growth factor 2,
F-box and WD repeat domain containing 7, mutS homolog 3, patched 1,
neurofibromin 1, ErbB-2 receptor tyrosine kinase 2 and checkpoint
kinase 2, and identified number of novel potential
cancer-associated genes, including KISS1 receptor, anti-Mullerian
hormone, motor neuron and pancreas homeobox 1, WNK lysine deficient
protein kinase 2, THAP domain containing 12; furthermore, certain
chromosomes were reported to have mutations in >30% of tumor
genes, including MACRO domain containing 2, fragile histidine triad
(FHIT) and parkin RBR E3 ubiquitin protein ligase (18). However, the application of whole
genome sequencing to detect mitochondrial changes in ESCC cell
lines has rarely been reported previously. Thus, identifying nDNA
and mtDNA mutation involved the occurrence and development of ESCC,
and investigating their impact on the occurrence and development of
ESCC is important for developing novel ESCC gene targeting
therapies.
Detecting SVs can help to investigate the mechanisms
of gene mutation in cancer, understand the biological differences
and identify novel therapeutic targets. However, the complexities
and mutation mechanisms of SVs throughout the whole ESCCs genome
are still unclear. Cheng et al (19) reported chromosomal abnormalities in
malignant metastatic ESCCs and the function of SVs-derived target
genes in these abnormal chromosomes are diverse, indicating that
the SVs map of the whole genome important for the prevention,
diagnosis and treatment of ESCCs. The present study detected many
SVs; however, most of the SVs may be common SVs present in normal
samples, or the false detections because of the limitation of NGS.
The categories of structural variants include insertions,
duplications, deletions, inversions, recurring mobile elements, and
other rearrangements. A total of 187 SVs were identified in the
current study, including seven inter-chromosomal translocations,
three intra-chromosomal translocations and 177 large insertions or
deletions. The loci with SVs on Chr 1–12 of the two ESCC cell lines
were ≥5%, whereas those with SVs on Chr 13–22, X and Y were ≤3%.
The SVs of mitochondrial genes detected in this study may provide
important information for further studies into the nDNA and mtDNA
mutations, and roles in the occurrence and development of ESCC.
DNA copy number alterations in tumor cells are key
genetic events in the development and progression of human cancers
(20). These alterations are
typically the result of genomic events producing gains and losses
of chromosomes or chromosomal regions. Losses and gains of DNA can
contribute to alterations in the expression of tumor suppressor
genes and oncogenes. Therefore, identifying DNA copy number
alterations in tumor genomes may help to identify critical genes
associated with cancer and, eventually, to improve therapeutic
approaches. Errors during mitosis and meiosis can result in
duplications or deletions of genes on a chromosomal level. These
differences are termed CNV, which may profoundly affect health and
lead to various disorders. In this study, CNVs were identified
based on read depth. Miyawaki et al (21) investigated the important roles of
MYC and FHIT gene CNV in selecting the optimal treatment strategy
in strategy in resected ESCC patients. The detection results of CNV
the current study demonstrated that >40% of genes exhibited gain
or loss. Almost half of loci on Chr 9 and 11 in WGC024118D and
WGC024119D exhibited gain or loss, and nearly half loci of on Chr
10 exhibited gain or loss, indicating that detecting CNV in ESCC is
important for identifying the oncogenic gene gains of the genome
and in genes involved in the development of tumors.
The results of SNPs and INDELs sequencing
demonstrate the potential functional impact. Cao et al
(22) used the whole exon
sequencing and revealed lots of genetic heterogeneities within
ESCC. Xu et al (23)
previously investigated the polymorphisms of ESCC phosphatase and
tensin homolog gene in the Chinese Han population, and the
interactions among genes. Similarly, detecting short insertion and
deletion in genes was applied to detect the gene mutations of ESCC
(24,25). The detection results of SNPs and
INDELs in the current study revealed potential adverse alterations
that have potentially important functions and are associated with
gene regulation, including transcription factor binding, microRNA
target, conserved elements. The inter-gene loci occurred INDEL and
had SNP features account for the majority, and these genes-encoded
proteins include mitochondrial electron respiratory
chain-associated proteins, such as CYP27A1, fatty acid
metabolism-associated proteins, such as HIBCH and ACAD9,
nDNA-encoding intra-mitochondrial-transporting proteins, such as
MECR, and ATP energy generation-related proteins, such as ATPAF1.
The sequencing results also demonstrate that the detection of SNPs
and INDELs can be used as the basis for the development of gene
therapy.
Under certain conditions, mtDNA can also have
effects on nDNA due to mitochondrial defects, thus resulting in
corresponding gene expression changes; this process is termed
reverse regulation (26). This
reverse regulation is associated with disturbances in nDNA
expressions, mtDNA mutations, respiratory function and
mitochondrial protein synthesis inhibition. This reverse regulation
is also involved in the process of apoptosis, during which
mitochondrial damages can cause the increasing of mitochondrial
membrane permeability; such apoptosis-inducing factors as
cytochrome c, apoptotic protease activating factor-1 and
apoptosis inducing factor are then released from mitochondria, and
can be directly transported to the nuclei, thus inducing the
expressions of certain genes in the nuclei and triggering the
apoptotic cascade. It is also suggested that the reverse regulation
may also be associated with the ROS pathway or the ratio of ATP/ADP
(26). In the present study, the
identified nDNA-encoded mtDNA with SVs, CNV, SNPs and INDELs
provides a good experimental basis and ideas for investigating the
relationships between nDNA and mtDNA in the occurrence and
development of ESCC.
Tumorigenesis not only depends on intranuclear
genetic materials, but also is also closely associated with
ectonuclear mtDNA. As the biological metabolism and energy
conversion center, mitochondrial genome replication, gene
expression, respiratory chain function, and
mitochondrion-associated cell function are inextricably associated
with nDNA. nDNA and mtDNA changes will cause corresponding changes
of mitochondrial functions. The transcription, replication and
translation of mtDNA need a variety of different nDNA products, and
the disorders of nDNA products are also associated with mtDNA
mutations and biosynthesis inhibition of mitochondrial proteins.
These changes can lead to various diseases, and play vital roles
particularly in the occurrence and development of tumors. Thus,
nDNA-encoded mtDNA protein variants in Ec9706 and Eca109 detected
in this study using high-throughput sequencing can provide very
reliable theoretical basis and data support for the gene targeting
therapies of ESCC.
In this study, the identified mtDNA with abnormal
transcription, duplication and translation in Ec9706 and Eca109
cells by high-throughput sequencing were closely associated with
the disorders of nDNA products. This study into these
nDNA-associated mtDNA and the interaction between the two is
important for basic research for ESCC, pointed out the direction
and provided a very reliable theoretical basis for the gene
targeting therapies of ESCC in clinical.
Acknowledgements
This study was supported by the First Batch of
Science and Technology Plan Projects of Zhengzhou in 2013 (grant
no. 131PCXTD628), the Fundamental and Advanced Technology Research
Project of Henan Province (grant no. 132300410409) and the Medical
Science and Technology Plan Program Grant of Henan Province (grant
no. 201401009).
References
|
1
|
Zhao P, Dai M, Chen W and Li N: Cancer
trends in China. Jpn J Clin Oncol. 40:281–285. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Ell C and Lorenz D: Diagnosis and
treatment of oesophageal carcinoma: Changes in every respect.
Viszeralmedizin. 31:3142015.PubMed/NCBI
|
|
3
|
Shigaki H, Baba Y, Watanabe M, Murata A,
Ishimoto T, Iwatsuki M, Iwagami S, Nosho K and Baba H: PIK3CA
mutation is associated with a favorable prognosis among patients
with curatively resected esophageal squamous cell carcinoma. Clin
Cancer Res. 19:2451–2459. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Kasar S, Kim J, Improgo R, Tiao G, Polak
P, Haradhvala N, Lawrence MS, Kiezun A, Fernandes SM, Bahl S, et
al: Whole-genome sequencing reveals activation-induced cytidine
deaminase signatures during indolent chronic lymphocytic leukaemia
evolution. Nat Commun. 6:88662015. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Knoppers BM, Zawati MH and Sénécal K:
Return of genetic testing results in the era of whole-genome
sequencing. Nat Rev Genet. 16:553–559. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Choi M, Scholl UI, Ji W, Liu T, Tikhonova
IR, Zumbo P, Nayir A, Bakkaloğlu A, Ozen S, Sanjad S, et al:
Genetic diagnosis by whole exome capture and massively parallel DNA
sequencing. Proc Natl Acad Sci USA. 106:pp. 19096–19101. 2009;
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Ng SB, Turner EH, Robertson PD, Flygare
SD, Bigham AW, Lee C, Shaffer T, Wong M, Bhattacharjee A, Eichler
EE, et al: Targeted capture and massively parallel sequencing of 12
human exomes. Nature. 461:272–276. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Frank K, Barta E, Bana NÁ, Nagy J, Horn P,
Orosz L and Stéger V: Complete mitochondrial genome sequence of a
Hungarian red deer (Cervus elaphus hippelaphus) from
high-throughput sequencing data and its phylogenetic position
within the family Cervidae. Acta Biol Hung. 67:133–147. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Brunelle JK, Shroff EH, Perlman H,
Strasser A, Moraes CT, Flavell RA, Danial NN, Keith B, Thompson CB
and Chandel NS: Loss of Mcl-1 protein and inhibition of electron
transport chain together induce anoxic cell death. Mol Cell Biol.
27:1222–1235. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Yang Ai SS, Hsu K, Herbert C, Cheng Z,
Hunt J, Lewis CR and Thomas PS: Mitochondrial DNA mutations in
exhaled breath condensate of patients with lung cancer. Respir Med.
107:911–918. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Cheng M, Guo Z, Li H, Li Z, Li C and Geng
C: Identification of sequence polymorphisms in the mitochondrial
displacement loop as risk factors for sporadic and familial breast
cancer. Tumour Biol. 35:4773–4777. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Li H and Durbin R: Fast and accurate
long-read alignment with Burrows-Wheeler transform. Bioinformatics.
26:589–595. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Hiyama T, Tanaka S, Shima H, Kose K,
Tuncel H, Ito M, Kitadai Y, Sumii M, Yoshihara M, Shimamoto F, et
al: Somatic mutation in mitochondrial DNA and nuclear
microsatellite instability in gastric cancer. Oncol Rep.
10:1837–1841. 2003.PubMed/NCBI
|
|
14
|
Liang BC: Evidence for association of
mitochondrial DNA sequence amplification and nuclear localization
in human low-grade gliomas. Mutat Res. 354:27–33. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Soares VY, Silva JC, Silva KR, Pires e
Cruz Mdo S, Santos MP, Ribolla PE, Alonso DP, Coelho LF, Costa DL
and Costa CH: Identification of blood meal sources of Lutzomyia
longipalpis using polymerase chain reaction-restriction fragment
length polymorphism analysis of the cytochrome B gene. Mem Inst
Oswaldo Cruz. 109:379–383. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Montebugnoli L, Leonardi E, Morandi L,
Farnedi A, Gissi DB, Marchetti C, Tarsitano A, Balbi T, Gentile L,
Cocchi R and Foschini MP: Genetic relationship between multiple
squamous cell carcinomas arising in the oral cavity. Head Neck.
36:94–100. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Shakhssalim N, Houshmand M, Kamalidehghan
B, Faraji A, Sarhangnejad R, Dadgar S, Mobaraki M, Rosli R and
Sanati MH: The mitochondrial C16069T polymorphism, not
mitochondrial D310 (D-loop) mononucleotide sequence variations, is
associated with bladder cancer. Cancer Cell Int. 13:1202013.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Hu N, Kadota M, Liu H, Abnet CC, Su H, Wu
H, Freedman ND, Yang HH, Wang C, Yan C, et al: Genomic landscape of
somatic alterations in esophageal squamous cell carcinoma and
gastric cancer. Cancer Res. 76:1714–1723. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Cheng C, Zhou Y, Li H, Xiong T, Li S, Bi
Y, Kong P, Wang F, Cui H, Li Y, et al: Whole-genome sequencing
reveals diverse models of structural variations in esophageal
squamous cell carcinoma. Am J Hum Genet. 98:256–274. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Gough DJ, Corlett A, Schlessinger K,
Wegrzyn J, Larner AC and Levy DE: Mitochondrial STAT3 supports
RAS-dependent oncogenic transformation. Science. 324:1713–1716.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Miyawaki Y, Kawachi H, Ooi A, Eishi Y,
Kawano T, Inazawa J and Imoto I: Genomic copy-number alterations of
MYC and FHIT genes are associated with survival in esophageal
squamous-cell carcinoma. Cancer Sci. 103:1558–1566. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Cao W, Wu W, Yan M, Tian F, Ma C, Zhang Q,
Li X, Han P, Liu Z, Gu J and Biddle FG: Multiple region whole-exome
sequencing reveals dramatically evolving intratumor genomic
heterogeneity in esophageal squamous cell carcinoma. Oncogenesis.
4:e1752015. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Xu X, Chen G, Wu L and Liu L: Association
of genetic polymorphisms in PTEN and additional gene-gene
interaction with risk of esophageal squamous cell carcinoma in
Chinese Han population. Dis Esophagus. 29:944–949. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Doucet-O'Hare TT, Sharma R, Rodić N,
Anders RA, Burns KH and Kazazian HH Jr: Somatically acquired LINE-1
insertions in normal esophagus undergo clonal expansion in
esophageal squamous cell carcinoma. Hum Mutat. 37:942–954. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Abnet CC, Huppi K, Carrera A, Armistead D,
McKenney K, Hu N, Tang ZZ, Taylor PR and Dawsey SM: Control region
mutations and the ‘common deletion’ are frequent in the
mitochondrial DNA of patients with esophageal squamous cell
carcinoma. BMC Cancer. 4:302004. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Rafael G and Carmen G: Animal
mitochondrial biogenesis and function: A regulatory cross-talk
between two genomes. Gene. 263:1–16. 2001. View Article : Google Scholar : PubMed/NCBI
|