Introduction
Liver fibrosis, characterized by excessive
deposition of extracellular matrix (ECM) and distortion of the
normal liver architecture, occurs as a result of liver injuries,
such as those induced by chronic hepatitis virus, ethanol, toxic
reagents or drugs (1). Liver
fibrosis, considered to be a serious global health problem, may
eventually develop into liver cirrhosis and, in certain cases,
liver cancer, and is thus associated with high morbidity and
mortality (2). Toxic reagents,
predominantly the metabolic products of toxicants, reactive oxygen
species, inflammatory cytokines and damage to DNA repair
mechanisms, are among the common risk factors for the development
of liver injury and fibrosis (3).
Carbon tetrachloride (CCl4) is widely known for its
toxic effects, resulting in liver injury and liver fibrosis.
Furthermore, CCl4 has been extensively used in studies
concerning liver fibrosis (4).
However, the exact molecular mechanisms underlying the toxic
effects of CCl4 have not yet been determined.
Long noncoding RNAs (lncRNAs) are commonly defined
as RNA molecules with a length >200 nucleotides with limited or
no protein-coding capacity (5).
Increasing evidence has revealed that lncRNAs are involved in
numerous biological processes, including cell proliferation,
apoptosis, cell cycle, cell differentiation (6–8),
nervous system development (9) and
cancer metastasis (10).
Furthermore, lncRNAs affect gene expression via the regulation of
transcriptional, post-transcriptional and/or epigenetic mechanisms
(11,12). Numerous studies have indicated the
involvement of several lncRNAs in liver fibrosis, including
Alu-mediated CDKN1A/p21 transcriptional regulator (13), metastasis-associated lung
adenocarcinoma transcript 1 (14),
maternally expressed 3 (15) and
growth arrest-specific 5 (16).
Despite these previous findings, other lncRNAs involved in the
occurrence and development of liver fibrosis require
identification.
High-throughput genome screening provides a
comprehensive picture of gene expression under pathological
conditions and may contribute novel insights into the mechanisms
underlying liver fibrosis. Rapid developments in systems biology
and bioinformatics means that the biological process and disease
complexity on a systems level can be elucidated (17).
Therefore, in order to determine potential
regulatory lncRNAs and investigate the expression pattern of
lncRNAs with regards to the occurrence and development of liver
fibrosis, the present study aimed to profile differentially
expressed lncRNAs and mRNAs in CCl4-induced liver
fibrotic tissue via RNA sequencing. Bioinformatics analyses,
including Gene Ontology (GO) and Kyoto Encyclopedia of Genes and
Genomes (KEGG) pathway analysis, were performed to improve the
classification of differentially expressed mRNAs between the
CCl4-treated and CCl4 untreated liver
tissues. In order to validate the RNA sequencing data, reverse
transcription-quantitative polymerase chain reaction (RT-qPCR) was
performed and a significantly differentially expressed lncRNA,
NR_002155.1, was identified, which, to the best of our knowledge,
has not been previously investigated. The present study aimed to
elucidate further candidate diagnostic biomarkers and treatment
targets for liver fibrosis.
Materials and methods
Rat liver fibrosis induced by
CCl4
Male Sprague-Dawley rats (n=20; 8 weeks old)
weighing 200–220 g were obtained from Guangdong Medical Laboratory
Animal Center (Guangzhou, China). The rats were housed in
polycarbonate cages under standard laboratory conditions at a
temperature of 22°C, a relative humidity of 55% and 12 h light/dark
cycle. Rats were fed with standard laboratory food pellets and
water ad libitum. Rats were housed in these conditions for 7
days prior to CCl4 administration. CCl4 was
purchased from Tianjin Fuyu Fine Chemical Co., Ltd. (Tianjin,
China). Rats were equally divided into two groups by random
sampling: Untreated group and CCl4-treated group. Rats
in the CCl4-treated group were injected
intraperitoneally with a 0.6 mg/kg mixture of CCl4 and
corn oil [1:1 (v/v)] twice a week for 8 weeks. Rats in the control
group were treated with equal volumes of corn oil. The body weight
and clinical symptoms of the rats were observed twice a week until
the termination of the experiment. Blood and liver tissues were
collected from each rat. Blood was collected for serum biochemistry
and cytokine investigation. Liver tissues were sliced into sections
and preserved in 4% paraformaldehyde for 48 h at room temperature
or TRIzol reagent at 4°C (Invitrogen; Thermo Fisher Scientific,
Inc., Waltham, MA, USA) for histological staining and RNA
extraction, respectively. The liver tissues of three rats from each
group were frozen in liquid nitrogen in preparation for RNA
sequencing. The animal experimental procedures in the present study
were performed in accordance with the Guide for Animal Care
(18) and Use and were approved by
the Animal Ethics Committee of Peking University Shenzhen Hospital
(Shenzhen, China).
Cell culture and overexpression of
NR_002155.1 in HSC-T6 cells
The HSC-T6 rat hepatic stellate cell (HSC) cell line
was purchased from the Institute of Cell Research, Chinese Academic
of Sciences (Shanghai, China). The HSC-T6 cells were cultured in
Dulbecco's modified Eagle's medium (DMEM; Gibco; Thermo Fisher
Scientific, Inc.) supplemented with 10% fetal bovine serum (FBS;
Gibco; Thermo Fisher Scientific, Inc.), 100 µg/ml streptomycin and
100 U/ml penicillin. They were incubated at 37°C with an atmosphere
of 5% CO2.
Ad-NR_002155.1 was purchased from Vigene
Biosciences, Inc. (Rockville, MD, USA). The NR_002155.1 sequence
(https://www.ncbi.nlm.nih.gov/nuccore/NR_002155.1?report=fasta)
or nonsense sequence (negative control;
ATGCCAACCACGGCGCCGCCCACAGCACTCCTGCTGCTGGCGGCCGCCCTTTAACCGACAAGGACCTGCGCGGGTGAGTGCCGGGTCCGGAGGGAAATGGCCTCTGAATTAGGGGCGAATTAGGCACCTGGGAAAAAAGGTCTCCACGTCGCACACCAGACCCTGCTCCTTCTCCACCGGCGTCCAGAACCCCGCGCCCTGCTCCCTGCCCGGCCCGCGCTTGGGCCCGGGAAAACGGGAGGAGATCAGGGTTTCACGCGCACCGCCCCCAGGCTCACACTCGGTTCGGTATTTCGTCACCGCGGTGTCCCGGCCCGGCCTCGGTTCACCACGGTTCATCTCTGTCG)
was subcloned into the adenovirus shuttle plasmid vector
pAd-EF1a-GFP (Vigene Biosciences, Inc.). Homologous recombination
was used with regards to the pBHGlox_E1, 3 Cre and shuttle plasmids
in order to generate Ad-NR_002155.1. HSC-T6 cells were subsequently
infected with Ad-NR_002155.1. Cells (1×105) were seeded
in 6 well plates. After 12 h, cells were infected by Ad-NR_002155.1
(multiplicity of infection=60) for 12 h. Cells were subsequently
cultured in normal DMEM with 10% FBS for 48 h prior to seeding for
CCK-8 and RT-qPCR assays. All cell incubations were at 37°C with 5%
CO2.
Detection of alanine aminotransferase
(Alt), aspartate transaminase (Ast), tumor necrosis factor-α
(Tnf-α) and transforming growth factor-β1 (Tgf-β1) expression
Serum was obtained from whole blood by
centrifugation at 12,000 × g for 15 min at 4°C. Alt (cat ml037336),
Ast (cat. no. ml12021), Tnf-α (cat. no. ml002859) and Tgf-β1 (cat.
no. ml002856) in serum were detected by enzyme linked-immunsorbent
assay kits purchased from Shanghai Enzyme-linked Biotechnology Co.,
Ltd. (Shanghai, China) according to the manufacturer's protocols.
The experiments were performed in triplicate.
Histological staining
The liver tissue samples from the rats were fixed in
4% paraformaldehyde at 4°C overnight, embedded in paraffin and cut
into 4 mm sections. The sections were deparaffinized in xylene and
rehydrated in a graded alcohol series (100% ethanol for 5 min, 80%
ethanol for 5 min; both at room temperature). The sectioned tissues
were stained by Hematoxylin and eosin (H&E) staining. In brief,
tissues were incubated with hematoxylin solution for 5 min and
eosin solution for 1 min at room temperature. Masson staining was
performed according to the manufacturer's protocols (Nanjing
SenBeiJia Biological Technology Co., Ltd., Nanjing, China).
Briefly, the sectioned tissues were stained with Celestine blue dye
for 2 min, Mayer's hematoxylin for 2 min, followed by Ponceau S and
acid fuchsin staining for 10 min, all at room temperature. The
sections were photographed with a light microscope at a
magnification of ×100 or ×200.
RNA extraction and RT-qPCR
Total RNA from tissues or cells was isolated using
TRIzol reagent. The purity and concentration of RNA were quantified
using the NanoDrop 2000 spectrophotometer (Thermo Fisher
Scientific, Inc.). High quality isolated RNA with an optical
density (OD) value 260/280>1.90 was determined as the cut-off
value for RNA sequencing and RT-qPCR. Total RNA was reverse
transcribed using a PrimeScript™ RT reagent Kit with gDNA Eraser
(Takara Bio, Inc., Otsu, Japan), according to the manufacturer's
protocol. Gene expression levels were determined via qPCR using the
SYBR Premix Ex Taq™ II kit (Takara Bio, Inc.) in an ABI PRISM 7500
system (Thermo Fisher Scientific, Inc.). According to the
manufacturer's protocol, the thermocycling conditions were 95°C for
5 min, 40 cycles at 95°C for 15 sec, 56°C for 30 sec and 72°C for
15 sec, followed by a final extension at 72°C for 7 min. The
housekeeping gene GAPDH was used for normalization and all
experiments were performed in triplicate. The relative fold-change
of the lncRNAs was calculated using the 2−ΔΔCq method
(19). The primer sequences used
in the present study are listed in Table I.
 | Table I.Rat primer sequences for reverse
transcription-quantitative polymerase chain reaction. |
Table I.
Rat primer sequences for reverse
transcription-quantitative polymerase chain reaction.
| Transcript | Forward primer | Reverse primer |
|---|
| NR_002155.1 |
5′-gatgacccggagcaagtg-3′ |
5′-ctccaggtaggccctctttc-3′ |
| XR_596496.1 |
5′-aaggaagaactcccatattgcat-3′ |
5′-acgtcagggcagaaaatgtc-3′ |
| XR_353556.2 |
5′-ggtgactgacaaagaagagaagc-3′ |
5′-ccaggcacaaggtcaacag-3′ |
| RGD1559724 |
5′-ggaaaccccgttgttttctt-3′ |
5′-tggttaacgtccaggcatc-3′ |
| XR_353723.1 |
5′-tgtggcttctccagaccag-3′ |
5′-tcctgtccttcgttgatgttc-3′ |
| XR_597361.1 |
5′-gcagagagctgtgtctgaacc-3′ |
5′-gcaaatggctgccctaga-3′ |
| XR_591985.1 |
5′-tactctctcctcggatggct-3′ |
5′-gctttctatgtggcctggttc-3′ |
| XR_591987.1 |
5′-cactggagcgtcaatcaaag-3′ |
5′-acttagccccacctccaagta-3′ |
| XR_352490.2 |
5′-caactaagctggacccttgg-3′ |
5′-ggggcctcagtcttgtgag-3′ |
| XR_361902.2 |
5′-tggcgtcagtgaacaacttc-3′ |
5′-acccgggccaaggtatag-3′ |
| Acp1-ps1 |
5′-tgagagaggagaacctggatg-3′ |
5′-gagtcactgtacagcccagga-3′ |
| XR_350045.1 |
5′-acatagatcaaaccccgagaag-3′ |
5′-tacagccagatgctcccagt-3′ |
| GAPDH |
5′-gacggcaagttcaacggcacagt-3′ |
5′-agcggaaggggcggagatgat-3′ |
RNA sequencing
Total RNA isolated from the liver tissues of three
rats from each group were selected for RNA sequencing. In order to
perform high-throughput sequencing, ribosomal RNA (rRNA) was
depleted from total RNA by using the Ribo-Zero rRNA Removal kit
(Epicentre; Illumina, Inc., San Diego, CA, USA) in accordance with
the manufacturer's protocol. cDNA libraries were subsequently
generated using a ScriptSeq v2 RNA-Seq Library Preparation kit
(Epicentre; Illumina, Inc.) and were sequenced on an Illumina HiSeq
3000 with 101 bp paired-end reads at Guangzhou RiboBio Co., Ltd.
(Guangzhou, China). The raw reads were first filtered to remove the
adapter sequences and the low quality sequences were removed by the
FASTX-Toolkit v0.0.13 (hannonlab.cshl.edu/fastx_toolkit/index.html.).
The subsequent clean reads were mapped to the reference genome
Rnor_6.0 (asia.ensembl.org/Rattus_norvegicus/Info/Index)
with TopHat v2.0.13 (20). The
number of reads mapped to each gene was determined using gfold
v1.1.2 (21). Differential
expression was assessed using Cufflinks (22) with fragments/kb of transcript per
million fragments mapped and read counts as the input.
Differentially expressed genes were identified according to the
following criteria: Fold-change >2 and adjusted P-value
<0.001.
GO and KEGG pathway analysis
Differentially expressed mRNAs in fibrotic liver
tissues were determined via GO analysis and KEGG pathway analysis.
With regards to GO analysis (http://geneontology.org/), the aberrantly expressed
mRNAs were classified into three items by enrichment analysis:
Biological process, cellular component and molecular function. KEGG
pathway analysis (www.genome.jp/kegg/) was performed in order to analyze
the potential pathways enriched by the differentially expressed
mRNAs.
Cell Counting Kit-8 (CCK-8) assay
A CCK-8 assay (Dojindo Molecular Technologies, Inc.,
Kumamoto, Japan) was used to perform the cell proliferation assay.
The transfected HSC-T6 cells were plated in 96-well plates at a
density of 1,000 cells/well. Following incubation at 37°C for 12,
24, 48 and 72 h time intervals, the OD values of absorbance were
determined using a microplate reader (Bio-Rad Laboratories, Inc.,
Hercules, CA, USA) at a wavelength of 450 nm, in accordance with
the manufacturer's protocol. All experiments were performed in
triplicate.
Statistical analysis
Data from at least three independent experiments
were presented as the mean ± standard deviation. Student's t-tests
were used to analyze the statistical significance of RT-qPCR, Alt,
Ast, Tnf-α, Tgf-β1 and cell proliferation analyses. Multiple
comparisons of the expression of NR_002155.1 among the three HSC-T6
cell groups were performed using one-way analysis of variance
followed by the Student-Newman-Keuls post-hoc test. P<0.05 was
considered to indicate a statistically significant difference. SPSS
v19.0 software (IBM Corp., Armonk, NY, USA) was used to analyze the
data.
Results
Liver injury and fibrosis
induction
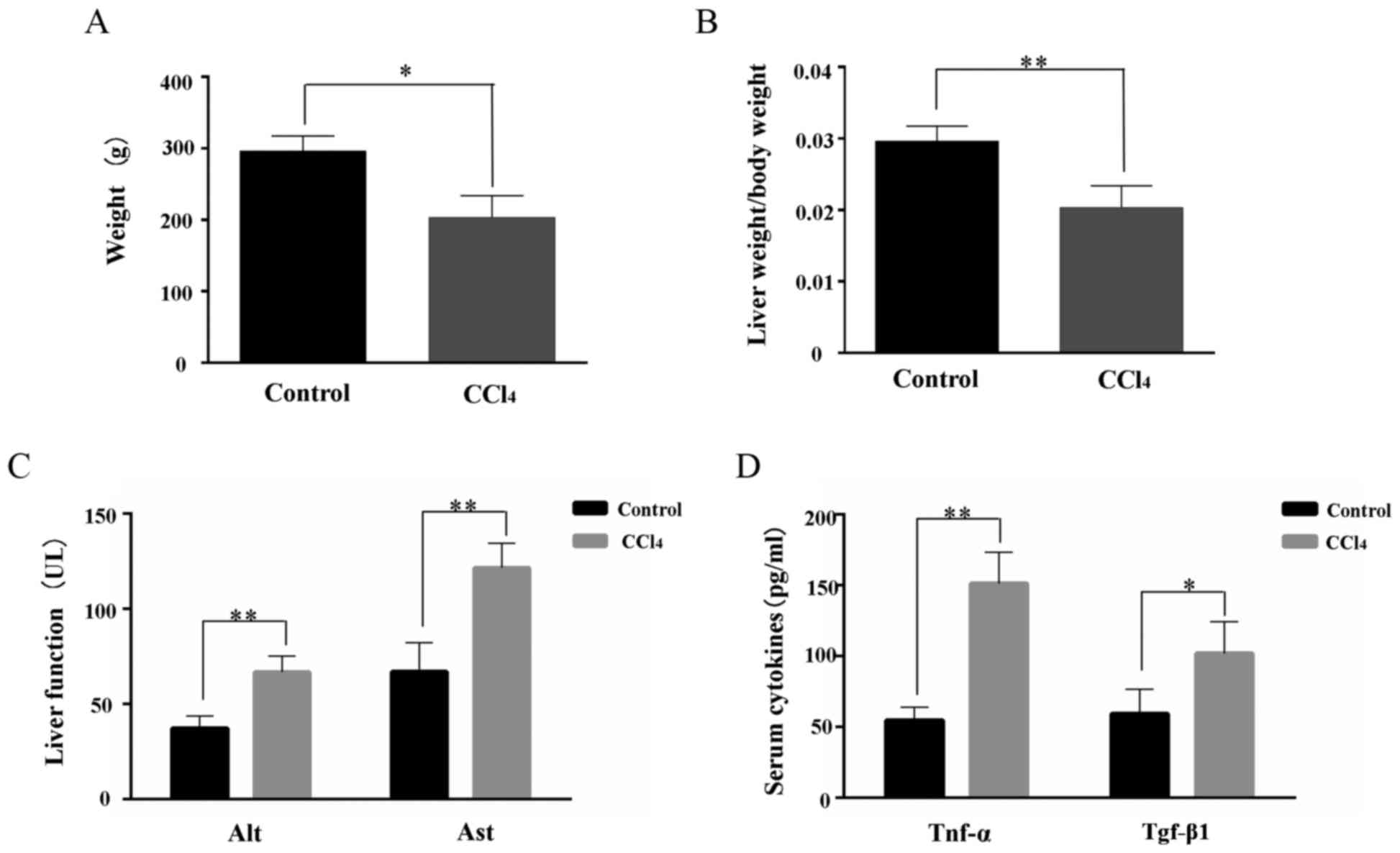
At the termination of the experiment, the body
weight and the liver/body weight ratio were significantly reduced
in the CCl4-treated rats compared with the untreated
group (Fig. 1A and B).
Liver functions, measured by Alt and Ast levels in
the serum, were also investigated. The results demonstrated that
Alt and Ast expression levels were significantly increased in the
CCl4-treated group compared with the untreated group
(Fig. 1C). Alterations in the
serum levels of Tnf-α and Tgf-β1 are presented in Fig. 1D. The expression levels of Tnf-α
and Tgf-β1 were significantly increased in the
CCl4-treated rats, indicating an increased inflammatory
response in the CCl4-treated rats following
treatment.
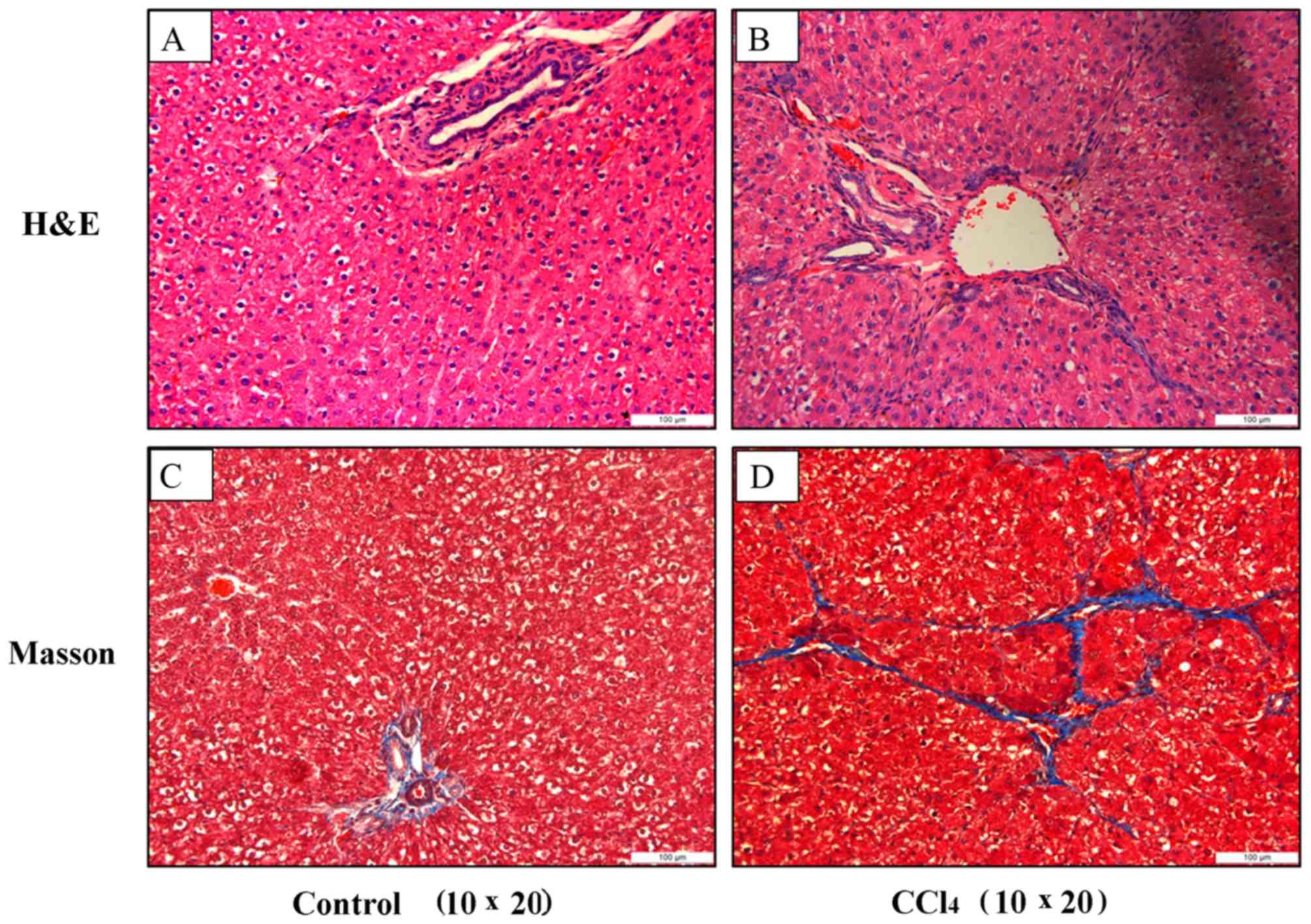
As demonstrated in Fig.
2A and B, H&E staining indicated that treatment with
CCl4 markedly increased levels of inflammation and cell
infiltration surrounding the liver cells, and also resulted in an
unclear hepatic lobular structure compared with that of the
untreated group. The results of the Masson staining indicated that
paraplastic connective tissues destroyed the structure of the liver
lobule and that marked levels of fibrosis developed in the liver
tissue of the CCl4-treated group, compared with the
untreated group where no marked liver fibrosis development was
observed (Fig. 2C and D).
Differentially expressed lncRNAs and
mRNAs
A total of 16,113 lncRNAs and 35,362 mRNAs were
identified by RNA sequencing. Compared with the untreated group,
the CCl4-treated group had 231 differentially expressed
lncRNAs (fold-change >2), which included 102 upregulated and 129
downregulated lncRNAs. The top 10 upregulated and top 10
downregulated lncRNAs with the largest fold-change are presented in
Table II. Expression profiling
data demonstrated that 1,306 mRNAs, consisting of 1,028 upregulated
and 278 downregulated mRNAs, were differentially expressed between
the two groups.
| Table II.Top 10 upregulated and downregulated
lncRNAs between untreated and carbon tetrachloride-treated liver
samples. |
Table II.
Top 10 upregulated and downregulated
lncRNAs between untreated and carbon tetrachloride-treated liver
samples.
| A, Upregulated
lncRNAs |
|---|
|
|---|
| Sequence name | Fold-change | P-value |
|---|
| LOC100364116 | 83.55 |
1.19×10−24 |
| NR_002155.1 | 36.61 |
1.29×10−10 |
| XR_595047.1 | 23.47 |
9.10×10−7 |
| LOC103692060 | 23.47 |
9.10×10−7 |
| XR_595045.1 | 23.47 |
9.10×10−7 |
| XR_595048.1 | 22.53 |
1.70×10−6 |
| XR_595044.1 | 22.53 |
1.70×10−6 |
| XR_595046.1 | 21.59 |
3.18×10−6 |
| XR_595680.1 | 19.71 |
2.11×10−10 |
| XR_355421.2 | 18.77 |
2.05×10−5 |
|
| B, Downregulated
lncRNAs |
|
| Sequence
name |
Fold-change | P-value |
|
| XR_594950.1 | 36.57 |
1.31×10−271 |
| XR_592258.1 | 32.31 |
7.51×10−163 |
| XR_591976.1 | 21.31 |
6.12×10−11 |
| LOC102554180 | 17.04 |
8.63×10−5 |
| XR_358112.2 | 12.25 |
5.14×10−6 |
| XR_356224.1 | 11.72 |
9.83×10−6 |
| XR_596548.1 | 10.65 |
3.56×10−5 |
| XR_350587.2 |
9.94 |
1.11×10−6 |
| NR_027324.1 |
9.87 |
1.63×10−120 |
| XR_352538.2 |
8.43 |
6.66×10−16 |
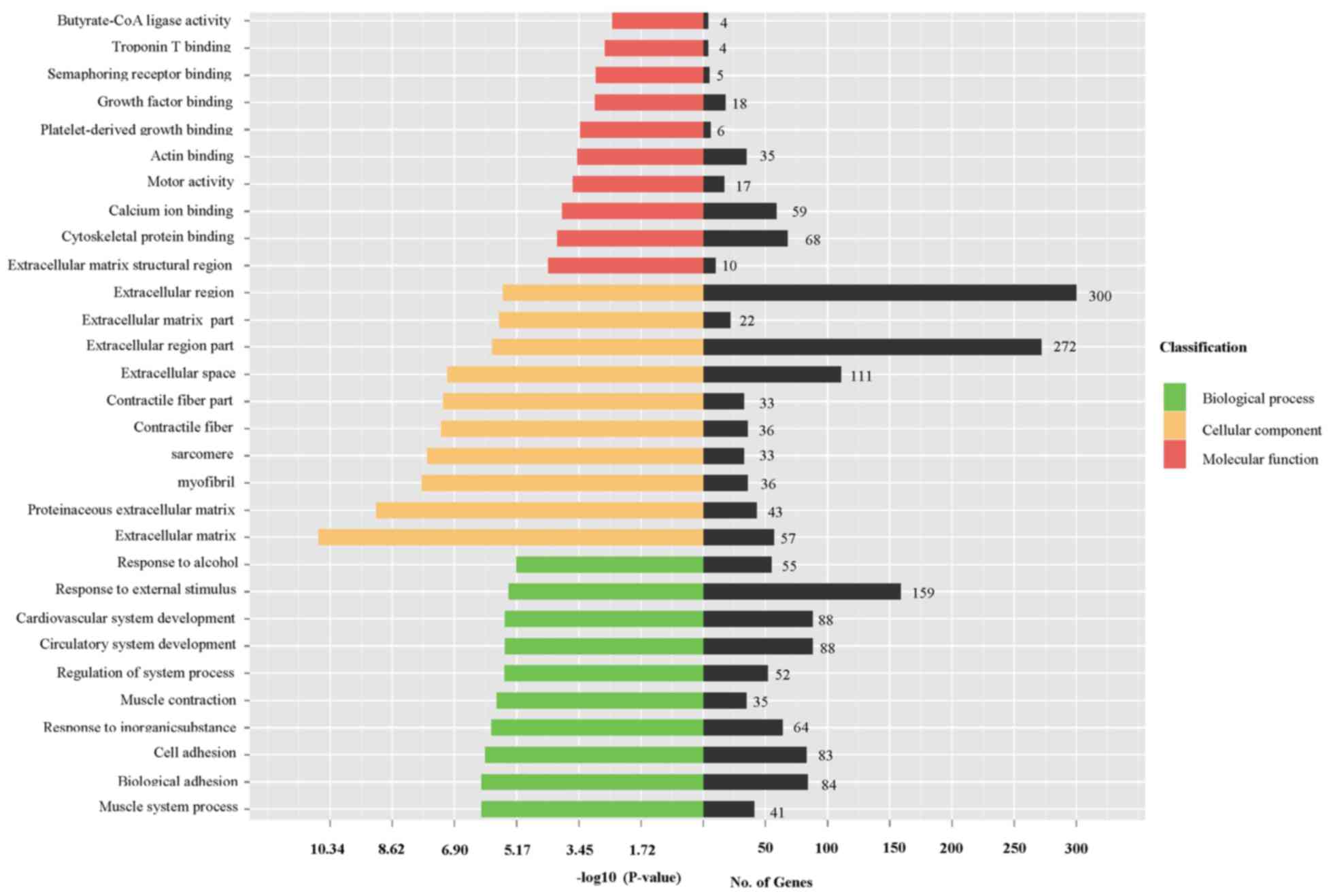
Bioinformatics analysis of
differentially expressed mRNAs
In order to identify the potential function of
dysregulated mRNAs in fibrotic rat liver tissues compared with
normal liver tissues, GO analysis was performed, during which three
items were described (biological process, cellular component and
molecular function). Differentially expressed mRNAs were
predominantly enriched in the following GO terms: ‘Muscle system
process’, ‘biological adhesion’ and ‘cell adhesion’ for biological
process terms; ‘proteinaceous extracellular matrix’, ‘extracellular
matrix’ and ‘myofibril’ for the cellular component terms; and
‘extracellular matrix structural region’, ‘cytoskeletal protein
binding’ and ‘calcium ion binding’ for molecular function terms.
The top 10 enriched GO terms for biological process, cellular
component and molecular function are presented in Fig. 3.
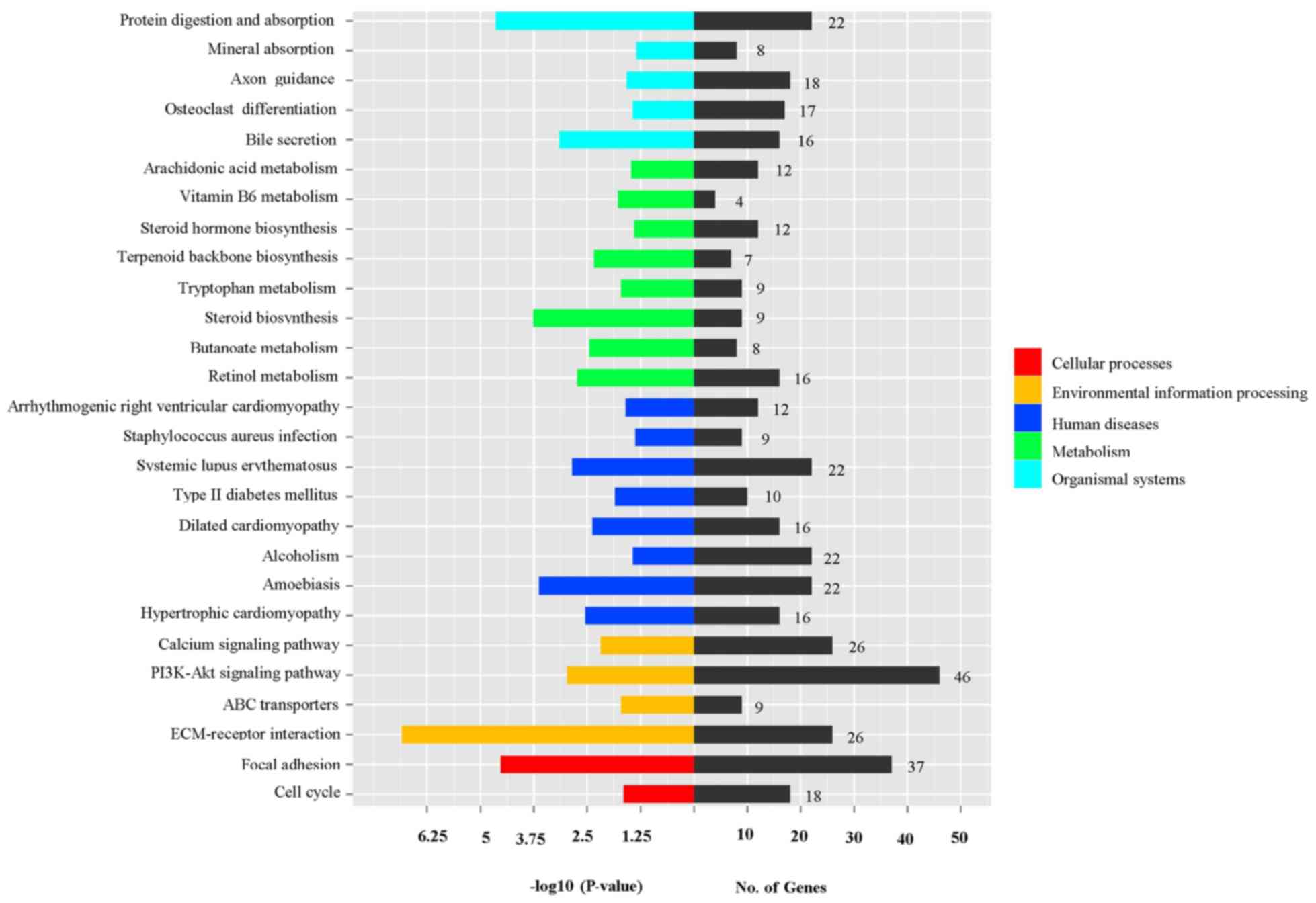
In addition, pathway analysis was performed with
reference to the KEGG database and 27 pathways presenting
significant differences (P<0.05) in gene expression were
identified between the CCl4-treated and untreated rat
liver tissues (data not shown). As revealed in Fig. 4, the differentially expressed genes
were predominantly enriched in three pathways, including
‘ECM-receptor interaction’, ‘PI3K-Akt signaling pathway’ and ‘focal
adhesion’.
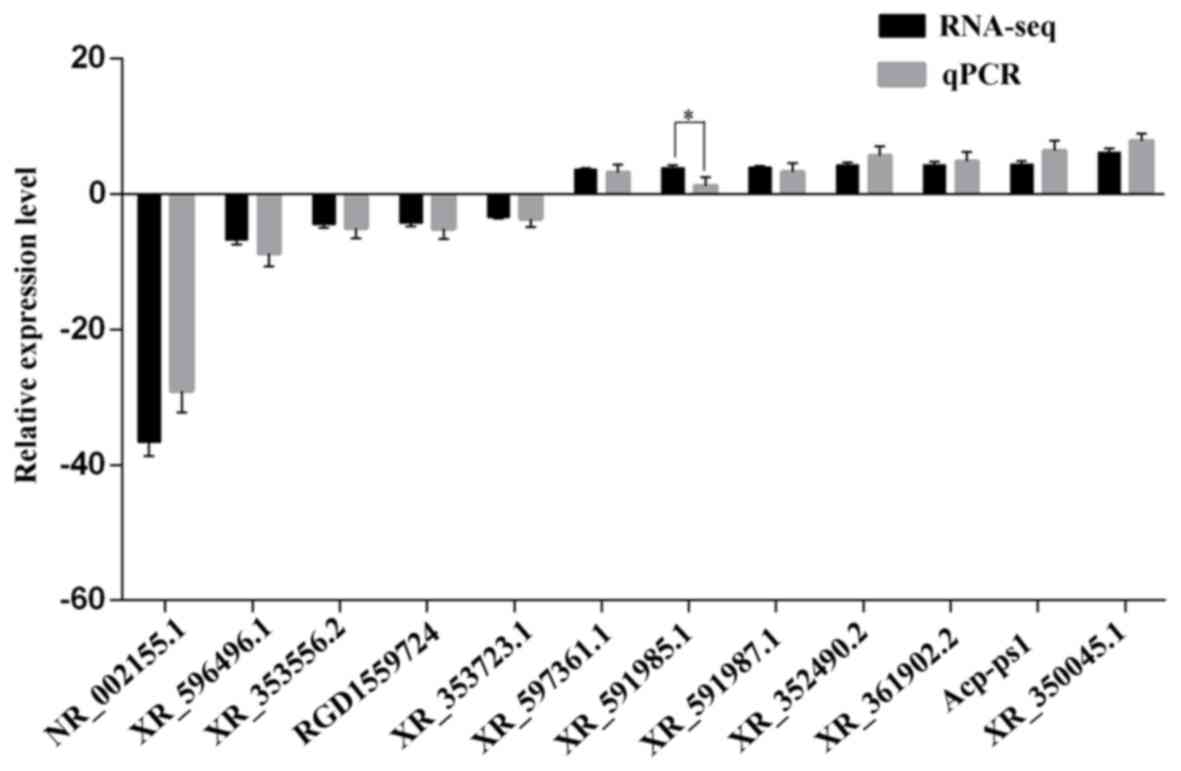
Validation by RT-qPCR
In order to verify the RNA sequencing data, 12
differentially expressed lncRNAs (nucleotides <5,000;
fold-change >3) were chosen for further analysis, 7 of which
were upregulated lncRNAs and 5 of which were downregulated lncRNAs.
RNA was extracted from all preserved liver tissues and RT-qPCR was
performed to validate the expression of selected lncRNAs. The
results demonstrated that the expression levels of 11 lncRNAs were
in accordance with the RNA sequencing data (Fig. 5); however, the expression level of
XR_591985.1 significantly differed from the results of the RNA
sequencing data. These results were considered to successfully
verify the reliability of the RNA sequencing results.
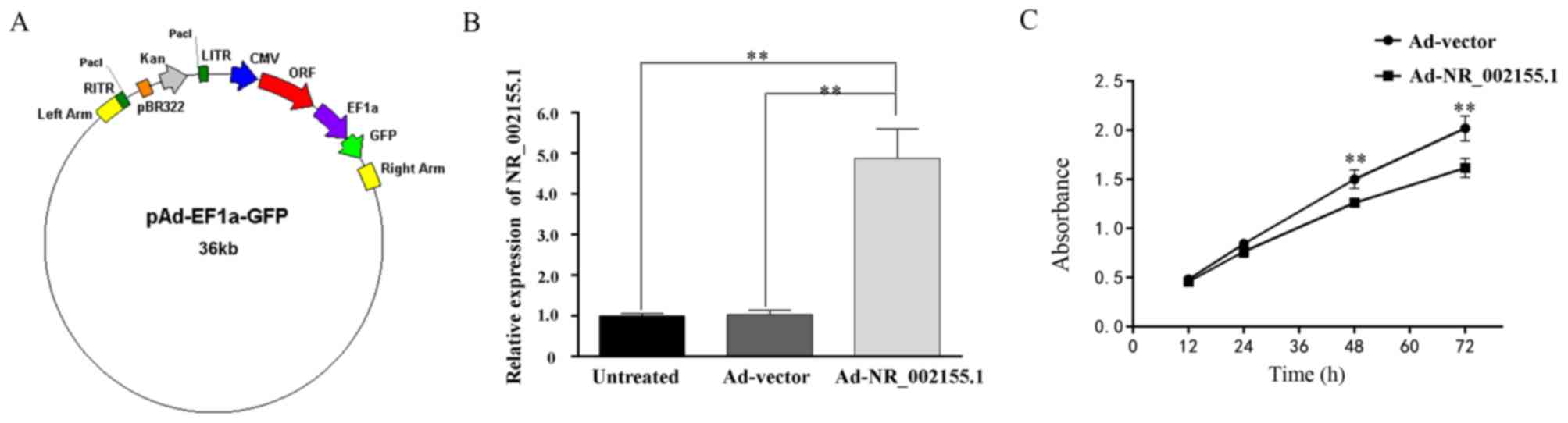
Overexpression of NR_002155.1 inhibits
HSC-T6 cell proliferation
In order to overexpress the novel lncRNA
NR_002155.1, its sequence was subcloned into the adenovirus shuttle
plasmid vector pAd-EF1a-GFP (Fig.
6A) and its successful overexpression in HSC-T6 cells was
confirmed using RT-qPCR (Fig. 6B).
Furthermore, the results of the CCK-8 assay demonstrated that
increasing the expression of NR_002155.1 significantly decreased
the proliferation of HSC-T6 cells at 48 and 72 h time intervals
(Fig. 6C), thus confirming that
NR_002155.1 may suppress the proliferation of HSCs and may
therefore inhibit liver fibrosis induced by CCl4.
Discussion
CCl4 administration leads to the
infiltration of inflammatory cells and subsequent liver damage
leads to the development of fibrosis. In order to investigate the
lncRNAs and mRNAs involved in the progression of liver fibrosis, an
animal model of liver injury and fibrosis was constructed using
male Sprague-Dawley rats treated with CCl4. The
predominant biological processes and pathways enriched in the
overlapping differentially expressed mRNAs were revealed. Following
this, the differentially expressed lncRNAs were analyzed and their
expression levels were verified using RT-qPCR. Finally, one of the
most significant differentially expressed lncRNAs, NR_002155.1, was
investigated further in order to determine its role in liver
fibrosis.
The results of the present study revealed that liver
tissues from rats in the CCl4-treated group exhibited
signs of inflammation and the occurrence of fibrosis. The body
weight and liver/body weight ratio of the rats were decreased
following CCl4 treatment for 8 weeks. Furthermore,
circulating levels of liver function indicators (Alt, Ast, Tgf-β1
and Tnf-α) were significantly enhanced in the CCl4
treated group. In addition, histological staining indicated that
there was marked inflammatory infiltration, collagen deposition and
liver fibrosis in the CCl4-treated liver tissues.
Therefore, it may be hypothesized that the aforementioned
consequences resulted from the basic toxicity of CCl4,
and this conclusion would be in accordance with the findings of
previous studies (23,24).
When liver tissues and cells are exposed to
CCl4, a complex process inducing resistance to toxicity
occurs. CCl4 metabolism in liver tissues or cells
induces the generation of free radicals (25) and thus triggers oxidative stress,
which results in the initiation and progression of liver damage
(26). Furthermore, oxidative
stress enhances the secretion of inflammatory cytokines, induces
inflammation and necrosis of hepatocytes, and promotes the
progression of liver fibrosis (27). These pathological consequences of
oxidative stress were demonstrated by the present study, in which
levels of Alt, Ast, Tgf-β1 and Tnf-α were increased in the serum of
CCl4-treated rats compared with untreated rats.
GO analysis in the present study revealed that 1,306
mRNAs were differentially expressed in liver tissues from
CCl4-treated rats compared with the untreated group.
These differentially expressed mRNAs were predominantly enriched in
‘extracellular matrix’, ‘response to external stimulus’, ‘cell
adhesion’ and ‘biological adhesion’. With regards to KEGG pathway
analysis, the differentially expressed mRNAs in the
CCl4-treated liver tissues were demonstrated to be
predominantly enriched in three pathways: ‘ECM-receptor
interaction’, ‘PI3K-Akt’ and ‘focal adhesion’.
HSCs are a type of hepatic nonparenchymal cell, and
their activation and proliferation are involved in liver fibrosis.
HSCs exposed to chronic injury are activated and undergo
proliferation. Excessive proliferation of HSCs leads to the
secretion of excessive levels of ECM, thus causing abnormal ECM
deposition. Abundant ECM has the potential to replace normal
hepatocytes (28), which
contributes to scar formation and fibrosis (29). These previous findings were
verified in the present study via Masson staining. ECM-receptor
interaction, associated with signaling pathways involved in
paracrine/endocrine functioning, may affect cell phenotype,
adhesion, migration, proliferation and differentiation (30). The focal adhesion signaling pathway
primarily involves integrin and focal adhesion kinase (FAK). As a
tyrosine kinase receptor, FAK is reported to be activated by
platelet-derived growth factor (PDGF) and, following interaction
with ECM proteins via association with integrin, FAK directly
senses the integrity of the extracellular environment (31–33).
The activation of FAK has been reported to be essential for
PDGF-induced HSC proliferation (34). As a key downstream kinase of
integrin/FAK, phosphoinositide 3-kinase (PI3K) is also activated by
PDGF stimulation and response to the proliferation of PDGF-induced
HSCs (34,35). Stimulation of the PI3K pathway has
been demonstrated to be essential for HSC proliferation in
vitro and in CCl4-induced rat liver fibrosis in
vivo (36). Akt, which is
positioned downstream of PI3K, is also reported to be involved in
the stimulation of cell proliferation and inhibition of cell
apoptosis in HSCs (37,38). Type I collagen gene expression was
demonstrated to be primarily regulated by the PI3K-Akt pathway
(34). The expression levels of
type I collagen mRNA and protein were significantly reduced by the
PI3K inhibitor LY294002 or by inhibition of Akt expression via
adenoviral-mediated transduction with a dominant negative form of
Akt (39). Therefore, the
ECM-receptor interaction, focal adhesion and the PI3K-Akt pathways
all interact with each other in order to regulate the process of
fibrogenesis.
In the present study, RNA sequencing demonstrated
that 231 lncRNAs were significantly differentially expressed
(fold-change >2) between CCl4-treated and untreated
rats' liver tissue. The majority of identified lncRNAs were not
previously reported and the functions of these lncRNAs remain
unknown. RT-qPCR analyses were performed to verify the reliability
of the RNA sequencing data. The expression levels of the selected
lncRNAs determined by RT-qPCR were consistent with RNA sequencing
data. In order to verify whether the differentially expressed
lncRNAs identified by RNA sequencing contribute to the progression
of liver fibrosis, the biological function of NR_002155.1 was
investigated (fold-change=36.6). NR_002155.1, a newly identified
long noncoding RNA with a length of 951 base pairs, to the best of
our knowledge, has not been previously investigated. In the present
study, it was demonstrated that NR_002155.1 may inhibit HSC-T6 cell
proliferation in vitro, which is consistent with the
findings of the RNA sequencing analysis.
In conclusion, the expression profile of lncRNAs in
CCl4-induced liver fibrosis was determined via RNA
sequencing and differentially expressed lncRNAs and mRNAs were
identified. Differentially expressed mRNAs between liver tissues
from CCl4-treated and untreated rats were predominantly
enriched in the ‘ECM-receptor interaction’, ‘focal adhesion’ and
‘PI3K-Akt signaling pathway’ path ways. Following validation of
these results by RT-qPCR analysis, the biological function of the
aberrantly expressed lncRNA NR_002155.1 was investigated. The
results of the in vitro function experiments indicated that
lncRNA NR_002155.1 may inhibit the proliferation of HSCs in
vitro and in vivo. However, the mechanisms of the
identified lncRNAs in liver fibrosis are yet to be determined.
Future studies should further investigate the association and
network construction between the lncRNAs and mRNAs identified in
the present study. In addition, further studies should explore the
biological functions and regulatory mechanisms of lncRNAs, which
may uncover novel diagnostic biomarkers and effective therapeutic
strategies for liver fibrosis.
Acknowledgements
Not applicable.
Funding
The present study was supported by the National
Natural Science Foundation of China (grant no. 81473473), the
Education Department of Jiangxi Province (grant no. GJJ160039), the
Science Technology and Innovation Commission of Shenzhen
Municipality in China (grant no. JCYJ20160428173252471) and the
Health and Family Planning Commission of Shenzhen Municipality in
China (grant no. SZSM201512026).
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors' contributions
ZG, JT, TX, JL and CD performed the experiments. ZG,
YP, JZ and GH analyzed and interpreted the RNA sequencing data. ZG
and GH were major contributors in writing the manuscript. All
authors read and approved the final manuscript.
Ethics approval and consent to
participate
Animal experiments were approved by the Animal
Ethics Committee of Peking University Shenzhen Hospital (Shenzhen,
China).
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
lncRNAs
|
long noncoding RNAs
|
|
CCl4
|
carbon tetrachloride
|
|
Alt
|
alanine aminotransferase
|
|
Ast
|
aspartate aminotransferase
|
|
ECM
|
extracellular matrix
|
|
GO
|
Gene Ontology
|
|
KEGG
|
Kyoto Encyclopedia of Genes and
Genomes
|
|
RT-qPCR
|
reverse transcription-quantitative
polymerase chain reaction
|
|
HSCs
|
hepatic stellate cells
|
|
H&E
|
hematoxylin and eosin
|
|
CCK-8
|
Cell Counting Kit-8
|
References
|
1
|
Duval F, Moreno-Cuevas JE, Gonzalez-Garza
MT, Maldonado-Bernal C and Cruz-Vega DE: Liver fibrosis and
mechanisms of the protective action of medicinal plants targeting
inflammation and the immune response. Int J Inflam.
2015:9434972015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Enomoto M, Morikawa H, Tamori A and Kawada
N: Noninvasive assessment of liver fibrosis in patients with
chronic hepatitis B. World J Gastroenterol. 20:12031–12038. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Xuan J, Chen S, Ning B, Tolleson WH and
Guo L: Development of HepG2-derived cells expressing cytochrome
P450s for assessing metabolism-associated drug-induced liver
toxicity. Chem Biol Interact. 255:63–73. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Huang ZG, Zhai WR, Zhang YE and Zhang XR:
Study of heteroserum-induced rat liver fibrosis model and its
mechanism. World J Gastroenterol. 4:206–209. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Novikova IV, Hennelly SP and Sanbonmatsu
KY: Sizing up long non-coding RNAs: Do lncRNAs have secondary and
tertiary structure? Bioarchitecture. 2:189–199. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Geisler S and Coller J: RNA in unexpected
places: Long non-coding RNA functions in diverse cellular contexts.
Nat Rev Mol Cell Biol. 14:699–712. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Gibb EA, Brown CJ and Lam WL: The
functional role of long non-coding RNA in human carcinomas. Mol
Cancer. 10:382011. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Li J, Tian H, Yang J and Gong Z: Long
noncoding RNAs regulate cell growth, proliferation, and apoptosis.
DNA Cell Biol. 35:459–470. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Briggs JA, Wolvetang EJ, Mattick JS, Rinn
JL and Barry G: Mechanisms of long Non-coding RNAs in mammalian
nervous system development, plasticity, disease, and evolution.
Neuron. 88:861–877. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Schmitt AM and Chang HY: Long noncoding
RNAs in cancer pathways. Cancer Cell. 29:452–463. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Roberts TC, Morris KV and Weinberg MS:
Perspectives on the mechanism of transcriptional regulation by long
non-coding RNAs. Epigenetics. 9:13–20. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Wang KC and Chang HY: Molecular mechanisms
of long noncoding RNAs. Mol Cell. 43:904–914. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Yu F, Zheng J, Mao Y, Dong P, Li G, Lu Z,
Guo C, Liu Z and Fan X: Long non-coding RNA APTR promotes the
activation of hepatic stellate cells and the progression of liver
fibrosis. Biochem Biophys Res Commun. 463:679–685. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Wu Y, Liu X, Zhou Q, Huang C, Meng X, Xu F
and Li J: Silent information regulator 1 (SIRT1) ameliorates liver
fibrosis via promoting activated stellate cell apoptosis and
reversion. Toxicol Appl Pharmacol. 289:163–176. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
He Y, Wu YT, Huang C, Meng XM, Ma TT, Wu
BM, Xu FY, Zhang L, Lv XW and Li J: Inhibitory effects of long
noncoding RNA MEG3 on hepatic stellate cells activation and liver
fibrogenesis. Biochim Biophys Acta. 1842:2204–2215. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Yu F, Zheng J, Mao Y, Dong P, Lu Z, Li G,
Guo C, Liu Z and Fan X: Long Non-coding RNA growth Arrest-specific
transcript 5 (GAS5) inhibits liver fibrogenesis through a mechanism
of competing endogenous RNA. J Biol Chem. 290:28286–28298. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Kong X, Zhou W, Wan JB, Zhang Q, Ni J and
Hu Y: An integrative thrombosis network: Visualization and
topological analysis. Evid Based Complement Alternat Med.
2015:2653032015. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
National Research Council, . Guide for the
Care and Use of Laboratory Animals. The National Academies Press;
Washington, DC: 1996, https://www.nap.edu/catalog/5140/guide-for-the-care-and-use-of-laboratory-animals
|
|
19
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Trapnell C, Pachter L and Salzberg SL:
TopHat: Discovering splice junctions with RNA-Seq. Bioinformatics.
25:1105–1111. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Feng J, Meyer CA, Wang Q, Liu JS, Liu
Shirley X and Zhang Y: GFOLD: A generalized fold-change for ranking
differentially expressed genes from RNA-seq data. Bioinformatics.
28:2782–2788. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Trapnell C, Roberts A, Goff L, Pertea G,
Kim D, Kelley DR, Pimentel H, Salzberg SL, Rinn JL and Pachter L:
Differential gene and transcript expression analysis of RNA-seq
experiments with TopHat and Cufflinks. Nat Protoc. 7:562–578. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Dong S, Chen QL, Song YN, Sun Y, Wei B, Li
XY, Hu YY, Liu P and Su SB: Mechanisms of CCl4-induced liver
fibrosis with combined transcriptomic and proteomic analysis. J
Toxicol Sci. 41:561–572. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Marques TG, Chaib E, da Fonseca JH,
Lourenço AC, Silva FD, Ribeiro MA Jr, Galvão FH and D'Albuquerque
LA: Review of experimental models for inducing hepatic cirrhosis by
bile duct ligation and carbon tetrachloride injection. Acta Cir
Bras. 27:589–594. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Sancheti S, Sancheti S and Seo SY:
Ameliorative effects of 7-methylcoumarin and 7-methoxycoumarin
against CCl4-induced hepatotoxicity in rats. Drug Chem Toxicol.
36:42–47. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Li S, Tan HY, Wang N, Zhang ZJ, Lao L,
Wong CW and Feng Y: The role of oxidative stress and antioxidants
in liver diseases. Int J Mol Sci. 16:26087–26124. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Heeba GH and Mahmoud ME: Therapeutic
potential of morin against liver fibrosis in rats: Modulation of
oxidative stress, cytokine production and nuclear factor kappa B.
Environ Toxicol Pharmacol. 37:662–671. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Duval F, Moreno-Cuevas JE, Gonzalez-Garza
MT, Rodriguez-Montalvo C and Cruz-Vega DE: Protective mechanisms of
medicinal plants targeting hepatic stellate cell activation and
extracellular matrix deposition in liver fibrosis. Chin Med.
9:272014. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Roy S, Benz F, Cardenas Vargas D, Vucur M,
Gautheron J, Schneider A, Hellerbrand C, Pottier N, Alder J, Tacke
F, et al: miR-30c and miR-193 are a part of the TGF-β-dependent
regulatory network controlling extracellular matrix genes in liver
fibrosis. J Dig Dis. 16:513–524. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Karsdal MA, Manon-Jensen T, Genovese F,
Kristensen JH, Nielsen MJ, Sand JM, Hansen NU, Bay-Jensen AC, Bager
CL, Krag A, et al: Novel insights into the function and dynamics of
extracellular matrix in liver fibrosis. Am J Physiol Gastrointest
Liver Physiol. 308:G807–G830. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Carloni V, Romanelli RG, Pinzani M, Laffi
G and Gentilini P: Focal adhesion kinase and phospholipase C gamma
involvement in adhesion and migration of human hepatic stellate
cells. Gastroenterology. 112:522–531. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Renshaw MW, Price LS and Schwartz MA:
Focal adhesion kinase mediates the integrin signaling requirement
for growth factor activation of MAP kinase. J Cell Biol.
147:611–618. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Wang Y, Ma J, Chen L, Xie XL and Jiang H:
Inhibition of focal adhesion kinase on hepatic Stellate-cell
adhesion and migration. Am J Med Sci. 353:41–48. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Reif S, Lang A, Lindquist JN, Yata Y,
Gabele E, Scanga A, Brenner DA and Rippe RA: The role of focal
adhesion kinase-phosphatidylinositol 3-kinase-akt signaling in
hepatic stellate cell proliferation and type I collagen expression.
J Biol Chem. 278:8083–8090. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Novo E, Cannito S, Paternostro C, Bocca C,
Miglietta A and Parola M: Cellular and molecular mechanisms in
liver fibrogenesis. Arch Biochem Biophys. 548:20–37. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Lin X, Bai F, Nie J, Lu S, Lu C, Zhu X,
Wei J, Lu Z and Huang Q: Didymin alleviates hepatic fibrosis
through inhibiting ERK and PI3K/Akt pathways via regulation of raf
kinase inhibitor protein. Cell Physiol Biochem. 40:1422–1432. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Kim AH, Khursigara G, Sun X, Franke TF and
Chao MV: Akt phosphorylates and negatively regulates apoptosis
signal-regulating kinase 1. Mol Cell Biol. 21:893–901. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Coffer PJ, Jin J and Woodgett JR: Protein
kinase B (c-Akt): A multifunctional mediator of
phosphatidylinositol 3-kinase activation. Biochem J. 335:1–13.
1998. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Ricupero DA, Poliks CF, Rishikof DC,
Cuttle KA, Kuang PP and Goldstein RH: Phosphatidylinositol
3-kinase-dependent stabilization of alpha1(I) collagen mRNA in
human lung fibroblasts. Am J Physiol Cell Physiol. 281:C99–C105.
2001. View Article : Google Scholar : PubMed/NCBI
|